
The Interaction between microRNAs and the Wnt/β-Catenin Signaling Pathway in Osteoarthritis

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. miRNA Biogenesis and Target Prediction
3. miRNAs Regulation in Osteoarthritis
4. Wnt Pathway Overview
5. Wnt Pathway Regulation in Osteoarthritis (OA)
6. miRNAs Modulate the Wnt Pathway in OA
7. Targeting Wnt Ligands/Receptors and Associated Proteins
8. Targeting β-catenin Destruction Complex and Associated Proteins
9. Targeting Wnt Pathway Transcription Factors and Associated Proteins
10. miRNAs Targeted by Canonical Wnt Pathway in OA
11. miRNA-Based Therapeutics in OA
12. Future Perspective
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| OA | Osteoarthritis |
| MiRNA | Micro ribonucleic acid |
| EVs | Extracellular vesicles |
| MSC | Mesenchymal stem cell |
| BMSC | Bone marrow-derived stem/stromal cell |
| SMSC | Synovial mesenchymal stem cell |
| FLS | Fibroblast-like synoviocyte |
| NSAIDs | non-steroidal anti-inflammatory drugs |
References
- Salmon, J.-H.; Rat, A.-C.; Charlot-Lambrecht, I.; Eschard, J.-P.; Jolly, D.; Fautrel, B. Cost Effectiveness of Intra-Articular Hyaluronic Acid and Disease-Modifying Drugs in Knee Osteoarthritis. Pharmaco. Econ. 2018, 36, 1321–1331. [Google Scholar] [CrossRef]
- Gunaratne, R.; Pratt, D.N.; Banda, J.; Fick, D.P.; Khan, R.J.K.; Robertson, B.W. Patient Dissatisfaction Following Total Knee Arthroplasty: A Systematic Review of the Literature. J. Arthroplast. 2017, 32, 3854–3860. [Google Scholar] [CrossRef]
- Stampella, A.; Monteagudo, S.; Lories, R. Wnt Signaling as Target for the Treatment of Osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2017, 31, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Malemud, C.J. MicroRNAs and Osteoarthritis. Cells 2018, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kowdley, K.V. MicroRNAs in Common Human Diseases. Genom. Proteom. Bioinform. 2012, 10, 246–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaki, S.; Asahara, H. Macro View of MicroRNA Function in Osteoarthritis. Nat. Rev. Rheumatol. 2012, 8, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Endisha, H.; Rockel, J.; Jurisica, I.; Kapoor, M. The Complex Landscape of MicroRNAs in Articular Cartilage: Biology, Pathology, and Therapeutic Targets. JCI Insight 2018, 3, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaki, S.; Sato, T.; Inoue, A.; Otsuki, S.; Ito, Y.; Yokoyama, S.; Kato, Y.; Takemoto, F.; Nakasa, T.; Yamashita, S.; et al. MicroRNA-140 Plays Dual Roles in Both Cartilage Development and Homeostasis. Genes Dev. 2010, 24, 1173–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Q.; Xu, X.; Zhang, Q.; Kang, L.; Xu, Y.; Zhang, K.; Li, L.; Liang, Y.; Hong, T.; Ye, Q.; et al. The IL-1β/AP-1/MiR-30a/ADAMTS-5 Axis Regulates Cartilage Matrix Degradation in Human Osteoarthritis. J. Mol. Med. Berl. Ger. 2016, 94, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.-K.; Meng, F.-G.; Zhang, Z.-Q.; Mao, G.-P.; Huang, Z.-Y.; Liao, W.-M.; He, A.-S. MicroRNA-193b-3p Regulates Matrix Metalloproteinase 19 Expression in Interleukin-1β-Induced Human Chondrocytes. J. Cell. Biochem. 2018, 119, 4775–4782. [Google Scholar] [CrossRef]
- Usui, T.; Okada, M.; Mizuno, W.; Oda, M.; Ide, N.; Morita, T.; Hara, Y.; Yamawaki, H. HDAC4 Mediates Development of Hypertension via Vascular Inflammation in Spontaneous Hypertensive Rats. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, 1894–1904. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Guan, Y.; Tian, S.; Wang, Y.; Sun, K.; Chen, Q. Mechanical and IL-1β Responsive MiR-365 Contributes to Osteoarthritis Development by Targeting Histone Deacetylase 4. Int. J. Mol. Sci. 2016, 17, 436. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Zhao, J.; Jing, W.; Yan, S.; Wang, X.; Xiao, C.; Ma, B. Role of MiR-146a in Human Chondrocyte Apoptosis in Response to Mechanical Pressure Injury in Vitro. Int. J. Mol. Med. 2014, 34, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Tuddenham, L.; Wheeler, G.; Ntounia-Fousara, S.; Waters, J.; Hajihosseini, M.K.; Clark, I.; Dalmay, T. The Cartilage Specific MicroRNA-140 Targets Histone Deacetylase 4 in Mouse Cells. FEBS Lett. 2006, 580, 4214–4217. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.-J.; Yang, X.; Wei, L.; Chen, Q. MiR-365: A Mechanosensitive MicroRNA Stimulates Chondrocyte Differentiation through Targeting Histone Deacetylase 4. FASEB J. 2011, 25, 4457–4466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Jin, E.-H.; Kim, D.; Kim, K.Y.; Chun, C.-H.; Jin, E.-J. MicroRNA-222 Regulates MMP-13 via Targeting HDAC-4 during Osteoarthritis Pathogenesis. BBA Clin. 2015, 3, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, N.; Hossain, U.; Mandal, A.; Sil, P.C. The Wnt Signaling Pathway: A Potential Therapeutic Target against Cancer. Ann. N.Y. Acad. Sci. 2019, 1443, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt Signaling in Cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, X.; Xing, L.; Tian, F. Wnt Signaling: A Promising Target for Osteoarthritis Therapy. Cell Commun. Signal. CCS 2019, 17, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, J.; Jerchow, B.-A.; Würtele, M.; Grimm, J.; Asbrand, C.; Wirtz, R.; Kühl, M.; Wedlich, D.; Birchmeier, W. Functional Interaction of an Axin Homolog, Conductin, with β-Catenin, APC, and GSK3β. Science 1998, 280, 596–599. [Google Scholar] [CrossRef]
- Azzolin, L.; Panciera, T.; Soligo, S.; Enzo, E.; Bicciato, S.; Dupont, S.; Bresolin, S.; Frasson, C.; Basso, G.; Guzzardo, V.; et al. YAP/TAZ Incorporation in the β-Catenin Destruction Complex Orchestrates the Wnt Response. Cell 2014, 158, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-S.; Zhou, P.; Zhang, Y. Abnormal Expression of Key Genes and Proteins in the Canonical Wnt/β-Catenin Pathway of Articular Cartilage in a Rat Model of Exercise-Induced Osteoarthritis. Mol. Med. Rep. 2016, 13, 1999–2006. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, C.; Mendoza-Topaz, C.; Mieszczanek, J.; Bienz, M. Stability Elements in the LRP6 Cytoplasmic Tail Confer Efficient Signalling upon DIX-Dependent Polymerization. J. Cell Sci. 2010, 123, 1588–1599. [Google Scholar] [CrossRef] [Green Version]
- Kahn, M. Can We Safely Target the WNT Pathway? Nat. Rev. Drug Discov. 2014, 13, 513–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruciat, C.-M.; Niehrs, C. Secreted and Transmembrane Wnt Inhibitors and Activators. Cold Spring Harb. Perspect. Biol. 2013, 5, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Funck-Brentano, T.; Bouaziz, W.; Marty, C.; Geoffroy, V.; Hay, E.; Cohen-Solal, M. Dkk-1-Mediated Inhibition of Wnt Signaling in Bone Ameliorates Osteoarthritis in Mice. Arthr. Rheumatol. 2014, 66, 3028–3039. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Niehrs, C. Kremen2 Modulates Dickkopf2 Activity during Wnt/LRP6 Signaling. Gene 2003, 302, 179–183. [Google Scholar] [CrossRef]
- Loughlin, J.; Dowling, B.; Chapman, K.; Marcelline, L.; Mustafa, Z.; Southam, L.; Ferreira, A.; Ciesielski, C.; Carson, D.A.; Corr, M. Functional Variants within the Secreted Frizzled-Related Protein 3 Gene Are Associated with Hip Osteoarthritis in Females. Proc. Natl. Acad. Sci. 2004, 101, 9757–9762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Tang, D.; Wu, Q.; Hao, S.; Chen, M.; Xie, C.; Rosier, R.N.; O′Keefe, R.J.; Zuscik, M.; Chen, D. Activation of β-Catenin Signaling in Articular Chondrocytes Leads to Osteoarthritis-Like Phenotype in Adult β-Catenin Conditional Activation Mice. J. Bone Miner. Res. 2009, 24, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Chen, M.; Zuscik, M.; Wu, Q.; Wang, Y.-J.; Rosier, R.N.; O’Keefe, R.J.; Chen, D. Inhibition of β-Catenin Signaling in Articular Chondrocytes Results in Articular Cartilage Destruction. Arthr. Rheum. 2008, 58, 2053–2064. [Google Scholar] [CrossRef] [Green Version]
- Lietman, C.; Wu, B.; Lechner, S.; Shinar, A.; Sehgal, M.; Rossomacha, E.; Datta, P.; Sharma, A.; Gandhi, R.; Kapoor, M.; et al. Inhibition of Wnt/β-Catenin Signaling Ameliorates Osteoarthritis in a Murine Model of Experimental Osteoarthritis. JCI Insight 2018, 3, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, T.; Kondo, N.; Yasuhara, R.; Shimono, K.; Mackem, S.; Pacifici, M.; Iwamoto, M.; Enomoto-Iwamoto, M. Transient Activation of Wnt/β-Catenin Signaling Induces Abnormal Growth Plate Closure and Articular Cartilage Thickening in Postnatal Mice. Am. J. Pathol. 2009, 175, 1993–2003. [Google Scholar] [CrossRef] [Green Version]
- Nalesso, G.; Thomas, B.L.; Sherwood, J.C.; Yu, J.; Addimanda, O.; Eldridge, S.E.; Thorup, A.-S.; Dale, L.; Schett, G.; Zwerina, J.; et al. WNT16 Antagonises Excessive Canonical WNT Activation and Protects Cartilage in Osteoarthritis. Ann. Rheum. Dis. 2017, 76, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Claudel, M.; Jouzeau, J.-Y.; Cailotto, F. Secreted Frizzled-Related Proteins (SFRPs) in Osteo-Articular Diseases: Much More than Simple Antagonists of Wnt Signaling? FEBS J. 2019, 286, 4832–4851. [Google Scholar] [CrossRef]
- Yazici, Y.; McAlindon, T.E.; Fleischmann, R.; Gibofsky, A.; Lane, N.E.; Kivitz, A.J.; Skrepnik, N.; Armas, E.; Swearingen, C.J.; DiFrancesco, A.; et al. A Novel Wnt Pathway Inhibitor, SM04690, for the Treatment of Moderate to Severe Osteoarthritis of the Knee: Results of a 24-Week, Randomized, Controlled, Phase 1 Study. Osteoarthr. Cartil. 2017, 25, 1598–1606. [Google Scholar] [CrossRef] [Green Version]
- Dell’Accio, F. Pharmacological Blockade of the WNT-Beta-Catenin Signaling: A Possible First-in-Kind DMOAD. Osteoarthr. Cartil. 2018, 3, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Shi, H.; Wang, M.; Liu, J.; Liu, G. MicroRNA-497-5p Attenuates IL-1β-Induced Cartilage Matrix Degradation in Chondrocytes via Wnt/β-Catenin Signal Pathway. Int. J. Clin. Exp. Pathol. 2019, 12, 3108–3118. [Google Scholar] [PubMed]
- Zhang, Y.; Huang, X.; Yuan, Y. MicroRNA-410 Promotes Chondrogenic Differentiation of Human Bone Marrow Mesenchymal Stem Cells through down-Regulating Wnt3a. Am. J. Transl. Res. 2017, 9, 136–145. [Google Scholar] [PubMed]
- Mao, G.; Zhang, Z.; Hu, S.; Zhang, Z.; Chang, Z.; Huang, Z.; Liao, W.; Kang, Y. Exosomes Derived from MiR-92a-3p-Overexpressing Human Mesenchymal Stem Cells Enhance Chondrogenesis and Suppress Cartilage Degradation via Targeting WNT5A. Stem Cell Res. Ther. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Yan, P.; Chen, Y.; Chen, Y.; Yang, J.; Xu, G.; Mao, H.; Qiu, Y. MicroRNA-26b Inhibits Cell Proliferation and Cytokine Secretion in Human RASF Cells via the Wnt/GSK-3β/β-Catenin Pathway. Diagn. Pathol. 2015, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Hu, C.; Han, L.; Liu, L.; Jing, W.; Tang, W.; Tian, W.; Long, J. MiR-154-5p Regulates Osteogenic Differentiation of Adipose-Derived Mesenchymal Stem Cells under Tensile Stress through the Wnt/PCP Pathway by Targeting Wnt11. Bone 2015, 78, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, Y.; Jin, H.; Yu, T. Knockdown of MicroRNA-203 Alleviates LPS-Induced Injury by Targeting MCL-1 in C28/I2 Chondrocytes. Exp. Cell Res. 2017, 359, 171–178. [Google Scholar] [CrossRef]
- Sun, T.; Li, X.; Song, H.; Gao, F.; Zhou, G.; Li, X.; Chen, Z.; Chen, L. MiR-146a Aggravates LPS-Induced Inflammatory Injury by Targeting CXCR4 in the Articular Chondrocytes. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 44, 1282–1294. [Google Scholar] [CrossRef] [PubMed]
- Le, L.T.T.; Swingler, T.E.; Crowe, N.; Vincent, T.L.; Barter, M.J.; Donell, S.T.; Delany, A.M.; Dalmay, T.; Young, D.A.; Clark, I.M. The MicroRNA-29 Family in Cartilage Homeostasis and Osteoarthritis. J. Mol. Med. 2016, 94, 583–596. [Google Scholar] [CrossRef] [Green Version]
- Xing, D.; Wang, B.; Ke, Y.; Lin, J. Overexpression of Microrna-1 Controls the Development of Osteoarthritis via Targeting FZD7 of WNT/β-Catenin Signaling. Osteoarthr. Cartil. 2017, 25, 1–19. [Google Scholar] [CrossRef]
- Zhang, J.; Tu, Q.; Bonewald, L.F.; He, X.; Stein, G.; Lian, J.; Chen, J. Effects of MiR-335-5p in Modulating Osteogenic Differentiation by Specifically Downregulating Wnt Antagonist DKK1. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2011, 26, 1953–1963. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Liang, D.; Li, B.; Liang, C.; Huang, W.; Lin, H. MicroRNA-320a Protects against Osteoarthritis Cartilage Degeneration by Regulating the Expressions of BMI-1 and RUNX2 in Chondrocytes. Pharmacology 2017, 72, 223–226. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Y.; Liang, C.; Chen, L.; Zhang, G.; Qian, A. Mechanosensitive MiRNAs and Bone Formation. Int. J. Mol. Sci. 2017, 18, 1684. [Google Scholar] [CrossRef] [Green Version]
- Chao, J.-R.; Wang, J.-M.; Lee, S.-F.; Peng, H.-W.; Lin, Y.-H.; Chou, C.-H.; Li, J.-C.; Huang, H.-M.; Chou, C.-K.; Kuo, M.-L.; et al. Mcl-1 Is an Immediate-Early Gene Activated by the Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) Signaling Pathway and Is One Component of the GM-CSF Viability Response. Mol. Cell. Biol. 1998, 18, 4883–4898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dkk-1–Mediated Inhibition of Wnt Signaling in Bone Ameliorates Osteoarthritis in Mice-Funck-Brentano-2014-Arthritis & Rheumatology-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/art.38799 (accessed on 11 December 2019).
- Cho, J.-H.; Dimri, M.; Dimri, G.P. A Positive Feedback Loop Regulates the Expression of Polycomb Group Protein BMI1 via WNT Signaling Pathway. J. Biol. Chem. 2013, 288, 3406–3418. [Google Scholar] [CrossRef] [Green Version]
- Ukai, T.; Sato, M.; Akutsu, H.; Umezawa, A.; Mochida, J. MicroRNA-199a-3p, MicroRNA-193b, and MicroRNA-320c Are Correlated to Aging and Regulate Human Cartilage Metabolism. J. Orthop. Res. 2012, 30, 1915–1922. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Zhao, Q.; Wang, J.; He, X. MicroRNA-10a Influences Osteoblast Differentiation and Angiogenesis by Regulating β-Catenin Expression. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 37, 2194–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, Y.; Zhang, H.; Gao, W.; Lu, M.; Liu, W.; Li, Y.; Yin, Z. Forkhead Box C1 Promotes the Pathology of Osteoarthritis by Upregulating β-Catenin in Synovial Fibroblasts. FEBS J. 2019, 287, 3065–3087. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Ye, Y.; Zhang, W.; Wang, J.; Chen, A.; Guo, F. MiR-142-3p Promotes Osteoblast Differentiation by Modulating Wnt Signaling. Mol. Med. Rep. 2013, 7, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Zhao, C.; Zhang, P.; Liu, Y.; Jiang, Y.; Wu, E.; Xue, H.; Liu, C.; Li, Z. MiR-26b Modulates OA Induced BMSC Osteogenesis through Regulating GSK3β/β-Catenin Pathway. Exp. Mol. Pathol. 2019, 107, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Long, J.; Guo, W.; Ye, W. MicroRNA-195-5p Inhibitor Prevents the Development of Osteoarthritis by Targeting REGγ. Mol. Med. Rep. 2019, 19, 4561–4568. [Google Scholar] [CrossRef]
- Wang, F.L.; Li, D.D.; Huang, X.; Liu, F.; Liu, Q.B.; Lan, J. miR-29c-3p targeted dishevelled 2 on osteogenesis differentiation of rat bone marrow mesenchymal stem cells in high-fat environment. Zhonghua Kou Qiang Yi Xue Za Zhi Zhonghua Kouqiang Yixue Zazhi Chin. J. Stomatol. 2018, 53, 694–700. [Google Scholar] [CrossRef]
- Zhang, H.-X.; Sun, C.; Yu, H.-C.; Song, B.; Pan, Z.-X. Targeted Inhibition of β-Catenin by MiR-320 and Decreased MMP-13 Expression in Suppressing Chondrocyte Collagen Degradation. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5828–5835. [Google Scholar] [CrossRef] [PubMed]
- Konsavage, W.M.; Kyler, S.L.; Rennoll, S.A.; Jin, G.; Yochum, G.S. Wnt/β-Catenin Signaling Regulates Yes-Associated Protein (YAP) Gene Expression in Colorectal Carcinoma Cells. J. Biol. Chem. 2012, 287, 11730–11739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Gao, X.; Yu, T.; Yuan, L.; Dai, J.; Wang, W.; Chen, G.; Jiao, C.; Zhou, W.; Huang, Q.; et al. REGγ Controls Hippo Signaling and Reciprocal NF-ΚB–YAP Regulation to Promote Colon Cancer. Clin. Cancer Res. 2018, 24, 2015–2025. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Castro-Piedras, I.; Simmons, G.E.; Pruitt, K. Dishevelled: A Masterful Conductor of Complex Wnt Signals. Cell. Signal. 2018, 47, 52–64. [Google Scholar] [CrossRef]
- Paik, S.; Jung, H.S.; Lee, S.; Yoon, D.S.; Park, M.S.; Lee, J.W. MiR-449a Regulates the Chondrogenesis of Human Mesenchymal Stem Cells Through Direct Targeting of Lymphoid Enhancer-Binding Factor-1. Stem Cells Dev. 2012, 21, 3298–3308. [Google Scholar] [CrossRef] [Green Version]
- Baek, D.; Lee, K.-M.; Park, K.W.; Suh, J.W.; Choi, S.M.; Park, K.H.; Lee, J.W.; Kim, S.-H. Inhibition of MiR-449a Promotes Cartilage Regeneration and Prevents Progression of Osteoarthritis in In Vivo Rat Models. Mol. Ther. Nucleic Acids 2018, 13, 322–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Chen, Z.; Shi, W.; Zhang, R.; Li, L.; Liu, H.; Wu, L. TMF Inhibits MiR-29a/Wnt/β-Catenin Signaling through Upregulating Foxo3a Activity in Osteoarthritis Chondrocytes. Drug Des. Devel. Ther. 2019, 13, 2009–2019. [Google Scholar] [CrossRef] [Green Version]
- Mao, G.; Zhang, Z.; Huang, Z.; Chen, W.; Huang, G.; Meng, F.; Zhang, Z.; Kang, Y. MicroRNA-92a-3p Regulates the Expression of Cartilage-Specific Genes by Directly Targeting Histone Deacetylase 2 in Chondrogenesis and Degradation. Osteoarthr. Cartil. 2017, 25, 521–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, F.; Li, Z.; Zhang, Z.; Yang, Z.; Kang, Y.; Zhao, X.; Long, D.; Hu, S.; Gu, M.; He, S.; et al. MicroRNA-193b-3p Regulates Chondrogenesis and Chondrocyte Metabolism by Targeting HDAC3. Theranostics 2018, 8, 2862–2883. [Google Scholar] [CrossRef]
- Xu, W.; Gao, P.; Zhang, Y.; Piao, L.; Dong, D. MicroRNA-138 Induces Cell Survival and Reduces WNT/β-Catenin Signaling of Osteoarthritis Chondrocytes through NEK2. IUBMB Life 2019, 71, 1355–1366. [Google Scholar] [CrossRef]
- Gao, F.; Peng, C.; Zheng, C.; Zhang, S.; Wu, M. MiRNA-101 Promotes Chondrogenic Differentiation in Rat Bone Marrow Mesenchymal Stem Cells. Exp. Ther. Med. 2019, 17, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.; Zhang, X.; Hu, X.; Zhou, C.; Ao, Y. Silencing of MicroRNA-101 Prevents IL-1β-Induced Extracellular Matrix Degradation in Chondrocytes. Arthr. Res. Ther. 2012, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Sanchez, A.; Dudek, K.A.; Murphy, C.L. Regulation of Human Chondrocyte Function through Direct Inhibition of Cartilage Master Regulator SOX9 by MicroRNA-145 (MiRNA-145). J. Biol. Chem. 2012, 287, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.-X.; Tian, Z.-G.; Zhu, L.-F.; Wu, W.-D.; Zhou, S.-L.; Zhao, Y.-T.; Huang, S. MicroRNA-615-3p Promotes the Osteoarthritis Progression by Inhibiting Chondrogenic Differentiation of Bone Marrow Mesenchymal Stem Cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6212–6220. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.-C.; Yuan, T.; Zhang, Y.-L.; Yin, W.-J.; Guo, S.-C.; Zhang, C.-Q. Exosomes Derived from MiR-140-5p-Overexpressing Human Synovial Mesenchymal Stem Cells Enhance Cartilage Tissue Regeneration and Prevent Osteoarthritis of the Knee in a Rat Model. Theranostics 2017, 7, 180–195. [Google Scholar] [CrossRef] [PubMed]
- Götze, S.; Coersmeyer, M.; Müller, O.; Sievers, S. Histone Deacetylase Inhibitors Induce Attenuation of Wnt Signaling and TCF7L2 Depletion in Colorectal Carcinoma Cells. Int. J. Oncol. 2014, 45, 1715–1723. [Google Scholar] [CrossRef]
- Chen, W.-P.; Bao, J.-P.; Tang, J.-L.; Hu, P.-F.; Wu, L.-D. Trichostatin A Inhibits Expression of Cathepsins in Experimental Osteoarthritis. Rheumatol. Int. 2011, 31, 1325–1331. [Google Scholar] [CrossRef]
- Ota, S.; Ishitani, S.; Shimizu, N.; Matsumoto, K.; Itoh, M.; Ishitani, T. NLK Positively Regulates Wnt/β-Catenin Signalling by Phosphorylating LEF1 in Neural Progenitor Cells. EMBO J. 2012, 31, 1904–1915. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cornelis, F.M.F.; Lories, R.J.; Monteagudo, S. Exostosin-1 Enhances Canonical Wnt Signaling Activity during Chondrogenic Differentiation. Osteoarthr. Cartil. 2019, 27, 1702–1710. [Google Scholar] [CrossRef] [PubMed]
- Topol, L.; Chen, W.; Song, H.; Day, T.F.; Yang, Y. Sox9 Inhibits Wnt Signaling by Promoting Beta-Catenin Phosphorylation in the Nucleus. J. Biol. Chem. 2009, 284, 3323–3333. [Google Scholar] [CrossRef] [Green Version]
- Cheleschi, S.; De Palma, A.; Pecorelli, A.; Pascarelli, N.; Valacchi, G.; Belmonte, G.; Carta, S.; Galeazzi, M.; Fioravanti, A. Hydrostatic Pressure Regulates MicroRNA Expression Levels in Osteoarthritic Chondrocyte Cultures via the Wnt/β-Catenin Pathway. Int. J. Mol. Sci. 2017, 18, 133. [Google Scholar] [CrossRef]
- Wang, J.H.; Shih, K.S.; Wu, Y.W.; Wang, A.W.; Yang, C.R. Histone Deacetylase Inhibitors Increase MicroRNA-146a Expression and Enhance Negative Regulation of Interleukin-1β Signaling in Osteoarthritis Fibroblast-like Synoviocytes. Osteoarthr. Cartil. 2013, 21, 1987–1996. [Google Scholar] [CrossRef] [Green Version]
- Kapinas, K.; Kessler, C.; Ricks, T.; Gronowicz, G.; Delany, A.M. MiR-29 Modulates Wnt Signaling in Human Osteoblasts through a Positive Feedback Loop. J. Biol. Chem. 2010, 285, 25221–25231. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Qin, S.; Yi, C.; Ma, G.; Zhu, H.; Zhou, W.; Xiong, Y.; Zhu, X.; Wang, Y.; He, L.; et al. MiR-140 Is Co-Expressed with Wwp2-C Transcript and Activated by Sox9 to Target Sp1 in Maintaining the Chondrocyte Proliferation. FEBS Lett. 2011, 585, 2992–2997. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Miao, Y.; Cao, Z.; Shi, J.; Li, J.; Kang, F.; Dou, C.; Xie, Z.; Xiang, Q.; Dong, S. MicroRNA-29b Regulates Hypertrophy of Murine Mesenchymal Stem Cells Induced toward Chondrogenesis. J. Cell. Biochem. 2019, 120, 8742–8753. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Li, Y.; Chopp, M. Exosomes/MiRNAs as Mediating Cell-Based Therapy of Stroke. Front. Cell. Neurosci. 2014, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lin, J.; Zhou, X.; Chen, X.; Chen, A.C.; Pi, B.; Pan, G.; Pei, M.; Yang, H.; Liu, T.; et al. Melatonin Prevents Osteoarthritis-Induced Cartilage Degradation via Targeting MicroRNA-140. Oxid. Med. Cell. Longev. 2019, 2019, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, N.; Janicot, M.; Perera, T. The Wnt-Dependent Signaling Pathways as Target in Oncology Drug Discovery. Invest. New Drugs 2006, 24, 263–280. [Google Scholar] [CrossRef] [Green Version]
- De Ferrari, G.V.; Inestrosa, N.C. Wnt Signaling Function in Alzheimer′s Disease. Brain Res. Brain Res. Rev. 2000, 33, 1–12. [Google Scholar] [CrossRef]
- Schiavone, M.L.; Millucci, L.; Bernardini, G.; Giustarini, D.; Rossi, R.; Marzocchi, B.; Santucci, A. Homogentisic Acid Affects Human Osteoblastic Functionality by Oxidative Stress and Alteration of the Wnt/β-Catenin Signaling Pathway. J. Cell. Physiol. 2020, 235, 6808–6816. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, V.; O′Green, A.L.; Bossard, C.; Seo, T.; Lamangan, L.; Ibanez, M.; Ghias, A.; Lai, C.; Do, L.; Cho, S.; et al. Modulation of the Wnt Pathway through Inhibition of CLK2 and DYRK1A by Lorecivivint as a Novel, Potentially Disease-Modifying Approach for Knee Osteoarthritis Treatment. Osteoarthr. Cartil. 2019, 27, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.; Liang, Y.; Xu, X.; Xiao, Y.; Wang, D. Recent Progress on the Role of MiR-140 in Cartilage Matrix Remodelling and Its Implications for Osteoarthritis Treatment. Arthr. Res. Ther. 2020, 22, 194. [Google Scholar] [CrossRef]
- Hecht, N.; Johnstone, B.; Angele, P.; Walker, T.; Richter, W. Mechanosensitive MiRs Regulated by Anabolic and Catabolic Loading of Human Cartilage. Osteoarthr. Cartil. 2019, 27, 1208–1218. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Z.; Liu, Z.; Shi, S.; Zhang, Z.; Zhang, J.; Lin, H. MiR-221/222 Activate the Wnt/β-Catenin Signaling to Promote Triple-Negative Breast Cancer. J. Mol. Cell Biol. 2018, 10, 302–315. [Google Scholar] [CrossRef] [Green Version]
- Garzon, R.; Marcucci, G.; Croce, C.M. Targeting MicroRNAs in Cancer: Rationale, Strategies and Challenges. Nat. Rev. Drug Discov. 2010, 9, 775–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sondag, G.R.; Haqqi, T.M. The Role of MicroRNAs and Their Targets in Osteoarthritis. Curr. Rheumatol. Rep. 2016, 18, 1–23. [Google Scholar] [CrossRef]
- Mathiessen, A.; Conaghan, P.G. Synovitis in Osteoarthritis: Current Understanding with Therapeutic Implications. Arthr. Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usami, Y.; Gunawardena, A.T.; Iwamoto, M.; Enomoto-Iwamoto, M. Wnt Signaling in Cartilage Development and Diseases: Lessons from Animal Studies. Lab. Invest. 2016, 96, 186–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Feng, Y.; Zhang, R.; Zhang, W.; Shu, Y.; Zeng, Z.; Huang, S.; Zhang, L.; Huang, B.; Wu, D.; et al. A Simplified System for the Effective Expression and Delivery of Functional Mature MicroRNAs in Mammalian Cells. Cancer Gene Ther. 2020, 27, 424–437. [Google Scholar] [CrossRef]
- Jin, H.Y.; Gonzalez-Martin, A.; Miletic, A.V.; Lai, M.; Knight, S.; Sabouri-Ghomi, M.; Head, S.R.; Macauley, M.S.; Rickert, R.C.; Xiao, C. Transfection of MicroRNA Mimics Should Be Used with Caution. Front. Genet. 2015, 6, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taheri, S.; Yoshida, T.; Böker, K.O.; Foerster, R.H.; Jochim, L.; Flux, A.L.; Grosskopf, B.; Lehmann, W.; Schilling, A.F. Investigating the Microchannel Architectures Inside the Subchondral Bone in Relation to Estimated Hip Reaction Forces on the Human Femoral Head. Calcif. Tissue Int. 2021, 1, 1–15. [Google Scholar] [CrossRef]
- Taheri, S.; Winkler, T.; Schenk, L.S.; Neuerburg, C.; Baumbach, S.F.; Zustin, J.; Lehmann, W.; Schilling, A.F. Developmental Transformation and Reduction of Connective Cavities within the Subchondral Bone. Int. J. Mol. Sci. 2019, 20, 770. [Google Scholar] [CrossRef] [Green Version]
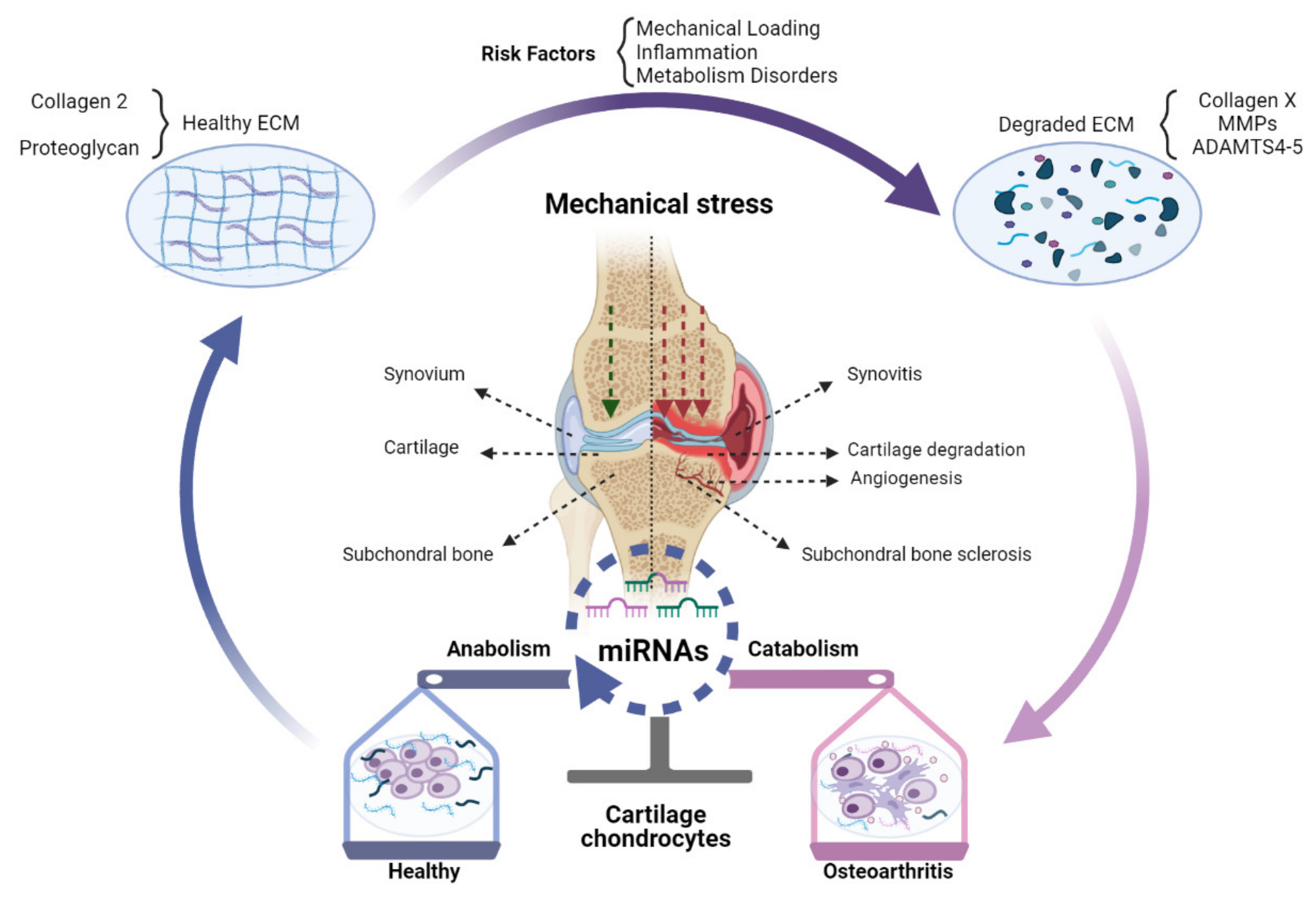

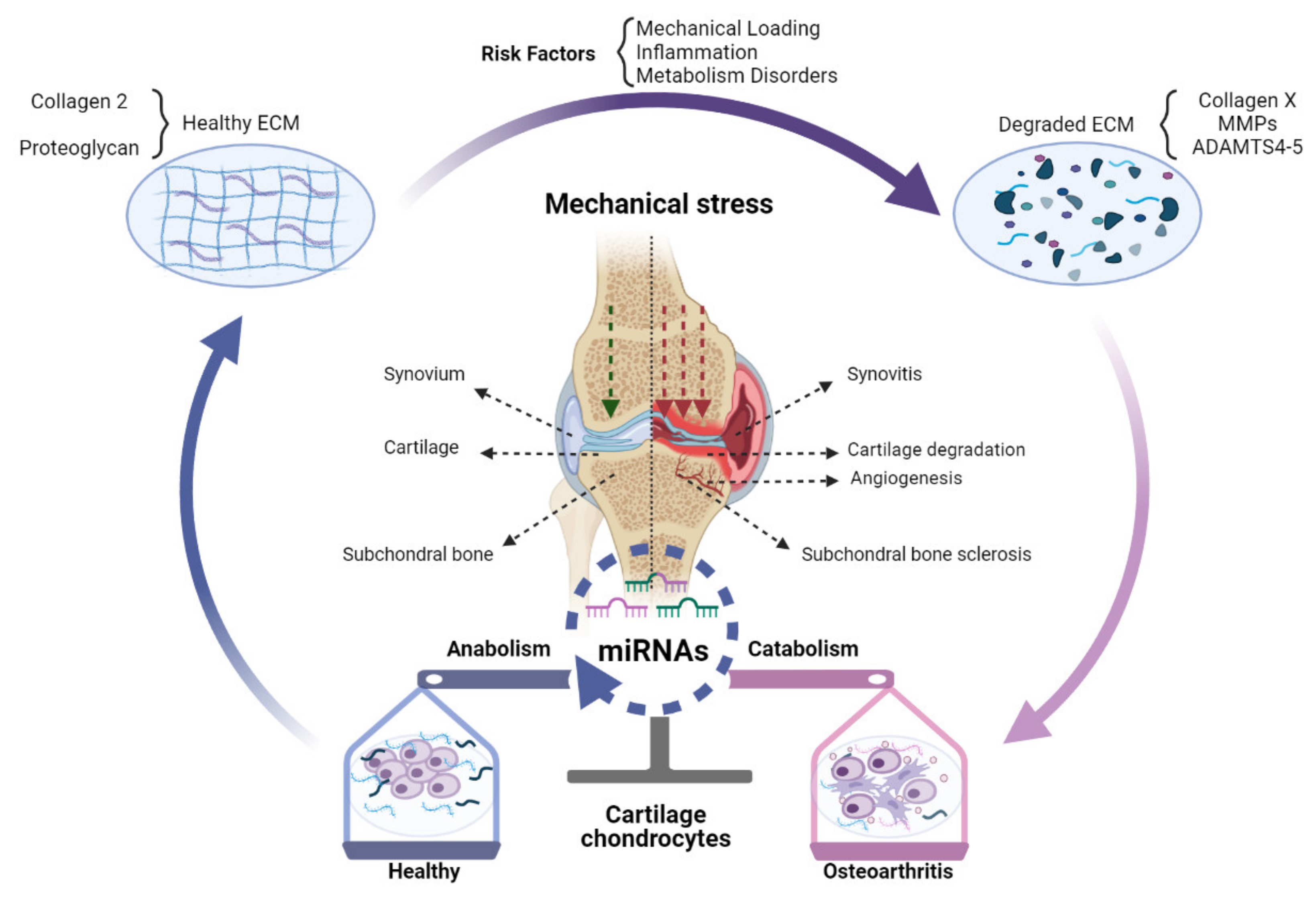{kind=link}
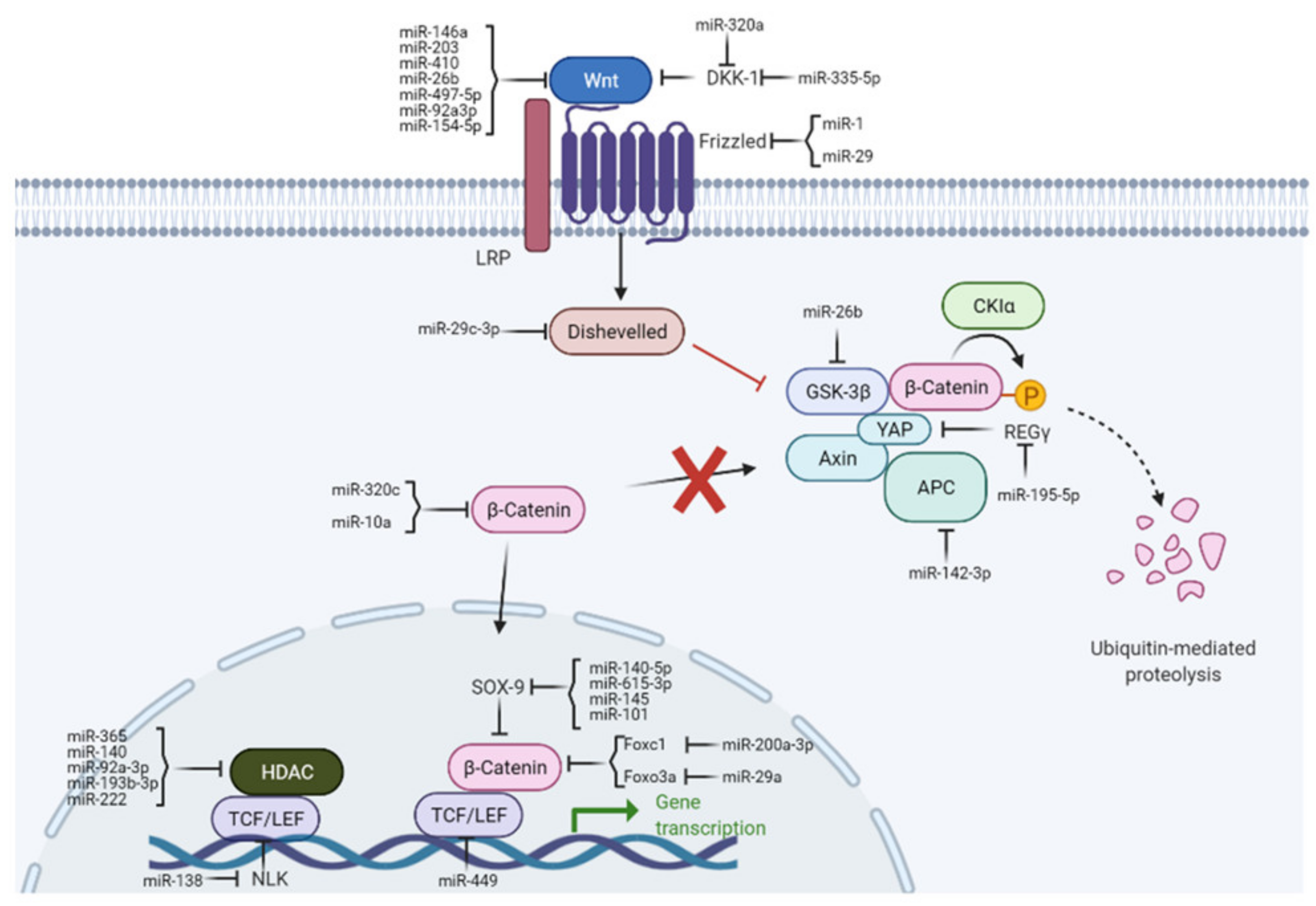
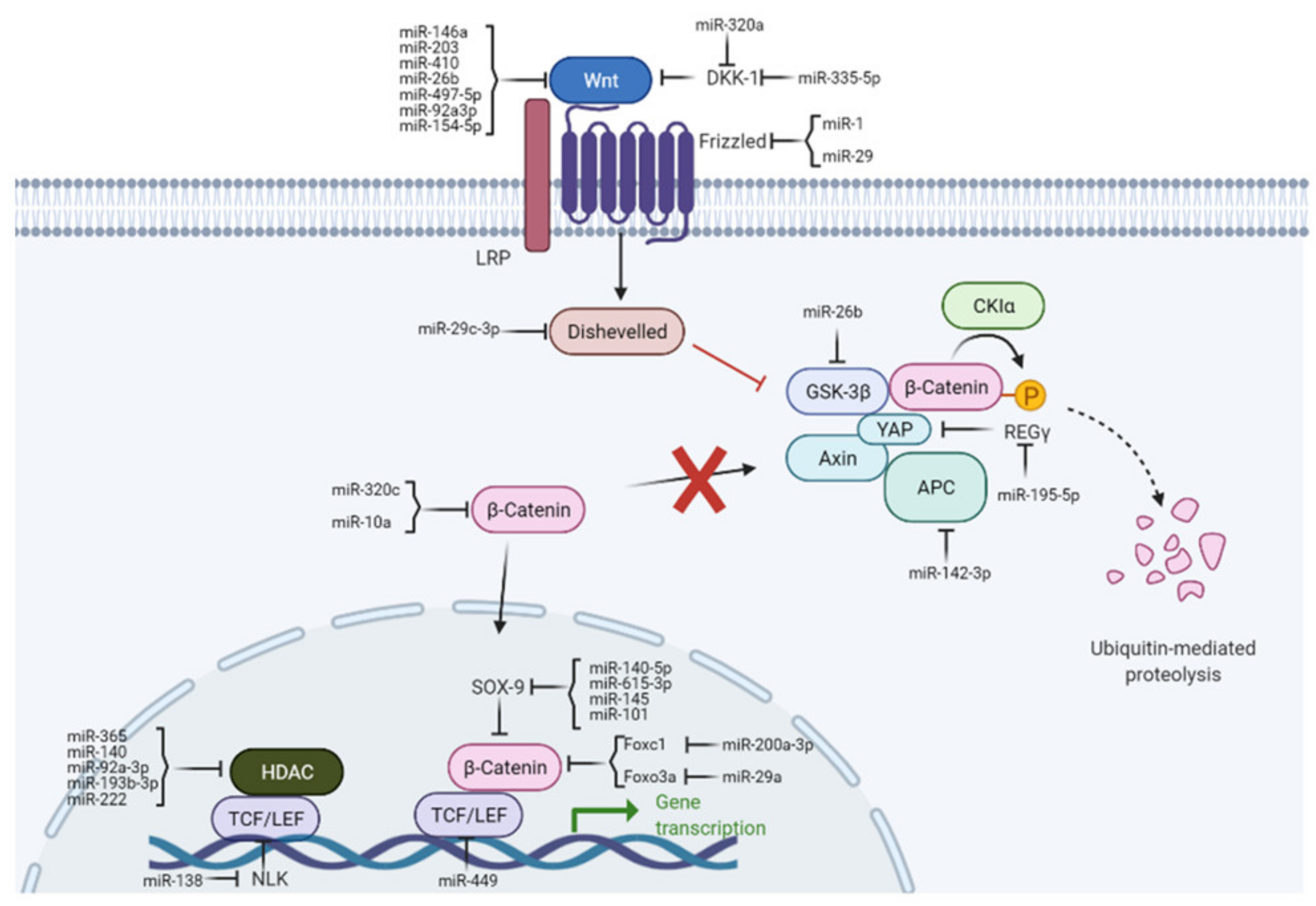{kind=link}
| miRNA | Target | Effect of miRNA | Reference |
|---|---|---|---|
| miR-497-5p | Wnt3a | Inhibits cartilage matrix degradation | Hou et al., (2019) [38] |
| miR-410 | Wnt3a | Promotes chondrogenic differentiation | Y. Zhang et al., (2017) [39] |
| miR-92a-3p | Wnt5a | Inhibits cartilage matrix degradation | Mao et al., (2018) [40] |
| miR-26b | Wnt | Inhibits chondrogenic differentiation | T. Huang et al., (2019) [41] |
| miR-154-5p | Wnt11 | Inhibits osteogenic differentiation | Li et al., (2015) [42] |
| miR-203 | Mcl-1 | Promotes inflammation | Zhao et al., (2017) [43] |
| miR-146a | CXCR4 | Promotes inflammation | Sun et al., (2017) [44] |
| miR-29 | FZD3, FZD5, DVL3, FRAT2, and CK2A2 | Complex role in cartilage homeostasis | Le et al., (2016) [45] |
| miR-1 | FZD7 | Inhibits cartilage matrix degradation | Xing et al., (2017) [46] |
| miR-335-5p | DKK1 | Promotes osteogenic differentiation | J. Zhang et al., (2011) [47] |
| miR-320a | BMI-1 | Inhibits cartilage matrix degradation | Peng et al., (2017) [48] |
| miRNA | Target | Effect of miRNA | Reference |
|---|---|---|---|
| miR-320c | β-catenin | Inhibits cartilage matrix degradation | Zhang et al., (2019) [53] |
| miR-10a | β-catenin | Inhibits osteogenic differentiation | Li et al., (2015) [54] |
| miR-200a-3p | FoxC1 | Inhibits cartilage matrix degradation | Wang et al., (2019) [55] |
| miR-142-3p | APC | Promotes osteogenic differentiation | Hu et al., (2013) [56] |
| miR-26b | GSK3β | Promotes osteogenic differentiation | Hu et al., (2019) [57] |
| miR-195-5p | YAP | Promotes chondrocytes apoptosis | Shu et al., (2019) [58] |
| miR-29c-3p | DVL2 | Inhibits osteogenic differentiation | Wang et al., (2018) [59] |
| miRNA | Target | Effect of miRNA | Reference |
|---|---|---|---|
| miR-449a | LEF-1 | Inhibits chondrogenesis | Paik et al., (2012) [64] Baek et al., (2018) [65] |
| miR-29a | Foxo3a | Promotes chondrocyte apoptosis | Huang et al., (2019) [66] |
| miR-365 | HDAC-4 | Promote chondrogenesis | Guan et al., (2011) [16] |
| miR-222 | HDAC-4 | Promote chondrogenesis | Song et al., (2015) [17] |
| miR-92a-3p | HDAC-2 | Promote chondrogenesis | Mao et al., (2017) [67] |
| miR-193b-3p | HDAC-3 | Promote chondrogenesis | Meng et al., (2018) [68] |
| miR-140 | HDAC-4 | Promote chondrogenesis | Tuddenham et al., (2006) [15] |
| miR-138 | NLK | Promotes chondrogenesis | Xu et al., (2019) [69] |
| miR-101 | Sox9 | Inhibits chondrogenesis | Gao et al., (2019) [70] Dai et al., (2012) [71] |
| miR-145 | Sox9 | Inhibits chondrogenesis | Sanchez et al., (2012) [72] |
| miR-615-3p | Sox9 | Inhibits chondrogenesis | Zhou et al., (2018) [73] |
| miR-140-5p | RaIA | Promotes chondrogenesis | Tao et al., (2017) [74] |
| Molecules | miRNA | Effect of miRNA | Reference |
|---|---|---|---|
| Wnt/β-catenin | miR-27a/b, miR-140, miR-146a/b miR-365 | Promotes cartilage matrix degradation | Cheleschi et al., (2017) [80] |
| HDAC inhibitor | miR-146a | Inhibits inflammation | Wang et al., (2013) [81] |
| β-catenin/TCF4/LEF1 | miR-29 | Promotes chondrocyte apoptosis | Kapinas et al., (2019) [82] |
| Wnt/β-catenin | miR-140 | Promotes chondrogenesis | Yang et al., (2011) [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shang, X.; Böker, K.O.; Taheri, S.; Hawellek, T.; Lehmann, W.; Schilling, A.F. The Interaction between microRNAs and the Wnt/β-Catenin Signaling Pathway in Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 9887. https://doi.org/10.3390/ijms22189887
Shang X, Böker KO, Taheri S, Hawellek T, Lehmann W, Schilling AF. The Interaction between microRNAs and the Wnt/β-Catenin Signaling Pathway in Osteoarthritis. International Journal of Molecular Sciences. 2021; 22(18):9887. https://doi.org/10.3390/ijms22189887
Chicago/Turabian StyleShang, Xiaobin, Kai Oliver Böker, Shahed Taheri, Thelonius Hawellek, Wolfgang Lehmann, and Arndt F. Schilling. 2021. "The Interaction between microRNAs and the Wnt/β-Catenin Signaling Pathway in Osteoarthritis" International Journal of Molecular Sciences 22, no. 18: 9887. https://doi.org/10.3390/ijms22189887
APA StyleShang, X., Böker, K. O., Taheri, S., Hawellek, T., Lehmann, W., & Schilling, A. F. (2021). The Interaction between microRNAs and the Wnt/β-Catenin Signaling Pathway in Osteoarthritis. International Journal of Molecular Sciences, 22(18), 9887. https://doi.org/10.3390/ijms22189887