
Environmental Enrichment Induces Meningeal Niche Remodeling through TrkB-Mediated Signaling

 ,
,  ,
, 
 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
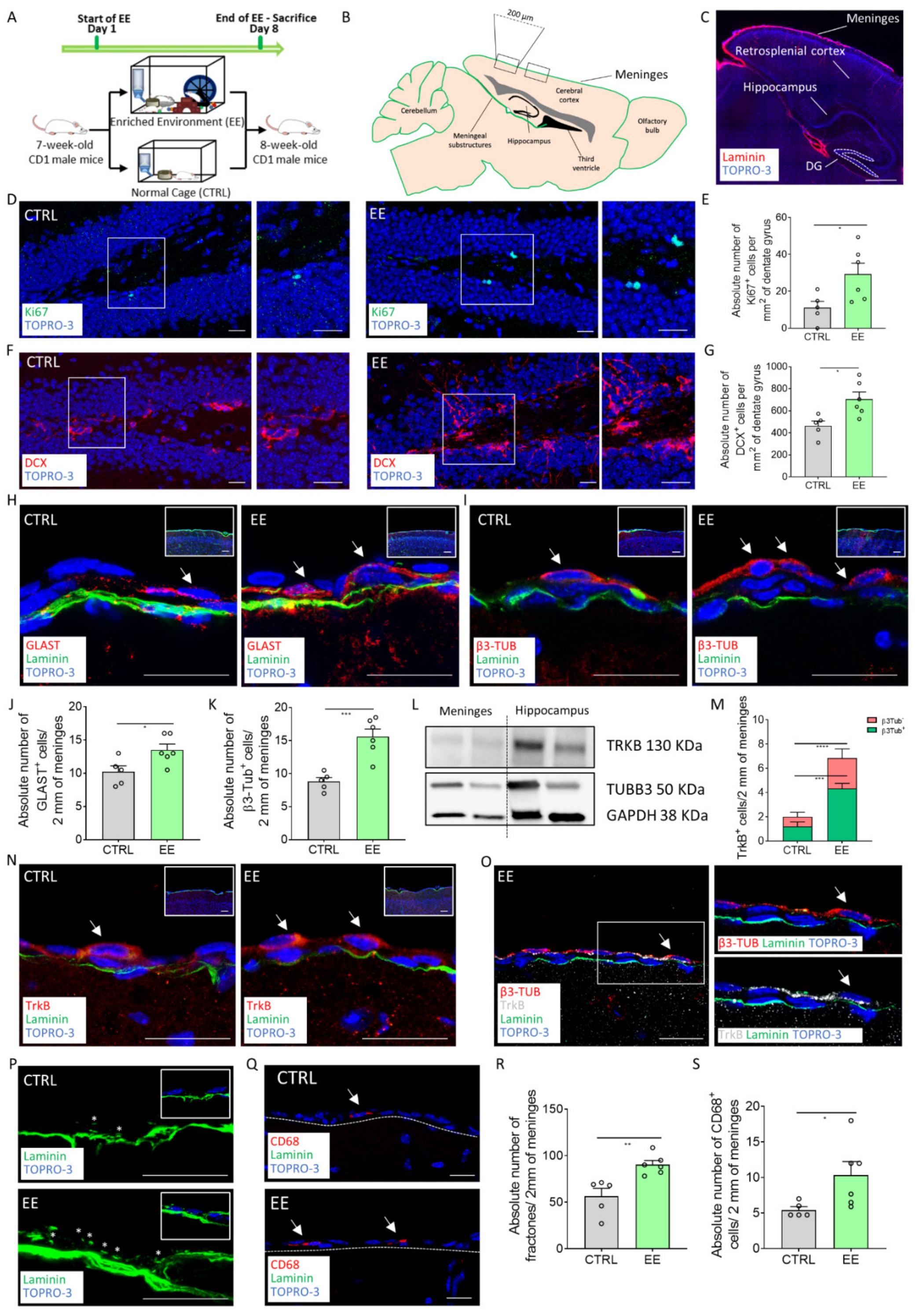
2.1. EE Exposure Induces Meningeal Niche Remodeling
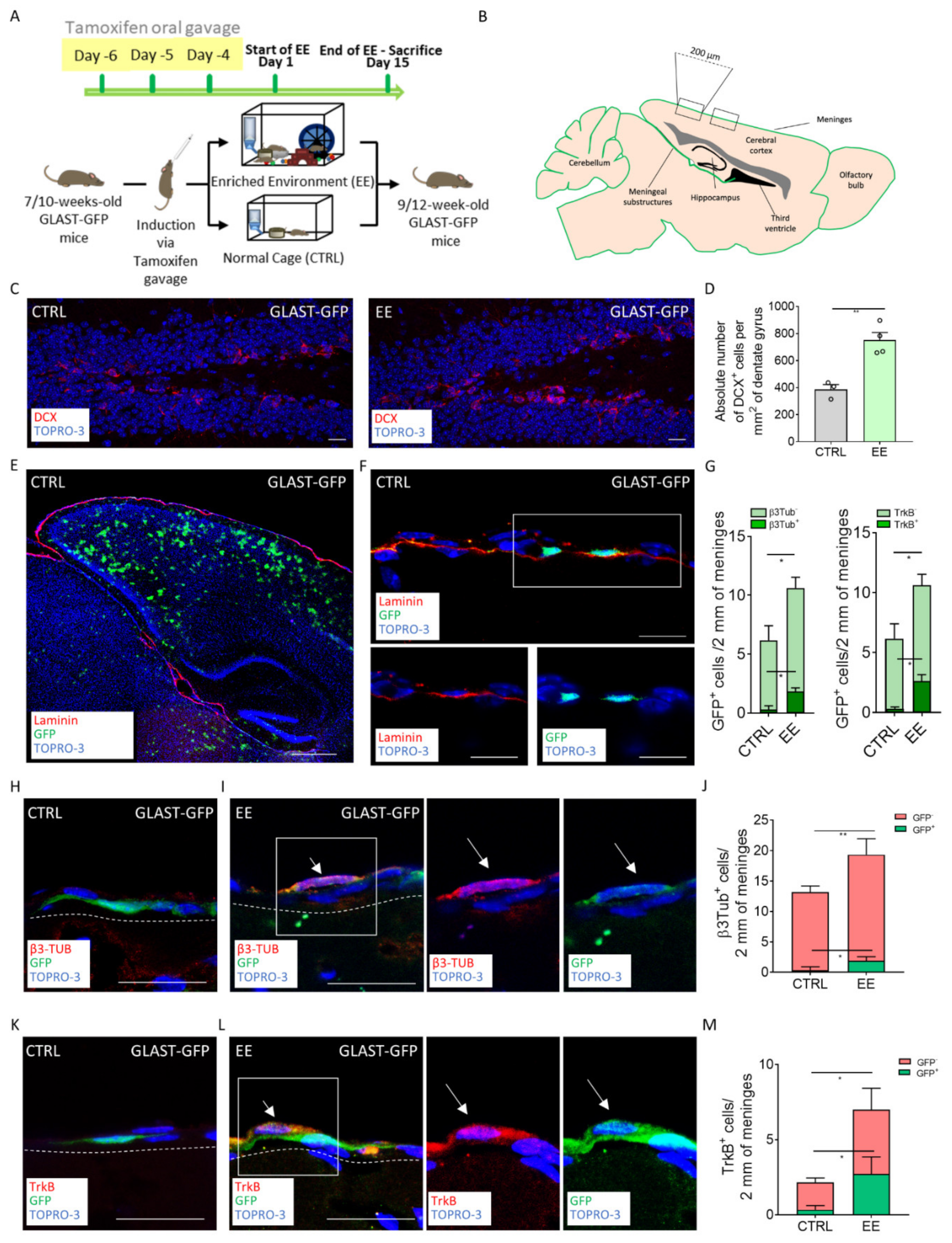
2.2. EE-Induced NPs in Meninges Derived from GLAST+ Radial Glia Progenitors
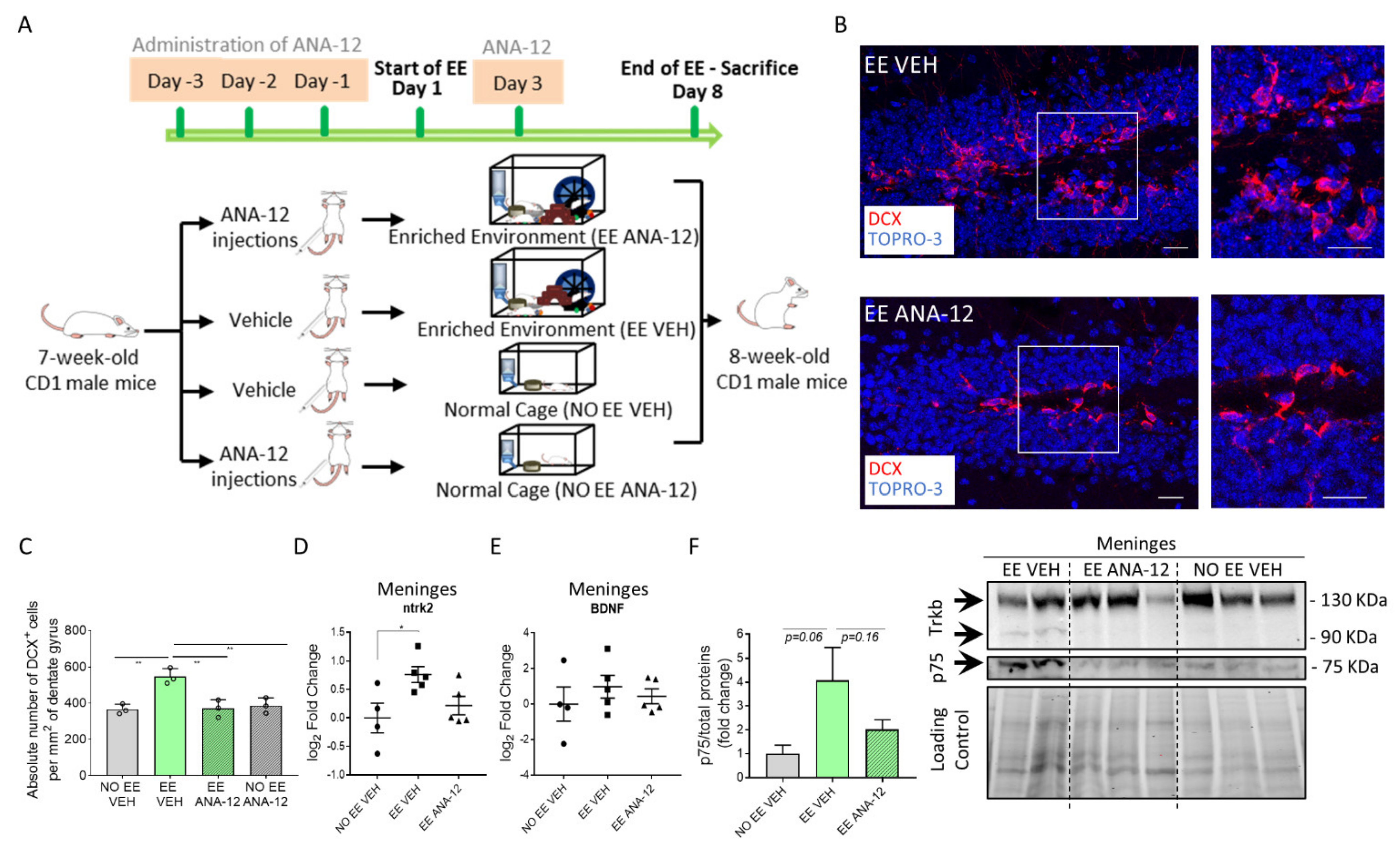
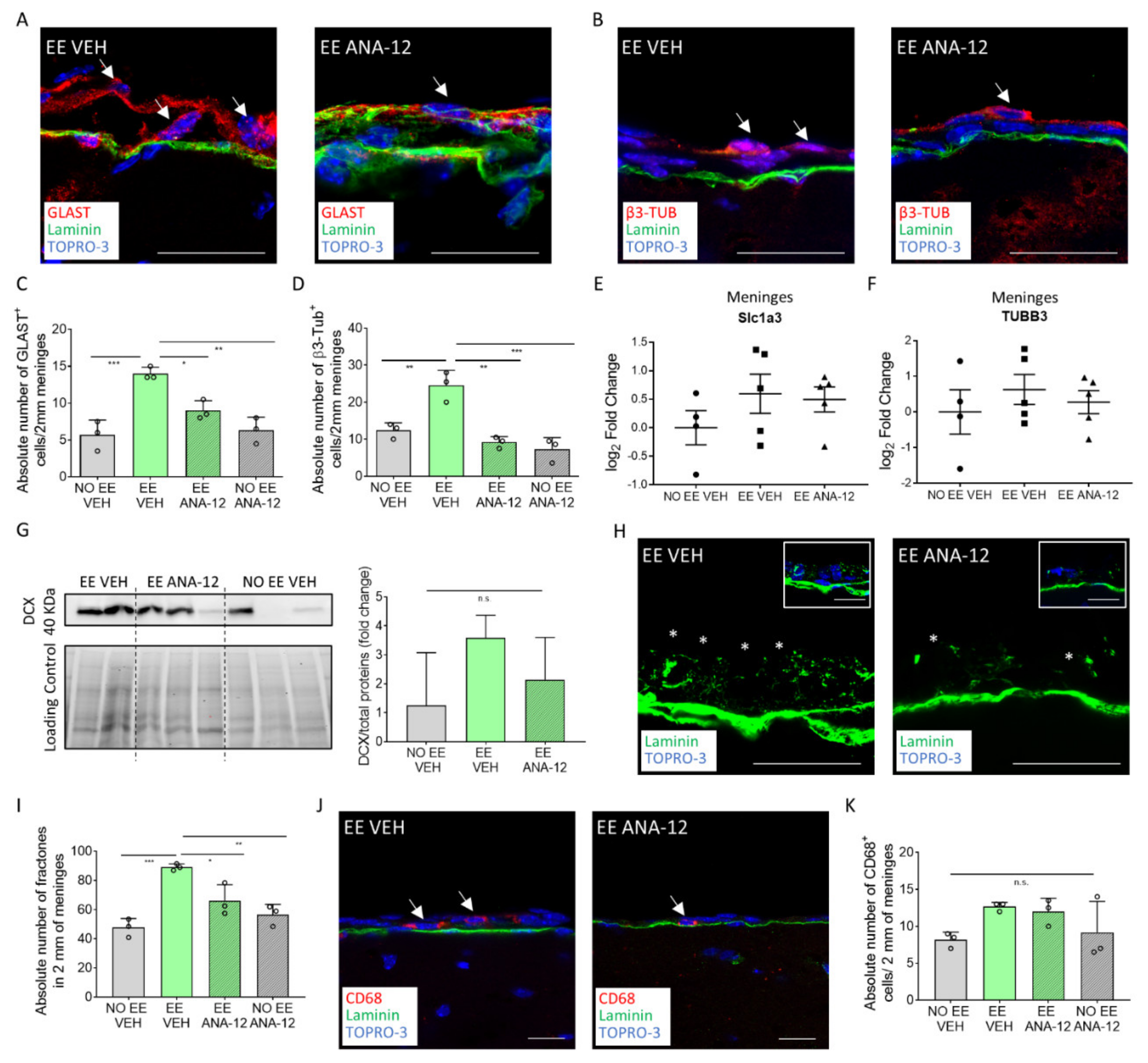
2.3. TrkB/BDNF Signaling Modulates the Meningeal Niche Response to Enriched Environment Exposure
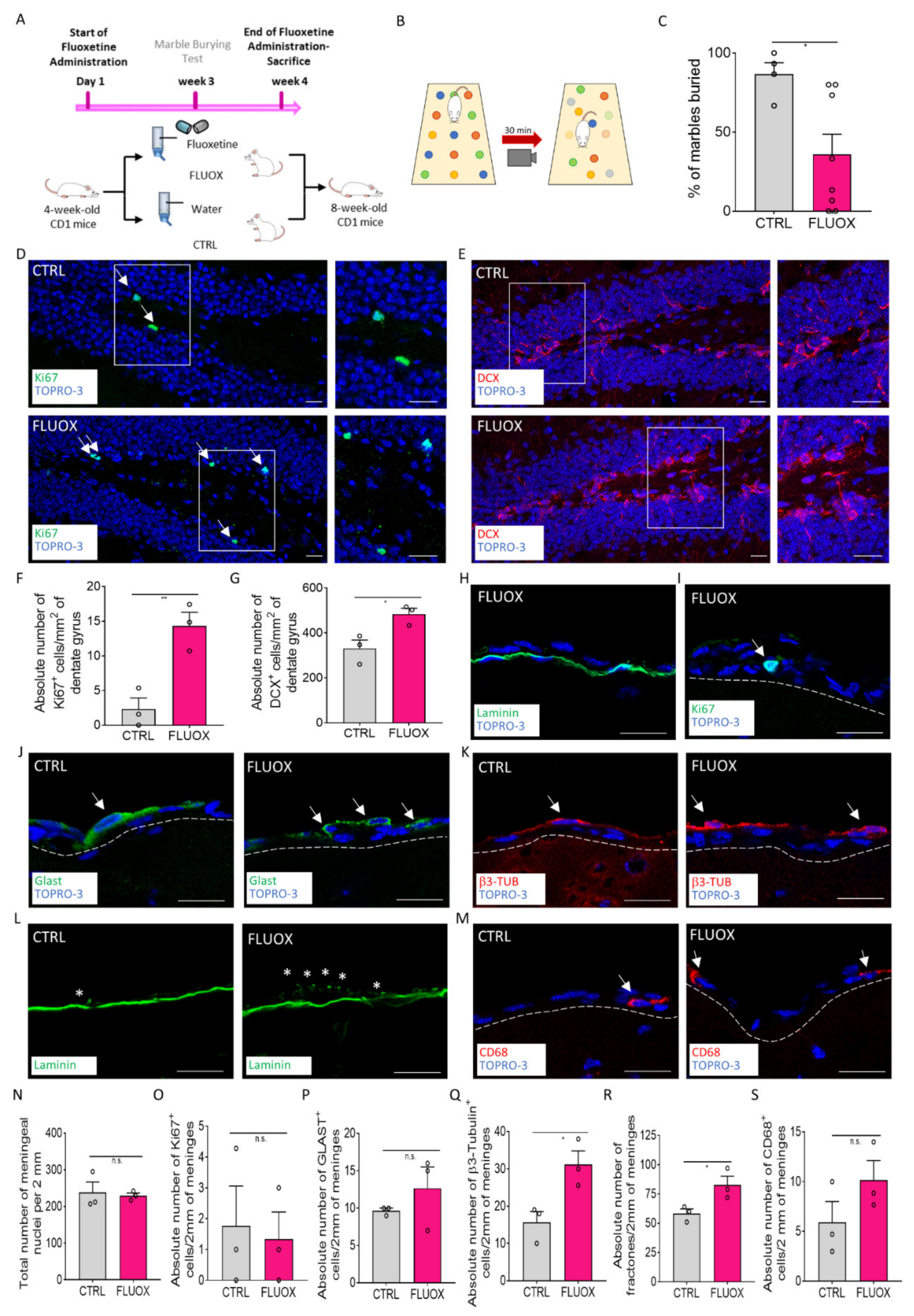
2.4. Fluoxetine Administration Induces the Meningeal Niche Response
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Exposure to an Enriched Environment
4.3. GLAST-GFP Exposure to EE
4.4. Exposure to the EE and TrkB Inhibitor ANA-12
4.5. Fluoxetine Administration
4.6. Marble Test Administration
4.7. Tissue Preparation and Immunofluorescence
4.8. Antibodies
4.9. Immunofluorescence Image Acquisition, Analysis and Quantification
4.10. Protein Extraction from the Mouse Meninges and Immunoblot Analysis
4.11. RNA Extraction from the Mouse Brain Meninges and Real-Time (rt) PCR Analysis
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BDNF | Brain-derived neurotrophic factors |
| BrdU | Bromodeoxyuridine |
| CD68 | Cluster of differentiation 68 |
| CNS | Central nervous system |
| CTRL | Control animal |
| DCX | Doublecortin |
| DG | Dentate gyrus |
| ECM | Extracellular matrix |
| EE | Enriched environment |
| FGF | Fibroblast growth factor |
| FGF2 | Fibroblast growth factor 2 |
| FLUOX | Animal treated with fluoxetine |
| GFP | Green fluorescent protein |
| GLAST | Glutamate and aspartate transporter |
| MBT | Marble burying test |
| NGF | Nerve growth factor |
| NP | Neural precursors |
| NSC | Neural stem cells |
| OCD | Obsessive compulsive disorder |
| RG | Radial glia |
| SCI | Spinal cord injury |
| scRNAseq | single-cell RNA sequencing |
| SSRI | Selective serotonin reuptake inhibitor |
| TrkB | Tropomyosin receptor kinase B |
| WT | Wild-type |
| β3-Tub | Tubulin β3 |
References
- Venna, V.R.; Xu, Y.; Doran, S.J.; Patrizz, A.; McCullough, L.D. Social interaction plays a critical role in neurogenesis and recovery after stroke. Transl. Psychiatry 2014, 4, e351. [Google Scholar] [CrossRef]
- Benarroch, E.E. Adult neurogenesis in the dentate gyrus: General concepts and potential implications. Neurology 2013, 81, 1443–1452. [Google Scholar] [CrossRef]
- Samuels, B.A.; Hen, R. Neurogenesis and affective disorders. Eur. J. Neurosci. 2011, 33, 1152–1159. [Google Scholar] [CrossRef]
- David, D.J.; Samuels, B.A.; Rainer, Q.; Wang, J.W.; Marsteller, D.; Mendez, I.; Drew, M.; Craig, D.A.; Guiard, B.P.; Guilloux, J.P.; et al. Neurogenesis-dependent and -independent effects of fluoxetine in an animal model of anxiety/depression. Neuron 2009, 62, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Ceccarelli, M.; D’Andrea, G.; Tirone, F. Depression and adult neurogenesis: Positive effects of the antidepressant fluoxetine and of physical exercise. Brain Res. Bull 2018, 143, 181–193. [Google Scholar] [CrossRef]
- Wang, J.W.; David, D.J.; Monckton, J.E.; Battaglia, F.; Hen, R. Chronic fluoxetine stimulates maturation and synaptic plasticity of adult-born hippocampal granule cells. J. Neurosci. 2008, 28, 1374–1384. [Google Scholar] [CrossRef]
- Forbes, T.A.; Goldstein, E.Z.; Dupree, J.L.; Jablonska, B.; Scafidi, J.; Adams, K.L.; Imamura, Y.; Hashimoto-Torii, K.; Gallo, V. Environmental enrichment ameliorates perinatal brain injury and promotes functional white matter recovery. Nat. Commun. 2020, 11, 964. [Google Scholar] [CrossRef]
- Ferguson, J.M. Mechanism of Action of Antidepressant Medications. Prim. Care Companion J. Clin. Psychiatry 2001, 3, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef]
- Eisinger, B.E.; Zhao, X. Identifying molecular mediators of environmentally enhanced neurogenesis. Cell Tissue Res. 2018, 371, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Bjorkholm, C.; Monteggia, L.M. BDNF—A key transducer of antidepressant effects. Neuropharmacology 2016, 102, 72–79. [Google Scholar] [CrossRef]
- Kozisek, M.E.; Middlemas, D.; Bylund, D.B. Brain-derived neurotrophic factor and its receptor tropomyosin-related kinase B in the mechanism of action of antidepressant therapies. Pharmacol. Ther. 2008, 117, 30–51. [Google Scholar] [CrossRef]
- Casarotto, P.C.; Girych, M.; Fred, S.M.; Kovaleva, V.; Moliner, R.; Enkavi, G.; Biojone, C.; Cannarozzo, C.; Sahu, M.P.; Kaurinkoski, K.; et al. Antidepressant drugs act by directly binding to TRKB neurotrophin receptors. Cell 2021, 184, 1299–1313.e19. [Google Scholar] [CrossRef]
- Pino, A.; Fumagalli, G.; Bifari, F.; Decimo, I. New neurons in adult brain: Distribution, molecular mechanisms and therapies. Biochem. Pharmacol. 2017, 141, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Bifari, F.; Berton, V.; Pino, A.; Kusalo, M.; Malpeli, G.; Di Chio, M.; Bersan, E.; Amato, E.; Scarpa, A.; Krampera, M.; et al. Meninges harbor cells expressing neural precursor markers during development and adulthood. Front. Cell Neurosci. 2015, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Decimo, I.; Dolci, S.; Panuccio, G.; Riva, M.; Fumagalli, G.; Bifari, F. Meninges: A Widespread Niche of Neural Progenitors for the Brain. Neuroscientist 2021, 5, 506–528. [Google Scholar] [CrossRef] [PubMed]
- Dolci, S.; Pino, A.; Berton, V.; Gonzalez, P.; Braga, A.; Fumagalli, M.; Bonfanti, E.; Malpeli, G.; Pari, F.; Zorzin, S.; et al. High Yield of Adult Oligodendrocyte Lineage Cells Obtained from Meningeal Biopsy. Front. Pharmacol. 2017, 8, 703. [Google Scholar] [CrossRef]
- Bifari, F.; Decimo, I.; Chiamulera, C.; Bersan, E.; Malpeli, G.; Johansson, J.; Lisi, V.; Bonetti, B.; Fumagalli, G.; Pizzolo, G.; et al. Novel stem/progenitor cells with neuronal differentiation potential reside in the leptomeningeal niche. J. Cell Mol. Med. 2009, 13, 3195–3208. [Google Scholar] [CrossRef]
- Bifari, F.; Dolci, S.; Bottani, E.; Pino, A.; di Chio, M.; Zorzin, S.; Ragni, M.; Zamfir, R.G.; Brunetti, D.; Bardelli, D.; et al. Complete neural stem cell (NSC) neuronal differentiation requires a branched chain amino acids-induced persistent metabolic shift towards energy metabolism. Pharmacol. Res. 2020, 158, 104863. [Google Scholar] [CrossRef]
- Bifari, F.; Decimo, I.; Pino, A.; Llorens-Bobadilla, E.; Zhao, S.; Lange, C.; Panuccio, G.; Boeckx, B.; Thienpont, B.; Vinckier, S.; et al. Neurogenic Radial Glia-like Cells in Meninges Migrate and Differentiate into Functionally Integrated Neurons in the Neonatal Cortex. Cell Stem Cell 2017, 20, 360–373.e7. [Google Scholar] [CrossRef]
- Dang, T.C.; Ishii, Y.; Nguyen, V.; Yamamoto, S.; Hamashima, T.; Okuno, N.; Nguyen, Q.L.; Sang, Y.; Ohkawa, N.; Saitoh, Y.; et al. Powerful Homeostatic Control of Oligodendroglial Lineage by PDGFRalpha in Adult Brain. Cell Rep. 2019, 27, 1073–1089.e5. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, T.; Molnar, Z.; Nakano-Doi, A.; Taguchi, A.; Saino, O.; Kubo, S.; Clausen, M.; Yoshikawa, H.; Nakagomi, N.; Matsuyama, T. Ischemia-induced neural stem/progenitor cells in the pia mater following cortical infarction. Stem Cells Dev. 2011, 20, 2037–2051. [Google Scholar] [CrossRef] [PubMed]
- Day-Lollini, P.A.; Stewart, G.R.; Taylor, M.J.; Johnson, R.M.; Chellman, G.J. Hyperplastic Changes within the Leptomeninges of the Rat and Monkey in Response to Chronic Intracerebroventricular Infusion of Nerve Growth Factor. Exp. Neurol. 1997, 145, 24–37. [Google Scholar] [CrossRef]
- Parr, A.M.; Tator, C.H.; Keating, A. Bone marrow-derived mesenchymal stromal cells for the repair of central nervous system injury. Bone Marrow Transpl. 2007, 40, 609–619. [Google Scholar] [CrossRef]
- Decimo, I.; Bifari, F.; Rodriguez, F.J.; Malpeli, G.; Dolci, S.; Lavarini, V.; Pretto, S.; Vasquez, S.; Sciancalepore, M.; Montalbano, A.; et al. Nestin- and doublecortin-positive cells reside in adult spinal cord meninges and participate in injury-induced parenchymal reaction. Stem Cells 2011, 29, 2062–2076. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Kuhn, H.G.; Gage, F.H. More hippocampal neurons in adult mice living in an enriched environment. Nature 1997, 386, 493–495. [Google Scholar] [CrossRef]
- Zhou, Q.G.; Lee, D.; Ro, E.J.; Suh, H. Regional-specific effect of fluoxetine on rapidly dividing progenitors along the dorsoventral axis of the hippocampus. Sci. Rep. 2016, 6, 35572. [Google Scholar] [CrossRef]
- Kempermann, G. Environmental enrichment, new neurons and the neurobiology of individuality. Nat. Rev. Neurosci. 2019, 20, 235–245. [Google Scholar] [CrossRef]
- Young, D.; Lawlor, P.A.; Leone, P.; Dragunow, M.; During, M.J. Environmental enrichment inhibits spontaneous apoptosis, prevents seizures and is neuroprotective. Nat. Med. 1999, 5, 448–453. [Google Scholar] [CrossRef]
- Mercier, F. Fractones: Extracellular matrix niche controlling stem cell fate and growth factor activity in the brain in health and disease. Cell Mol. Life Sci. 2016, 73, 4661–4674. [Google Scholar] [CrossRef]
- Ziv, Y.; Ron, N.; Butovsky, O.; Landa, G.; Sudai, E.; Greenberg, N.; Cohen, H.; Kipnis, J.; Schwartz, M. Immune cells contribute to the maintenance of neurogenesis and spatial learning abilities in adulthood. Nat. Neurosci. 2006, 9, 268–275. [Google Scholar] [CrossRef]
- Mori, T.; Tanaka, K.; Buffo, A.; Wurst, W.; Kuhn, R.; Gotz, M. Inducible gene deletion in astroglia and radial glia—A valuable tool for functional and lineage analysis. Glia 2006, 54, 21–34. [Google Scholar] [CrossRef]
- Nakamura, T.; Colbert, M.C.; Robbins, J. Neural crest cells retain multipotential characteristics in the developing valves and label the cardiac conduction system. Circ. Res. 2006, 98, 1547–1554. [Google Scholar] [CrossRef]
- Balcombe, J.; Barnard, N.; Sandusky, C. Laboratory Routines Cause Animal Stress. Contemp. Top. Lab. Anim. Sci. 2004, 6, 42–51. [Google Scholar]
- Bonnichsen, M.; Dragsted, N.; Hansen, A.K. The welfare impact of gavaging laboratory rats. Anim. Welf. 2005, 14, 223–227. [Google Scholar]
- Brown, A.P.; Dinger, N.; Levine, B.S. Stress Produced by Gavage Administration in the Rat. Contemp. Top. Lab. Anim. Sci. 2000, 39, 17–21. [Google Scholar] [PubMed]
- Cazorla, M.; Premont, J.; Mann, A.; Girard, N.; Kellendonk, C.; Rognan, D. Identification of a low-molecular weight TrkB antagonist with anxiolytic and antidepressant activity in mice. J. Clin. Investig. 2011, 121, 1846–1857. [Google Scholar] [CrossRef] [PubMed]
- Moy, J.K.; Szabo-Pardi, T.; Tillu, D.V.; Megat, S.; Pradhan, G.; Kume, M.; Asiedu, M.N.; Burton, M.D.; Dussor, G.; Price, T.J. Temporal and sex differences in the role of BDNF/TrkB signaling in hyperalgesic priming in mice and rats. Neurobiol. Pain 2019, 5, 100024. [Google Scholar] [CrossRef]
- Tomassoni-Ardori, F.; Fulgenzi, G.; Becker, J.; Barrick, C.; Palko, M.E.; Kuhn, S.; Koparde, V.; Cam, M.; Yanpallewar, S.; Oberdoerffer, S.; et al. Rbfox1 up-regulation impairs BDNF-dependent hippocampal LTP by dysregulating TrkB isoform expression levels. eLife 2019, 8, e49673. [Google Scholar] [CrossRef]
- Formaggio, E.; Fazzini, F.; Dalfini, A.C.; di Chio, M.; Cantu, C.; Decimo, I.; Fiorini, Z.; Fumagalli, G.; Chiamulera, C. Nicotine increases the expression of neurotrophin receptor tyrosine kinase receptor A in basal forebrain cholinergic neurons. Neuroscience 2010, 166, 580–589. [Google Scholar] [CrossRef]
- Meeker, R.B.; Williams, K.S. The p75 neurotrophin receptor: At the crossroad of neural repair and death. Neural Regen. Res. 2015, 10, 721–725. [Google Scholar] [CrossRef]
- Khodanovich, M.; Kisel, A.; Kudabaeva, M.; Chernysheva, G.; Smolyakova, V.; Krutenkova, E.; Wasserlauf, I.; Plotnikov, M.; Yarnykh, V. Effects of Fluoxetine on Hippocampal Neurogenesis and Neuroprotection in the Model of Global Cerebral Ischemia in Rats. Int. J. Mol. Sci. 2018, 19, 162. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. Object Burying Test for Assessment of Obsessive Compulsive Behaviors in Mice. Methods Mol. Biol. 2019, 1916, 81–85. [Google Scholar]
- De Brouwer, G.; Fick, A.; Harvey, B.H.; Wolmarans, W. A critical inquiry into marble-burying as a preclinical screening paradigm of relevance for anxiety and obsessive-compulsive disorder: Mapping the way forward. Cogn. Affect. Behav. Neurosci. 2019, 19, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Sohur, U.S.; Emsley, J.G.; Mitchell, B.D.; Macklis, J.D. Adult neurogenesis and cellular brain repair with neural progenitors, precursors and stem cells. Philos. Trans. R. Soc. Lond B Biol. Sci. 2006, 361, 1477–1497. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.L.; Song, H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Csaba, Z.; Peineau, S.; Srivastava, R.; Rasika, S.; Mani, S.; Gressens, P.; El Ghouzzi, V. Endogenous cerebellar neurogenesis in adult mice with progressive ataxia. Ann. Clin. Transl. Neurol. 2014, 1, 968–981. [Google Scholar] [CrossRef]
- Nakagomi, T.; Molnar, Z.; Taguchi, A.; Nakano-Doi, A.; Lu, S.; Kasahara, Y.; Nakagomi, N.; Matsuyama, T. Leptomeningeal-derived doublecortin-expressing cells in poststroke brain. Stem Cells Dev. 2012, 21, 2350–2354. [Google Scholar] [CrossRef]
- Slaker, M.; Barnes, J.; Sorg, B.A.; Grimm, J.W. Impact of Environmental Enrichment on Perineuronal Nets in the Prefrontal Cortex following Early and Late Abstinence from Sucrose Self-Administration in Rats. PLoS ONE 2016, 11, e0168256. [Google Scholar] [CrossRef] [PubMed]
- Kerever, A.; Schnack, J.; Vellinga, D.; Ichikawa, N.; Moon, C.; Arikawa-Hirasawa, E.; Efird, J.T.; Mercier, F. Novel extracellular matrix structures in the neural stem cell niche capture the neurogenic factor fibroblast growth factor 2 from the extracellular milieu. Stem Cells 2007, 25, 2146–2157. [Google Scholar] [CrossRef]
- Bergami, M.; Rimondini, R.; Santi, S.; Blum, R.; Götz, M.; Canossa, M. Deletion of TrkB in adult progenitors alters newborn neuron integration into hippocampal circuits and increases anxiety-like behavior. PNAS 2008, 105, 15570–15575. [Google Scholar] [CrossRef]
- Benedetti, B.; Dannehl, D.; Konig, R.; Coviello, S.; Kreutzer, C.; Zaunmair, P.; Jakubecova, D.; Weiger, T.M.; Aigner, L.; Nacher, J.; et al. Functional Integration of Neuronal Precursors in the Adult Murine Piriform Cortex. Cereb. Cortex 2020, 30, 1499–1515. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, C.; Ghibaudi, M.; Bonfanti, L. Newly Generated and Non-Newly Generated “Immature” Neurons in the Mammalian Brain: A Possible Reservoir of Young Cells to Prevent Brain Aging and Disease? J. Clin. Med. 2019, 8, 685. [Google Scholar] [CrossRef]
- La Rosa, C.; Cavallo, F.; Pecora, A.; Chincarini, M.; Ala, U.; Faulkes, C.G.; Nacher, J.; Cozzi, B.; Sherwood, C.C.; Amrein, I.; et al. Phylogenetic variation in cortical layer II immature neuron reservoir of mammals. eLife 2020, 9, e55456. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, L.C.; Rompani, S.B.; Parraguez, J.I.; Obernier, K.; Romero, R.; Cepko, C.L.; Alvarez-Buylla, A. Embryonic Origin of Postnatal Neural Stem Cells. Cell 2015, 161, 1644–1655. [Google Scholar] [CrossRef]
- Jhaveri, D.J.; O’Keeffe, I.; Robinson, G.J.; Zhao, Q.Y.; Zhang, Z.H.; Nink, V.; Narayanan, R.K.; Osborne, G.W.; Wray, N.R.; Bartlett, P.F. Purification of neural precursor cells reveals the presence of distinct, stimulus-specific subpopulations of quiescent precursors in the adult mouse hippocampus. J. Neurosci. 2015, 35, 8132–8144. [Google Scholar] [CrossRef] [PubMed]
- Ottone, C.; Krusche, B.; Whitby, A.; Clements, M.; Quadrato, G.; Pitulescu, M.E.; Adams, R.H.; Parrinello, S. Direct cell-cell contact with the vascular niche maintains quiescent neural stem cells. Nat. Cell Biol. 2014, 16, 1045–1056. [Google Scholar] [CrossRef]
- Luzzati, F.; Nato, G.; Oboti, L.; Vigna, E.; Rolando, C.; Armentano, M.; Bonfanti, L.; Fasolo, A.; Peretto, P. Quiescent neuronal progenitors are activated in the juvenile guinea pig lateral striatum and give rise to transient neurons. Development 2014, 141, 4065–4075. [Google Scholar] [CrossRef]
- Paredes, M.F.; James, D.; Gil-Perotin, S.; Kim, H.; Cotter, J.A.; Ng, C.; Sandoval, K.; Rowitch, D.H.; Xu, D.; McQuillen, P.S.; et al. Extensive migration of young neurons into the infant human frontal lobe. Science 2016, 354, aaf7073. [Google Scholar] [CrossRef]
- Kempermann, G. A Back Door to Cortical Development. Cell Stem Cell 2017, 20, 295–296. [Google Scholar] [CrossRef][Green Version]
- Lopez, M.F.; Laber, K. Impact of social isolation and enriched environment during adolescence on voluntary ethanol intake and anxiety in C57BL/6J mice. Physiol. Behav. 2015, 148, 151–156. [Google Scholar] [CrossRef]
- Baek, I.S.; Park, J.Y.; Han, P.L. Chronic Antidepressant Treatment in Normal Mice Induces Anxiety and Impairs Stress-coping Ability. Exp. Neurobiol. 2015, 24, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Procedure | Mouse Strain | Sex | Age at Time of Sacrifice | IF | WB | Rt-PCR |
|---|---|---|---|---|---|---|
| Enriched Environment | CD1 | M | 8 weeks | CTRL n = 5 EE n = 6 | N.A. | N.A. |
| Enriched Environment on Lineage Tracing Model | GLAST-GFP [32,33] | M | 9–12 weeks | CTRL n = 3 EE n = 4 | N.A. | N.A. |
| Enriched Environment with Administration of TrkB Inhibitor ANA-12 | CD1 | M | 8 weeks | NO EE VEH n = 3 EE VEH n = 3 EE ANA-12 n = 3 NO EE ANA-12 n = 3 | NO EE VEH n = 3 EE VEH n = 2 EE ANA-12 n = 3 | NO EE VEH n = 4 EE VEH n = 5 EE ANA-12 n = 5 |
| Fluoxetine Administration | CD1 | M | 8 weeks | CTRL n = 3 FLUOX n = 3 | N.A. | N.A. |
| Toys | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | Day 7 |
|---|---|---|---|---|---|---|---|
| Running Wheel | X | X | X | X | X | X | X |
| Nesting Material | X | X | X | X | X | X | X |
| Carboard Rolls | X | X | X | ||||
| Marbles | X | X | X | ||||
| Stairs | X | X | X |
| Gene Name | Forward Sequence | Reverse Sequence |
|---|---|---|
| BDNF | CACATTACCTTCCTGCATCTGTTG | CTGGTGGAACATTGTGGCTTT |
| TUBB3 | ACAATGAGGCCTCCTCTCACA | TCCATCGTTCCAGGTTCCAA |
| ntrk2 | CACACACAGGGCTCCTTAAGG | TGGCGCAAAATGCACAGT |
| Slc1a3 | CGCGGTGATAATGTGGTATGC | GAGGCCGACAATGACTGTCA |
| Gapdh | GTCCGTCGTGGATCTGA | GATGCCTGCTTCACCACCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorzin, S.; Corsi, A.; Ciarpella, F.; Bottani, E.; Dolci, S.; Malpeli, G.; Pino, A.; Amenta, A.; Fumagalli, G.F.; Chiamulera, C.; et al. Environmental Enrichment Induces Meningeal Niche Remodeling through TrkB-Mediated Signaling. Int. J. Mol. Sci. 2021, 22, 10657. https://doi.org/10.3390/ijms221910657
Zorzin S, Corsi A, Ciarpella F, Bottani E, Dolci S, Malpeli G, Pino A, Amenta A, Fumagalli GF, Chiamulera C, et al. Environmental Enrichment Induces Meningeal Niche Remodeling through TrkB-Mediated Signaling. International Journal of Molecular Sciences. 2021; 22(19):10657. https://doi.org/10.3390/ijms221910657
Chicago/Turabian StyleZorzin, Stefania, Andrea Corsi, Francesca Ciarpella, Emanuela Bottani, Sissi Dolci, Giorgio Malpeli, Annachiara Pino, Alessia Amenta, Guido Franceso Fumagalli, Cristiano Chiamulera, and et al. 2021. "Environmental Enrichment Induces Meningeal Niche Remodeling through TrkB-Mediated Signaling" International Journal of Molecular Sciences 22, no. 19: 10657. https://doi.org/10.3390/ijms221910657
APA StyleZorzin, S., Corsi, A., Ciarpella, F., Bottani, E., Dolci, S., Malpeli, G., Pino, A., Amenta, A., Fumagalli, G. F., Chiamulera, C., Bifari, F., & Decimo, I. (2021). Environmental Enrichment Induces Meningeal Niche Remodeling through TrkB-Mediated Signaling. International Journal of Molecular Sciences, 22(19), 10657. https://doi.org/10.3390/ijms221910657