
Specific Incorporation of Polyunsaturated Fatty Acids into the sn-2 Position of Phosphatidylglycerol Accelerates Photodamage to Photosystem II under Strong Light

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
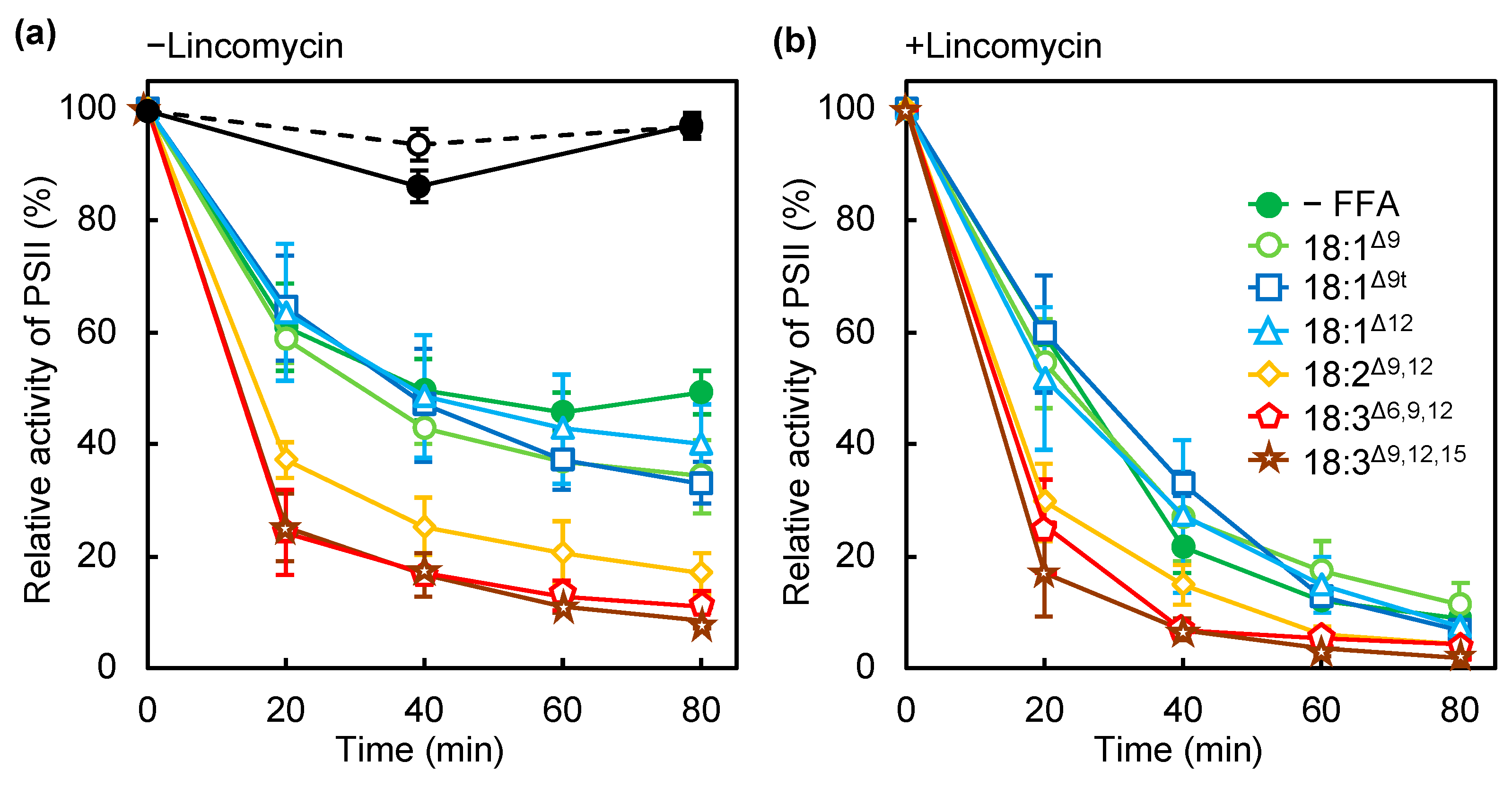
2.1. PUFAs Accelerate Photodamage by Inhibiting Electron Transfer at the Oxygen-Evolving Complex
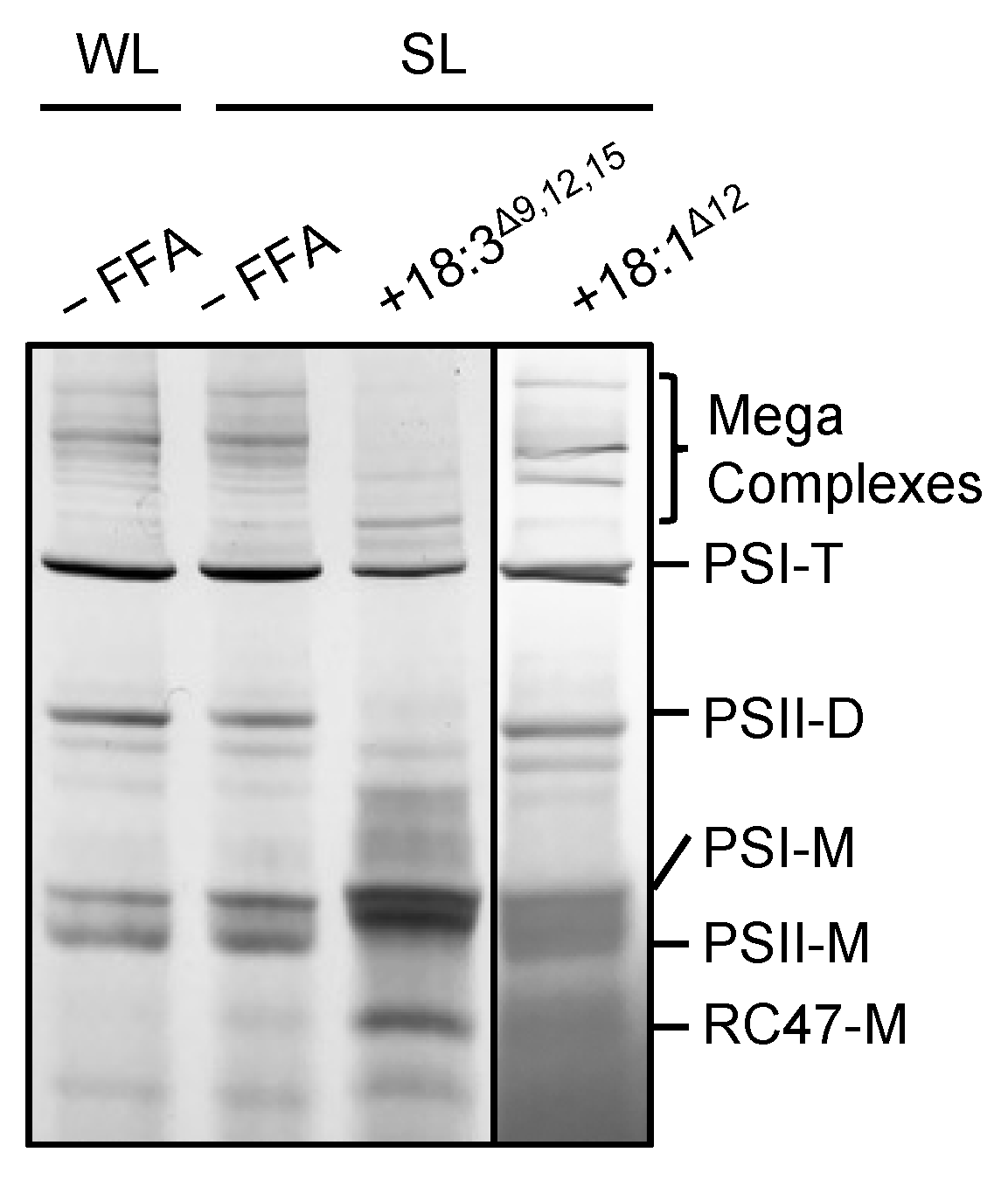
2.2. 18:3Δ9,12,15 Destabilizes Photosynthetic Complexes under Strong Light
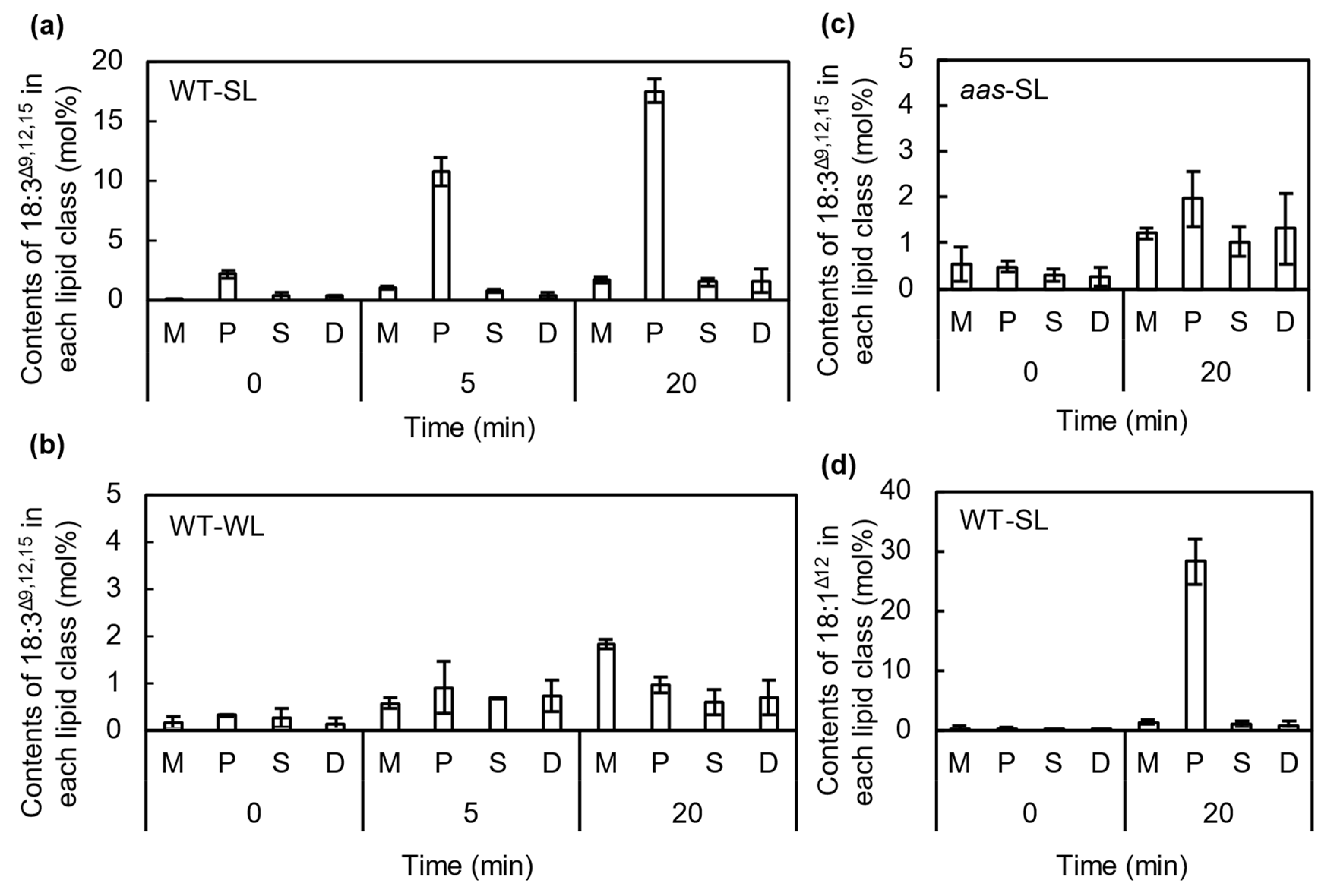
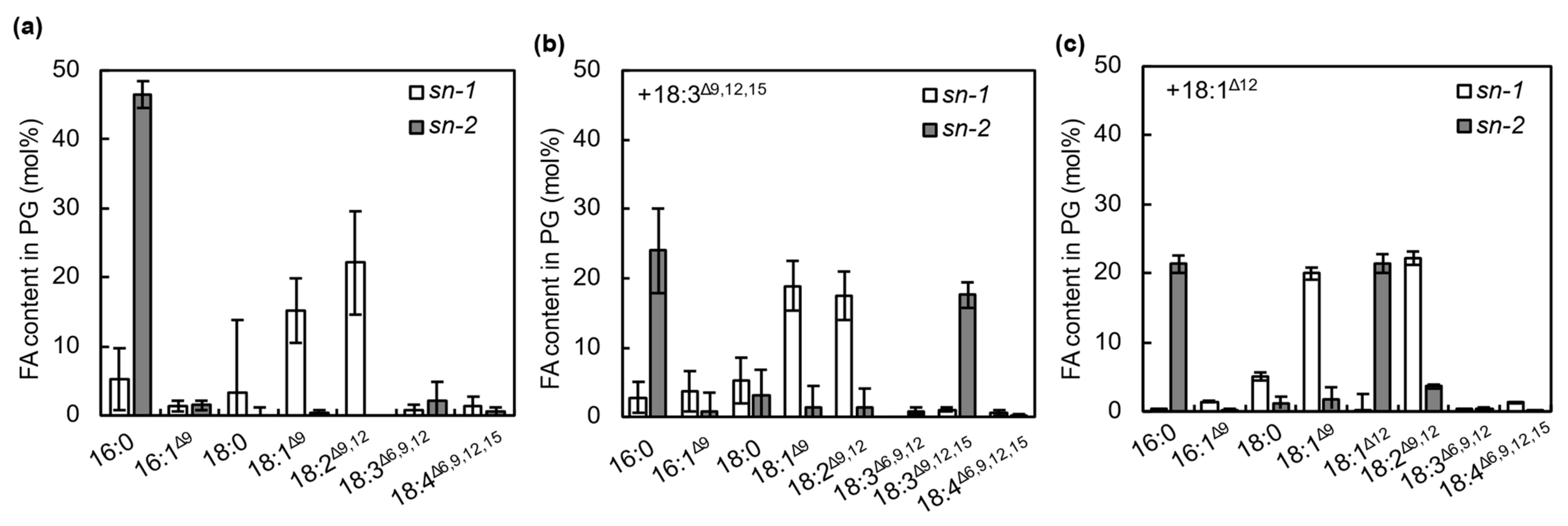
2.3. FFAs Are Incorporated into the sn-2 Position of PG Molecules by the Action of AAS under Strong Light
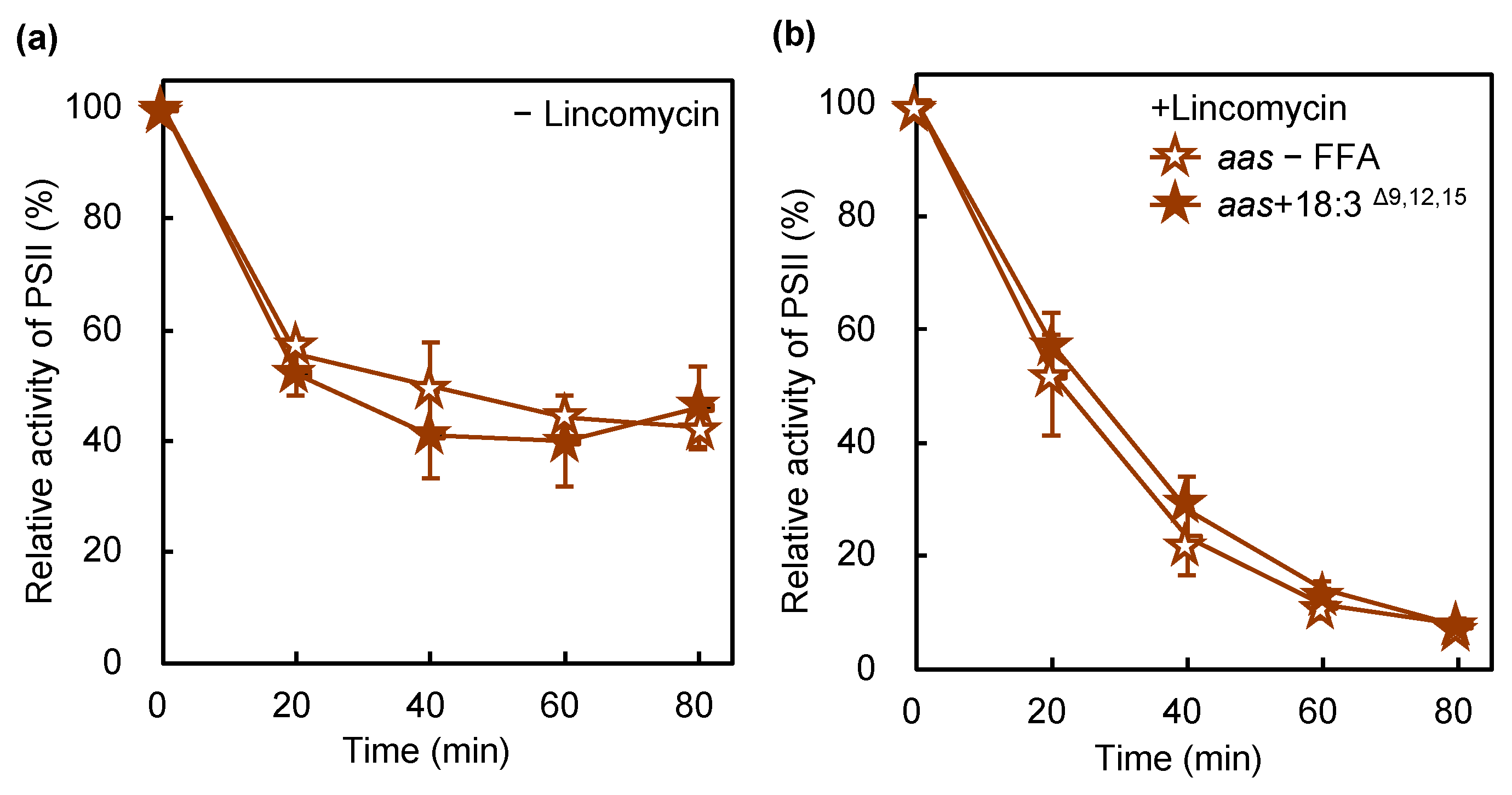
2.4. Disruption of the Gene for AAS Protects PSII against PUFA-Induced Photoinhibition
3. Discussion
3.1. Effect of PUFAs on Photodamage to PSII
3.2. Incorporation of PUFAs into PG
4. Materials and Methods
4.1. Strains and Culture Conditions
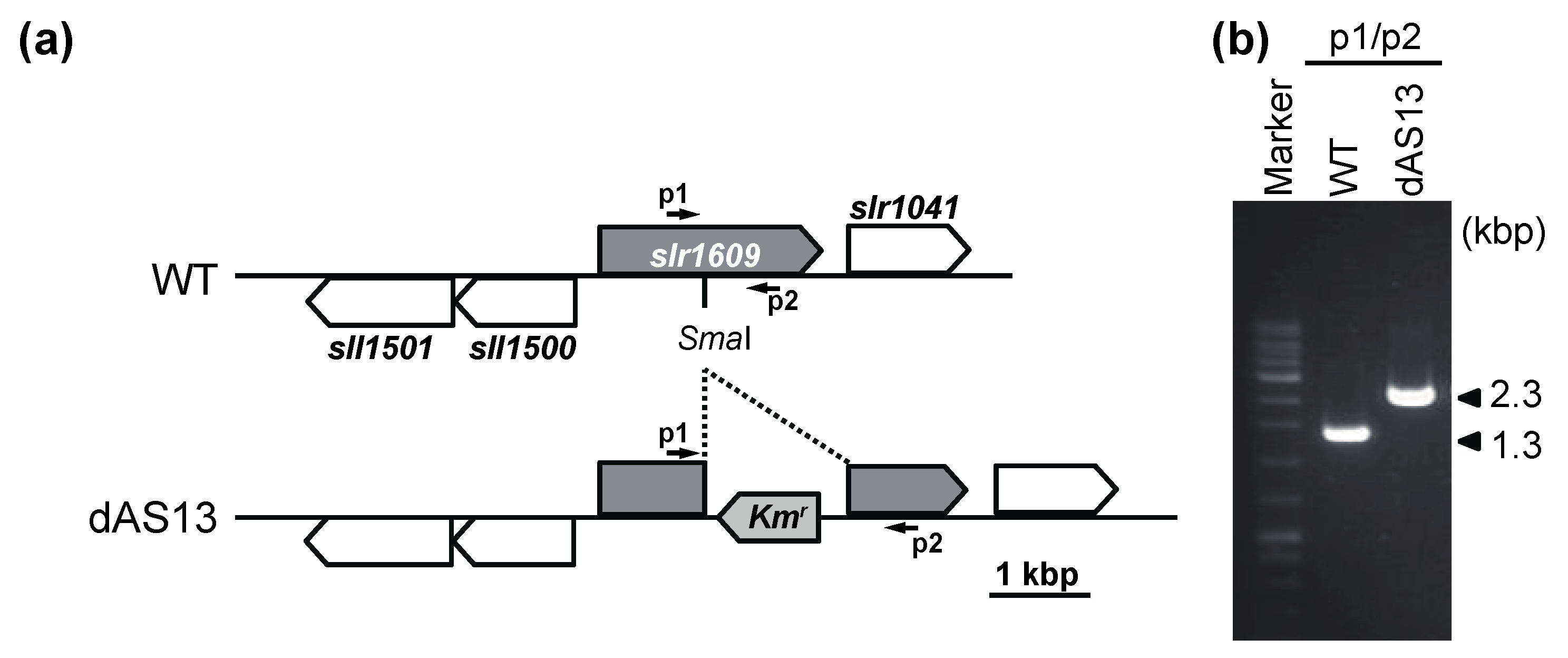
4.2. Targeted Inactivation of aas in Synechocystis
4.3. Photosynthetic Activity of PSII
4.4. Analysis of Photosynthetic Complexes by Blue-Native Polyacrylamide Gel Electrophoresis (PAGE)
4.5. Lipid Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jimbo, H.; Takagi, K.; Hirashima, T.; Nishiyama, Y.; Wada, H. Long-chain saturated fatty acids, palmitic and stearic acids, enhance the repair of photosystem II. Int. J. Mol. Sci. 2020, 21, 7509. [Google Scholar] [CrossRef]
- Ruffing, A.M.; Trahan, C.A. Biofuel toxicity and mechanisms of biofuel tolerance in three model cyanobacteria. Algal Res. 2014, 5, 121–132. [Google Scholar] [CrossRef]
- Von Berlepsch, S.; Kunz, H.H.; Brodesser, S.; Fink, P.; Marin, K.; Flugge, U.I.; Gierth, M. The acyl–acyl carrier protein synthetase from Synechocystis sp. PCC 6803 mediates fatty acid import. Plant Physiol. 2012, 159, 606–617. [Google Scholar] [CrossRef] [Green Version]
- Kojima, K.; Keta, S.; Uesaka, K.; Kato, A.; Takatani, N.; Ihara, K.; Omata, T.; Aichi, M. A simple method for isolation and construction of markerless cyanobacterial mutants defective in acyl-acyl carrier protein synthetase. Appl. Microbiol. Biotechnol. 2016, 100, 10107–10113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarzyk, D.; Fulda, M. Fatty acid activation in cyanobacteria mediated by acyl–acyl carrier protein synthetase enables fatty acid recycling. Plant Physiol. 2010, 152, 1598–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimbo, H.; Izuhara, T.; Hihara, Y.; Hisabori, T.; Nishiyama, Y. Light-inducible expression of translation factor EF-Tu during acclimation to strong light enhances the repair of photosystem II. Proc. Natl. Acad. Sci. USA 2019, 116, 21268–21273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, B.Y.; Higashi, S.; Gombos, Z.; Murata, N. Unsaturation of the membrane lipids of chloroplasts stabilizes the photosynthetic machinery against low-temperature photoinhibition in transgenic tobacco plants. Proc. Natl. Acad. Sci. USA 1995, 92, 6219–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyystjarvi, E.; Aro, E.M. The rate constant of photoinhibition, measured in lincomycin-treated leaves, is directly proportional to light intensity. Proc. Natl. Acad. Sci. USA 1996, 93, 2213–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kok, B.; Businger, J.A. Kinetics of photosynthesis and photo-inhibition. Nature 1956, 177, 135–136. [Google Scholar] [CrossRef]
- Vass, I. Molecular mechanisms of photodamage in the Photosystem II complex. Biochim. Biophys. Acta 2012, 1817, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Kusama, Y.; Inoue, S.; Jimbo, H.; Takaichi, S.; Sonoike, K.; Hihara, Y.; Nishiyama, Y. Zeaxanthin and echinenone protect the repair of photosystem II from inhibition by singlet oxygen in Synechocystis sp. PCC 6803. Plant Cell Physiol. 2015, 56, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Jimbo, H.; Yutthanasirikul, R.; Nagano, T.; Hisabori, T.; Hihara, Y.; Nishiyama, Y. Oxidation of translation factor EF-Tu inhibits the repair of photosystem II. Plant Physiol. 2018, 176, 2691–2699. [Google Scholar] [CrossRef] [Green Version]
- Jimbo, H.; Noda, A.; Hayashi, H.; Nagano, T.; Yumoto, I.; Orikasa, Y.; Okuyama, H.; Nishiyama, Y. Expression of a highly active catalase VktA in the cyanobacterium Synechococcus elongatus PCC 7942 alleviates the photoinhibition of photosystem II. Photosynth. Res. 2013, 117, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Hakala, M.; Tuominen, I.; Keranen, M.; Tyystjarvi, T.; Tyystjarvi, E. Evidence for the role of the oxygen-evolving manganese complex in photoinhibition of photosystem II. Biochim. Biophys. Acta 2005, 1706, 68–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, N.; Allakhverdiev, S.I.; Takahashi, S.; Higashi, S.; Watanabe, M.; Nishiyama, Y.; Murata, N. Two-step mechanism of photodamage to photosystem II: Step 1 occurs at the oxygen-evolving complex and step 2 occurs at the photochemical reaction center. Biochemistry 2005, 44, 8494–8499. [Google Scholar] [CrossRef]
- Zavafer, A.; Cheah, M.H.; Hillier, W.; Chow, W.S.; Takahashi, S. Photodamage to the oxygen evolving complex of photosystem II by visible light. Sci. Rep. 2015, 5, 16363. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, N.; Kashino, Y.; Satoh, K.; Ozawa, S.; Takahashi, Y. Chloroplast-encoded polypeptide PsbT is involved in the repair of primary electron acceptor QA of photosystem II during photoinhibition in Chlamydomonas reinhardtii. J. Biol. Chem. 2007, 282, 7107–7115. [Google Scholar] [CrossRef] [Green Version]
- Umena, Y.; Kawakami, K.; Shen, J.R.; Kamiya, N. Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 A. Nature 2011, 473, 55–60. [Google Scholar] [CrossRef]
- Endo, K.; Kobayashi, K.; Wang, H.T.; Chu, H.A.; Shen, J.R.; Wada, H. Site-directed mutagenesis of two amino acid residues in cytochrome b559 α subunit that interact with a phosphatidylglycerol molecule (PG772) induces quinone-dependent inhibition of photosystem II activity. Photosynth. Res. 2018, 139, 267–279. [Google Scholar] [CrossRef]
- Wada, H.; Murata, N. Temperature-induced changes in the fatty acid composition of the cyanobacterium, Synechocystis PCC6803. Plant Physiol. 1990, 92, 1062–1069. [Google Scholar] [CrossRef] [Green Version]
- Wada, H.; Murata, N. Synechocystis PCC 6803 mutants defective in desaturation of fatty acids. Plant Cell Physiol. 1989, 30, 971–978. [Google Scholar] [CrossRef]
- Murata, N.; Wada, H.; Gombos, Z. Modes of fatty-acid desaturation in cyanobacteria. Plant Cell Physiol. 1992, 33, 933–941. [Google Scholar]
- Laczko-Dobos, H.; Frycak, P.; Ughy, B.; Domonkos, I.; Wada, H.; Prokai, L.; Gombos, Z. Remodeling of phosphatidylglycerol in Synechocystis PCC 6803. Biochim. Biophys. Acta 2010, 1801, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Takatani, N.; Use, K.; Kato, A.; Ikeda, K.; Kojima, K.; Aichi, M.; Maeda, S.; Omata, T. Essential role of acyl-ACP synthetase in acclimation of the cyanobacterium Synechococcus elongatus strain PCC 7942 to high-light conditions. Plant Cell Physiol. 2015, 56, 1608–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, K.; Sato, N.; Tsuji, N.; Tsuzuki, M.; Nishida, I. The significance of C16 fatty acids in the sn-2 positions of glycerolipids in the photosynthetic growth of Synechocystis sp. PCC 6803. Plant Physiol. 2006, 141, 546–556. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; He, Q.; Peng, Z.; Yu, J.; Bian, F.; Li, Y.; Bi, Y. Production of γ-linolenic acid and stearidonic acid by Synechococcus sp. PCC7002 containing cyanobacterial fatty acid desaturase genes. Chin. J. Oceanol. Limnol. 2016, 34, 772–780. [Google Scholar] [CrossRef]
- Murata, N.; Wada, H. Acyl-lipid desaturases and their importance in the tolerance and acclimatization to cold of cyanobacteria. Biochem. J. 1995, 308, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kis, M.; Zsiros, O.; Farkas, T.; Wada, H.; Nagy, F.; Gombos, Z. Light-induced expression of fatty acid desaturase genes. Proc. Natl. Acad. Sci. USA 1998, 95, 4209–4214. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, I.; Hagio, M.; Gombos, Z.; Tyystjarvi, T.; Paakkarinen, V.; Aro, E.M.; Wada, H. Requirement of phosphatidylglycerol for maintenance of photosynthetic machinery. Plant Physiol. 2003, 133, 1376–1384. [Google Scholar] [CrossRef] [Green Version]
- Hagio, M.; Gombos, Z.; Varkonyi, Z.; Masamoto, K.; Sato, N.; Tsuzuki, M.; Wada, H. Direct evidence for requirement of phosphatidylglycerol in photosystem II of photosynthesis. Plant Physiol. 2000, 124, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Jimbo, H.; Izuhara, T.; Hirashima, T.; Endo, K.; Nakamura, Y.; Wada, H. Membrane lipid remodeling is required for photosystem II function under low CO2. Plant J. 2021, 105, 245–253. [Google Scholar] [CrossRef]
- Mizusawa, N.; Wada, H. The role of lipids in photosystem II. Biochim. Biophys. Acta 2012, 1817, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Domonkos, I.; Malec, P.; Sallai, A.; Kovacs, L.; Itoh, K.; Shen, G.; Ughy, B.; Bogos, B.; Sakurai, I.; Kis, M.; et al. Phosphatidylglycerol is essential for oligomerization of photosystem I reaction center. Plant Physiol. 2004, 134, 1471–1478. [Google Scholar] [CrossRef] [Green Version]
- Kruse, O.; Hankamer, B.; Konczak, C.; Gerle, C.; Morris, E.; Radunz, A.; Schmid, G.H.; Barber, J. Phosphatidylglycerol is involved in the dimerization of photosystem II. J. Biol. Chem. 2000, 275, 6509–6514. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, I.; Mizusawa, N.; Ohashi, S.; Kobayashi, M.; Wada, H. Effects of the lack of phosphatidylglycerol on the donor side of photosystem II. Plant Physiol. 2007, 144, 1336–1346. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Endo, K.; Wada, H. Specific distribution of phosphatidylglycerol to photosystem complexes in the thylakoid membrane. Front. Plant Sci. 2017, 8, 1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankaran, K.; Wu, H.C. Lipid modification of bacterial prolipoprotein. Transfer of diacylglyceryl moiety from phosphatidylglycerol. J. Biol. Chem. 1994, 269, 19701–19706. [Google Scholar] [CrossRef]
- Fagerlund, R.D.; Eaton-Rye, J.J. The lipoproteins of cyanobacterial photosystem II. J. Photochem. Photobiol. B 2011, 104, 191–203. [Google Scholar] [CrossRef]
- Sveshnikov, D.; Funk, C.; Schroder, W.P. The PsbP-like protein (sll1418) of Synechocystis sp. PCC 6803 stabilises the donor side of Photosystem II. Photosynth. Res. 2007, 93, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Kashino, Y.; Inoue-Kashino, N.; Roose, J.L.; Pakrasi, H.B. Absence of the PsbQ protein results in destabilization of the PsbV protein and decreased oxygen evolution activity in cyanobacterial photosystem II. J. Biol. Chem. 2006, 281, 20834–20841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoppová, J.; Yu, J.; Janouškovec, J.; Halada, P.; Nixon, P.J.; Whitelegge, J.P.; Komenda, J. The Photosystem II assembly factor Ycf48 from the cyanobacterium Synechocystis sp. PCC 6803 is lipidated using an atypical lipobox sequence. Int. J. Mol. Sci. 2021, 22, 3733. [Google Scholar] [CrossRef]
- Weier, D.; Muller, C.; Gaspers, C.; Frentzen, M. Characterisation of acyltransferases from Synechocystis sp. PCC 6803. Biochem. Biophys. Res. Commun. 2005, 334, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Eungrasamee, K.; Miao, R.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Improved lipid production via fatty acid biosynthesis and free fatty acid recycling in engineered Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2019, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Xu, J.; Lu, T.; Zhang, Q.; Qu, Q.; Yang, Z.; Pan, X. Responses of unicellular alga Chlorella pyrenoidosa to allelochemical linoleic acid. Sci. Total Environ. 2018, 625, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Huang, A.H. Plastid lysophosphatidyl acyltransferase is essential for embryo development in Arabidopsis. Plant Physiol. 2004, 134, 1206–1216. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Wakao, S.; Fan, J.; Benning, C. Loss of plastidic lysophosphatidic acid acyltransferase causes embryo-lethality in Arabidopsis. Plant Cell Physiol. 2004, 45, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Ajjawi, I.; Manoli, A.; Sawin, A.; Xu, C.; Froehlich, J.E.; Last, R.L.; Benning, C. Fatty Acid Desaturase4 of Arabidopsis encodes a protein distinct from characterized fatty acid desaturases. Plant J. 2009, 60, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Browse, J.; Warwick, N.; Somerville, C.R.; Slack, C.R. Fluxes through the prokaryotic and eukaryotic pathways of lipid synthesis in the ‘16:3’ plant Arabidopsis thaliana. Biochem. J. 1986, 235, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Benning, C. Chloroplast lipid synthesis and lipid trafficking through ER-plastid membrane contact sites. Biochem. Soc. Trans. 2012, 40, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arondel, V.; Lemieux, B.; Hwang, I.; Gibson, S.; Goodman, H.M.; Somerville, C.R. Map-based cloning of a gene controlling omega-3 fatty acid desaturation in Arabidopsis. Science 1992, 258, 1353–1355. [Google Scholar] [CrossRef]
- Okuley, J.; Lightner, J.; Feldmann, K.; Yadav, N.; Lark, E.; Browse, J. Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell 1994, 6, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Li, Y.; Huang, A.H. Ubiquitous and endoplasmic reticulum-located lysophosphatidyl acyltransferase, LPAT2, is essential for female but not male gametophyte development in Arabidopsis. Plant Cell 2005, 17, 1073–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.P.; Wolk, C.P. Use of a conditionally lethal gene in Anabaena sp. strain PCC 7120 to select for double recombinants and to entrap insertion sequences. J. Bacteriol. 1990, 172, 3138–3145. [Google Scholar] [CrossRef] [Green Version]
- Bricker, T.M.; Morvant, J.; Masri, N.; Sutton, H.M.; Frankel, L.K. Isolation of a highly active Photosystem II preparation from Synechocystis 6803 using a histidine-tagged mutant of CP 47. Biochim. Biophys. Acta 1998, 1409, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Ichihara, K.; Shibahara, A.; Yamamoto, K.; Nakayama, T. An improved method for rapid analysis of the fatty acids of glycerolipids. Lipids 1996, 31, 535–539. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimbo, H.; Yuasa, K.; Takagi, K.; Hirashima, T.; Keta, S.; Aichi, M.; Wada, H. Specific Incorporation of Polyunsaturated Fatty Acids into the sn-2 Position of Phosphatidylglycerol Accelerates Photodamage to Photosystem II under Strong Light. Int. J. Mol. Sci. 2021, 22, 10432. https://doi.org/10.3390/ijms221910432
Jimbo H, Yuasa K, Takagi K, Hirashima T, Keta S, Aichi M, Wada H. Specific Incorporation of Polyunsaturated Fatty Acids into the sn-2 Position of Phosphatidylglycerol Accelerates Photodamage to Photosystem II under Strong Light. International Journal of Molecular Sciences. 2021; 22(19):10432. https://doi.org/10.3390/ijms221910432
Chicago/Turabian StyleJimbo, Haruhiko, Koki Yuasa, Kensuke Takagi, Takashi Hirashima, Sumie Keta, Makiko Aichi, and Hajime Wada. 2021. "Specific Incorporation of Polyunsaturated Fatty Acids into the sn-2 Position of Phosphatidylglycerol Accelerates Photodamage to Photosystem II under Strong Light" International Journal of Molecular Sciences 22, no. 19: 10432. https://doi.org/10.3390/ijms221910432
APA StyleJimbo, H., Yuasa, K., Takagi, K., Hirashima, T., Keta, S., Aichi, M., & Wada, H. (2021). Specific Incorporation of Polyunsaturated Fatty Acids into the sn-2 Position of Phosphatidylglycerol Accelerates Photodamage to Photosystem II under Strong Light. International Journal of Molecular Sciences, 22(19), 10432. https://doi.org/10.3390/ijms221910432