
Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms


 ,
,  , ,
, ,  , , ,
, , ,  ,
,  , and
, and 
Abstract
1. Introduction
2. Plant Growth Promotion by Plant Growth-Promoting Rhizobacteria (PGPR): An Overview
3. Mechanisms of Action of PGPR in Plant Growth Promotion
3.1. Biological Nitrogen Fixation
3.2. Phosphorus Solubilization
3.3. Potassium Solubilization
3.4. Siderophore Production
3.5. Production of Phytohormones
3.6. Root Colonization and Increased Uptake of Plant Nutrients
4. Plant Growth-Promoting Rhizobacteria (PGPR) as Biocontrol Agents: An Overview
5. Mechanisms of Action of PGPR as Biocontrol Agents
5.1. Production of Antibiotics
5.2. Induction of Plant Systemic Resistance
5.3. Rhizosphere Competence and Root Colonization
5.4. Outcompetition and Direct Antagonism Against Pathogens
5.5. Synthesis of Hydrogen Cyanide (HCN)
5.6. Synthesis of Cell Wall Degrading Enzymes
6. Plant Growth-Promoting Rhizobacteria (PGPR) in Bioremediation: An Overview
7. Mechanisms of Action of PGPR in Bioremediation
7.1. Siderophores and Heavy Metal Removal
7.2. Biosurfactants in Heavy Metal Removal
7.3. Biosorption
7.4. ACC Deaminase Activity and Reduction in Ethylene Levels
7.5. Production of Exopolysaccharides and Polymeric Substances
8. Pros and Cons of Rhizosphere Bacteria for Agricultural Sustainability
8.1. Pros of Rhizosphere Bacterial Application
8.2. Cons of Rhizosphere Bacterial Application
8.2.1. Research and Development Constraints
8.2.2. Regulatory and Marketing Cnstraints
8.2.3. Field–Level Constraints
9. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bharti, N.; Barnawal, D. Amelioration of salinity stress by PGPR: ACC deaminase and ROS scavenging enzymes activity. In PGPR Amelioration in Sustainable Agriculture; Singh, A.K., Kumar, A., Singh, P.K., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 85–106. [Google Scholar] [CrossRef]
- Barrow, C. Biochar: Potential for countering land degradation and for improving agriculture. Appl. Geogr. 2012, 34, 21–28. [Google Scholar] [CrossRef]
- Niamat, B.; Naveed, M.; Ahmad, Z.; Yaseen, M.; Ditta, A.; Mustafa, A.; Rafique, M.; Bibi, R.; Sun, N.; Xu, M. Calcium-Enriched Animal Manure Alleviates the Adverse Effects of Salt Stress on Growth, Physiology and Nutrients Homeostasis of Zea mays L. Plants 2019, 8, 480. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Saharan, B.S.; Nehra, V. Plant growth promoting rhizobacteria: A critical review. Life Sci. Med. Res. 2011, 21, 1–30. [Google Scholar]
- Danish, S.; Zafar-Ul-Hye, M.; Mohsin, F.; Hussain, M. ACC-deaminase producing plant growth promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS ONE 2020, 15, e0230615. [Google Scholar] [CrossRef]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The good, the bad, and the ugly of rhizosphere microbiome. In Probiotics and Plant Health; Springer: Singapore, 2017; pp. 253–290. [Google Scholar]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.D.; Mahgoub, H.A.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; Elkelish, A. Comparative Study between Exogenously Applied Plant Growth Hormones versus Metabolites of Microbial Endophytes as Plant Growth-Promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Shahid, M.A.; Mustafa, A.; Sayyed, R.Z.; Curá, J.A. Insights into the Interactions among Roots, Rhizosphere, and Rhizobacteria for Improving Plant Growth and Tolerance to Abiotic Stresses: A Review. Cells 2021, 10, 1551. [Google Scholar] [CrossRef]
- Mustafa, A.; Naveed, M.; Saeed, Q.; Ashraf, M.N.; Hussain, A.; Abbas, T.; Kamran, M.; Minggang, X. Application potentials of plant growth promoting rhizobacteria and fungi as an alternative to conventional weed control methods. In Sustainable Crop Production; IntechOpen: London, UK, 2019. [Google Scholar]
- Mustafa, A.; Naveed, M.; Abbas, T.; Saeed, Q.; Hussain, A.; Ashraf, M.N.; Minggang, X. Growth response of wheat and associated weeds to plant antagonistic rhizobacteria and fungi. Ital. J. Agron. 2019, 14, 191–198. [Google Scholar] [CrossRef]
- Abbas, T.; Zahir, Z.A.; Naveed, M.; Abbas, S.; Alwahibi, M.S.; Elshikh, M.S.; Mustafa, A. Large scale screening of rhizospheric allelopathic bacteria and their potential for the biocontrol of wheat-associated weeds. Agronomy 2020, 10, 1469. [Google Scholar] [CrossRef]
- Ray, P.; Lakshmanan, V.; Labbé, J.L.; Craven, K.D. Microbe to microbiome: A paradigm shift in the application of microorganisms for sustainable agriculture. Front. Microbiol. 2020, 11, 3323. [Google Scholar] [CrossRef]
- Ahmad, M.; Naseer, I.; Hussain, A.; Zahid Mumtaz, M.; Mustafa, A.; Hilger, T.H.; Ahmad Zahir, Z.; Minggang, X. Appraising endophyte–plant symbiosis for improved growth, nodulation, nitrogen fixation and abiotic stress tolerance: An experimental investigation with chickpea (Cicer arietinum L.). Agronomy 2019, 9, 621. [Google Scholar] [CrossRef]
- Nazli, F.; Mustafa, A.; Ahmad, M.; Hussain, A.; Jamil, M.; Wang, X.; Shakeel, Q.; Imtiaz, M.; El-Esawi, M.A. A Review on Practical Application and Potentials of Phytohormone-Producing Plant Growth-Promoting Rhizobacteria for Inducing Heavy Metal Tolerance in Crops. Sustainability 2020, 12, 9056. [Google Scholar] [CrossRef]
- Haider, F.U.; Ejaz, M.; Cheema, S.A.; Khan, M.I.; Zhao, B.; Cai, L.; Salim, M.A.; Naveed, M.; Khan, N.; Núñez-Delgado, A.; et al. Phytotoxicity of petroleum hydrocarbons: Sources, impacts and remediation strategies. Environ. Res. 2021, 197, 111031. [Google Scholar] [CrossRef]
- Goudaa, S.; Kerryb, R.G.; Dasc, G.; Paramithiotisd, S.; Shine, H.S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Umar, W.; Ayub, M.A.; ur Rehman, M.Z.; Ahmad, H.R.; Farooqi, Z.U.R.; Shahzad, A.; Rehman, U.; Mustafa, A.; Nadeem, M. Nitrogen and phosphorus use efficiency in agroecosystems. In Resources Use Efficiency in Agriculture; Springer: Singapore, 2020; pp. 213–257. [Google Scholar]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef]
- Raza, W.; Ling, N.; Yang, L.; Huang, Q.; Shen, Q. Response of tomato wilt pathogen Ralstonia solanacearum to the volatile organic compounds produced by a biocontrol strain Bacillus amyloliquefaciens SQR-9. Sci. Rep. 2016, 6, 24856. [Google Scholar] [CrossRef]
- Raza, W.; Yousaf, S.; Rajer, F.U. Plant growth promoting activity of volatile organic compounds produced by Bio-control strains. Sci. Lett. 2016, 4, 40–43. [Google Scholar]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzianatronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef]
- Ramakrishna, W.; Rathore, P.; Kumari, R.; Yadav, R. Brown gold of marginal soil: Plant growth promoting bacteria to overcome plant abiotic stress for agriculture, biofuels and carbon sequestration. Sci. Total Environ. 2020, 711, 135062. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in Chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef]
- Gupta, A.; Gopal, M.; Thomas, G.V.; Manikandan, V.; Gajewski, J.; Thomas, G.; Seshagiri, S.; Schuster, S.C.; Rajesh, P.; Gupta, R. Whole genome sequencing and analysis of plant growth promoting bacteria isolated from the rhizosphere of plantation crops coconut, cocoa and arecanut. PLoS ONE 2014, 9, e104259. [Google Scholar] [CrossRef]
- Verma, P.P.; Shelake, R.M.; Das, S.; Sharma, P.; Kim, J.Y. Plant growth-promoting rhizobacteria (PGPR) and fungi (PGPF): Potential biological control agents of diseases and pests. In Microbial Interventions in Agriculture and Environment; Springer: Singapore, 2019; pp. 281–311. [Google Scholar]
- Kumari, A.; Kumar, R. Exploring phyllosphere bacteria for growth promotion and yield of potato (Solanum tuberosum L.). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1065–1071. [Google Scholar] [CrossRef]
- Shivakumar, B. Biocontrol Potential and Plant Growth Promotional Activity of Fluorescent Pseudomonads of Western Ghats. Ph.D. Dissertation, UAS, Dharwad, India, 2007. [Google Scholar]
- Mohammed, A.F.; Oloyede, A.R.; Odeseye, A.O. Biological control of bacterial wilt of tomato caused by Ralstonia solanacearum using Pseudomonas species isolated from the rhizosphere of tomato plants. Arch. Phytopathol. Plant Prot. 2020, 53, 1–16. [Google Scholar] [CrossRef]
- Larkin, R.P. Biological control of soil borne diseases in organic potato production using hypovirulent strains of Rhizoctonia solani. Biol. Agric. Hortic. 2020, 36, 1–11. [Google Scholar] [CrossRef]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef]
- Kim, K.Y.D.; Jordan, D.; McDonald, G.A. Solubilization of hydroxyapatite. Enterobacter agglomerans and cloned Escherichia coli in culture medium. Biol. Fertil. Soils 1997, 24, 347–352. [Google Scholar] [CrossRef]
- Ladha, J.K.; de Bruijn, F.J.; Malik, K.A. Introduction: Assessing opportunities for nitrogen fixation in rice-a frontier project. Plant Soil 1997, 124, 1–10. [Google Scholar] [CrossRef]
- Raymond, J.; Siefert, J.L.; Staples, C.R.; Blankenship, R.E. The natural history of nitrogen fixation. Mol. Biol. Evol. 2004, 21, 541–554. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Ahmad, M.; Khan, M.S. Effects of pesticides on plant growth promoting traits of Mesorhizobium strain MRC4. J. Saudi Soc. Agric. Sci. 2012, 11, 63–71. [Google Scholar]
- Zahran, H.H. Rhizobia from wild legumes: Diversity, taxonomy, ecology, nitrogen fixation and biotechnology. J. Biotechnol. 2001, 91, 143–153. [Google Scholar] [CrossRef]
- Giordano, W.; Hirsch, A.M. The expression of MaEXP1, a Melilotus alba expansin gene, is upregulated during the sweetclover-Sinorhizobium meliloti interaction. Mol. Plant Microbe Interact. 2004, 17, 613–622. [Google Scholar] [CrossRef]
- Marroquí, S.; Zorreguieta, A.; Santamaría, C. Enhanced symbiotic performance by Rhizobium tropici glycogen synthase mutants. J. Bacteriol. 2001, 183, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. The enhancement of plant growth by free living bacteria. Can. J. Microbiol. 1995, 41, 109114. [Google Scholar] [CrossRef]
- Parray, J.A.; Jan, S.; Kamili, A.N.; Qadri, R.A.; Egamberdieva, D.; Ahmad, P. Current perspectives on plant growth-promoting rhizobacteria. J. Plant Growth Regul. 2016, 35, 877–902. [Google Scholar] [CrossRef]
- Ma, W.; Penrose, D.M.; Glick, B.R. Strategies used by rhizobia to lower plant ethylene levels and increase nodulation. Can. J. Microbiol. 2002, 48, 947–954. [Google Scholar] [CrossRef]
- Ma, W.; Charles, T.C.; Glick, B.R. Expression of an exogenous 1 aminocyclopropane-1-carboxylate deaminase gene in Sinorhizobium meliloti increases its ability to nodulate alfalfa. Appl. Environ. Microbiol. 2004, 100, 5891–5897. [Google Scholar] [CrossRef] [PubMed]
- Dubeikovsky, A.N.; Mordukhova, E.A.; Kochetkov, V.V.; Polikarpova, F.Y.; Boronin, A.M. Growth promotion of blackcurrant softwood cuttings by recombinant strain Pseudomonas uorescens BSP53a synthesizing an increased amount of indole3-acetic acid. Soil Biol. Biochem. 1993, 25, 1277–1281. [Google Scholar] [CrossRef]
- Ke, X.; Feng, S.; Wang, J.; Lu, W.; Zhang, W.; Chen, M.; Lin, M. Effect of inoculation with nitrogen-fixing bacterium Pseudomonas stutzeri A1501 on maize plant growth and the microbiome indigenous to the rhizosphere. Syst. Appl. Microbiol. 2019, 42, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Valderrama, B.; Arredondo-Peter, R.; Soberón, M.; Mora, J.; Hernández, G. Rhizobium etli genetically engineered for the heterologous expression of Vitreoscilla sp. hemoglobin: Effects on free-living and symbiosis. Mol. Plant-Microbe Interact. 1999, 12, 1008–1015. [Google Scholar] [CrossRef]
- Liu, D.; Yan, R.; Fu, Y.; Wang, X.; Zhang, J.; Xiang, W. Antifungal, Plant Growth-Promoting, and Genomic Properties of an Endophytic Actinobacterium Streptomyces sp. NEAU-S7GS2. Front. Microbiol. 2019, 10, 2077. [Google Scholar] [CrossRef] [PubMed]
- Rima, F.S.; Biswas, S.; Sarker, P.K. Bacteria endemic to saline coastal belt and their ability to mitigate the effects of salt stress on rice growth and yields. Ann. Microbiol. 2018, 68, 525–535. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K. Fluorescent Pseudomonas sp. PF17 as an efficient plant growth regulator and biocontrol agent for sunflower crop under saline conditions. Symbiosis 2016, 68, 99–108. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef]
- Parmar, P.; Sindhu, S.S. Potassium solubilization by rhizosphere bacteria: Influence of nutritional and environmental conditions. J. Microbiol. Res. 2013, 3, 25–31. [Google Scholar]
- Gururani, M.A.; Upadhyaya, C.P.; Baskar, V.; Venkatesh, J.; Nookaraju, A.; Park, S.W. Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J. Plant Growth Regul. 2013, 32, 245–258. [Google Scholar] [CrossRef]
- Robin, A.; Mougel, C.; Siblot, S.; Vansuyt, G.; Mazurier, S.; Lemanceau, P. Effect of ferritin overexpression in tobacco on the structure of bacterial and pseudomonad communities associated with the roots. FEMS Microbiol. Ecol. 2006, 58, 492–502. [Google Scholar] [CrossRef]
- Vansuyt, G.; Robin, A.; Briat, J.F.; Curie, C.; Lemanceau, P. Iron acquisition from Fe-pyoverdine by Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2007, 20, 441–447. [Google Scholar] [CrossRef]
- Mounir, A.M.; Osman, Y.M.M.; Khalil, O.A. Impact of potassium solubilizing bacteria on growth and yield of garlic. Plant Arch. 2020, 20, 8374–8388. [Google Scholar]
- Elkhatib, H.A.; Saed, M.G.; Ramadan, A.M.; Ahmed, F.M. Impact of potassium fertilizer rates and potassium solubilizing bacteria inoculation on the growth, yield and quality of potato (Solanum tuberosum L.). Alex. Sci. Exch. J. 2019, 40, 127–141. [Google Scholar]
- Liu, D.; Lian, B.; Dong, H. Isolation of Paenibacillus sp. and assessment of its potential for enhancing mineral weathering. Geomicrobiol. J. 2012, 29, 413–421. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K. Role of salicylic acid from Pseudomonas aeruginosa PF23EPS+ in growth promotion of sunflower in saline soils infested with phytopathogen Macrophomina phaseolina. Environ. Sustain. 2018, 1, 49–59. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Jabborova, D.; Räsänen, L.A.; Liao, H. Coordination between Bradyrhizobium and Pseudomonas alleviates salt stress in soybean through altering root system architecture. J. Plant Interact. 2017, 12, 100–107. [Google Scholar] [CrossRef]
- Asari, S.; Danuse, T.; Jakub, R.; Ondrej, N.; David, V.P.; Sarosh, B.; Johan, M. Analysis of plant growth-promoting properties of Bacillus amyloliquefaciens UCMB5113 using Arabidopsis thaliana as host plant. Planta 2017, 245, 15–30. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.A.; Ali, H.M.; Alayafi, A.A. Serratia liquefaciens KM4 improves salt stress tolerance in maize by regulating redox potential, ion homeostasis, leaf gas exchange and stress-related gene expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [PubMed]
- López-Reyes, L.; Carcaño-Montiel, M.G.; Lilia, T.L.; Medina-de la Rosa, G.; Armando, T.H.R. Antifungal and growth-promoting activity of Azospirillumbrasilense in Zea mays L. ssp. mexicana. Arch. Phytopathol. Plant Prot. 2017, 50, 727–743. [Google Scholar] [CrossRef]
- Cavite, H.J.M.; Mactal, A.G.; Evangelista, E.V. Growth and Yield Response of Upland Rice to Application of Plant Growth-Promoting Rhizobacteria. J. Plant Growth Regul. 2021, 40, 494–508. [Google Scholar] [CrossRef]
- Sheng, M.; Jia, H.; Zhang, G.; Zeng, L.; Zhang, L.; Long, Y.; Lan, J.; Hu, Z.; Zeng, Z.; Wang, B.; et al. Siderophore Production by Rhizosphere Biological Control Bacteria Brevibacillus brevis GZDF3 of Pinellia ternata and Its Antifungal Effects on Candida albicans. J. Microbiol. Biotechnol. 2020, 30, 689–699. [Google Scholar] [CrossRef]
- Ghazy, N.; El Nahrawy, S. Siderophore production by Bacillus subtilis MF497446 and Pseudomonas koreensis MG209738 and their efficacy in controlling Cephalosporium maydis in maize plant. Arch. Microbiol. 2021, 203, 1195–1209. [Google Scholar] [CrossRef]
- Billah, M.; Khan, M.; Bano, A.; Hassan, T.U.; Munir, A.; Gurmani, A.R. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Li, M.; Whelan, M. Phosphorus activators contribute to legacy phosphorus availability in agricultural soils: A review. Sci. Total Environ. 2018, 612, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Fahad, S.; Khan, I.A.; Saeed, M.; Ihsan, M.Z.; Saud, S.; Riaz, M.; Wang, D.; Wu, C. Integration of poultry manure and phosphate solubilizing bacteria improved availability of Ca bound P in calcareous soils. 3 Biotech 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Alaylar, B.; Egamberdieva, D.; Gulluce, M.; Karadayi, M.; Arora, N.K. Integration of molecular tools in microbial phosphate solubilization research in agriculture perspective. World J. Microbiol. Biotechnol. 2020, 36, 1–12. [Google Scholar] [CrossRef]
- Pereira, S.I.; Castro, P.M. Phosphate-solubilizing rhizobacteria enhance Zea mays growth in agricultural P-deficient soils. Ecol. Eng. 2014, 73, 526–535. [Google Scholar] [CrossRef]
- FAO. World Fertilizer Trends and Outlook to 2020; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Dash, S.; Borah, S.S.; Kalamdhad, A.S. Study of the limnology of wetlands through a one-dimensional model for assessing the eutrophication levels induced by various pollution sources. Ecol. Model. 2020, 416, 108907. [Google Scholar] [CrossRef]
- Younas, F.; Mustafa, A.; Farooqi, Z.U.R.; Wang, X.; Younas, S.; Mohy-Ud-Din, W.; Ashir Hameed, M.; Mohsin Abrar, M.; Maitlo, A.A.; Noreen, S.; et al. Current and Emerging Adsorbent Technologies for Wastewater Treatment: Trends, Limitations, and Environmental Implications. Water 2021, 13, 215. [Google Scholar] [CrossRef]
- Loper, J.E.; Schroth, M.N. Influence of bacterial sources of indole-3-acetic acid on root elongation of sugar beet. Phytopathology 1986, 76, 386–389. [Google Scholar] [CrossRef]
- Muleta, A.; Tesfaye, K.; Selassie, T.H.; Cook, D.R.; Assefa, F. Phosphate solubilization and multiple plant growth promoting properties of Mesorhizobium species nodulating chickpea from acidic soils of Ethiopia. Arch. Microbiol. 2021, 203, 2129–2137. [Google Scholar] [CrossRef]
- Kafle, A.; Cope, K.; Raths, R.; Krishna, Y.J.; Subramanian, S.; Bucking, H.; Garcia, K. Harnessing Soil Microbes to Improve Plant Phosphate Efficiency in Cropping Systems. Agronomy 2019, 9, 127. [Google Scholar] [CrossRef]
- Singh, D.; Prasanna, R. Potential of microbes in the biofortification of Zn and Fe in dietary food grains. A review. Agron. Sustain. Dev. 2020, 40, 1–21. [Google Scholar] [CrossRef]
- Kalayu, G. Phosphate Solubilizing Microorganisms: Promising Approach as Biofertilizers. Int. J. Agron. 2019, 2019, 4917256. [Google Scholar] [CrossRef]
- Satyaprakash, M.; Nikitha, T.; Reddi, E.U.B.; Sadhana, B.; Vani, S.S. Phosphorous and phosphate solubilising bacteria and their role in plant nutrition. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2133–2144. [Google Scholar]
- Patel, S.T.; Minocheherhomji, F.P. Review: Plant Growth Promoting Rhizobacteria: Blessing to Agriculture. Int. J. Pure Appl. Biosci. 2018, 6, 481–492. [Google Scholar] [CrossRef]
- Pradhan, A.; Pahari, A.; Mohapatra, S.; Mishra, B.B. Phosphate-Solubilizing Microorganisms in Sustainable Agriculture: Genetic Mechanism and Application. In Advances in Soil Microbiology: Recent Trends and Future Prospects; Springer Nature Singapore Pte Ltd.: Singapore, 2017; pp. 81–97. [Google Scholar] [CrossRef]
- Castagno, L.N.; Sannazzaro, A.I.; Gonzalez, M.E.; Pieckenstain, F.L.; Estrella, M.J. Phosphobacteria as key actors to overcome phosphorus deficiency in plants. Ann. Appl. Biol. 2021, 178, 256–267. [Google Scholar] [CrossRef]
- Walpola, B.C.; Yoon, M.H. Isolation and characterization of phosphate solubilizing bacteria and their co-inoculation efficiency on tomato plant growth and phosphorous uptake. Afr. J. Microbiol. Res. 2013, 7, 266–275. [Google Scholar]
- Kumar, A.; Singh, V.K.; Tripathi, V.; Singh, P.P.; Singh, A.K. Plant growth-promoting rhizobacteria (PGPR): Perspective in agriculture under biotic and abiotic stress. In Crop Improvement through Microbial Biotechnology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 333–342. [Google Scholar]
- Nahas, E. Factors determining rock phosphate solubilization by microorganism isolated from soil. World J. Microbiol. Biotechnol. 1996, 12, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by rootinduced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Kim, J.; Rees, D.C. Nitrogenase and biological nitrogen fixation. Biochemistry 1994, 33, 389–397. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, W.H. Regulation of potassium transport and signaling in plants. Curr. Opin. Plant Biol. 2017, 39, 123–128. [Google Scholar] [CrossRef]
- Adams, E.; Shin, R. Transport, signaling, and homeostasis of potassium and sodium in plants. J. Integr. Plant Biol. 2014, 56, 231–249. [Google Scholar] [CrossRef]
- Zahoor, R.; Zhao, W.; Dong, H.; Snider, J.L.; Abid, M.; Iqbal, B.; Zhou, Z. Potassium improves photosynthetic tolerance to and recovery from episodic drought stress in functional leaves of cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 119, 21–32. [Google Scholar] [CrossRef]
- Filho, A.B.C.; Dutra, A.F.; Da Silva, G.S. Phosphate and potassium fertilization for radish grown in a latosol with a high content of these nutrients. Rev. Caatinga 2017, 30, 412–419. [Google Scholar] [CrossRef]
- Sparks, D.L.; Huang, P.M. Physical chemistry of soil potassium. In Potassium in Agriculture; Soil Science Society of America: Madison, WI, USA, 1985; pp. 201–276. [Google Scholar]
- Radulov, I.; Berbecea, A.; Imbrea, F.; Lato, A.; Crista, F.; Mergheş, P. Potassium in soil-plant-human system. Res. J. Agric. Sci. 2014, 46, 47–52. [Google Scholar]
- Sindhu, S.S.; Parmar, P.; Phour, M. Nutrient cycling: Potassium solubilization by microorganisms and improvement of crop growth. In Geomicrobiology and Biogeochemistry; Springer: Berlin/Heidelberg, Germany, 2014; pp. 175–198. [Google Scholar]
- Saiyad, S.A.; Jhala, Y.K.; Vyas, R.V. Comparative efficiency of five potash and phosphate solubilizing bacteria and their key enzymes useful for enhancing and improvement of soil fertility. Int. J. Sci. Res. 2015, 5, 1–6. [Google Scholar]
- Archana, D.S.; Nandish, M.S.; Savalagi, V.P.; Alagawadi, A.R. Characterization of potassium solubilizing bacteria (KSB) from rhizosphere soil. Bioinfolet-A Q. J. Life Sci. 2013, 10, 248–257. [Google Scholar]
- Rajawat, M.V.S.; Singh, S.; Tyagi, S.P.; Saxena, A.K. A modified plate assay for rapid screening of potassium-solubilizing bacteria. Pedosphere 2016, 26, 768–773. [Google Scholar] [CrossRef]
- Hu, X.; Chen, J.; Guo, J. Two phosphate-and potassium-solubilizing bacteria isolated from Tianmu Mountain, Zhejiang, China. World J. Microbiol. Biotechnol. 2006, 22, 983–990. [Google Scholar] [CrossRef]
- Sheng, X.F. Growth promotion and increased potassium uptake of cotton and rape by a potassium releasing strain of Bacillus edaphicus. Soil Biol. Biochem. 2005, 37, 1918–1922. [Google Scholar] [CrossRef]
- Prajapati, K.; Sharma, M.C.; Modi, H.A. Growth promoting effect of potassium solubilizing microorganisms on okra (Abelmoscus Esculantus). Int. J. Agri. Sci. Res. 2013, 1, 181–188. [Google Scholar]
- Huang, Z.; He, L.; Sheng, X.; He, Z. Weathering of potash feldspar by Bacillus sp. L11. Wei sheng wuxue bao. Acta Microbiol. Sin. 2013, 53, 1172–1178. [Google Scholar]
- Meena, V.S.; Maurya, B.R.; Verma, J.P. Does a rhizospheric microorganism enhance K+ availability in agricultural soils? Microbiol. Res. 2014, 169, 337–347. [Google Scholar] [CrossRef]
- Abou-el-Seoud, I.; Abdel-Megeed, A. Impact of rock materials and biofertilizations on P and K availability for maize (Zea Maize) under calcareous soil conditions. Saudi J. Biol. Sci. 2012, 19, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Shelobolina, E.; Xu, H.; Konishi, H.; Kukkadapu, R.; Wu, T.; Blöthe, M.E. Microbial lithotrophic oxidation of structural Fe (II) in biotite. Appl. Environ. Microbiol. 2012, 78, 5746–5752. [Google Scholar] [CrossRef]
- Sheng, X.F.; Gong, J.X. Increased degradation of phenanthrene in soil by Pseudomonas sp. GF3 in the presence of wheat. Soil Biol. Biochem. 2006, 38, 2587–2592. [Google Scholar] [CrossRef]
- Padhan, D.; Sen, A.; Adhikary, S.; Kundu, R.; Yadav, V.K. Chapter-3 Potassium Solubilisation in Soils: Mechanisms, Effect on Plant Growth and Future Prospects. Curr. Res. Soil Fertil. 2019, 19, 37. [Google Scholar]
- Spaepen, S.; Vanderleyden, J. Auxin and plant-microbe interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef]
- Wang, J.G.; Zhang, F.S.; Zhang, X.L.; Cao, Y.P. Release of potassium from K-bearing minerals: Effect of plant roots under P deficiency. Nutr. Cycl. Agroecosyst. 2000, 56, 45–52. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A.; Hosseini, H.M. Indole-3-acetic acid and 1-aminocyclopropane-1-carboxylate deaminase: Bacterial traits required in rhizosphere, rhizoplane and/or endophytic competence by beneficial bacteria. In Bacterial Metabolites in Sustainable Agroecosystem; Springer: Berlin/Heidelberg, Germany, 2015; pp. 183–258. [Google Scholar]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Rasouli-Sadaghiani, M.; Malakouti, M.J.; Khavazi, K.; Miransari, M. Siderophore Efficacy of Fluorescent Pseudomonades Affecting Labeled Iron (59Fe) Uptake by Wheat (Triticumaestivum L.) Genotypes Differing in Fe Efficiency. In Use of Microbes for the Alleviation of Soil Stresses; Springer: New York, NY, USA, 2014; pp. 121–132. [Google Scholar]
- Subramanium, N.; Sundaram, L. Siderophore producing Pseudomonas spp. isolated from rhizospheric soil and enhancing iron content in Arachis hypogaea L. plant. J. Agric. Technol. 2020, 16, 429–442. [Google Scholar]
- Reed, S.C.; Yang, X.; Thornton, P.E. Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor. New Phytol. 2015, 208, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering with Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Grobelak, A.; Hiller, J. Bacterial siderophores promote plant growth: Screening of catechol and hydroxamate siderophores. Int. J. Phytorem. 2017, 19, 825–833. [Google Scholar] [CrossRef]
- Sharma, A.; Johri, B.; Sharma, A.; Glick, B. Plant growth-promoting bacterium Pseudomonas spp. strain GRP3 influences iron acquisition in mung bean (Vigna radiata L. Wilzeck). Soil Biol. Biochem. 2003, 35, 887–894. [Google Scholar] [CrossRef]
- Braud, A.; Jézéquel, K.; Léger, M.A. Lebeau T, Siderophore production by using free and immobilized cells of two pseudomonads cultivated in a medium enriched with Fe and/or toxic metals (Cr, Hg, Pb). Biotechnol. Bioeng. 2006, 94, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Damam, M.; Kaloori, K.; Gaddam, B.; Kausar, R. Plant growth promoting substances (phytohormones) produced by rhizobacterial strains isolated from the rhizosphere of medicinal plants. Int. J. Pharm. Sci. Rev. 2016, 37, 130–136. [Google Scholar]
- Sureshbabu, K.; Amaresan, N.; Kumar, K. Amazing multiple function properties of plant growth promoting rhizobacteria in the rhizosphere soil. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 661–683. [Google Scholar] [CrossRef]
- Spence, C.; Bais, H. Role of plant growth regulators as chemical signals in plant–microbe interactions: A double edged sword, Curr. Opin. Plant Biol. 2015, 27, 52–58. [Google Scholar] [CrossRef]
- Kumar, K.V.; Srivastava, S.; Singh, N.; Behl, H. Role of metal resistant plant growth promoting bacteria in ameliorating fly ash to the growth of Brassica juncea. J. Hazard. Mater. 2009, 170, 51–57. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Iqrar, I. Selective isolation and characterization of agriculturally beneficial endophytic bacteria from wild hemp using canola. Pak. J. Bot. 2015, 47, 1999–2008. [Google Scholar]
- Spaepen, S.; Bossuyt, S.; Engelen, K.; Marchal, K.; Vanderleyden, J. Phenotypical and molecular responses of A rabidopsis thaliana roots as a result of inoculation with the auxin-producing bacterium A zospirillumbrasilense. New Phytol. 2014, 201, 850–861. [Google Scholar] [CrossRef]
- Llorente, B.E.; Alasia, M.A.; Larraburu, E.E. Biofertilization with Azospirillumbrasilense improves in vitro culture of Handroanthusochraceus, a forestry, ornamental and medicinal plant. New Biotechnol. 2016, 33, 32–40. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Kumar, A.; Vandana, R.S.; Singh, M.; Pandey, K.D. Plant growth promoting rhizobacteria (PGPR). A promising approach for disease management. In Microbes and Environmental Management; Singh, J.S., Singh, D.P., Eds.; Studium Press: New Delhi, India, 2015; pp. 195–209. [Google Scholar]
- Jha, C.K.; Saraf, M. Plant growth promoting rhizobacteria (PGPR): A review. E3 J. Agric. Res. Dev. 2015, 5, 108–119. [Google Scholar]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moenne-Loccoz, Y.; Muller, D. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability–A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Alías-Villegas, C.; Bellogín, R.; Del Cerro, P.; Espuny, M.; Jiménez-Guerrero, I. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef]
- Heydarian, Z.; Yu, M.; Gruber, M.; Glick, B.R.; Zhou, R.; Hegedus, D.D. Inoculation of soil with plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression of the corresponding acds gene in transgenic plants increases salinity tolerance in camelina sativa. Front. Microbiol. 2016, 7, 1966. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Zhang, R.; Vivanco, J.M.; Shen, Q. The unseen rhizosphere root–soil–microbe interactions for crop production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef]
- Von Wiren, N.; Lauter, F.R.; Ninnemann, O.; Gillissen, B.; Walch-Liu, P.; Engels, C. Differential regulation of three functional ammonium transporter genes by nitrogen in root hairs and by light in leaves of tomato. Plant J. 2000, 21, 167–175. [Google Scholar] [CrossRef]
- Ahn, S.J.; Shin, R.; Schachtman, D.P. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake. Plant Physiol. 2004, 134, 1135–1145. [Google Scholar] [CrossRef]
- Desbrosses, G.J.; Stougaard, J. Root nodulation: A paradigm for how plant-microbe symbiosis influences host developmental pathways. Cell Host Microbe 2011, 10, 348–358. [Google Scholar] [CrossRef]
- Traxler, M.F.; Kolter, R. Natural products in soil microbe interactions and evolution. Nat. Prod. Rep. 2015, 32, 956–970. [Google Scholar] [CrossRef]
- Combes-Meynet, E.; Pothier, J.F.; Moënne-Loccoz, Y.; Prigent-Combaret, C. The Pseudomonas secondary metabolite 2,4-diacetylphloroglucinol is a signal inducing rhizoplane expression of Azospirillum genes involved in plant-growth promotion. Mol. Plant-Microbe Interact. 2011, 24, 271–284. [Google Scholar] [CrossRef]
- Hassan, M. The Role of Pectin Utilization in Root Colonization and Plant Growth-Promotion by Bacillus amyloliquefaciens subsp. plantarum (Bap). Master’s Thesis, Auburn University, Auburn, ME, USA, 2016. [Google Scholar]
- Ramaekers, L.; Remans, R.; Rao, I.M.; Blair, M.W.; Vanderleyden, J. Strategies for improving phosphorus acquisition efficiency of crop plants. Field Crops Res. 2010, 117, 169–176. [Google Scholar] [CrossRef]
- Lakshmanan, V.; Castaneda, R.; Rudrappa, T.; Bais, H.P. Root transcriptome analysis of Arabidopsis thaliana exposed to beneficial Bacillus subtilis FB17 rhizobacteria revealed genes for bacterial recruitment and plant defense independent of malate. Planta 2013, 238, 657–668. [Google Scholar] [CrossRef]
- Hiltner, L. About recent experiences and problems the field of soil bacteriology with special consideration of green manure and fallow. Arbeiten der Deutschen Landwirtschaftlichen Gesellschaft 1904, 98, 59–78. [Google Scholar]
- Guo, R.F.; Yuan, G.F.; Wang, Q.M. Effect of NaCl treatments on glucosinolate metabolism in broccoli sprouts. J. Zhejiang Univ. Sci. B 2013, 14, 124. [Google Scholar] [CrossRef]
- He, D.C.; Zhan, J.S.; Xie, L.H. Problems, challenges and future of plant disease management: From an ecological point of view. J. Integr. Agric. 2016, 15, 705–715. [Google Scholar] [CrossRef]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Barka, E.A. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-containing soil bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Dobereiner, J. Review article History and New Perspectives of Diazotrophs in Association with Non-Leguminous Plants. Symbiosis 1992, 13, 1–13. [Google Scholar]
- Podile, A.R.; Kishore, G.K. Plant growth-promoting rhizobacteria. In Plant-Associated Bacteria; Springer: Dordrecht, The Netherlands, 2007; pp. 195–230. [Google Scholar]
- Gray, E.J.; Smith, D.L. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant–bacterium signaling processes. Soil Biol. Biochem. 2005, 37, 395–412. [Google Scholar] [CrossRef]
- Cook, R.J.; Weller, D.M.; El-Banna, A.Y.; Vakoch, D.; Zhang, H. Yield responses of direct-seeded wheat to rhizobacteria and fungicide seed treatments. Plant Dis. 2002, 86, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Amein, T.; Omer, Z.; Welch, C. Application and evaluation of Pseudomonas strains for biocontrol of wheat seedling blight. Crop Prot. 2008, 27, 532–536. [Google Scholar] [CrossRef]
- Ryder, M.H.; Yan, Z.; Terrace, T.E.; Rovira, A.D.; Tang, W.; Correll, R.L. Use of strains of Bacillus isolated in China to suppress take-all and rhizoctonia root rot, and promote seedling growth of glasshouse-grown wheat in Australian soils. Soil Biol. Biochem. 1999, 31, 19–29. [Google Scholar] [CrossRef]
- Cavaglieri, L.; Orlando, J.R.M.I.; Rodríguez, M.I.; Chulze, S.; Etcheverry, M. Biocontrol of Bacillus subtilis against Fusarium verticillioides in vitro and at the maize root level. Res. Microbiol. 2005, 156, 748–754. [Google Scholar] [CrossRef]
- Moustafa, H.E.; Abo-Zaid, G.A.; Abd-Elsalam, H.E.; Hafez, E.E. Antagonistic and inhibitory effect of Bacillus subtilis against certain plant pathogenic fungi, I. Biotechnology 2009, 8, 53–61. [Google Scholar]
- Das, A.J.; Lal, S.; Kumar, R.; Verma, C. Bacterial biosurfactants can be an ecofriendly and advanced technology for remediation of heavy metals and co-contaminated soil. Int. J. Environ. Sci. Technol. 2016, 14, 1343–1354. [Google Scholar] [CrossRef]
- Khan, M.R.; Fischer, S.; Egan, D.; Doohan, F.M. Biological control of Fusarium seedling blight disease of wheat and barley. Phytopathology 2006, 96, 386–394. [Google Scholar] [CrossRef]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.K.; Kumar, A. Disease management of tomato through PGPB: Current trends and future perspective. 3 Biotech 2017, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech 2017, 7, 315. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Verma, H.; Singh, V.K.; Singh, P.P.; Singh, S.K.; Ansari, W.A.; Yadav, A.; Singh, P.K.; Pandey, K.D. Role of Pseudomonas sp. in sustainable agriculture and disease management. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V., Mishra, P., Bisht, J., Pattanayak, A., Eds.; Springer: Singapore, 2017; pp. 195–215. [Google Scholar]
- Kumar, A.; Singh, A.K.; Kaushik, M.S.; Mishra, S.K.; Raj, P.; Singh, P.K.; Pandey, K.D. Interaction of turmeric (Curcuma longa L.) with beneficial microbes: A review. 3 Biotech 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Pandey, D.K.; Kumar, A.; Singh, M. PGPR isolates from the rhizosphere of vegetable crop Momordica charantia: Characterization and application as biofertilizer. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1789–1802. [Google Scholar]
- Haas, D.; Keel, C. Regulation of antibiotic production in root-colonizing Pseudomonas spp. and relevance for biological control of plant disease. Annu. Rev. Phytopathol. 2003, 41, 117–153. [Google Scholar] [CrossRef] [PubMed]
- Dowling, D.N.; O’Gara, F. Metabolites of Pseudomonas involved in the biocontrol of plant disease. Trends Biotechnol. 1994, 12, 133–141. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Duffy, B. Pathogen self-defense: Mechanisms to counteract microbial antagonism. Annu. Rev. Phytopathol. 2003, 41, 501–538. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- de Bruijn, I.; de Kock, M.J.D.; Yang, M.; de Waard, P.; van Beek, T.A.; Raaijmakers, J.M. Genome-based discovery, structure prediction and functional analysis of cyclic lipopeptide antibiotics in Pseudomonas species. Mol. Microbiol. 2007, 63, 417–428. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; de Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef]
- Abbas, T.; Zahir, Z.A.; Naveed, M.; Kremer, R.J. Limitations of existing weed control practices necessitate development of alternative techniques based on biological approaches. Adv. Agron. 2018, 147, 239–280. [Google Scholar]
- Maksimov, I.V.; Abizgil’dina, R.R.; Pusenkova, L.I. Plant growth promoting rhizobacteria as alternative to chemical crop protectors from pathogens (Review). Appl. Biochem. Microbiol. 2011, 47, 333–345. [Google Scholar] [CrossRef]
- Hill, D.S.; Stein, J.I.; Torkewitz, N.R.; Morse, A.M.; Howell, C.R.; Pachlatko, J.P.; Becker, J.O.; Ligon, J.M. Cloning of genes involved in the synthesis of pyrrolnitrin from Pseudomonas fluorescens and role of pyrrolnitrin synthesis in biological control of plant disease. Appl. Environ. Microbiol. 1994, 60, 78–85. [Google Scholar] [CrossRef]
- Silo-Suh, L.A.; Lethbridge, B.J.; Raffel, S.J.; He, H.; Clardy, J.; Handelsman, J. Biological activities of two fungistatic antibiotics produced by Bacillus cereus UW85. Appl. Environ. Microbiol. 1994, 60, 2023–2030. [Google Scholar] [CrossRef]
- He, H.; Silo-Suh, L.A.; Handelsman, J.; Clardy, J. Zwittermicin A, an antifungal and plant protection agent from Bacillus cereus. Tetrahedron Lett. 1994, 35, 2499–2502. [Google Scholar] [CrossRef]
- Emmert, E.A.; Handelsman, J. Biocontrol of plant disease: A (Gram-) positive perspective. FEMS Microbiol. Lett. 1999, 171, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kokalis-Burelle, N.; Kloepper, J.W.; Reddy, M.S. Plant growth-promoting rhizobacteria as transplant amendments and their effects on indigenous rhizosphere microorganisms. Appl. Soil Ecol. 2005, 31, 91–100. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Bakker, P.A.H.M.; Pieterse, C.M.J. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [PubMed]
- Van Peer, R.; Niemann, G.J.; Schippers, B. Induced resistance and phytoalexin accumulation in biological control of Fusarium wilt of carnation by Pseudomonas sp. strain WCS 417 r. Phytopathology 1991, 81, 728–734. [Google Scholar] [CrossRef]
- Wei, G.; Kloepper, J.W.; Tuzun, S. Induction of systemic resistance of cucumber to Colletotrichum orbiculare by select strains of plant growth-promoting rhizobacteria. Phytopathology 1991, 81, 1508–1512. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef]
- Van der Ent, S.; Verhagen, B.W.; Van Doorn, R.; Bakker, D.; Verlaan, M.G.; Pel, M.J.; Joosten, R.G.; Proveniers, M.C.; Van Loon, L.C.; Ton, J.; et al. MYB72 is required in early signaling steps of rhizobacteria-induced systemic resistance in Arabidopsis. Plant Physiol. 2008, 146, 1293–1304. [Google Scholar] [CrossRef]
- Somers, E.; Vanderleyden, J.; Srinivasan, M. Rhizosphere bacterial signalling: A love parade beneath our feet. Crit. Rev. Microbiol. 2004, 30, 205–240. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.M.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Vleesschauwer, D.; Höfte, M. Rhizobacteria-induced systemic resistance. Adv. Bot. Res. 2009, 51, 223–281. [Google Scholar]
- Van Loon, L.C. Plant responses to plant growth-promoting rhizobacteria. In New Perspectives and Approaches in Plant Growth-Promoting Rhizobacteria Research; Springer: Dordrecht, The Netherlands, 2007; pp. 243–254. [Google Scholar]
- Van Peer, R.; Schippers, B. Lipopolysaccharides of plant-growth promoting Pseudomonas sp. strain WCS417r induce resistance in carnation to Fusarium wilt. Neth. J. Plant Pathol. 1992, 98, 129–139. [Google Scholar] [CrossRef]
- Leeman, M.; Van Pelt, J.A.; Den Ouden, F.M.; Heinsbroek, M.; Bakker, P.A.H.M.; Schippers, B. Induction of systemic resistance by Pseudomonas fluorescens in radish cultivars differing in susceptibility to fusarium wilt, using a novel bioassay. Eur. J. Plant Pathol. 1995, 101, 655–664. [Google Scholar] [CrossRef]
- Silva, H.S.A.; Romeiro, R.D.S.; Mounteer, A. Development of a root colonization bioassay for rapid screening of rhizobacteria for potential biocontrol agents. J. Phytopathol. 2003, 151, 42–46. [Google Scholar] [CrossRef]
- Handelsman, J.; Stabb, E.V. Biocontrol of soilborne plant pathogens. Plant Cell 1996, 8, 1855. [Google Scholar] [CrossRef] [PubMed]
- Benizri, E.; Baudoin, E.; Guckert, A. Root colonization by inoculated plant growth-promoting rhizobacteria. Biocontrol Sci. Technol. 2001, 11, 557–574. [Google Scholar] [CrossRef]
- Montealegre, J.R.; Reyes, R.; Pérez, L.M.; Herrera, R.; Silva, P.; Besoain, X. Selection of bioantagonistic bacteria to be used in biological control of Rhizoctonia solani in tomato. Electron. J. Biotechnol. 2003, 6, 115–127. [Google Scholar] [CrossRef]
- Rangarajan, S.; Saleena, L.M.; Vasudevan, P.; Nair, S. Biological suppression of rice diseases by Pseudomonas spp. under saline soil conditions. Plant Soil 2003, 251, 73–82. [Google Scholar] [CrossRef]
- Bloemberg, G.V.; Lugtenberg, B.J. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Beattie, G.A. Plant-associated bacteria: Survey, molecular phylogeny, genomics and recent advances. In Plant-Associated Bacteria; Springer: Dordrecht, The Netherlands, 2007; pp. 1–56. [Google Scholar]
- Neeraja, C.; Anil, K.; Purushotham, P.; Suma, K.; Sarma, P.V.S.R.N.; Moerschbacher, B.M.; Podile, A.R. Biotechnological approaches to develop bacterial chitinases as a bioshield against fungal diseases of plants. Crit. Rev. Biotechnol. 2010, 30, 231–241. [Google Scholar] [CrossRef]
- Stephens, P.M.; Crowley, J.J.; O’Connell, C. Selection of pseudomonad strains inhibiting Pythium ultimum on sugarbeet seeds in soil. Soil Biol. Biochem. 1993, 25, 1283–1288. [Google Scholar] [CrossRef]
- Kamilova, F.; Validov, S.; Azarova, T.; Mulders, I.; Lugtenberg, B. Enrichment for enhanced competitive plant root tip colonizers selects for a new class of biocontrol bacteria. Environ. Microbiol. 2005, 7, 1809–1817. [Google Scholar] [CrossRef]
- Glick, B.R.; Bashan, Y. Genetic manipulation of plant growth-promoting bacteria to enhance biocontrol of phytopathogens. Biotechnol. Adv. 1997, 15, 353–378. [Google Scholar] [CrossRef]
- Dakora, F.D. Defining new roles for plant and rhizobial molecules in sole and mixed plant cultures involving symbiotic legumes. New Phytol. 2003, 158, 39–49. [Google Scholar] [CrossRef]
- Campanoni, P.; Blasius, B.; Nick, P. Auxin transport synchronizes the pattern of cell division in a tobacco cell line. Plant Physiol. 2003, 133, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Chabot, R.; Antoun, H.; Cescas, M.P. Growth promotion of maize and lettuce by phosphate-solubilizing Rhizobium leguminosarum biovar. phaseoli. Plant Soil 1996, 184, 311–321. [Google Scholar] [CrossRef]
- Yanni, Y.G.; Rizk, R.Y.; Corich, V.; Squartini, A.; Ninke, K.; Philip-Hollingsworth, S.; Orgambide, G.; De Bruijn, F.; Stoltzfus, J.; Buckley, D.; et al. Natural endophytic association between Rhizobium leguminosarum bv. trifolii and rice roots and assessment of its potential to promote rice growth. In Opportunities for Biological Nitrogen Fixation in Rice and Other Non-Legumes; Springer: Dordrecht, The Netherlands, 1997; pp. 99–114. [Google Scholar]
- Validov, S. Biocontrol of Tomato Foot and Root Rot by Pseudomonas bacteria in Stonewool. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 2007. [Google Scholar]
- Thomashow, L.S.; Weller, D.M. Current concepts in the use of introduced bacteria for biological disease control: Mechanisms and antifungal metabolites. In Plant-Microbe Interactions; Springer: Boston, MA, USA, 1996; pp. 187–235. [Google Scholar]
- Dekkers, L.C.; Mulders, I.H.; Phoelich, C.C.; Chin-A-Woeng, T.F.; Wijfjes, A.H.; Lugtenberg, B.J. The sss colonization gene of the tomato-Fusarium oxysporum f. sp. radicis-lycopersici biocontrol strain Pseudomonas fluorescens WCS365 can improve root colonization of other wild-type Pseudomonas spp. bacteria. Mol. Plant Microbe Interact. 2000, 13, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.J.; Dekkers, L.; Bloemberg, G.V. Molecular determinants of rhizosphere colonization by Pseudomonas. Annu. Rev. Phytopathol. 2001, 39, 461–490. [Google Scholar] [CrossRef] [PubMed]
- Kremer, R.J.; Souissi, T. Cyanide production by rhizobacteria and potential for suppression of weed seedling growth. Curr. Microbiol. 2001, 43, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Chet, I.; Ordentlich, A.; Shapira, R.; Oppenheim, A. Mechanisms of biocontrol of soil-borne plant pathogens by rhizobacteria. Plant Soil 1990, 129, 85–92. [Google Scholar] [CrossRef]
- Kobayashi, D.Y.; Reedy, R.M.; Bick, J.; Oudemans, P.V. Characterization of a chitinase gene from Stenotrophomonas maltophilia strain 34S1 and its involvement in biological control. Appl. Environ. Microbiol. 2002, 68, 1047–1054. [Google Scholar] [CrossRef]
- Sadfi, N.; Cherif, M.; Fliss, I.; Boudabbous, A.; Antoun, H. Evaluation of bacterial isolates from salty soils and Bacillus thuringiensis strains for the biocontrol of Fusarium dry rot of potato tubers. J. Plant Pathol. 2001, 83, 101–117. [Google Scholar]
- Neiendam Nielsen, M.; Sørensen, J. Chitinolytic activity of Pseudomonas fluorescens isolates from barley and sugar beet rhizosphere. FEMS Microbiol. Ecol. 1999, 30, 217–227. [Google Scholar] [CrossRef]
- Budi, S.W.; van Tuinen, D.; Arnould, C.; Dumas-Gaudot, E.; Gianinazzi-Pearson, V.; Gianinazzi, S. Hydrolytic enzyme activity of Paenibacillus sp. strain B2 and effects of the antagonistic bacterium on cell integrity of two soil-borne pathogenic fungi. Appl. Soil Ecol. 2000, 15, 191–199. [Google Scholar] [CrossRef]
- Someya, N.; Kataoka, N.; Komagata, T.; Hirayae, K.; Hibi, T.; Akutsu, K. Biological control of cyclamen soilborne diseases by Serratia marcescens strain B2. Plant Dis. 2000, 84, 334–340. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kurek, E.; Jaroszuk-Ściseł, J. Rye (Secale cereale) growth promotion by Pseudomonas fluorescens strains and their interactions with Fusarium culmorum under various soil conditions. Biol. Control 2003, 26, 48–56. [Google Scholar] [CrossRef]
- Mulla, S.I.; Bharagava, R.N.; Belhaj, D.; Saratale, G.D.; Kumar, A.; Rajasekar, A.; Tallur, P.N.; Edalli, V.A.; Hu, A.; Yu, C.P. Microbial degradation of phenolic compounds. In Microbes and Enzymes in Soil Health and Bioremediation; Microorganisms for sustainability; Kumar, A., Sharma, S., Eds.; Springer: Singapore, 2019; Volume 16. [Google Scholar] [CrossRef]
- Phulpoto, A.H.; Maitlo, M.A.; Kanhar, N.A. Culture dependent to culture independent approaches for the bioremediation of paints: A review. Int. J. Environ. Sci. Technol. 2021, 18, 241–262. [Google Scholar] [CrossRef]
- Muratova, A.Y.; Turkovskaya, O.V.; Antonyuk, L.P.; Makarov, O.E.; Pozdnyakova, L.I.; Ignatov, V.V. Oil-oxidizing potential of associative rhizobacteria of the genus Azospirillum. Microbiology 2005, 74, 210–215. [Google Scholar] [CrossRef]
- Ali, M.H.; Sattar, M.T.; Khan, M.I.; Naveed, M.; Rafique, M.; Alamri, S.; Siddiqui, M.H. Enhanced Growth of Mungbean and Remediation of Petroleum Hydrocarbons by Enterobacter sp. MN17 and Biochar Addition in Diesel Contaminated Soil. Appl. Sci. 2020, 10, 8548. [Google Scholar]
- Ejaz, M.; Zhao, B.; Wang, X.; Bashir, S.; Haider, F.U.; Aslam, Z.; Khan, M.I.; Shabaan, M.; Naveed, M.; Mustafa, A. Isolation and characterization of oil degrading Enterobacter sp. from naturally hydrocarbon contaminated soils and their potential against bioremediation of crude oil. Appl. Sci. 2021, 11, 3504. [Google Scholar] [CrossRef]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Alam Cheema, S.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef]
- Varjani, S.; Kumar, G.; Rene, E.R. Developments in biochar application for pesticide remediation: Current knowledge and future research directions. J. Environ. Manag. 2019, 232, 505–513. [Google Scholar] [CrossRef]
- Mahjoubi, M.; Cappello, S.; Souissi, Y.; Jaouani, A.; Cherif, A. Microbial bioremediation of petroleum hydrocarbon–contaminated marine environments. In Recent Insights in Petroleum Science and Engineering; Zoveidavianpoor, M., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Lu, L.; Zhang, J.; Peng, C. Shift of Soil Polycyclic Aromatic Hydrocarbons (PAHs) dissipation pattern and microbial community composition due to rhamnolipid supplementation. Water Air Soil Pollut. 2019, 230, 107. [Google Scholar] [CrossRef]
- Kumar, V.; Chandra, R. Bacterial-assisted phytoextraction mechanism of heavy metals by native hyperaccumulator plants from distillery waste–contaminated site for eco-restoration. In Microbes for Sustainable Development and Bioremediation; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar] [CrossRef]
- Macías-Almazán, A.; Lois-Correa, J.A.; Domínguez-Crespo, M.A.; López-Oyama, A.B.; Torres-Huerta, A.M.; Brachetti-Sibaja, S.B.; Rodríguez-Salazar, A.E. Influence of operating conditions on proton conductivity of nanocellulose films using two agroindustrial wastes: Sugarcane bagasse and pinewood sawdust. Carbohydr. Polym. 2020, 238, 116171. [Google Scholar] [CrossRef]
- Russo, F.; Ceci, A.; Pinzari, F.; Siciliano, A.; Guida, M.; Malusà, E.; Tartanus, M.; Miszczak, A.; Maggi, O.; Persiani, A.M. Bioremediation of dichlorodiphenyltrichloroethane (DDT)-contaminated agricultural soils: Potential of two autochthonous saprotrophic fungal strains. Appl. Environ. Microbiol. 2019, 85, 01720-19. [Google Scholar] [CrossRef]
- Chandra, R.; Kumar, V. Phytoextraction of heavy metals by potential native plants and their microscopic observation of root growing on stabilised distillery sludge as a prospective tool for in situ phytoremediation of industrial waste. Environ. Sci. Pollut. Res. 2017, 24, 2605–2619. [Google Scholar] [CrossRef]
- Chouychai, W.; Thongkukiatkul, A.; Upatham, S.; Pokethitiyook, P.; Kruatrachue, M.; Lee, H. Effect of corn plant on survival and phenanthrene degradation capacity of Pseudomonas sp. UG14Lr in two soils. Int. J. Phytoremediat. 2012, 14, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Malik, Z.A.; Ahmed, S. Degradation of petroleum hydrocarbons by oil field isolated bacterial consortium. Afr. J. Biotechnol. 2012, 11, 650–658. [Google Scholar]
- Thomas, F.; Cébron, A. Short-term rhizosphere effect on available carbon sources, phenanthrene degradation, and active microbiome in an aged contaminated industrial soil. Front. Microbiol. 2016, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Akhtar, M.J.; Zahir, Z.A.; Naveed, M.; Mitter, B.; Sessitsch, A. Cadmium-tolerant bacteria induce metal stress tolerance in cereals. Environ. Sci. Pollut. Res. 2014, 21, 11054–11065. [Google Scholar] [CrossRef] [PubMed]
- Lata, S.; Kaur, H.P.; Mishra, T. Cadmium bioremediation: A review. Int. J. Pharm. Sci. Res. 2019, 10, 4120–4128. [Google Scholar] [CrossRef]
- Yaghoubian, Y.; Siadat, S.A.; Telavat, M.R.M.; Pirdashti, H.; Yaghoubia, I. Bio-removal of cadmium from aqueous solutions by filamentous fungi: Trichoderma spp. and Piriformospora indica. Environ. Sci. Pollut. Res. 2019, 26, 7863–7872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, M.; Li, Y.; Che, Y.; Xiao, Y. Effects of arbuscularmycorrhizal fungi, biochar and cadmium on the yield and element uptake of Medicago sativa. Sci. Total Environ. 2019, 655, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Panchenko, L.; Muratova, A.; Dubrovskaya, E.; Golubev, S.; Turkovskaya, O. Dynamics of natural revegetation of hydrocarbon-contaminated soil and remediation potential of indigenous plant species in the steppe zone of the southern Volga Uplands. Environ. Sci. Pollut. Res. 2018, 25, 3260–3274. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, G.; Naushad, M.; Ala’a, H.; Kumar, A.; Hira, I.; Ahamad, T.; Ghfar, A.A.; Stadler, F.J. Visible photodegradation of ibuprofen and 2, 4-D in simulated waste water using sustainable metal free-hybrids based on carbon nitride and biochar. J. Environ. Manag. 2019, 231, 1164–1175. [Google Scholar] [CrossRef]
- Valderrama, J.A.; Durante-Rodríguez, G.; Blázquez, B.; García, J.L.; Carmona, M.; Díaz, E. Bacterial Degradation of Benzoate cross-regulation between aerobic and anaerobic pathways. J. Biol. Chem. 2012, 287, 10494–10508. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.L.; Maguire, K.L.; Anderson, D.R.; McGrath, S.P. Enhanced dissipation of chrysene in planted soil: The impact of a rhizobial inoculum. Soil Biol. Biochem. 2004, 36, 33–38. [Google Scholar] [CrossRef]
- Al-Wasify, R.S.; Hamed, S.R. Bacterial biodegradation of crude oil using local isolates. Int. J. Bacteriol. 2014, 2014, 863272. [Google Scholar] [CrossRef] [PubMed]
- Ichor, T.; Okerentugba, P.O.; Okpokwasili, G.C. Biodegradation of total petroleum hydrocarbon by aerobic heterotrophic bacteria isolated from crude oil contaminated brackish waters of Bodo Creek. J. Bioremed. Biodeg. 2014, 5, 1000236. [Google Scholar] [CrossRef]
- Jilani, S. Bioremediation application for textile effluent treatment. Middle East J. Sci. Res. 2015, 23, 26–34. [Google Scholar] [CrossRef]
- Verma, J.P.; Jaiswal, D.K.; Sagar, R. Pesticide relevance and their microbial degradation: A-state-of-art. Rev. Environ. Sci. Biotechnol. 2014, 13, 429–466. [Google Scholar] [CrossRef]
- Koksoy, H.; Aslim, B. Determination of herbicide resistance in aquatic cyanobacteria by probit analysis. J. Appl. Biol. Sci. 2013, 7, 37–41. [Google Scholar]
- Begum, S.S.; Arundhati, A. A study of bioremediation of methyl parathion in vitro India. Int. J. Curr. Microbiol. App. Sci. 2016, 5, 464–474. [Google Scholar] [CrossRef]
- Baraldi, E.A.; Damianovic, M.; Manfio, G.R.; Foresti, E.; Vazoller, R.F. Performance of a horizontal-flow anaerobic immobilized biomass (HAIB) reactor and dynamics of the microbial community during degradation of pentachlorophenol (PCP). Anaerobe 2008, 14, 268–274. [Google Scholar] [CrossRef]
- Tripathi, A.; Dixit. S. Bioremediation of phenolic compounds by higher fungi—A review. Int. J. Curr. Adv. Res. 2016, 4, 14–35. [Google Scholar] [CrossRef]
- Thakurta, G.S.; Aakula, M.; Chakrabarty, J.; Dutta, S. Bioremediation of phenol from synthetic and real wastewater using Leptolyngbya sp.: A comparison and assessment of lipid production. 3 Biotech 2018, 8, 206. [Google Scholar] [CrossRef]
- Sheng, X.F.; He, L.Y. Solubilization of potassium-bearing minerals by a wild-type strain of Bacillus edaphicus and its mutants and increased potassium uptake by wheat. Can. J. Microbiol. 2006, 52, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Gao, Y.; Wu, S.C.; Cheung, K.C.; Wang, X.R.; Wong, M.H. Physiological and biochemical responses of rice (Oryza sativa L.) to phenanthrene and pyrene. Int. J. Phytorem. 2008, 10, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Chouychai, W.; Thongkukiatkul, A.; Upatham, S.; Lee, H.; Pokethitiyook, P.; Kruatrachue, M. Plant-enhanced phenanthrene and pyrene biodegradation in acidic soil. J. Environ. Biol. 2009, 30, 139–144. [Google Scholar] [PubMed]
- Lechner, U.; Turkowsky, D.; Dinh, T.T.H.; Al-Fathi, H.; Schwoch, S.; Franke, S.; Gerlach, M.S.; Koch, M.; Bergen, M.V.; Jehmlich, N.; et al. Desulfitobacterium contributes to the microbial transformation of 2,4,5-T by methanogenic enrichment cultures from a Vietnamese active landfill. Microb. Biotechnol. 2018, 11, 1137–1156. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.N.; Yong, R.N.; Gibbs, B.F. Surfactant enhanced remediation of contaminated soil: A review. Eng. Geol. 2001, 60, 371–380. [Google Scholar] [CrossRef]
- Kaczorek, E.; Chrzanowski, Ł.; Pijanowska, A.; Olszanowski, A. Yeast and bacteria cell hydrophobicity and hydrocarbon biodegradation in the presence of natural surfactants: Rhamnolipides and saponins. Bioresour. Technol. 2008, 99, 4285–4291. [Google Scholar] [CrossRef]
- Mohanty, S.; Mukherji, S. Surfactant-aided biodegradation of NAPLs by Burkholderia multivorans: Comparison between Triton X-100 and rhamnolipid JBR 515. Colloids Surf. B 2012, 102, 644–652. [Google Scholar] [CrossRef]
- Selberg, A.; Juuram, K.; Budashova, J.; Tenno, T. Biodegradation and leaching of surfactants during surfactant-amended bioremediation of oil-polluted soil. In Applied Bioremediation—Active and Passive Approaches; Patil, Y.B., Rao, P., Eds.; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef]
- Babu, A.G.; Shim, J.; Shea, P.J.; Oh, B.T. Penicillium aculeatum PDR-4 and Trichoderma sp. PDR-16 promote phytoremediation of mine tailing soil and bioenergy production with sorghum-sudan grass. Ecol. Eng. 2014, 69, 186–191. [Google Scholar] [CrossRef]
- Mahajan, P.; Kaushal, J. Role of phytoremediation in reducing cadmium toxicity in soil and water. Hindawi J. Toxicol. 2018, 2018, 4864365. [Google Scholar] [CrossRef]
- Belimov, A.A.; Hontzeas, N.; Safronova, V.I.; Demchinskaya, S.V.; Piluzza, G.; Bullitta, S.; Glick, B.R. Cadmium-tolerant plant growth promoting rhizobacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol. Biochem. 2005, 37, 241–250. [Google Scholar] [CrossRef]
- Robinson, B.; Russell, C.; Hedley, M.J.; Clothier, B. Cadmium adsorption by rhizobacteria: Implications for New Zealand Pastureland. Agric. Ecosyst. Environ. 2001, 87, 315–321. [Google Scholar] [CrossRef]
- Gupta, A.; Meyer, J.M.; Goel, R. Development of heavy metal resistant mutants of phosphate solubilizing Pseudomonas sp. NBRI4014 and their characterization. Curr. Microbiol. 2002, 45, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Marques, G.C.; Moreira, H.; Franco, A.R.; Rangel, A.O.S.S.; Castro, P.M.L. Inoculating Helianthus annuus (sunflower) grown in zinc and cadmium contaminated soils with plant growth promoting bacteria—Effects on phytoremediation strategies. Chemosphere 2013, 92, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Prapagdee, B.; Khonsue, N. Bacterial-assisted cadmium phytoremediation by Ocimum gratissimum L. in polluted agricultural soil: A field trial experiment. Int. J. Environ. Sci. Technol. 2015, 12, 3843–3852. [Google Scholar] [CrossRef]
- Yuan, Z.; Yi, H.; Wang, T.; Zhang, Y.; Zhu, X.; Yao, J. Application of phosphate solubilizing bacteria in immobilization of Pb and Cd in soil. Environ. Sci. Pollut. Res. 2017, 27, 21877–21884. [Google Scholar] [CrossRef]
- Martín-González, A.; Díaz, S.; Borniquel, S.; Gallego, A.; Gutiérrez, J.C. Cytotoxicity and bioaccumulation of heavy metals by ciliated protozoa isolated from urban wastewater treatment plants. Res. Microbiol. 2006, 157, 108–118. [Google Scholar] [CrossRef]
- Saranya, K.; Sundaramanickam, A.; Shekhar, S.; Meena, M.; Sathishkumar, R.S.; Balasubramanian, T. Biosorption of multiheavy metals by coral associated phosphate solubilising bacteria Cronobacter muytjensii KSCAS2. J. Environ. Manag. 2018, 222, 396–401. [Google Scholar] [CrossRef]
- Choińska-Pulit, A.; Sobolczyk-Bednarek, J.; Łaba, W. Optimization of copper, lead and cadmium biosorption onto newly isolated bacterium using a Box-Behnken design. Ecotoxicol. Environ Saf. 2018, 149, 275–283. [Google Scholar] [CrossRef]
- Vishan, I.; Saha, B.; Sivaprakasam, S.; Kalamdhad, A. Evaluation of Cd(II) biosorption in aqueous solution by using lyophilized biomass of novel bacterial strain Bacillus badius AK: Biosorption kinetics, thermodynamics and mechanism. Environ. Technol. Innov. 2019, 14, 100323. [Google Scholar] [CrossRef]
- Wu, S.C.; Cheung, K.C.; Luo, Y.M.; Wong, M.H. Effects of inoculation of plant growth-promoting rhizobacteria on metal uptake by Brassica juncea. Environ. Pollut. 2006, 140, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Arunakumara, K.K.; Walpola, B.C.; Song, J.S.; Shin, M.; Lee, C.; Yoon, M. Phytoextraction of heavy metals induced by bioaugmentation of a phosphate solubilising bacterium. Korean J. Environ. Agric. 2014, 33, 220–230. [Google Scholar] [CrossRef]
- Faisal, M.; Hasnain, S. Bacterial Cr (VI) reduction concurrently improves sunflower (Helianthus annuus L.) growth. Biotechnol. Lett. 2005, 27, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shanab, R.A.; Ghanem, K.; Ghanem, N.; Al-Kolaibe, A. The role of bacteria on heavy metal extraction and uptake by plants growing on multi-metal-contaminated soils. World J. Microbiol. Biotechnol. 2008, 24, 253–262. [Google Scholar] [CrossRef]
- Oves, M.; Khan, M.S.; Zaidi, A. Chromium reducing and plant growth promoting novel strain Pseudomonas aeruginosa OSG41 enhance chickpea growth in chromium amended soils. Eur. J. Soil Biol. 2013, 56, 72–83. [Google Scholar] [CrossRef]
- Naseem, H.; Bano, A. Role of plant growth-promoting rhizobacteria and their exopolysaccharide in drought tolerance of maize. J. Plant Interact. 2014, 9, 689–701. [Google Scholar] [CrossRef]
- Khan, M.U.; Sessitsch, A.; Harris, M.; Fatima, K.; Imran, A.; Arslan, M.; Shabir, G.; Khan, Q.M.; Afzal, M. Cr-resistant rhizo-and endophytic bacteria associated with Prosopis juliflora and their potential as phytoremediation enhancing agents in metal-degraded soils. Front. Plant Sci. 2015, 5, 755. [Google Scholar] [CrossRef]
- Srinath, T.; Verma, T.; Ramteke, P.; Garg, S. Chromium (VI) biosorption and bioaccumulation by chromate resistant bacteria. Chemosphere 2002, 48, 427–435. [Google Scholar] [CrossRef]
- Ksheminska, H.; Fedorovych, D.; Babyak, L.; Yanovych, D.; Kaszycki, P.; Koloczek, H. Chromium (III) and (VI) tolerance and bioaccumulation in yeast: A survey of cellular chromium content in selected strains of representative genera. Process Biochem. 2005, 40, 1565–1572. [Google Scholar] [CrossRef]
- Li, K.; Ramakrishna, W. Effect of multiple metal resistant bacteria from contaminated lake sediments on metal accumulation and plant growth. J. Hazard. Mater. 2011, 189, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.D.; He, Z.L.; Yang, X.E. Role of soil rhizobacteria in phytoremediation of heavy metal contaminated soils. J. Zhejiang Univ. Sci. B 2007, 8, 192–207. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, W.W. Rhizosphere processes and management in plant-assisted bioremediation (phytoremediation) of soils. Plant Soil 2009, 321, 385–408. [Google Scholar] [CrossRef]
- Płociniczak, T.; Sinkkonen, A.; Romantschuk, M.; Sułowicz, S.; Piotrowska-Seget, Z. Rhizospheric bacterial strain Brevibacterium casei MH8a colonizes plant tissues and enhances Cd, Zn, Cu phytoextraction by white mustard. Front. Plant Sci. 2016, 16, 101. [Google Scholar] [CrossRef]
- Lin, M.; Jin, M.; Xu, K.; He, L.; Cheng, D. Phosphate solubilising bacteria improve the phytoremediation efficiency of Wedeliatrilobata for Cu-contaminated soil. Int. J. Phytoremediat. 2018, 20, 813–822. [Google Scholar] [CrossRef]
- Iram, S.; Shabbir, R.; Zafar, H.; Javaid, M. Biosorption and bioaccumulation of copper and lead by heavy metal-resistant fungal isolates. Arab. J. Sci. Eng. 2015, 40, 1867–1873. [Google Scholar] [CrossRef]
- De-Silóniz, M.-I.; Balsalobre, L.; Alba, C.; Valderrama, M.-J.; Peinado, J.M. Feasibility of copper uptake by the yeast Pichia guilliermondii isolated from sewage sludge. Res. Microbiol. 2002, 153, 173–180. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Rocha, I.; Oliveira, R.; Freitas, H. Serpentine bacteria influence metal translocation and bioconcentration of Brassica juncea and Ricinus communis growing in multi-pollute soils. Front. Plant Sci. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis strain SJ 101 as a bioinoculant for concurrent plant growth promotion and nickel accumulation in Brassica juncea. Chemosphere 2006, 64, 991–997. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Prieto-Fernandez, A.; Alvarez-Lopez, V.; Monterroso, C.; Cabello-Conejo, M.I.; Acea, M.J. Nickel solubilising capacity of rhizobacteria isolated from hyperaccumulating and non-hyperaccumulating subspecies of Alyssum serpyllifolium. Int. J. Phytoremediat. 2011, 13, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Aboudrar, W.; Schwartz, C.; Morel, J.L.; Boularbah, A. Effect of nickel-resistant rhizosphere bacteria on the uptake of nickel by the hyperaccumulator Noccaea caerulescens under controlled conditions. J. Soils Sediments 2013, 13, 501–507. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Aamer, M.; Nawaz, M.; Ali, A.; Khan, M.A.U.; Khan, T.A. Nickel toxicity in plants: Reasons, toxic effects, tolerance mechanisms, and remediation possibilities—A review. Environ. Sci. Pollut. Res. 2019, 26, 12673–12688. [Google Scholar] [CrossRef]
- Burd, G.I. Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol. 2000, 46, 237–245. [Google Scholar] [CrossRef]
- Govarthanan, M.; Mythili, R.; Selvakumar, T.; Kamala-Kannan, S.; Kim, H. Myco-phytoremediation of arsenic- and lead-contaminated soils by Helianthus annuus and wood rot fungi, Trichoderma sp. isolated from decayed wood. Ecotoxicol. Environ. Saf. 2018, 151, 279–284. [Google Scholar] [CrossRef]
- Mohapatra, R.K.; Parhi, P.K.; Pandey, S.; Bindhani, B.K.; Thatoi, H.; Panda, C.R. Active and passive biosorption of Pb(II)using live and dead biomass of marine bacterium Bacillus xiamenensis PbRPSD202: Kinetics and isotherm studies. J. Environ. Manag. 2019, 247, 121–134. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Z.; Ma, Y.; Zhou, Y.; Zhao, S.; Wang, L.; Zhai, H. Influence of elevated Zn (II) on Anammox system: Microbial variation and zinc tolerance. Bioresour. Technol. 2018, 251, 108–113. [Google Scholar] [CrossRef]
- Kramer, J.; Özkaya, Ö.; Kümmerli, R. Bacterial siderophores in community and host interactions. Nat. Rev. Microbiol. 2020, 18, 152–163. [Google Scholar] [CrossRef]
- Prabhu, N.; Borkar, S.; Garg, S. Phosphate solubilization by microorganisms. In Advances in Biological Science Research; Academic Press: Cambridge, MA, USA, 2019; pp. 161–176. [Google Scholar] [CrossRef]
- Rajkumar, M.; Ae, N.; Prasad, M.N.; Freitas, H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef]
- Gaonkar, T.; Bhosle, S. Effect of metals on a siderophore producing isolate and its implications on microbial assisted bioremediation of metal contaminated soils. Chemosphere 2013, 93, 9. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Saxena, G.; Kumar, V. Phytoremediation of environmental pollutants: An eco-sustainable green technology to environmental management. In Advances in Biodegradation and Bioremediation of Industrial Waste; Chandra, R., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 1–29. [Google Scholar]
- Ma, Y.; Prasad, M.N.V.; Rajkumar, M.; Freitas, H. Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol. Adv. 2011, 29, 248–258. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Mechanisms used by plant growth-promoting bacteria. In Bacteria in Agrobiology: Plant Nutrient Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 17–46. [Google Scholar]
- Violante, A.; Zhu, J.; Pigna, M.; Jara, A.; Cozzolino, V.; Mora, M.L. Role of Biomolecules in Influencing Transformation Mechanisms of Metals and Metalloids in Soil Environments. In Molecular Environmental Soil Science; Springer: Dordrecht, The Netherlands, 2013; pp. 167–191. [Google Scholar]
- Meyer, J.M. Pyoverdines: Pigments siderophores and potential taxonomic markers of fluorescent Pseudomonas species. Arch. Microbiol. 2000, 174, 135–142. [Google Scholar] [CrossRef]
- Jing, Y.X.; Yan, J.L.; He, H.D.; Yang, D.J.; Xiao, L. Characterization of bacteria in the rhizosphere soils of Polygonum pubescens and their potential in promoting growth and Cd, Pb, Zn uptake by Brassica napus. Int. J. Phytoremediat. 2014, 16, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Sifour, M.; Al-Jilawi, M.H.; Aziz, G.M. Emulsification properties of biosurfactant produced from Pseudomonas aeruginosa RB 28. Pak. J. Biol. Sci. 2007, 10, 1331–1335. [Google Scholar] [CrossRef]
- Reddy, M.S.; Naresh, B.; Leela, T.; Prashanthi, M.; Madhusudhan, N.C.; Dhanasri, G. Biodegradation of phenanthrene with biosurfactant production by a new strain of Brevibacillus sp. Bioresour. Technol. 2010, 101, 79807983. [Google Scholar] [CrossRef] [PubMed]
- Geys, R.; Soetaert, W.; Bogaer, I.V. Biotechnological opportunities in biosurfactant production. Curr. Opin. Biotechnol. 2014, 30, 66–72. [Google Scholar] [CrossRef]
- Ławniczak, Ł.; Marecik, R.; Chrzanowski, Ł. Contributions of biosurfactants to natural or induced bioremediation. Appl. Microbiol. Biotechnol. 2013, 97, 2327–2339. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.N.; Aglave, B.A.; Pethkar, A.V.; Gaikwad, V.B. Stenotrophomonas koreensis a novel biosurfactant producer for abatement of heavy metals from the environment. Afr. J. Microbiol. Res. 2012, 6, 51735178. [Google Scholar]
- Lima, T.M.S.; Procopio, L.C.; Brandao, F.D.; Carvalho, A.M.X.; Totola, M.R.; Borges, A.C. Simultaneous phenanthrene and cadmium removal from contaminated soil by a ligand/biosurfactant solution. Biodegradation 2011, 22, 10071015. [Google Scholar] [CrossRef]
- Qazi, M.A.; Malik, Z.A.; Qureshi, G.D.; Hameed, A.; Ahmed, S. Yeast extract as the most preferable substrate for optimized biosurfactant production by rhlB gene positive Pseudomonas putida SOL-10 isolate. J. Bioremed. Biodeg. 2013, 4, 2. [Google Scholar]
- Tambekar, D.H.; Gadakh, P.V. Biochemical and molecular detection of biosurfactant producing bacteria from soil. Int. J. Life Sci. Biotech. Pharma Res 2013, 2, 204–211. [Google Scholar]
- Juwarkar, A.A.; Nair, A.; Dubey, K.V.; Singh, S.K.; Devotta, S. Biosurfactant technology for remediation of cadmium and lead contaminated soils. Chemosphere 2007, 68, 1996–2002. [Google Scholar] [CrossRef]
- Chakraborty, J.; Das, S. Biosurfactant-based bioremediation of toxic metals. In Microbial Biodegradation and Bioremediation; Elsevier: Amsterdam, The Netherlands, 2014; pp. 167–201. [Google Scholar] [CrossRef]
- Miller, R.M. Biosurfactant facilitated remediation of contaminated soil. Environ. Health Perspect. 1995, 103, 59–62. [Google Scholar] [PubMed]
- Kang, S.W.; Kim, Y.B.; Shin, J.D.; Kim, E.K. Enhanced biodegradation of hydrocarbons in soil by microbial biosurfactant, sophorolipid. Appl. Biochem. Biotechnol. 2010, 160, 780790. [Google Scholar] [CrossRef]
- Kapadia, S.G.; Yagnik, B.N. Current trend and potential for microbial biosurfactants. Asian J. Exp. Biol. Sci. 2013, 4, 1–8. [Google Scholar]
- Mrvčić, J.; Stanzer, D.; Šolić, E.; Stehlik-Tomas, V. Interaction of lactic acid bacteria with metal ions: Opportunities for improving food safety and quality. World J. Microbiol. Biotechnol. 2012, 28, 2771–2782. [Google Scholar] [CrossRef] [PubMed]
- Ahalya, N.; Ramachandra, T.V.; Kanamadi, R.D. Biosorption of heavy metals. Res. J. Chem. Environ. 2003, 7, 71–79. [Google Scholar]
- Escudero, L.B.; Quintas, P.Y.; Wuilloud, R.G.; Dotto, G.L. Recent advances on elemental biosorption. Environ. Chem. Lett. 2019, 17, 409–427. [Google Scholar] [CrossRef]
- Davis, T.A.; Volesky, B.; Mucci, A. A review of the biochemistry of heavy metal biosorption by brown algae. Water Res. 2003, 37, 4311–4330. [Google Scholar] [CrossRef]
- Volesky, B. Biosorption for the next century. In Process Metallurgy; Elsevier: Amsterdam, The Netherlands, 1999; Volume 9, pp. 161–170. [Google Scholar] [CrossRef]
- Chojnacka, K. Biosorption and bioaccumulation–the prospects for practical applications. Environ. Int. 2010, 36, 299–307. [Google Scholar] [CrossRef]
- Yilmazer, P.; Saracoglu, N. Bioaccumulation and biosorption of copper (II) and chromium (III) from aqueous solutions by Pichia stipitis yeast. J. Chem. Technol. Biotechnol. 2009, 84, 604–610. [Google Scholar] [CrossRef]
- Ramrakhiani, L.; Majumder, R.; Khowala, S. Removal of hexavalent chromium by heat inactivated fungal biomass of Termitomycesclypeatus: Surface characterization and mechanism of biosorption. Chem. Eng. J. 2011, 171, 1060–1068. [Google Scholar] [CrossRef]
- Ding, Y.; Jing, D.; Gong, H.; Zhou, L.; Yang, X. Biosorption of aquatic cadmium (II) by unmodified rice straw. Bioresour. Technol. 2012, 114, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K. Biosorption and Bioaccumulation in Practice; Nova Science Publishers: Hauppauge, New York, USA, 2009; p. 137. [Google Scholar]
- Chojnacka, K.; Chojnacki, A.; Gorecka, H. Biosorption of Cr3+, Cd2+ and Cu2+ ions by blue–green algae Spirulina sp.: Kinetics, equilibrium and the mechanism of the process. Chemosphere 2005, 59, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kanamarlapudi, S.L.R.K.; Chintalpudi, V.K.; Muddada, S. Application of biosorption for removal of heavy metals from wastewater. Biosorption 2018, 18, 69. [Google Scholar] [CrossRef]
- Calderón, O.A.R.; Abdeldayem, O.M.; Pugazhendhi, A.; Rene, E.R. Current updates and perspectives of biosorption technology: An alternative for the removal of heavy metals from wastewater. Curr. Pollut. Rep. 2020, 6, 8–27. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Recent trends in microbial biosorption of heavy metals: A review. Biochem. Mol. Biol. 2013, 1, 19–26. [Google Scholar] [CrossRef]
- Villen-Guzman, M.; Gutierrez-Pinilla, D.; Gomez-Lahoz, C.; Vereda-Alonso, C.; Rodriguez-Maroto, J.M.; Arhoun, B. Optimization of Ni (II) biosorption from aqueous solution on modified lemon peel. Environ. Res. 2019, 179, 108849. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A.; Hamouda, R.A.; Mousa, I.E.; Abdel-Hamid, M.S.; Rabei, N.H. Biosorption optimization, characterization, immobilization and application of Gelidiumamansii biomass for complete Pb2+ removal from aqueous solutions. Sci. Rep. 2018, 8, 13456. [Google Scholar] [CrossRef]
- Carlos, M.H.J.; Stefani, P.V.Y.; Janette, A.M.; Melani, M.S.S.; Gabriela, P.O. Assessing the effects of heavy metals in ACC deaminase and IAA production on plant growth-promoting bacteria. Microbiol. Res. 2016, 188, 53–61. [Google Scholar] [CrossRef]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov, A.Y.; Tikhonovich, I.A.; Kluge, C.; Preisfeld, A.; et al. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2001, 47, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, H.; Vessely, S.; Shah, S.; Glick, B.R. Effect of a nickel-tolerant ACC deaminase-producing Pseudomonas strain on growth of non transformed and transgenic canola plants. Curr. Microbiol. 2008, 57, 170–174. [Google Scholar] [CrossRef]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef]
- Strong, P.J.; Xie, S.; Clarke, W.P. Methane as a resource: Can the methanotrophs add value? Environ. Sci. Technol. 2015, 49, 4001–4018. [Google Scholar] [CrossRef]
- Wilshusen, J.H.; Hettiaratchi, J.P.; De Visscher, A.; Saint-Fort, R. Methane oxidation and formation of EPS in compost: Effect of oxygen concentration. Environ. Pollut. 2004, 129, 305–314. [Google Scholar] [CrossRef]
- Glick, B. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 1–15. Available online: https://www.scielo.br/scielo.php?script=sci_nlinks&pid=S1415-4757201500040040100093&lng=en (accessed on 1 October 2012). [CrossRef]
- Etesami, H.; Emami, S.; Alikhani, H.A. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects: A review. J. Soil Sci. Plant Nutr. 2017, 17, 897–911. [Google Scholar] [CrossRef]
- Etesami, H. Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues: Mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 147, 175–191. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Plant-growth-promoting rhizobacteria: Drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 2016, 66, 35–42. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Diwan, B. Bacterial exopolysaccharide mediated heavy metal removal: A review on biosynthesis, mechanism and remediation strategies. Biotechnol. Rep. 2017, 13, 58–71. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, V.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol. Fertil. Soils 2009, 46, 17–26. [Google Scholar] [CrossRef]
- Bhaskar, P.V.; Bhosle, N.B. Bacterial extracellular polymeric substance (EPS): A carrier of heavy metals in the marine food-chain. Environ. Int. 2006, 32, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Artyszak, A.; Gozdowski, D. The effect of growth activators and plant growth-promoting rhizobacteria (PGPR) on the soil properties, root yield, and technological quality of sugar beet. Agronomy 2020, 10, 1262. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press Inc.: Cambridge, MA, USA, 2020; Volume 162, pp. 31–87. [Google Scholar]
- Uribe, D.; Sánchez-Nieves, J.; Vanegas, J. Role of microbial biofertilizers in the development of a sustainable agriculture in the Tropics. In Soil Biology and Agriculture in the Tropics; Dion, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 235–250. [Google Scholar]
- Mehnaz, S. An overview of globally available bioformulations. In Bioformulations: For Sustainable Agriculture; Arora, N., Mehnaz, S., Balestrini, R., Eds.; Springer: New Delhi, India, 2016; pp. 268–281. [Google Scholar]
- García-Fraile, P.; Menéndez, E.; Celador-Lera, L.; Díez-Méndez, A.; Jiménez-Gómez, A.; Marcos-García, M.; Cruz-González, X.A.; Martínez-Hidalgo, P.; Mateos, P.F.; Rivas, R. Bacterial probiotics: A truly green revolution. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 131–162. [Google Scholar]
- Adeleke, R.A.; Raimi, A.R.; Roopnarain, A.; Mokubedi, S.M. Status and prospects of bacterial inoculants for sustainable management of agroecosystems. In Biofertilizers for Sustainable Agriculture and Environment; Giri, B., Prasad, R., Wu, Q.-S., Varma, A., Eds.; Springer: Cham, Switzerland, 2019; pp. 137–172. [Google Scholar]
- Aloo, B.N.; Makumba, B.A.; Mbega, E.R. Plant growth promoting rhizobacterial biofertilizers for sustainable crop production: The past, present, and future. Preprints 2020, 2020090650. [Google Scholar] [CrossRef]
- Dal Cortivo, C.; Ferrari, M.; Visioli, G.; Lauro, M.; Fornasier, F.; Barion, G.; Panozzo, A.; Vamerali, T. Effects of seed-applied biofertilizers on rhizosphere biodiversity and growth of common wheat (Triticum aestivum L.) in the field. Front. Plant Sci. 2020, 11, 72. [Google Scholar] [CrossRef]
- Mahajan, A.; Gupta, R.D. Bio-fertilizers: Their kinds and requirement in India. In Integrated Nutrient Management (INM) in a Sustainable Rice—Wheat Cropping System; Mahajan, A., Gupta, R.D., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 75–100. [Google Scholar]
- Malathi, S.; Sriramulu, M. Laboratory efficacy of biotic insecticides against lepidopterous pests fed on treated cabbage leaves. Shashpa 2000, 7, 63–66. [Google Scholar]
- Apurva, K.; Singh, C.; Kumar, V.; Jha, V.B. Bacterial biopesticides and their use in agricultural production. In Biofertilizers and Biopesticides in Sustainable Agriculture; Apple Academic Press: London, UK, 2019; pp. 23–42. [Google Scholar]
- Schwenk, V.; Riegg, J.; Lacroix, M.; Märtlbauer, E.; Jessberger, N. Enteropathogenic potential of Bacillus thuringiensis isolates from soil, animals, food and biopesticides. Foods 2020, 9, 1484. [Google Scholar] [CrossRef]
- Cardinale, M.; Ratering, S.; Suarez, C.; Zapata Montoya, A.M.; Geissler-Plaum, R.; Schnell, S. Paradox of plant growth promotion potential of rhizobacteria and their actual promotion effect on growth of barley (Hordeum vulgare L.) under salt stress. Microbiol. Res. 2015, 181, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, O.; Koren, A.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [PubMed]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G. An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evolut. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M.A.; Santoyo, G. Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J. Environ. Manag. 2020, 273, 111118. [Google Scholar] [CrossRef]
- Kumar, S.S.; Kadier, A.; Malyan, S.K.; Ahmad, A.; Bishnoi, N.R. Phytoremediation and rhizoremediation: Uptake, mobilization and sequestration of heavy metals by plants. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Springer: New Delhi, India, 2017; pp. 367–394. [Google Scholar]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.S.; Saxena, A.K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatal. Agric. Biotechnol. 2020, 23, 101487. [Google Scholar] [CrossRef]
- Raj, S.N.; Shetty, H.S.; Reddy, M.S. Plant growth promoting rhizobacteria: Potential green alternative for plant productivity. In PGPR: Biocontrol and Biofertilization; Springer: Dordrecht, The Netherlands, 2005; pp. 197–216. [Google Scholar]
- Son, J.S.; Sumayo, M.; Hwang, Y.J.; Kim, B.S.; Ghim, S.Y. Screening of plant growth-promoting rhizobacteria as elicitor of systemic resistance against gray leaf spot disease in pepper. Appl. Soil Ecol. 2014, 73, 1–8. [Google Scholar] [CrossRef]
- Liu, J.; Dang, P.; Gao, Y.; Zhu, H.; Zhu, H.; Zhao, F.; Zhao, Z. Effects of tree species and soil properties on the composition and diversity of the soil bacterial community following afforestation. For. Ecol. Manag. 2018, 427, 342–349. [Google Scholar] [CrossRef]
- Bhattacharjee, R.B.; Singh, A.; Mukhopadhyay, S.N. Use of nitrogen-fixing bacteria as biofertiliser for non-legumes: Prospects and challenges. Appl. Microbiol. Biotechnol. 2008, 80, 199–209. [Google Scholar] [CrossRef]
- Chen, J.H. The combined use of chemical and organic fertilizers and/or biofertilizer for crop growth and soil fertility. In Proceedings of the International Workshop on Sustained Management of the Soilrhizosphere System for Efficient Crop Production and Fertilizer Use, Land Development Department, Bangkok, Thailand, 16–20 October 2006; Volume 16, p. 20. [Google Scholar]
- Kumawat, N.; Kumar, R.; Kumar, S.; Meena, V.S. Nutrient solubilizing microbes (NSMs): Its role in sustainable crop production. In Agriculturally Important Microbes for Sustainable Agriculture; Springer: Singapore, 2017; pp. 25–61. [Google Scholar]
- Ortiz-Castro, R.; López-Bucio, J. Phytostimulation and root architectural responses to quorum-sensing signals and related molecules from rhizobacteria. Plant Sci. 2019, 284, 135–142. [Google Scholar] [CrossRef]
- Meena, M.; Swapnil, P.; Divyanshu, K.; Kumar, S.; Tripathi, Y.N.; Zehra, A.; Marwal, A.; Upadhyay, R.S. PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: Current perspectives. J. Basic Microbiol. 2020, 60, 828–861. [Google Scholar] [PubMed]
- Zandi, P.; Basu, S.K. Role of plant growth-promoting rhizobacteria (PGPR) as biofertilizers in stabilizing agricultural ecosystems. Org. Farming Sustain. Agric. 2016, 71–87. [Google Scholar] [CrossRef]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A potential approach for sustainable agriculture development. Environ. Sci. Pollut. Res. 2017, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Aka, R.J.N.; Babalola, O.O. Effect of bacterial inoculation of strains of Pseudomonas aeruginosa, Alcaligenes feacalis and Bacillus subtilis on germination, growth and heavy metal (Cd, Cr, and Ni) uptake of Brassica juncea. Int. J. Phytoremediat. 2016, 18, 200–209. [Google Scholar]
- Dangi, S.R.; Tirado-Corbalá, R.; Gerik, J.; Hanson, B.D. Effect of long-term continuous fumigation on soil microbial communities. Agronomy 2017, 7, 37. [Google Scholar] [CrossRef]
- Dupuy, L.X.; Mimault, M.; Patko, D.; Ladmiral, V.; Ameduri, B.; MacDonald, M.P.; Ptashnyk, M. Micromechanics of root development in soil. Curr. Opin. Genet. Dev. 2018, 51, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Gundel, P.E.; Omacini, M.; Sadras, V.O.; Ghersa, C.M. The interplay between the effectiveness of the grass-endophyte mutualism and the genetic variability o the host plant. Evol. Appl. 2010, 3, 538–546. [Google Scholar] [CrossRef] [PubMed]




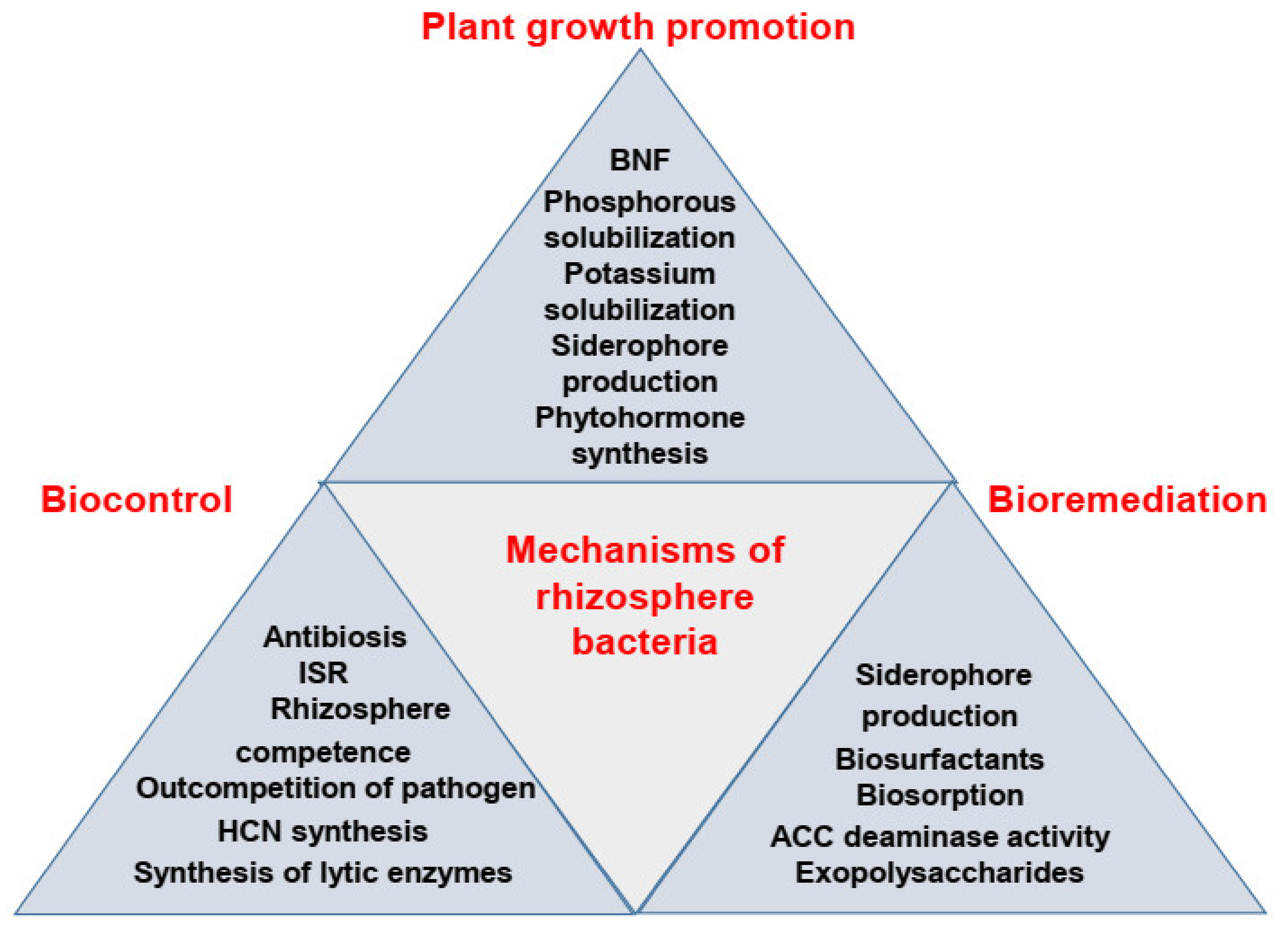{kind=link}
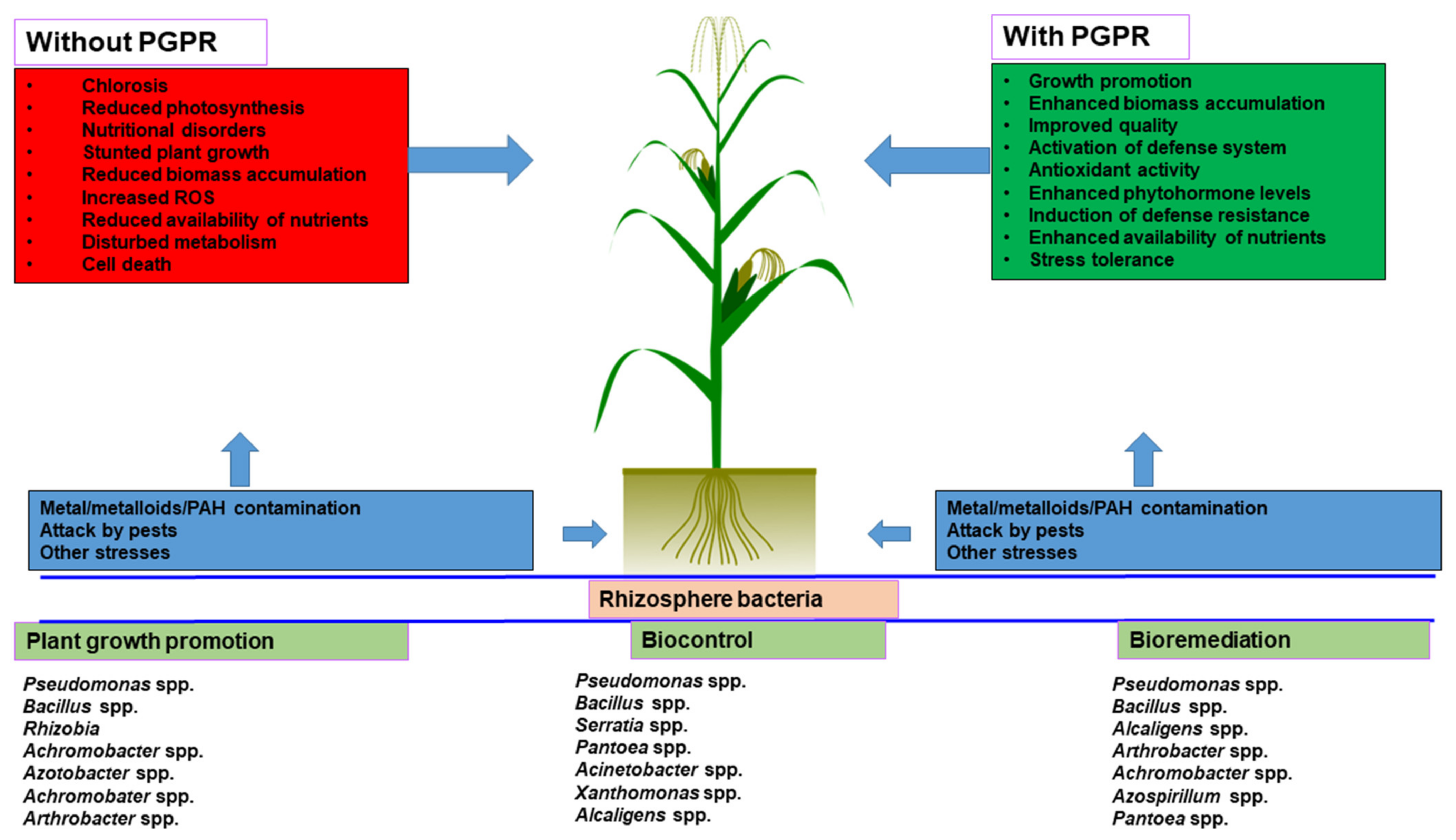{kind=link}
{kind=link}
| PGPR Candidate | Host Plant | Growth Condition | Proposed Mechanism(s) | Plant Response | References |
|---|---|---|---|---|---|
| Rhizobium tropici | Bean | Pot | N2 fixation and IAA production | Increase in number of nodules and plant dry weight | [45] |
| Pseudomonas stutzeri A1501 | Maize | Field | N2 fixation and ACC deaminase production | Better growth and more nitrogen accumulation | [46] |
| Rhizobium etli | Bean | Pot | Nitrogenase activity | A 16% increase in the N2 content of the resultant seeds and a 25–30% increase in leaf nitrogen | [47] |
| Pseudomonas fluorescens BSP53 | Blackcurrant cuttings | Pot and field | IAA overproduction | Root development | [45] |
| Streptomyces sp. NEAU-S7GS2 | Soybean | Pot | IAA production and inorganic phosphate solubilization | A 77% increase in plant growth and a 38% decrease in sclerotinia stem rot of soybean | [48] |
| Halobacillus dabanensis strain SB-26 and Halobacillus sp. GSP 34 | Rice | Pot | IAA production and nitrogen fixation | Enhanced plant height and yield | [49] |
| Fluorescent Pseudomonas sp. PF17 | Sunflower | Pot | IAA production and phosphate solubilization | Promoted overall plant growth | [50] |
| wild-type P. putida GR12-2 | Mung bean | Pot | IAA-overproducing and pH-lowering mechanism | Root and shoot development | [51] |
| R. leguminosarum bv. Viciae | Alfalfa | Pot | ACC deaminase | Increase in nodule number and biomass | [44] |
| Bacillus sp. HWP47 | Wheat | Pot | ACC deaminase activity and chelation mechanism | Caused 51.46% increase in root dry weight (RDW) | [52] |
| Rhizobacteria sp. | Solanum tuberosum | Pot | ACC deaminase activity and phytohormone production | ROS antioxidant enzyme expression and improved photosynthesis | [53] |
| Pseudomonads | Tobacco | Field | Ferric siderophore production | Adventitious root development | [54] |
| Pseudomonas fluorescens C7 | Arabidopsis thaliana | Pot and Field Trials | Fe-pyoverdine complex | Increase in iron inside plant tissues and hence enhanced plant growth | [55] |
| Bacillus mucilaginosus HQ013329 | Garlic | Pot | Various extracellular polymers are produced (primarily proteins and polysaccharides) | Promoted garlic plant growth characteristics and avoided environmental pollution hazards caused by heavy application of potassium fertilizers | [56] |
| Bacillus circulans | Pepper Potato | Pot | Acidolysis of the surrounding area of microorganism | Highest total yield of pepper and potato tuber yield per plant and average tuber weight | [57] |
| B. mucilaginosus strain RCBC13 | Tomato | Pot | Production of α-ketogluconic acid | 125% increase in biomass, whereas K and P uptake were more than 150% in tomato | [58] |
| Pseudomonas aeruginosa PF23EPS+ | Sunflower | Pot and field | Salicylic acid production | Improved growth of crop in salinized soil | [59] |
| Bradyrhizobium japonicum USDA 110 with salt-tolerant Pseudomonas putida TSAU1 | Soybean | Hydroponic | Formation of nodules | Increased plant growth and nitrogen and phosphorus uptake | [60] |
| Bacillus amyloliquefaciens subsp. plantarum UCMB5113 | Arabidopsis thaliana | Lab | Improved root growth and configuration | Increased plant growth and reduced disease effect | [61] |
| Bacillus firmus SW5 | Soybean | Pot | Improved root structure | Increased biomass, nutrient uptake, and gas exchange parameters | [62] |
| Azospirillum brasilense | Maize | Pot and field | Siderophore production | Increased biomass and improved plant health | [63] |
| Acidovorax delafieldii | Rice | Pot | Siderophore production and tricalcium phosphate solubilization | Enhanced biomass, grain yield, and NPK uptake | [64] |
| Brevibacillus brevis GZDF3 | Pinellia ternata (an important herb in traditional Chinese medicine) | Lab | Siderophore production | Helpful in the development of new biological agent | [65] |
| Bacillus subtilis MF497646 and Pseudomonas koreensis MG209738 | Maize | Pot and field | Siderophore production | Increased antioxidant activities and yield parameters | [66] |
| Biocontrol Agent | Plant Pathogen | Host Plant | Proposed Mechanism(s) | Reference |
|---|---|---|---|---|
| Pseudomonas fluorescens | Fusarium culmorum | Rye | Fe(III)-chelating compounds (including siderophores) | [159] |
| Acinetobacter, Pseudomonas, Staphylococcus, Bacillus, Enterobacter, Pantoea, Alcaligenes | Fusarium oxysporum, Alternaria alternate, F. culmorum, F. solani, Botrytis cinerea, Pythium ultimum, Phytophthora cryptogea | Wheat | Antagonism and growth promotion | [155] |
| Bacillus sp. L324-92 | Gaeumannomyces graminis var tritici, Rhizoctonia root rot, R. solani AG8, Pythium root rot, Pythium irregulare P. ultimum. | Wheat | Not specified | [90] |
| Bacillus sp., Pseudomonas fluorescens | R. oryzae, P. ultimum, G. graminis, R. solani | Wheat | Not specified | [160] |
| Pseudomonas fluorescens | Microconidium nivale/ Fusarium nivale | Wheat | Growth promotion, siderophore production, in vitro antibiosis | [161] |
| Bacillus subtilis and B. cereus | Take all (G. graminis var tritici) Rhizoctonia root rot (R. solaniAG8) | Wheat | Growth promotion | [162] |
| Bacillus subtilis CE1 | Fusarium verticillioides | Maize | Not specified | [163] |
| Pseudomonas chlororaphis | Macrophomina phaseolina (charcoal rot of sorghum) | Sorghum | Extracellular antibiotics, production of volatiles, siderophores, effective root colonization | [164] |
| Pseudomonas fluorescens MKB 100 and MKB 249, P. frederiksbergensis 202, Pseudomonas spp. MKB 158 | Fusarium culmorum | Wheat and barley | Induced resistance, antibiotic production, pathogenesis-related proteins (induced resistance) in wheat | [165] |
| PGPR Candidate | Pollutants | Pollutant Concentration | Remediation Efficiency (%) | References |
|---|---|---|---|---|
| Azoarcus sp., Escherichia coli L. | Benzoates | 1 mM | 90.0 | [248] |
| Pseudomonas fluorescens L. and Pseudomonas putida L. | Benzoates and related substances | 1 mM | 60.0 | [227] |
| Rhizobium leguminosarum L. | Chrysene | 500 mg kg-1 | 28.0 | [249] |
| Azospirillum lipoferum L. and Azospirillum brasilense L. | Crude oil | 1% v/v | 57.0 | [228] |
| Acinetobacter lwoffii L., Bacillus subtilis L., and Pseudomonas aeruginosa L. | Crude oil | 1% v/v | 89.0 | [250] |
| Alcaligenes faecalis L., Citrobacter murliniae L., Dietzia papillomatosis L., Nocardioides deserti L. | Crude oil | 1174 mg/L | 90.0 | [251] |
| Enterobacter sp. MN17 | Crude oil | 5000 mg kg−1 | 63.0 | [229] |
| Enterobacter cloacae | Crude oil | 2000 ppm | 54.0 | [230,231] |
| Pseudomonas cepacia L. and Arthrobacter sp. | 2,4-Dichlorphenoxyessigsäure (kurz 2,4-D) | 5% v/v | 80.0 | [236,252] |
| Rhizopus arrhizus L. and Pseudomonas aeruginosa L. | Dichlorodiphenyltrichloroethane (DDT) | 1 mg L−1 | 34.0 | [237,253] |
| Chroococcus sp. and Synechocystis sp. | Linurin | 1 mg L−1 | >98 | [254] |
| Pseudomonas sp., Pseudomonas aeruginosa L., Pseudomonas stutzeri L. | Parathion | 3800 µg/mL | >49 | [255] |
| Archaea sp., Acinetobacter sp., Achromobacter sp., Alcaligenes sp., Aspergillus sp., Arthrobacter sp., Azotobacter sp., Bacillus cereus sp., Flavobacterium sp., Neurospora sp. | Phenolic compounds | 13.0 mg L−1 | 99.0 | [256,257] |
| Leptolyngbya sp., Penicillium sp., Candida tropicalis L., Debaryomyces subglobosus L., Pseudomonas aeruginosa L., Pseudomonas putida L., and Trichosporon cutaneum L. | Phenolic compounds | 150.0 mg L−1 | 99.0 | [258] |
| Pseudomonas sp. | Phenanthrene | 200 mg kg-1 | >62 | [259] |
| Actinobacteria sp. | Phenanthrene | 100 mg L−1 | 50.0 | [260] |
| Pseudomonas putida L. | Phenanthrene | 100 mg kg-1 | 89.0 | [261] |
| Pseudomonas sp. | Phenanthrene | 100 mg kg−1 | 73.0 | [228] |
| Pseudomonas sp. | Phenanthrene | 100 mg kg-1 | 50.0 | [239] |
| Actinobacteria sp., Caulobacterales sp., Rhizobiales sp., Rhodococcus sp., Xanthomonadales sp. | Phenanthrene | 1260 mg kg−1 | 48.0 | [241] |
| Pseudomonas cepacia L. | 2,4,5-Trichlorophenoxyacetic acid (2,4,5-T) | 200 µM | 27.0 | [262] |
| Alcaligene sp., Bacillus sp., Citrobacter sp., Corynebacterium sp., Flavobacterium sp., and Pseudomonas sp. | Surfactants | 200 mg L−1 | 50.0 | [263] |
| Bacillus sp., Candia sp., Pseudomonas sp. | Surfactants | 300 mg L-1 | [264] | |
| Burkholderia multivorans L. | Surfactants | 150 mg L-1 | 41.0 | [265] |
| Pseudomonas sp. | Surfactants | 2% v/v | 94.0 | [266] |
| Heavy Metals | Microbes | Host Plant(s) | Proposed Mechanism(s) | References |
|---|---|---|---|---|
| As | Penicillium aculeatum PDR-4 | Sunflower | Phosphatase, siderophore | [267] |
| Cd | Bacillus sp., Endophytic sp., Rahnella sp., Enterobacter sp., Klebsiella spp., Pseudomonas sp., Arthrobacter sp. TISTR 2220, Agromyces AR33, Brevundimonas Kro13, Ralstonia sp. TISTR 2219, Streptomyces AR17, Variovorax paradoxus L. | Wheat, Mustard, Soybean, Mung bean, Willow, Amaranthus, Sunflower, Basil, Goat willow | Exopolysaccharide production, siderophores | [231,268,269,270,271,272,273,274] |
| Cd | Bacillus badius L., Cronobacter muytjensii L., Drepanomonas revolute L., Euplotes sp., Pseudomonas azotoformans L., Uronema nigricans L. | – | Biosorption | [275,276,277,278] |
| Cd | Alcaligenes sp., Bacillus sp., Pseudomonas sp., Rhodococcus sp., and Variovorax sp. | Canola, Sunflower | ACC deaminase activity and reduction in ethylene levels | [68,279] |
| Co | Enterobacter ludwigii L. | Sunflower | Exopolysaccharide production | [280] |
| Cr | Aerococcus sp., Bacillus sp., Endophytic sp., Rhizobacteria sp., Staphylococcus sp., Ochrobactrum intermedium L., Pseudomonas aeruginosa L., Pseudomonas pseudoalcaligenes L. | Chickpea, Mesquite, Prosopis, Maize, Sorghum, Wheat, Mung bean, Soybean, Sunflower | Exopolysaccharide production | [271,281,282,283,284,285] |
| Cr | Bacillus circulans L., Bacillus megaterium L., Cronobacter muytjensii L., Saccharomyces cerevisiae L. | – | Biosorption | [276,286,287] |
| Cu | Pseudomonas sp., Rhizobacteria sp., Bacillus subtilis L., Brevibacterium halotolerans L., Bacillus pumilus L., Brevibacterium casei MH8a, Paenibacillus polymyxa L., Pseudomonas pseudoalcaligenes L. | Maize, Sunflower, Sorghum, Mustard, Trailing daisy | Exopolysaccharide production, siderophores | [282,288,289,290,291,292] |
| Cu | Aspergillus flavus L., Cronobacter muytjensii L., Pseudomonas azotoformans L., Drepanomonas revolute L., Euplotes sp., Uronema nigricans L., Pichia guilliermondii L. | Maize, Wheat, Sorghum | Biosorption | [275,276,277,293,294] |
| Fe | Pseudomonas sp., Psychrobacter sp. | Castor bean | Exopolysaccharide production, siderophores | [295] |
| Ni | Micrococcus sp., Pseudomonas sp., Psychrobacter sp., Bacillus subtilis SJ-101, Bacillus pumilus L., Rhodococcus erythropolis X79289, Rhodococcus globerulus X80619 | Mustard, Alyssum sp., Alpine pennycress, Castor bean, Mung bean, Wheat, Soybean | Exopolysaccharide production, siderophores | [271,295,296,297,298,299] |
| Ni | Kluyvera ascorbate L. (SUD165), Pseudomonas putida L. (UW4) | Tomato, Canola, Tobacco | ACC deaminase activity and reduction in ethylene levels | [288,300] |
| Pb | Enterobacter sp., Klebsiella sp., Enterobacter ludwigii L., AgromycesAR33, Bacillus subtilis L., Brevibacterium halotolerans L., Bacillus megaterium HKP-1, Bacillus pumilus L., Penicillium aculeatum PDR-4, Pseudomonas pseudoalcaligenes L., Streptomyces AR17 | Sunflower, Mustard, Maize, Sorghum, Goat willow | Exopolysaccharide production, siderophores | [91,231,272,274, 279,280,282,301] |
| Pb | Aspergillus niger L., Bacillus xiamenensis L., Pseudomonas azotoformans L. | – | Biosorption | [277,293,302] |
| Zn | Enterobacter sp., Klebsiella sp., Pseudomonas sp., Rahnella sp., Rhizobacteria sp., Psychrobacter sp. Agromyces AR33, Bacillus megaterium HKP-1, Bacillus subtilis L., Brevibacterium halotolerans L., BrevibacteriumcaseiMH8a, Bacilluspumilus L., Enterobacter ludwigii L., Streptomyces AR17 | Mustard, Sunflower, Castor bean, Maize, Sorghum, Goat willow, | Exopolysaccharide production, siderophores | [272,279,280,282,291,295,303] |
| Zn | Cronobacter muytjensii L., Drepanomonas revolute L., Euplotes sp., Uronema nigricans L. | – | Biosorption | [275,276] |
| Pro’s Types | Product (Bacterial Composition) | References |
|---|---|---|
| Nitrogen fixers | AgriLife NitroFix (A. chroococcum, A. vinelandii, A. diazotrophicus, A. lipoferum, R. japonicum), Ajay Azospirillum (Azospirillum sp.), Azofer (A. brasilense), Azo-N (A. brasilense + A. lipoferum), Azo-N Plus (A. brasilense + A. lipoferum + A. chroococcum), Azoter (A. chroococcum, A. brasilense, B. megaterium), Azotobacterin (A. brasilense B-4485), BactoFil A10 (B. Megaterium + A. brasilense, A. vinelandii), BactoFil Soya (B. japonicum), BiAgro 10 (B. japonicum), Bioboots (Bradyrhizobium sp. + D. acidovorans), Biofix (Rhizobia), BioGro (C. freundii, K. pneumoniae, P. fluorescens), Bio-N (Azospirillum spp.), Cell-Tech (Rhizobia), Custom N2 (P. polymyxa), Dimargon (A. chroococcum), Legume Fix (B. japonicum + Rhizobium sp.), Mamezo (Rhizobia), Nitragin Gold (Rhizobia), Nitrasec (Rhizobium sp.), Nitrofix (Azospirillum sp.), Nodulator (B. Japonicum), Nodulator PRO (B. subtilis + B. Japonicum), Nodulest 10 (B. japonicum), Nodumax (Bradyrhizobium spp.), Phylazonit M (A. chroococcum + B. megaterium), Rhizofer (R. etli), Rhizosum Aqua (Azospirillum sp.), Rhizosum N (A. vinelandii + R. irregularis), Rizo-Liq (Bradyrhizobium sp., + M. ciceri, + Rhizobium spp.), Rizo-Liq Top (B. japonicum), Symbion N (Azospirillum sp. + Rhizobium sp. + Acetobacter sp. + Azotobacter sp.), TagTeam (P. bilaii + Rhizobia), TwinN and TripleN (Azorhizobium sp. + Azoarcus sp. + Azospirillum sp.), Zadspirillum (A. brasilense), | [11,359,360,361,362,363,364,365,366] |
| Nutrient solubilizers | Bio Phos (B. megaterium), Biozink (PGPR consortia), CataPult (Bacillus spp. + G. intraradices), CBF (B. mucilaginosus, + B. subtilis), Fosforina (P. Fluorescens), K Sol B (F. aurantia), P Sol B (P. striata, + B. polymyxa, B. megaterium), Phosphobacterin (B. megaterium), Rhizosum K (F. aurantia), Rhizosum PK (B. megaterium, + F. aurantia, + R. irregularis), Symbion van Plus (B. megaterium), Zn Sol B (T. thiooxidans), | [360,362,363,366,367] |
| Biopesticides | Biobit, Dipel, and Delfin (Bacillus thuringiensis var. kurstaki), Certan (Bacillus thuringiensis var. aizawai), Acrobe, Skectal, Vectobac (Bacillus thuringiensis var. israelensis), Trident, Novodor (Bacillus thuringiensis var. tenebrionis), Ciba-Foil, Agree, Cutlass (Bacillus thuringiensis var. conjugates), MVP, M-Trak (Pseudomonas fluorescens (Bt toxin), Doom (Bacillus papilliae), Invade (Serratia entomophila) | [368,369,370] |
| Other biofertilizers | Amase (P. azotoformans), Bioativo (PGPR consortia), EVL Coating (PGPR consortia), Biotilis (B. subtilis), Cedomon (P. chlororaphis), Cedress (P. chlororaphis) | [360,362] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. https://doi.org/10.3390/ijms221910529
Saeed Q, Xiukang W, Haider FU, Kučerik J, Mumtaz MZ, Holatko J, Naseem M, Kintl A, Ejaz M, Naveed M, et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. International Journal of Molecular Sciences. 2021; 22(19):10529. https://doi.org/10.3390/ijms221910529
Chicago/Turabian StyleSaeed, Qudsia, Wang Xiukang, Fasih Ullah Haider, Jiří Kučerik, Muhammad Zahid Mumtaz, Jiri Holatko, Munaza Naseem, Antonin Kintl, Mukkaram Ejaz, Muhammad Naveed, and et al. 2021. "Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms" International Journal of Molecular Sciences 22, no. 19: 10529. https://doi.org/10.3390/ijms221910529
APA StyleSaeed, Q., Xiukang, W., Haider, F. U., Kučerik, J., Mumtaz, M. Z., Holatko, J., Naseem, M., Kintl, A., Ejaz, M., Naveed, M., Brtnicky, M., & Mustafa, A. (2021). Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. International Journal of Molecular Sciences, 22(19), 10529. https://doi.org/10.3390/ijms221910529