
The Small Molecule Fractions of Floccularia luteovirens Induce Apoptosis of NSCLC Cells through Activating Caspase-3 Activity


Abstract
:
1. Introduction
2. Results
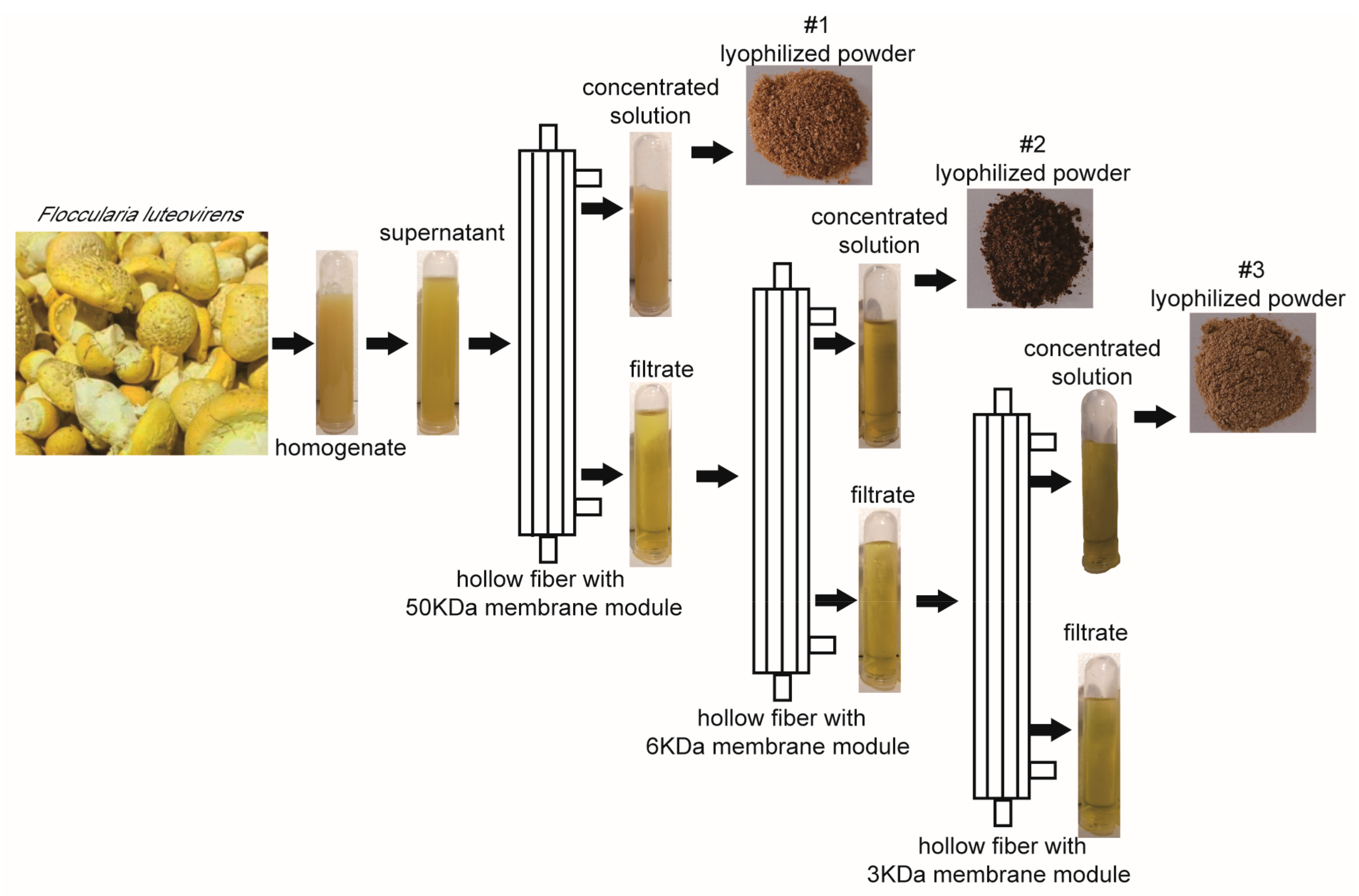
2.1. Extraction of Different Fractions of F. luteovirens
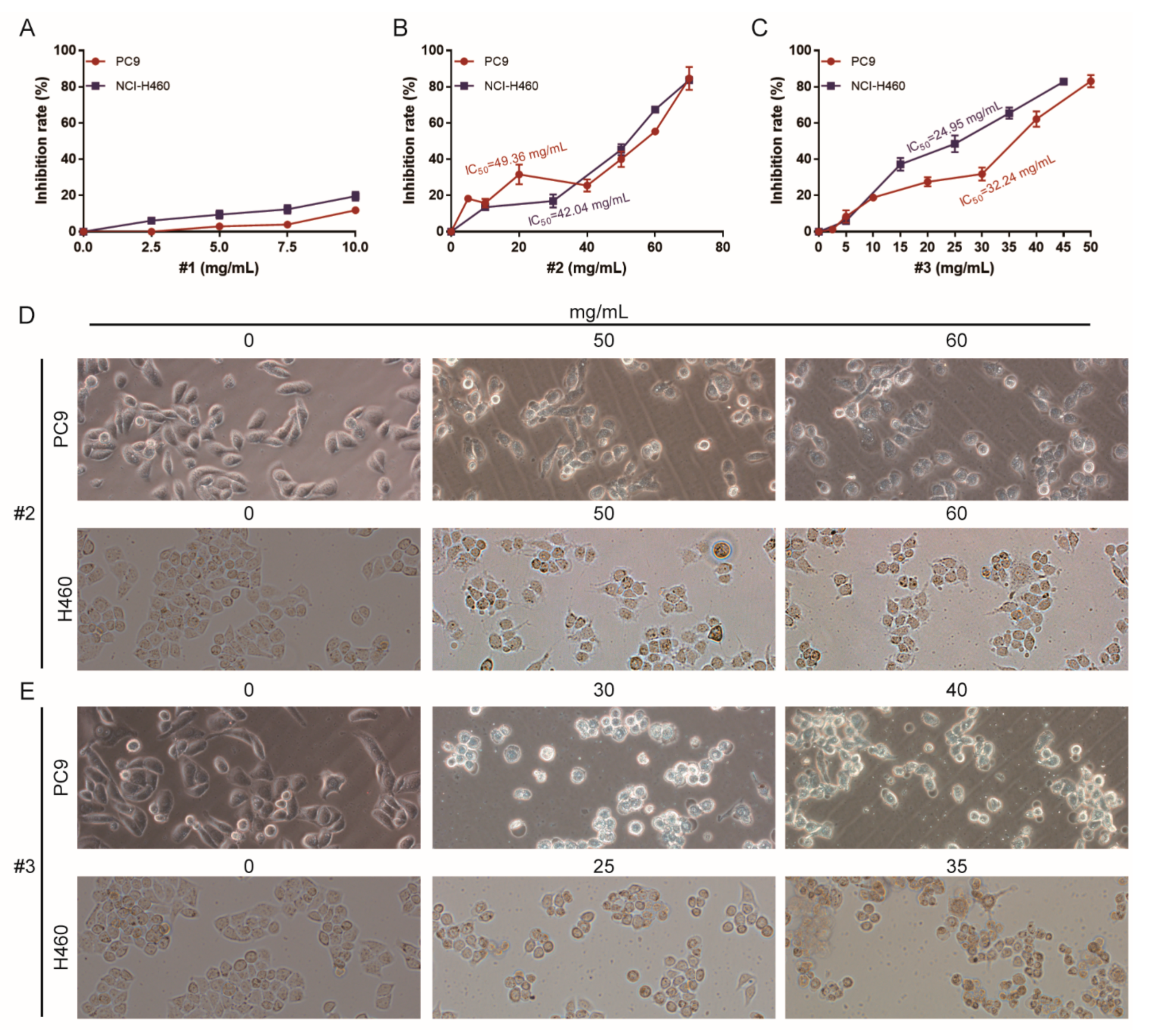
2.2. Cytotoxicity Analysis of Different Fractions of F. luteovirens on NSCLC Cells
2.3. The Apoptosis Effect of Different Fractions of F. luteovirens on NSCLC Cells
2.4. The Apoptosis Mechanism of Different Fractions of F. luteovirens on NSCLC Cells
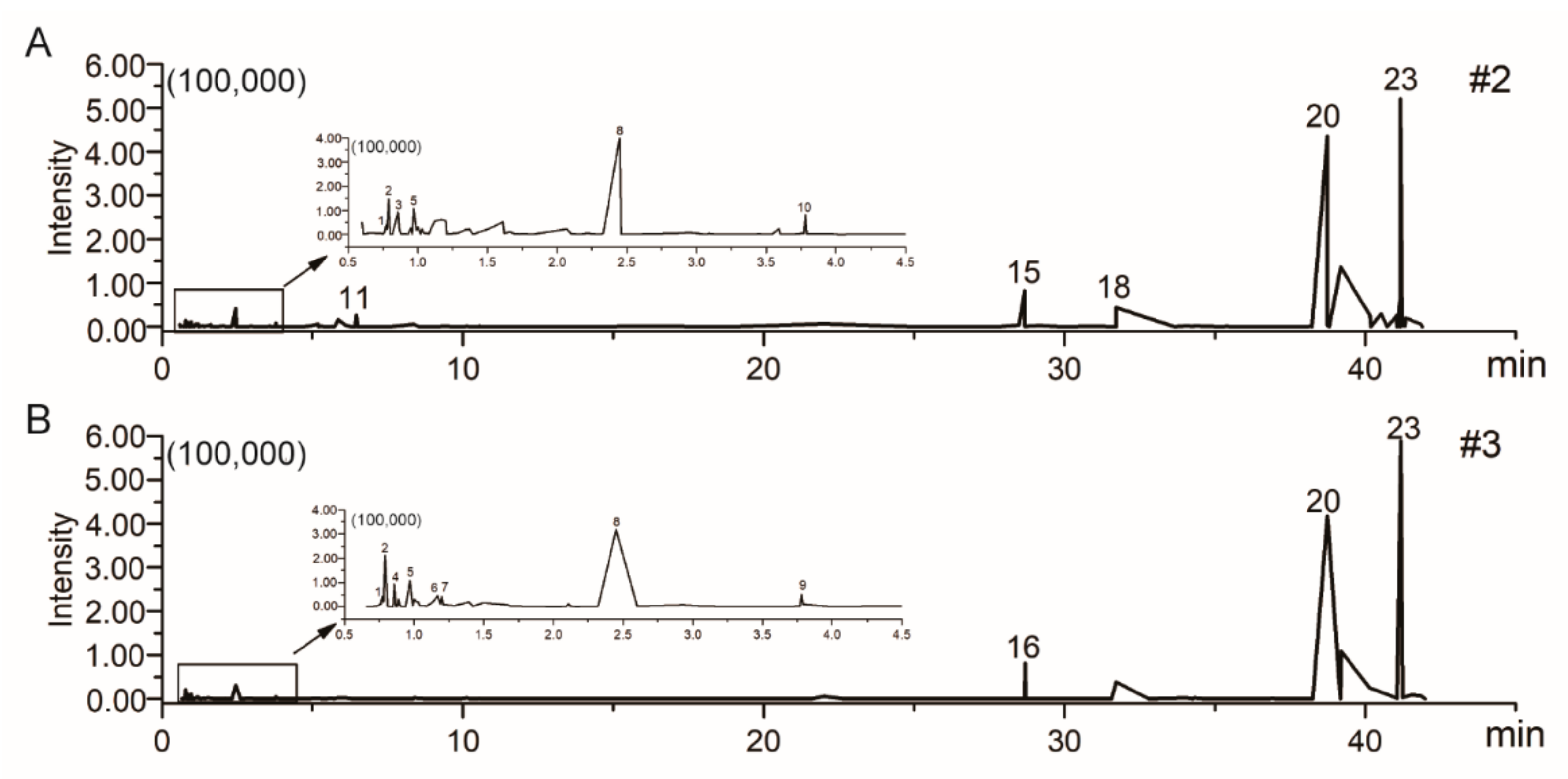
2.5. Chemical Profiling of Different Fractions of F. luteovirens
3. Discussion
4. Materials and Methods
4.1. Extraction of Different Fractions of F. luteovirens
4.2. Tumor Cell Culture
4.3. Sample Preparation for the Anti-Lung Cancer Detections
4.4. Tumor Cell Proliferative Inhibitory Assay
4.5. Tumor Cell Morphological Observation
4.6. Tumor Cell Apoptosis Assay
4.7. Western Blot Analysis
4.8. UFLC-Q-TOF-MS/MS Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nozaki, M.; Nishizuka, M. Repression of RhoJ expression promotes TGF-b-mediated EMT in human non-small-cell lung cancer A549 cells. Biochem. Biophys. Res. Commun. 2021, 566, 94–100. [Google Scholar] [CrossRef]
- Hu, Q.Y.; Zhang, X.K.; Wang, J.N.; Chen, H.X.; He, L.P.; Tang, J.S.; Yao, S.Y.; Liu, J. Malayoside, a cardenolide glycoside extracted from AntiaristoxicariaLesch, induces apoptosis in human non-small lung cancer cells via MAPK-Nur77 signaling pathway. Biochem. Pharmacol. 2021, 190, 114622. [Google Scholar] [CrossRef]
- Zhang, H.F.; Jiang, H.; Zhu, L.C.; Li, J.W.; Ma, S.L. Cancer-associated fibroblasts in non-small cell lung cancer: Recent advances and future perspectives. Cancer Lett. 2021, 514, 38–47. [Google Scholar] [CrossRef]
- Zhang, X.F.; Liu, M.; Yang, F.; Ma, L. Antiproliferative effect of cryptotanshinone against human non-small cell lung cancer cells through inactivation of lncRNA HOTAIR /p-Akt signaling Pathway. Arab. J. Chem. 2021, 14, 103150. [Google Scholar] [CrossRef]
- Roupas, P.; Keogh, J.; Noakes, M.; Margetts, C.; Taylor, P. The role of edible mushrooms in health: Evaluation of the evidence. J. Funct. Foods 2012, 4, 687–709. [Google Scholar] [CrossRef]
- Xue, Y.; Xie, J.; Xu, X.S.; Li, Y.; Hu, B.; Liang, J.; Li, X.D.; Qing, L.S. UPLC-QqQ/MS combined with similarity assessment of 17 nucleic acid constituents in 147 edible fungi from Sichuan Basin, China. Food Res. Int. 2019, 120, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Jacinto-Azevedo, B.; Valderrama, N.; Henriquez, K.; Aranda, M.; Aqueveque, P. Nutritional value and biological properties of Chilean wild and commercial edible mushrooms. Food Chem. 2021, 356, 129651. [Google Scholar] [CrossRef] [PubMed]
- Asaduzzaman, K.; Mousumi, T. Nutritional and medicinal importance of Pleurotus mushrooms: An overview. Food Rev. Int. 2012, 28, 313–329. [Google Scholar]
- Reis, F.S.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Chemical composition and nutritional value of the most widely appreciated cultivated mushrooms: An inter species comparative study. Food Chem. Toxicol. 2012, 50, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.K.; Ding, Z.C.; Gao, X.L.; Wang, Y.Y.; Yang, Y.; Wu, D.; Zhang, H.N. Comparative study of physicochemical properties and bioactivity of Hericium erinaceus polysaccharides at different solvent extractions. Carbohydr. Polym. 2018, 193, 373–382. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinial mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2012, 60, 258–274. [Google Scholar]
- Friedman, M. Mushroom polysaccharides: Chemistry and antiobesity, antidiabetes, anticancer, and antibiotic properties in cells, rodents, and humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.C.; Chen, Q.H.; Zhou, J.S.; Zhang, H.F.; Chen, H.Y. Improvement of exo-polysaccharides production and modeling kinetics by Armillaria luteovirens Sacc in submerged cultivation. LWT-Food Sci. Technol. 2008, 41, 1694–1700. [Google Scholar] [CrossRef]
- Chen, C.; Shao, Y.; Tao, Y.D.; Wen, H.X. Optimization of dynamic microwave-assisted extraction of Armillaria polysaccharides using RSM, and their biological activity. LWT-Food Sci. Technol. 2015, 64, 1263–1269. [Google Scholar] [CrossRef]
- Feng, K.; Liu, Q.H.; Ng, T.B.; Liu, H.Z.; Li, J.Q.; Chen, G.; Sheng, H.Y.; Xie, Z.L.; Wang, H.X. Isolation and characterization of a novel lectin from the mushroom Armillaria luteovirens. Biochem. Biophys. Res. Commun. 2006, 345, 1573–1578. [Google Scholar] [CrossRef]
- Yan, Y.; Yin, X.J.; Guo, H.M.; Wang, R.L.; Song, R.; Tian, Y.; Zhang, Z.J. Identification and comparative analysis of the major chemical constituents in the extracts of single Fuzi herb and Fuzi-Gancao herb-pair by UFLC-IT-TOF/MS. Chin. J. Nat. Med. 2014, 12, 542–553. [Google Scholar]
- Li, Y.F.; Ouyang, S.H.; Chang, Y.Q.; Wang, T.M.; Li, W.X.; Tian, H.Y.; Cao, H.; Kurihara, H.; He, R.R. A comparative analysis of chemical compositions in Camellia sinensis var. puanensis Kurihara, a novel Chinese tea, by HPLC and UFLC-Q-TOF-MS/MS. Food Chem. 2017, 216, 282–288. [Google Scholar]
- Zeng, X.; Su, W.W.; Bai, Y.; Chen, T.B.; Yan, Z.H.; Wang, J.W.; Su, M.; Zheng, Y.Y.; Peng, W.; Yao, H.L. Urinary metabolite profiling of flavonoids in Chinese volunteers after consumption of orange juice by UFLC-Q-TOF-MS/MS. J. Chromatogr. B 2017, 1061–1062, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.J.; Xie, G.Y.; Qin, S.H. The hydrophilic polypropylene/poly (ethylene-co-vinyl alcohol) hollow fiber membrane with bimodal microporous structure prepared by melt-spinning and stretching. Sep. Purif. Technol. 2021, 274, 118890. [Google Scholar] [CrossRef]
- Liu, Z.J.; Jiao, Y.C.; Lu, H.Y.; Shu, X.L.; Chen, Q.H. Chemical characterization, antioxidant properties and anticancer activity of exopolysaccharides from Floccularia luteovirens. Carbohydr. Polym. 2020, 229, 115432. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.Y.; Jiao, Y.D.; Yang, X.B.; Yuan, L.; Guo, J.J.; Zhao, Y. Antioxidant and antitumor effects of polysaccharides from the fungus Pleurotus abalonus. Chem.-Biol. Interact. 2015, 237, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Y.; Lu, Y.; Li, W.; Guo, J.L.; Dong, X.; Lin, B.; Chen, Y.; Xie, X.J.; Li, M.S. Hepatitis B virus X protein driven alpha fetoprotein expression to promote malignant behaviors of normal liver cells and hepatoma cells. J. Cancer 2016, 7, 935–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.M.; Chen, Y.; Chen, J.C.; Lin, T.Y.; Tseng, S.H. Tetrandrine induces apoptosis and growth suppression of colon cancer cells in mice. Cancer Lett. 2010, 287, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Katiyar, S.K. Honokiol Inhibits Non-Small Cell Lung Cancer Cell Migration by Targeting PGE2-Mediated Activation of β-Catenin Signaling. PLoS ONE 2013, 8, e60749. [Google Scholar] [CrossRef] [Green Version]
- Lin, B.; Zhu, M.Y.; Wang, W.T.; Li, W.; Dong, X.; Chen, Y.; Lu, Y.; Guo, J.L.; Li, M.S. Structural basis for alpha fetoprotein-mediated inhibition of caspase-3 activity in hepatocellular carcinoma cells. Int. J. Cancer 2017, 141, 1413–1421. [Google Scholar] [CrossRef]
- Li, J.N.; Wang, Q.H.; Wang, Z.B.; Cui, N.; Yang, B.Y.; Niu, W.Y.; Kuang, H.X. Tetrandrine inhibits colon carcinoma HT-29 cells growth via the Bcl-2/Caspase 3/PARP pathway and G1/S phase. Biosci. Rep. 2019, 39, BSR20182109. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Zhang, H.; Cao, M.; Wang, L.; Wu, S.; Fang, B. Auranofin Enhances Ibrutinib’s Anticancer Activity in EGFR-Mutant Lung Adenocarcinoma. Mol. Cancer Ther. 2018, 17, 2156–2163. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.C.; Zhu, Z.Y.; Liu, Y.L.; Sun, H.Q. Comparisons of the antitumor activity of polysaccharides from fermented mycelia and cultivated fruiting bodies of Cordyceps militaris in vitro. Int. J. Biol. Macromol. 2019, 130, 307–314. [Google Scholar] [CrossRef]
- Chen, T.B.; Su, W.W.; Yan, Z.H.; Wu, H.; Zeng, X.; Peng, W.; Gan, L.; Zhang, Y.H.; Yao, H.L. Identification of naringin metabolites mediated by human intestinal microbes with stable isotope-labeling method and UFLC-Q-TOF-MS/MS. J. Pharm. Biomed. Anal. 2018, 161, 262–272. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | [M + H]+(m/z) | Proposed Formula | Identification | Fragment Ions (MS2) m/z | Source | Type | ||
|---|---|---|---|---|---|---|---|---|
| Measured | Predicted | Error (ppm) | ||||||
| 1 | 156.0771 | 156.0770 | 0.64 | C6H9N3O2 | Histidine | 110.0706 | #2, #3 | amino acid |
| 2 | 175.1186 | 175.1184 | 1.14 | C6H14N4O2 | Arginine | 116.0706,129.1030 | #2, #3 | |
| 3 | 148.0613 | 148.0614 | −0.68 | C5H9NO4 | Glutamic acid | 130.0509,114.0555 | #2 | |
| 4 | 182.0813 | 182.0810 | 1.64 | C9H11NO3 | Tyrosine | 91.0542 | #3 | |
| 5 | 166.0866 | 166.0863 | 1.8 | C9H11NO2 | Phenylalanine | 120.0803 | #2, #3 | |
| 6 | 205.0969 | 205.0972 | 0.9 | C11H12N2O2 | Tryptophane | 188.0695 | #2 | |
| 7 | 267.1387 | 267.1379 | 2.9 | C18H18O2 | Honokiol | 115.0540 | #3 | phenylpropanoid |
| 8 | 268.1041 | 268.1040 | 0.1 | C5H5N5 | Adenine | 136.0599 | #2, #3 | nucleoside |
| 9 | 269.0870 | 269.0881 | 3.9 | C5H4N4O | Hypoxanthine | 137.0444 | #3 | |
| 10 | 279.1614 | 279.1617 | −1.07 | C16H22O4 | Dibutyl phthalate | 149.0230 | #3 | ester |
| 11 | 315.1074 | 315.1085 | 3.61 | C14H20O8 | Cimidahurinine | 149.0233 | #2 | glycoside |
| 12 | 827.4469 | 827.4424 | 5.4 | C42H66O16 | Esculentoside A | 425.2153 | #2 | |
| 13 | 336.3289 | 336.3294 | −1.5 | C22H41NO | N-Isobutyl-(2E,4E)- octadecadienamide | 135.1173 | #2, #3 | alkaloid |
| 14 | 339.2540 | 339.2530 | 2.8 | C20H34O4 | Kirenol | 221.2268 | #2, #3 | terpenoid |
| 15 | 437.2353 | 437.2322 | 6.9 | C27H32O5 | 5-O-Benzoyl-20- deoxyingenol | 133.0851 | #2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Gao, J.; Hou, L.; Gao, Y.; Sun, J.; Zhang, N.; Fan, B.; Wang, F. The Small Molecule Fractions of Floccularia luteovirens Induce Apoptosis of NSCLC Cells through Activating Caspase-3 Activity. Int. J. Mol. Sci. 2021, 22, 10609. https://doi.org/10.3390/ijms221910609
Li S, Gao J, Hou L, Gao Y, Sun J, Zhang N, Fan B, Wang F. The Small Molecule Fractions of Floccularia luteovirens Induce Apoptosis of NSCLC Cells through Activating Caspase-3 Activity. International Journal of Molecular Sciences. 2021; 22(19):10609. https://doi.org/10.3390/ijms221910609
Chicago/Turabian StyleLi, Shuying, Jie Gao, Lizhen Hou, Yaxin Gao, Jing Sun, Nana Zhang, Bei Fan, and Fengzhong Wang. 2021. "The Small Molecule Fractions of Floccularia luteovirens Induce Apoptosis of NSCLC Cells through Activating Caspase-3 Activity" International Journal of Molecular Sciences 22, no. 19: 10609. https://doi.org/10.3390/ijms221910609
APA StyleLi, S., Gao, J., Hou, L., Gao, Y., Sun, J., Zhang, N., Fan, B., & Wang, F. (2021). The Small Molecule Fractions of Floccularia luteovirens Induce Apoptosis of NSCLC Cells through Activating Caspase-3 Activity. International Journal of Molecular Sciences, 22(19), 10609. https://doi.org/10.3390/ijms221910609