
Sexual Experience Induces the Expression of Gastrin-Releasing Peptide and Oxytocin Receptors in the Spinal Ejaculation Generator in Rats

 ,
,
Abstract
1. Introduction
2. Results
2.1. Experiment 1: Changes in the Activation of dXL Neurons between Naïve and Sexually Experienced Male Rats
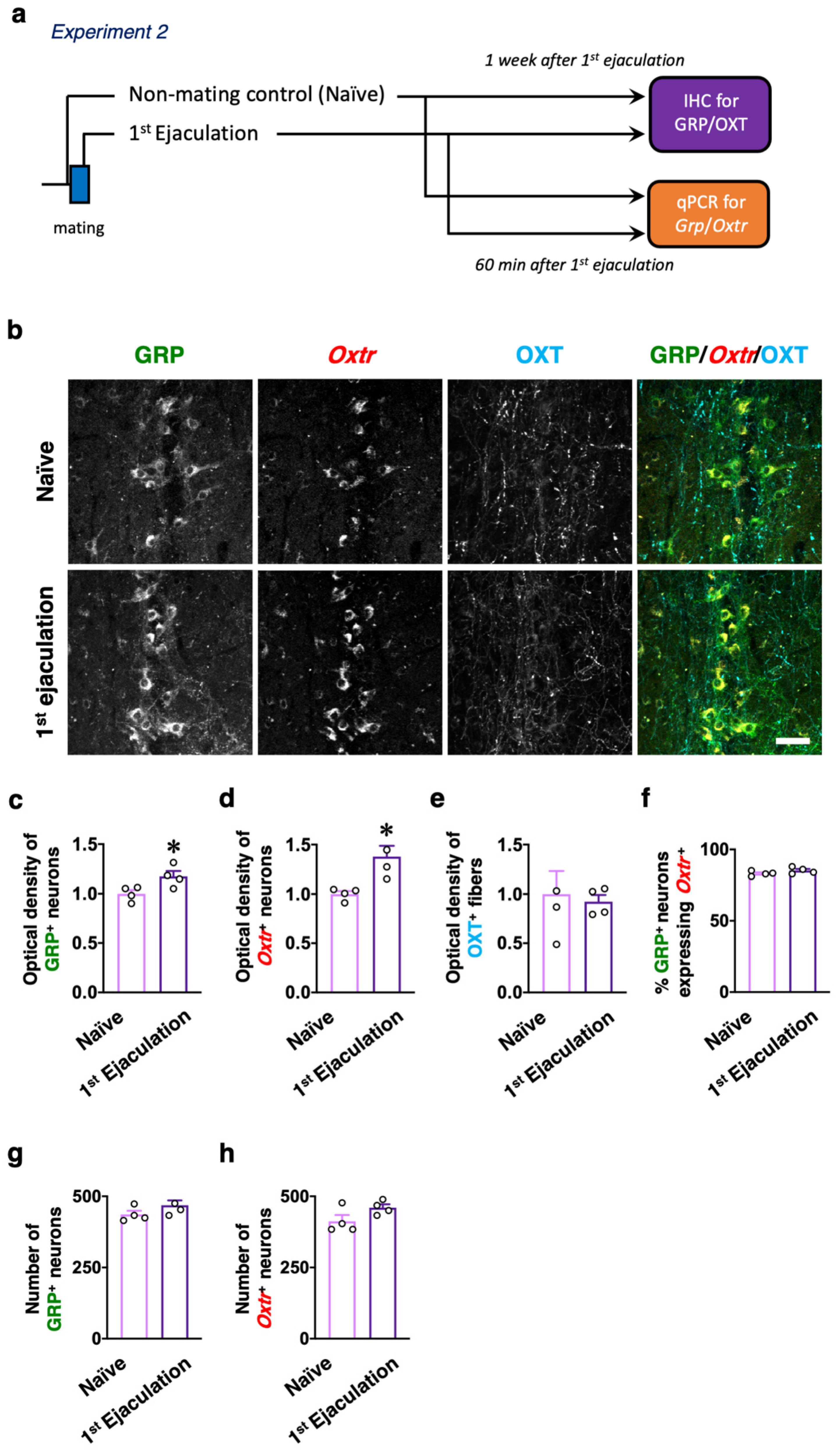
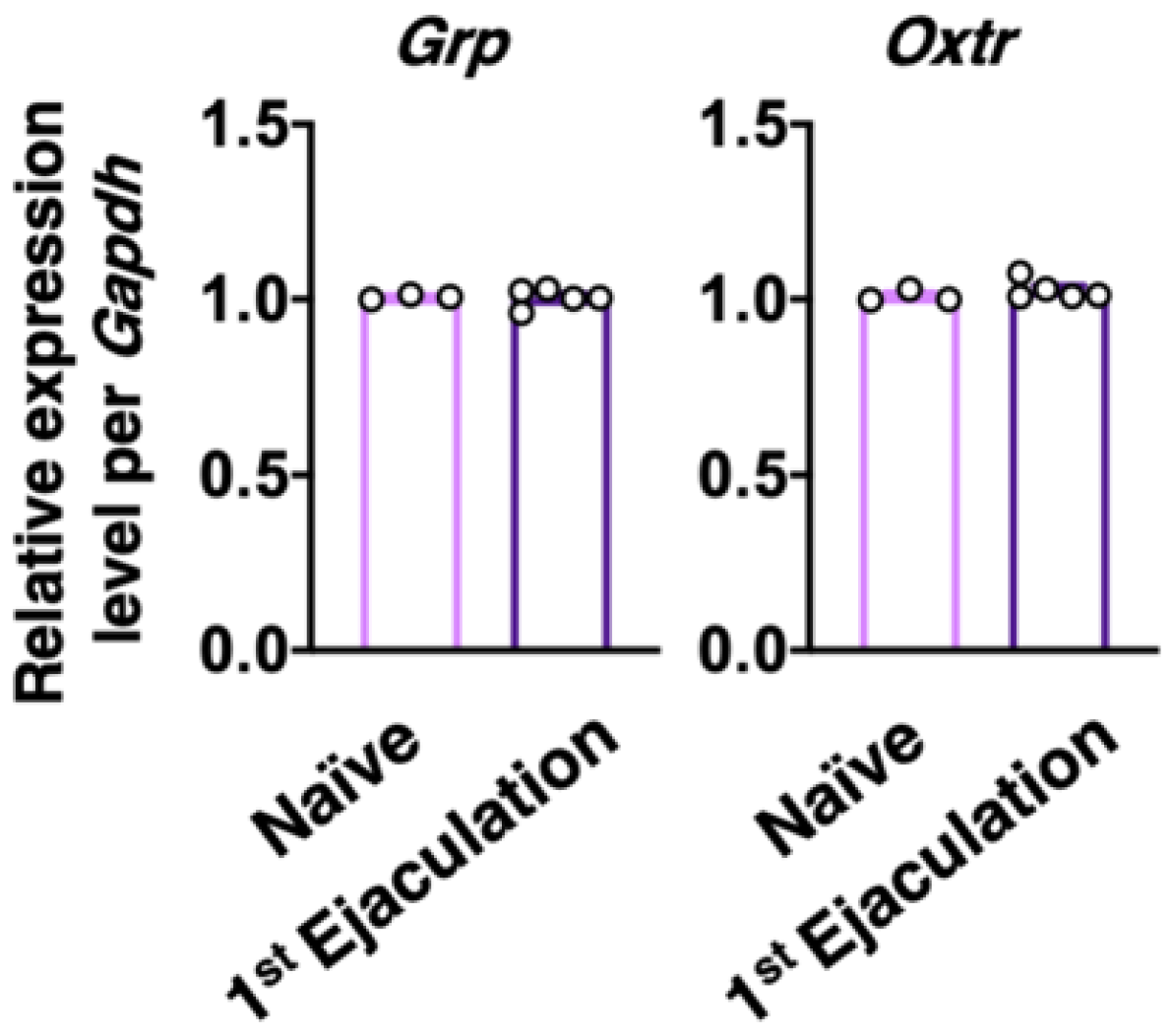
2.2. Experiment 2: Effects of Sexual Experience on the dXL Neuron System
2.3. Experiment 3: Evaluation of Whether Reflexive Erection or Ejaculation Activates dXL Neurons
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Effects of Sexual Experience on the Activation of dXL Neurons
4.3. Protein Expression of GRP, OXT, and YFP in the Lumbar Spinal Cord
4.4. Effects of Sexual Experience on the Grp and Oxtr mRNA Expressions in the Upper Lumbar Spinal Cord
4.5. The Activation of dXL Neurons after Reflexive Erection or PCA-Induced Ejaculation
4.6. Sexual Behavior Test
4.7. Immunohistochemistry and Immunofluorescence
4.8. Morphological Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SEG | spinal ejaculation generator |
| LSt | lumbar spinothalamic |
| GRP | gastrin-releasing peptide |
| dXL | dorsal lamina X of the lumbar spinal cord |
| SAN | sacral autonomic nucleus |
| OXT | oxytocin |
| SNB | spinal nucleus of the bulbocavernosus |
| OXTR | oxytocin receptor |
| mPOA | medial preoptic area |
| IML | intermediolateral cell column |
| SPFp | subparafascicular thalamic nucleus |
| pERK | phosphorylated extracellular signal-related kinases 1 and 2 |
| ANOVA | analysis of variance |
| YFP | yellow fluorescent protein |
| Tg | transgenic |
| + | positive |
| qPCR | quantitative PCR |
| PCA | p-chloroamphetamine |
| SCI | spinal cord injury |
References
- Truitt, W.A.; Coolen, L.M. Identification of a potential ejaculation generator in the spinal cord. Science 2002, 297, 1566–1569. [Google Scholar] [CrossRef]
- Newton, B.W. Galanin-like immunoreactivity in autonomic regions of the rat lumbosacral spinal cord is sexually dimorphic and varies with the estrous cycle. Brain Res. 1992, 589, 69–83. [Google Scholar] [CrossRef]
- Truitt, W.A.; Shipley, M.T.; Veening, J.G.; Coolen, L.M. Activation of a subset of lumbar spinothalamic neurons after copulatory behavior in male but not female rats. J. Neurosci. 2003, 23, 325–331. [Google Scholar] [CrossRef]
- Ju, G.; Melander, T.; Ceccatelli, S.; Hokfelt, T.; Frey, P. Immunohistochemical evidence for a spinothalamic pathway co-containing cholecystokinin- and galanin-like immunoreactivities in the rat. Neuroscience 1987, 20, 439–456. [Google Scholar] [CrossRef]
- Phan, D.C.; Newton, B.W. Cholecystokinin-8-like immunoreactivity is sexually dimorphic in a midline population of rat lumbar neurons. Neurosci. Lett. 1999, 276, 165–168. [Google Scholar] [CrossRef]
- Nicholas, A.P.; Zhang, X.; Hokfelt, T. An immunohistochemical investigation of the opioid cell column in lamina X of the male rat lumbosacral spinal cord. Neurosci. Lett. 1999, 270, 9–12. [Google Scholar] [CrossRef]
- Sakamoto, H.; Matsuda, K.-I.; Zuloaga, D.G.; Hongu, H.; Wada, E.; Wada, K.; Jordan, C.L.; Breedlove, S.M.; Kawata, M. Sexually dimorphic gastrin releasing peptide system in the spinal cord controls male reproductive functions. Nat. Neurosci. 2008, 11, 634–636. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Saito, K.; Marie-Luce, C.; Raskin, K.; Oti, T.; Satoh, K.; Tamura, K.; Sakamoto, T.; Mhaouty-Kodja, S. Androgen regulates development of the sexually dimorphic gastrin-releasing peptide neuron system in the lumbar spinal cord: Evidence from a mouse line lacking androgen receptor in the nervous system. Neurosci. Lett. 2014, 558, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Oti, T.; Satoh, K.; Saito, K.; Murata, K.; Kawata, M.; Sakamoto, T.; Sakamoto, H. Three-dimensional evaluation of the spinal local neural network revealed by the high-voltage electron microscopy: A double immunohistochemical study. Histochem. Cell Biol. 2012, 138, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Kobayashi, Y.; Hirooka, A.; Takanami, K.; Oti, T.; Jogahara, T.; Oda, S.; Sakamoto, T.; Sakamoto, H. Identification of the sexually dimorphic gastrin-releasing peptide system in the lumbosacral spinal cord that controls male reproductive function in the mouse and Asian house musk shrew (Suncus murinus). J. Comp. Neurol. 2017, 525, 1586–1598. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Oti, T.; Takanami, K.; Satoh, K.; Ueda, Y.; Sakamoto, T.; Sakamoto, H. A sexually dimorphic peptidergic system in the lower spinal cord controlling penile function in non-human primates. Spinal Cord 2017, 56, 57–62. [Google Scholar] [CrossRef]
- Baskerville, T.A.; Allard, J.; Wayman, C.; Douglas, A.J. Dopamine-oxytocin interactions in penile erection. Eur. J. Neurosci. 2009, 30, 2151–2164. [Google Scholar] [CrossRef] [PubMed]
- Veronneau-Longueville, F.; Rampin, O.; Freund-Mercier, M.J.; Tang, Y.; Calas, A.; Marson, L.; McKenna, K.E.; Stoeckel, M.E.; Benoit, G.; Giuliano, F. Oxytocinergic innervation of autonomic nuclei controlling penile erection in the rat. Neuroscience 1999, 93, 1437–1447. [Google Scholar] [CrossRef]
- Oti, T.; Satoh, K.; Uta, D.; Nagafuchi, J.; Tateishi, S.; Ueda, R.; Takanami, K.; Young, L.J.; Galione, A.; Morris, J.F.; et al. Oxytocin Influences Male Sexual Activity via Non-synaptic Axonal Release in the Spinal Cord. Curr. Biol. 2021, 31, 103–114.e5. [Google Scholar] [CrossRef] [PubMed]
- Nutsch, V.L.; Will, R.G.; Hattori, T.; Tobiansky, D.J.; Dominguez, J.M. Sexual experience influences mating-induced activity in nitric oxide synthase-containing neurons in the medial preoptic area. Neurosci. Lett. 2014, 579, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, N.; Chiba, A. Effects of sexual experience on conspecific odor preference and estrous odor-induced activation of the vomeronasal projection pathway and the nucleus accumbens in male rats. Brain Res. 2005, 1066, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Giantonio, G.W.; Lund, N.L.; Gerall, A.A. Effect of diencephalic and rhinencephalic lesions on the male rat’s sexual behavior. J. Comp. Physiol. Psychol. 1970, 73, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y. Lesions of the medial amygdala produce severe impairment of copulatory behavior in sexually inexperienced male rats. Physiol. Behav. 1992, 51, 939–943. [Google Scholar] [CrossRef]
- Jean, A.; Bonnet, P.; Liere, P.; Mhaouty-Kodja, S.; Hardin-Pouzet, H. Revisiting medial preoptic area plasticity induced in male mice by sexual experience. Sci. Rep. 2017, 7, 17846. [Google Scholar] [CrossRef]
- Sala, C.; Segal, M. Dendritic spines: The locus of structural and functional plasticity. Physiol. Rev. 2014, 94, 141–188. [Google Scholar] [CrossRef]
- Gil, M.; Bhatt, R.; Picotte, K.B.; Hull, E.M. Sexual experience increases oxytocin receptor gene expression and protein in the medial preoptic area of the male rat. Psychoneuroendocrinology 2013, 38, 1688–1697. [Google Scholar] [CrossRef]
- Yonezawa, A.; Yoshizumi, M.; Ebiko, M.; Iwanaga, T.; Kimura, Y.; Sakurada, S. Evidence for an involvement of peripheral serotonin in p-chloroamphetamine-induced ejaculation of rats. Pharmacol. Biochem. Behav. 2005, 82, 744–750. [Google Scholar] [CrossRef]
- Sakamoto, H. Sexually dimorphic nuclei in the spinal cord control male sexual functions. Front. Neurosci. 2014, 8, 184. [Google Scholar] [CrossRef]
- Maejima, S.; Abe, Y.; Yamaguchi, S.; Musatov, S.; Ogawa, S.; Kondo, Y.; Tsukahara, S. VGF in the Medial Preoptic Nucleus Increases Sexual Activity Following Sexual Arousal Induction in Male Rats. Endocrinology 2018, 159, 3993–4005. [Google Scholar] [CrossRef]
- Wiggins, J.W.; Kozyrev, N.; Sledd, J.E.; Wilson, G.G.; Coolen, L.M. Chronic Spinal Cord Injury Reduces Gastrin-Releasing Peptide in the Spinal Ejaculation Generator in Male Rats. J. Neurotrauma 2019, 36, 3378–3393. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, J.W.; Sledd, J.E.; Coolen, L.M. Spinal Cord Injury Causes Reduction of Galanin and Gastrin Releasing Peptide mRNA Expression in the Spinal Ejaculation Generator of Male Rats. Front. Neurol. 2021, 12, 982. [Google Scholar] [CrossRef]
- Sachs, B.D.; Garinello, L.D. Hypothetical spinal pacemaker regulating penile reflexes in rats: Evidence from transection of spinal cord and dorsal penile nerves. J. Comp. Physiol. Psychol. 1980, 94, 530–535. [Google Scholar] [CrossRef]
- Kozyrev, N.; Lehman, M.N.; Coolen, L.M. Activation of gastrin-releasing peptide receptors in the lumbosacral spinal cord is required for ejaculation in male rats. J. Sex. Med. 2012, 9, 1303–1318. [Google Scholar] [CrossRef]
- Oti, T.; Takanami, K.; Ito, S.; Ueda, T.; Matsuda, K.I.; Kawata, M.; Soh, J.; Ukimura, O.; Sakamoto, T.; Sakamoto, H. Effects of Sex Steroids on the Spinal Gastrin-Releasing Peptide System Controlling Male Sexual Function in Rats. Endocrinology 2018, 159, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Oti, T.; Takanami, K.; Katayama, N.; Edey, T.; Satoh, K.; Sakamoto, T.; Sakamoto, H. Perinatal testosterone exposure is critical for the development of the male-specific sexually dimorphic gastrin-releasing peptide system in the lumbosacral spinal cord that mediates erection and ejaculation. Biol. Sex Differ. 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Takanami, K.; Sakamoto, H.; Matsuda, K.I.; Satoh, K.; Tanida, T.; Yamada, S.; Inoue, K.; Oti, T.; Sakamoto, T.; Kawata, M. Distribution of gastrin-releasing peptide in the rat trigeminal and spinal somatosensory systems. J. Comp. Neurol. 2014, 522, 1858–1873. [Google Scholar] [CrossRef]
- Katayama, N.; Oti, T.; Takanami, K.; Sakamoto, T.; Sakamoto, H. Postnatal development of the gastrin-releasing peptide system in the lumbosacral spinal cord controlling male reproductive function in rats. Proc. Jpn. Acad. Ser. B 2016, 92, 69–75. [Google Scholar] [CrossRef]
- Sun, Y.G.; Chen, Z.F. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature 2007, 448, 700–703. [Google Scholar] [CrossRef]
- Staudt, M.D.; de Oliveira, C.V.; Lehman, M.N.; McKenna, K.E.; Coolen, L.M. Activation of MAP kinase in lumbar spinothalamic cells is required for ejaculation. J. Sex. Med. 2010, 7, 2445–2457. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K.; Oti, T.; Katoh, A.; Ueta, Y.; Morris, J.F.; Sakamoto, T.; Sakamoto, H. In vivo processing and release into the circulation of GFP fusion protein in arginine vasopressin enhanced GFP transgenic rats: Response to osmotic stimulation. FEBS J. 2015, 282, 2488–2499. [Google Scholar] [CrossRef] [PubMed]
- Castel, M.; Morris, J.F.; Whitnall, M.H.; Sivan, N. Improved visualization of the immunoreactive hypothalamo-neurohypophysial system by use of immuno-gold techniques. Cell Tissue Res. 1986, 243, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ward, A.R.; Morris, J.F. Oestradiol acutely stimulates exocytosis of oxytocin and vasopressin from dendrites and somata of hypothalamic magnocellular neurons. Neuroscience 1995, 68, 1179–1188. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Intromission | Ejaculation | |
|---|---|---|---|
| Naïve | 23.3 ± 3.3 | 27.3 ± 2.2 | 56.1 ± 3.2 |
| Experienced | 32.5 ± 5.0 | 45.3 ± 3.5 | 51.3 ± 3.4 |
| Control | Erection | Ejaculation | |
|---|---|---|---|
| Reflexive erection | 37.8 ± 5.6 | 42.5 ± 7.8 | - |
| PCA-induced ejaculation | 42.1 ± 5.0 | - | 52.6 ± 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oti, T.; Ueda, R.; Kumagai, R.; Nagafuchi, J.; Ito, T.; Sakamoto, T.; Kondo, Y.; Sakamoto, H. Sexual Experience Induces the Expression of Gastrin-Releasing Peptide and Oxytocin Receptors in the Spinal Ejaculation Generator in Rats. Int. J. Mol. Sci. 2021, 22, 10362. https://doi.org/10.3390/ijms221910362
Oti T, Ueda R, Kumagai R, Nagafuchi J, Ito T, Sakamoto T, Kondo Y, Sakamoto H. Sexual Experience Induces the Expression of Gastrin-Releasing Peptide and Oxytocin Receptors in the Spinal Ejaculation Generator in Rats. International Journal of Molecular Sciences. 2021; 22(19):10362. https://doi.org/10.3390/ijms221910362
Chicago/Turabian StyleOti, Takumi, Ryota Ueda, Ryoko Kumagai, Junta Nagafuchi, Takashi Ito, Tatsuya Sakamoto, Yasuhiko Kondo, and Hirotaka Sakamoto. 2021. "Sexual Experience Induces the Expression of Gastrin-Releasing Peptide and Oxytocin Receptors in the Spinal Ejaculation Generator in Rats" International Journal of Molecular Sciences 22, no. 19: 10362. https://doi.org/10.3390/ijms221910362
APA StyleOti, T., Ueda, R., Kumagai, R., Nagafuchi, J., Ito, T., Sakamoto, T., Kondo, Y., & Sakamoto, H. (2021). Sexual Experience Induces the Expression of Gastrin-Releasing Peptide and Oxytocin Receptors in the Spinal Ejaculation Generator in Rats. International Journal of Molecular Sciences, 22(19), 10362. https://doi.org/10.3390/ijms221910362