
Renal Ischemia/Reperfusion Early Induces Myostatin and PCSK9 Expression in Rat Kidneys and HK-2 Cells

 , ,
, ,
Abstract
1. Introduction
2. Results
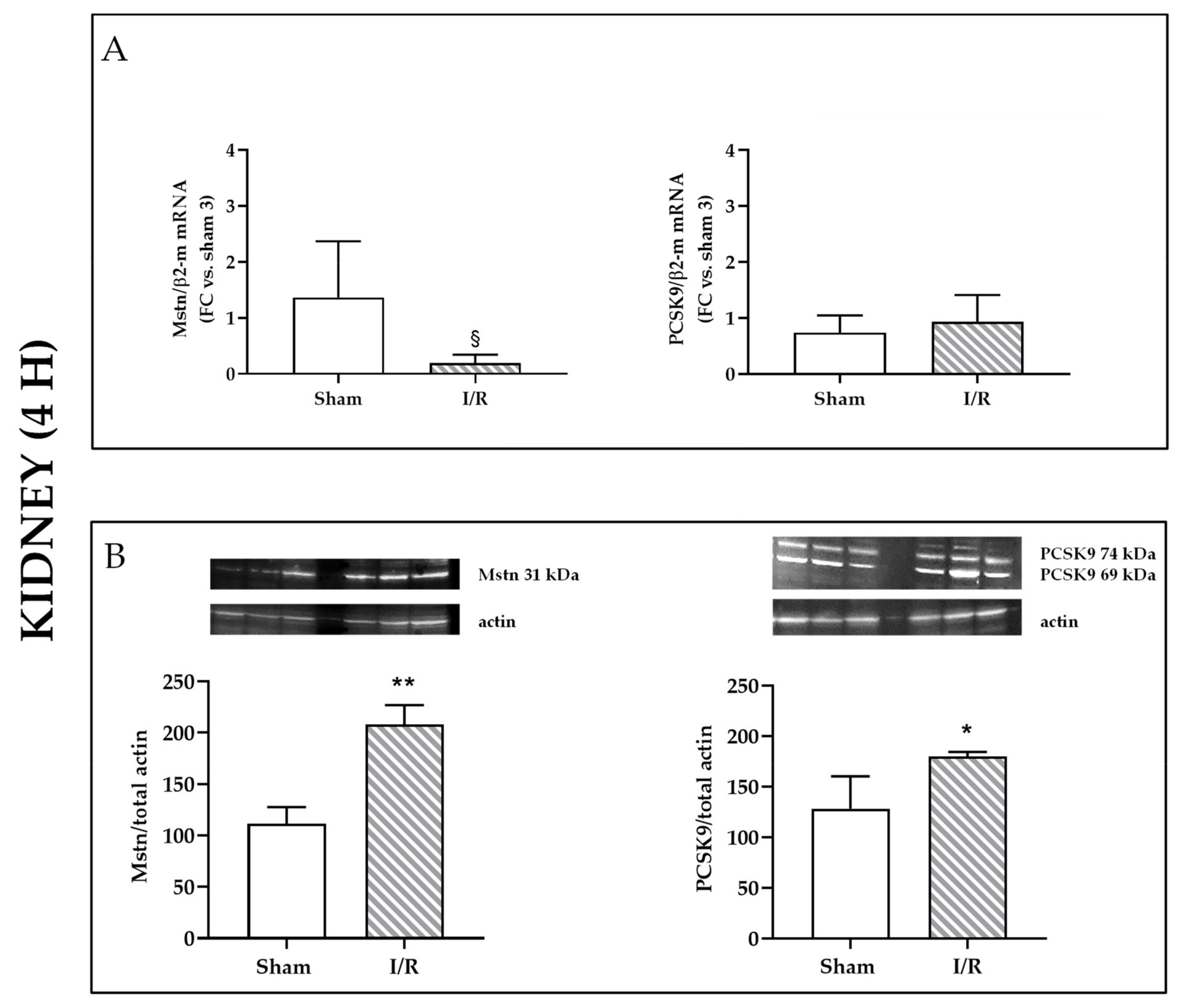
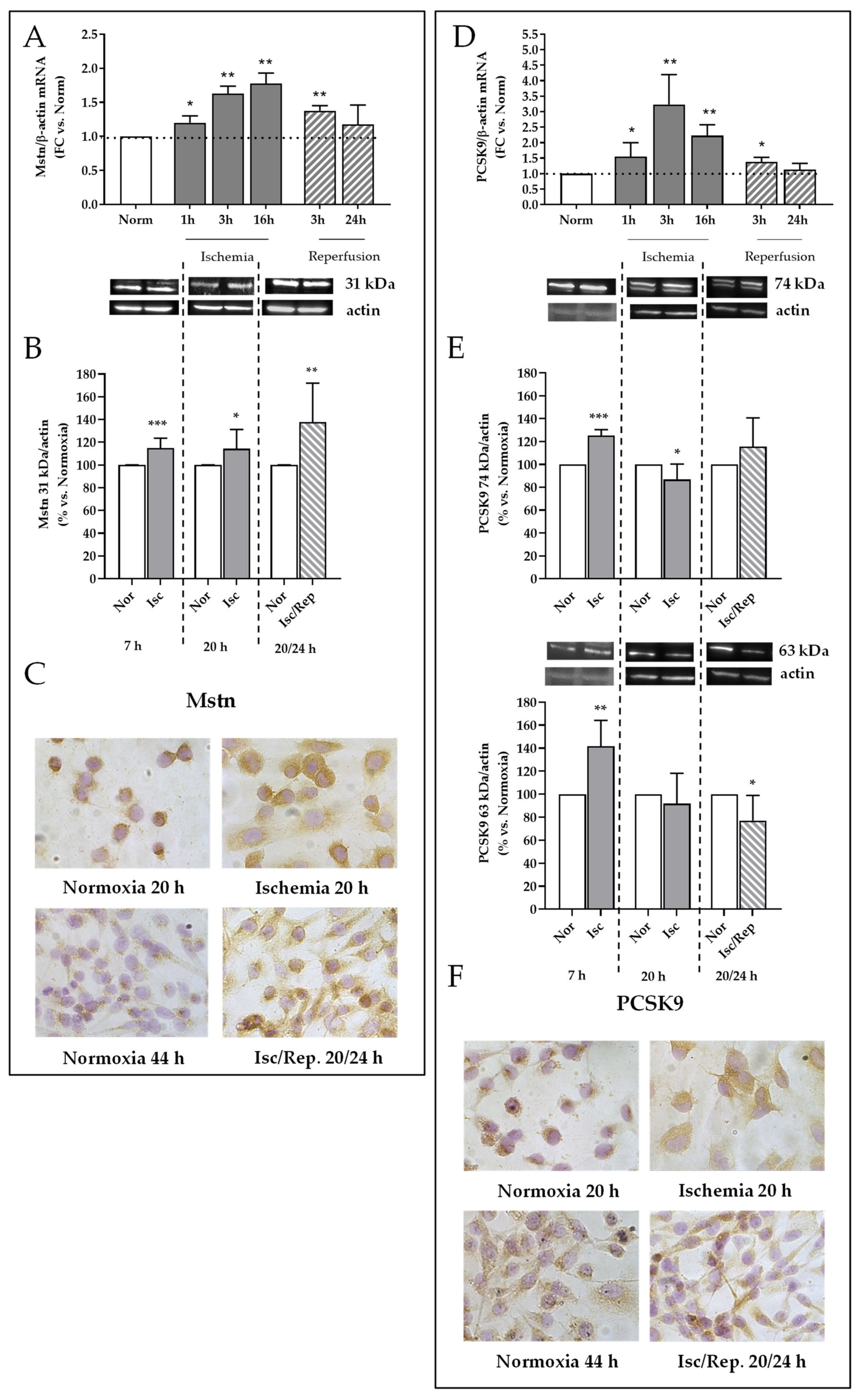
2.1. I/R Induces Mstn and PCSK9 Protein Expression in Rat Kidneys 4 h after Reflow
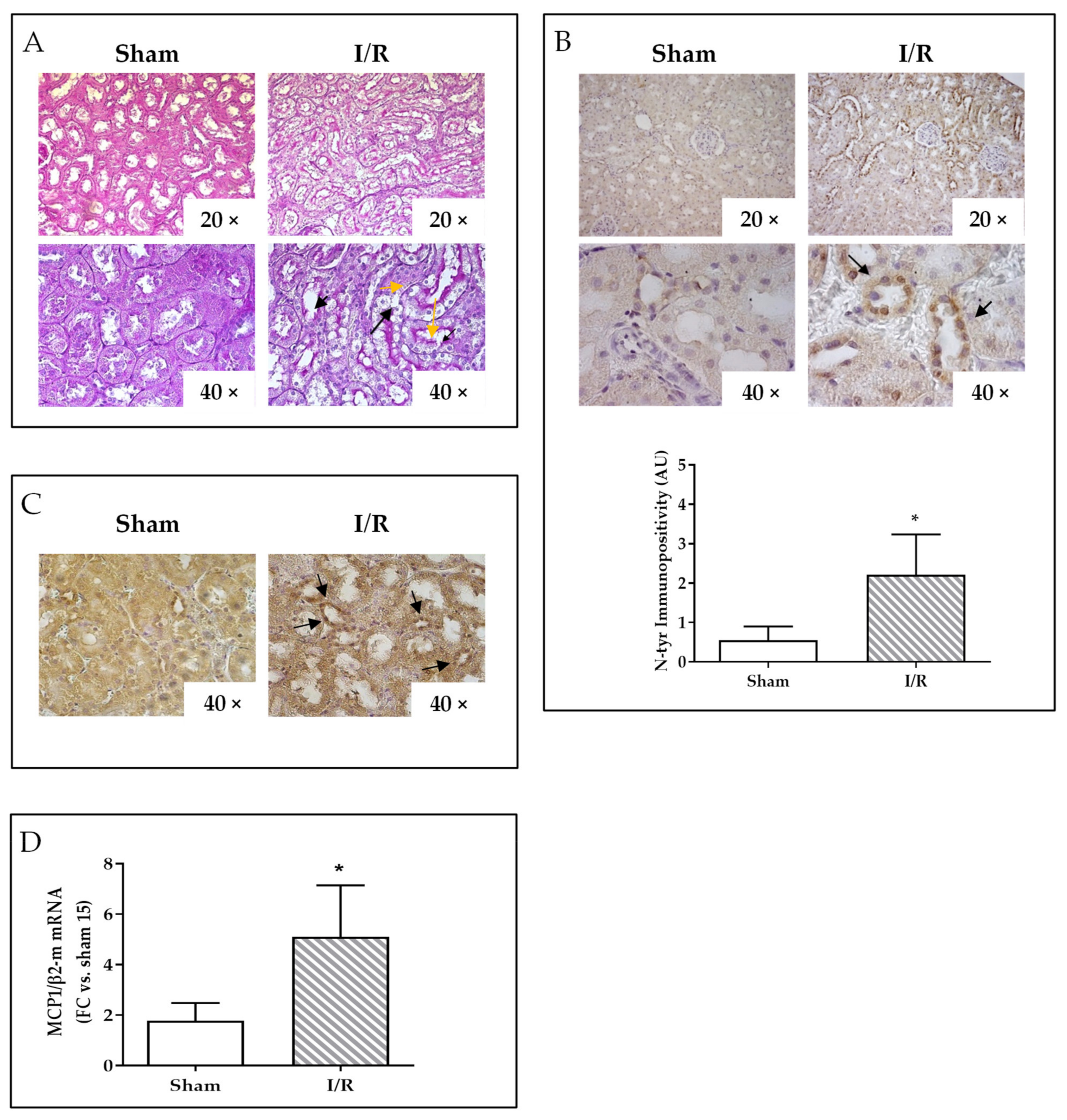
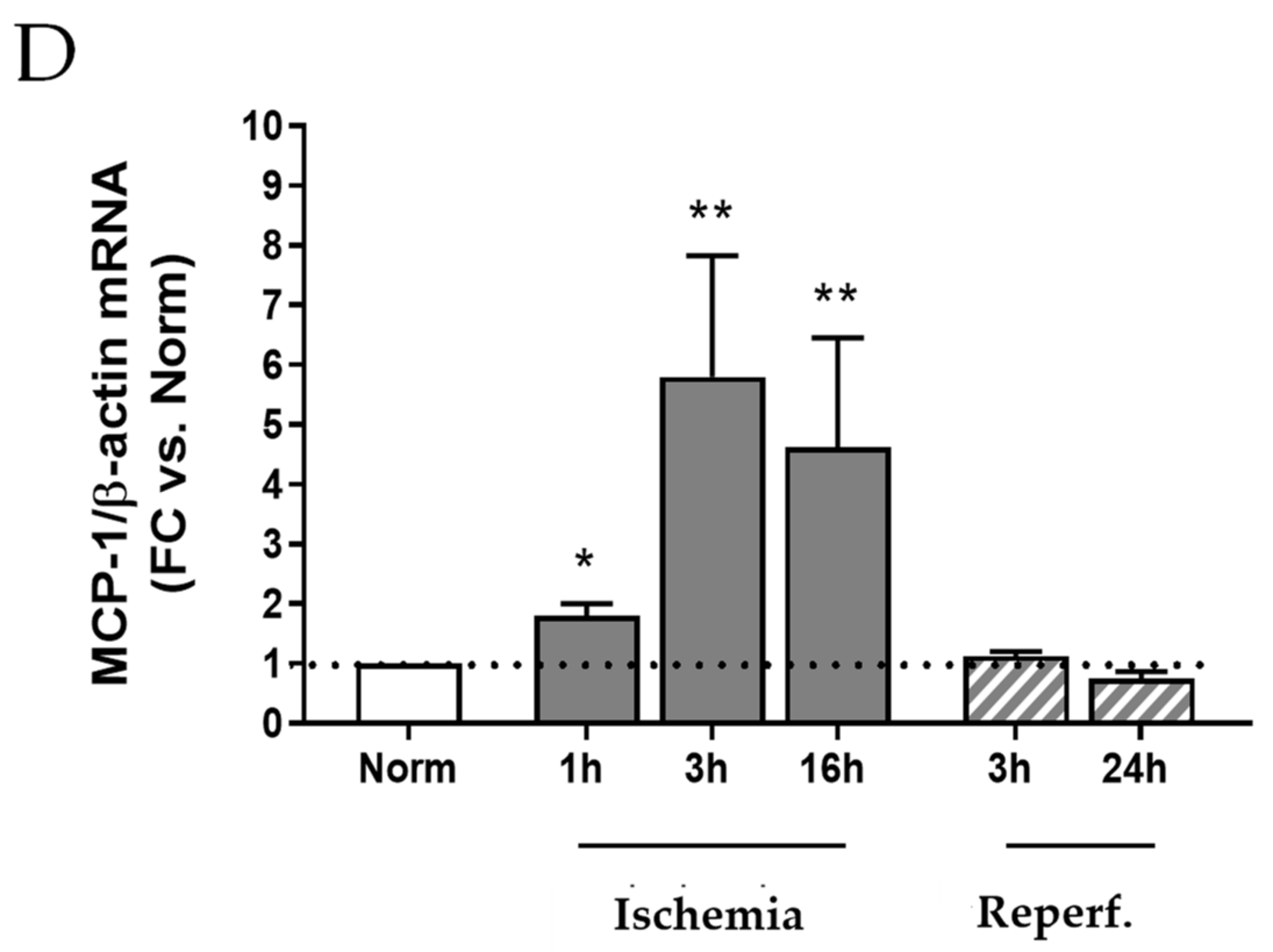
2.2. I/R Induces Focal Tubular Necrosis, Oxidative Stress, Peroxisome Proliferator-Activated Receptor Gamma Coactivator-1alpha (PGC-1α) Activation and Increased MCP-1 Gene Expression in Rat Kidneys 24 h after Reflow
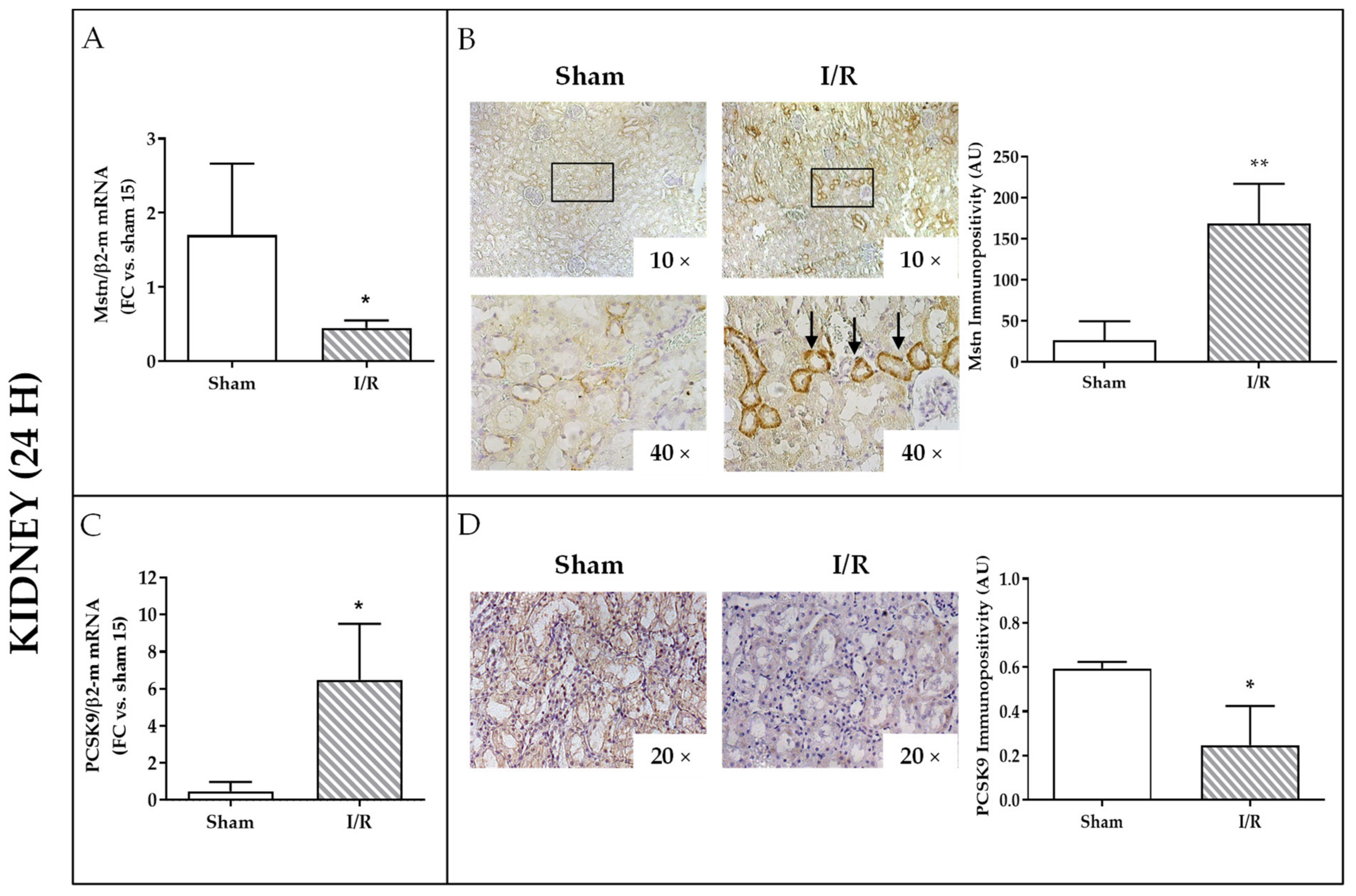
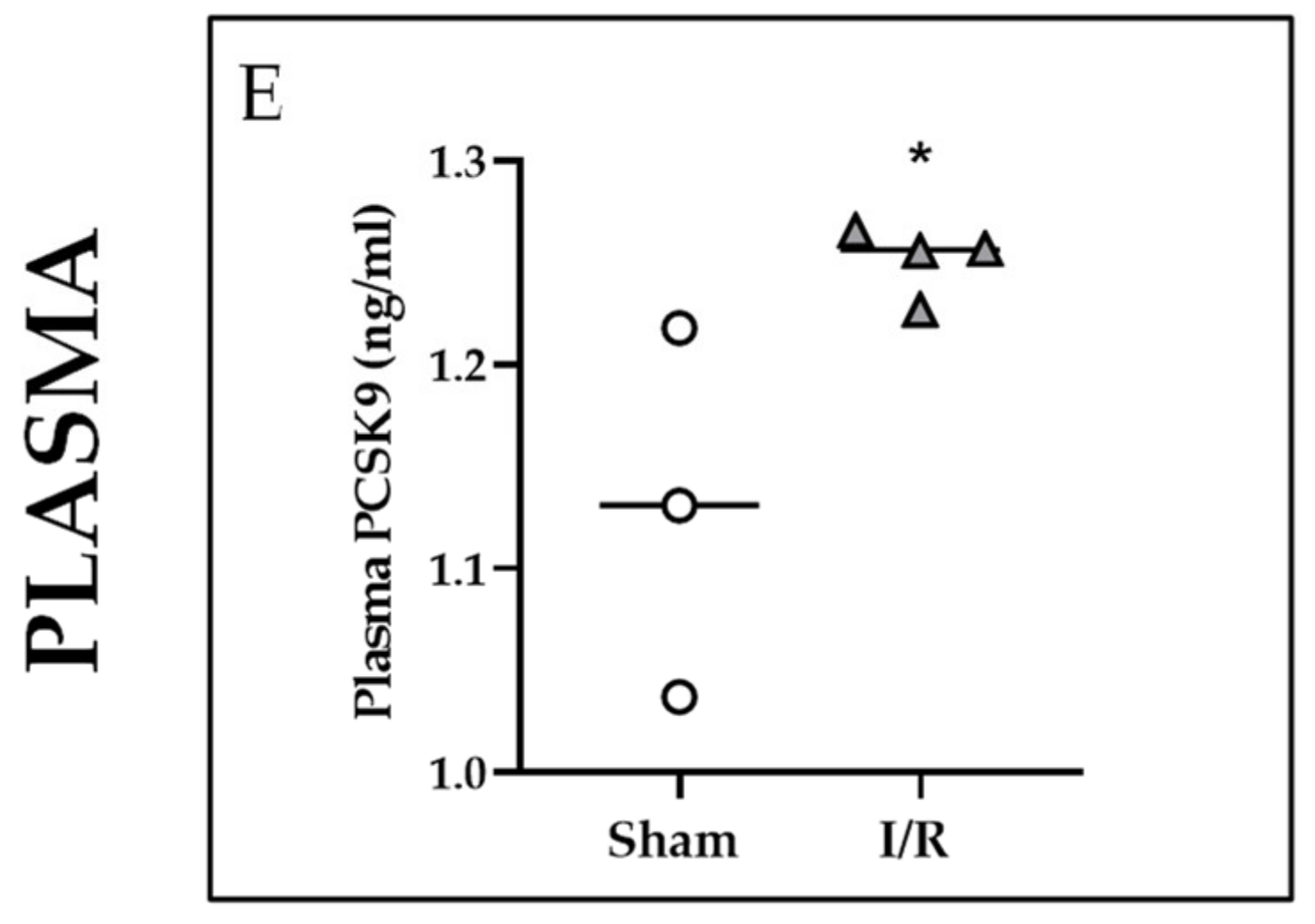
2.3. Effects of I/R on Mstn and PCSK9 in Rat Kidneys and on PCSK9 in Rat Plasma 24 h after Reflow
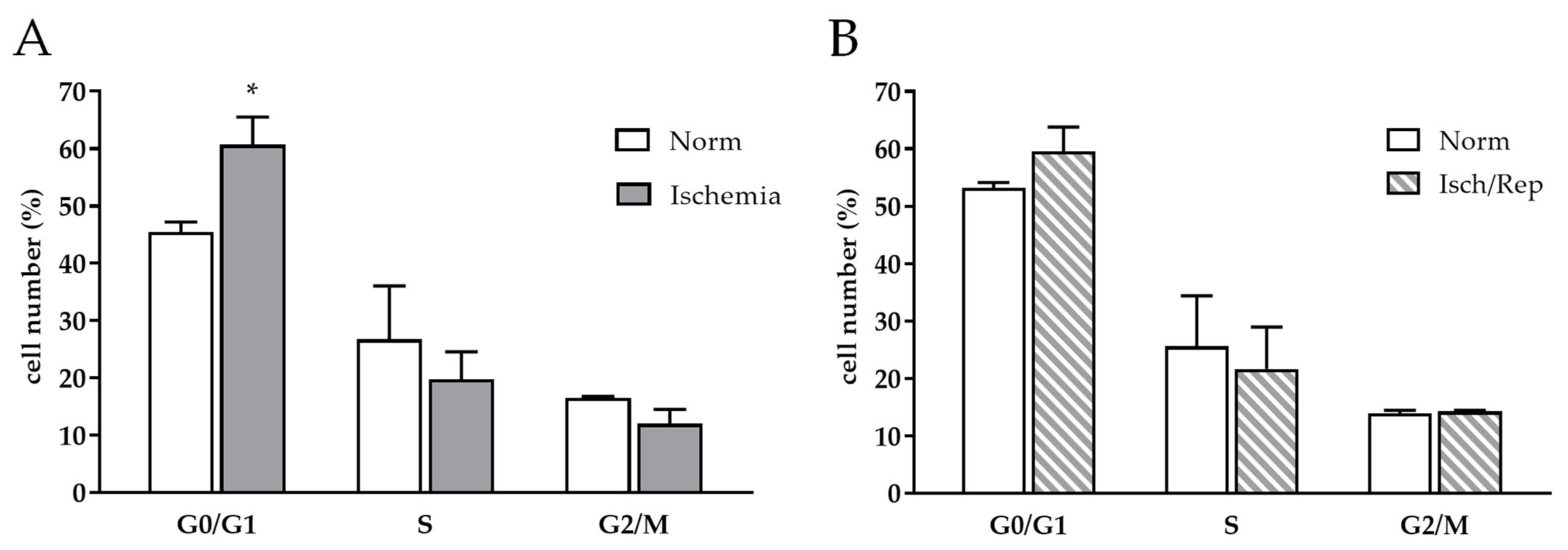
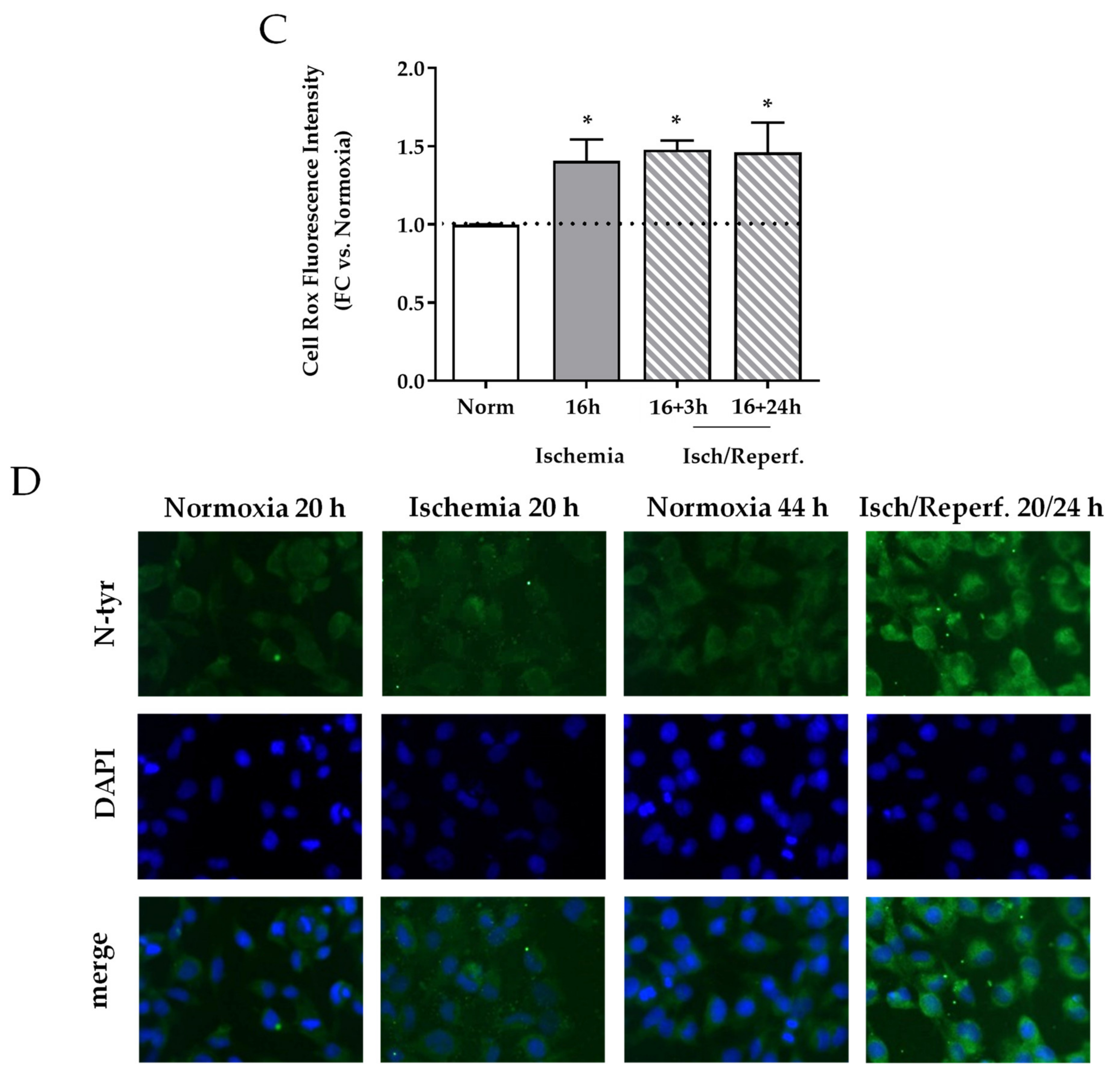
2.4. Effects of I/R on Cell Cycle and Oxidative Stress in HK-2 Renal Tubular Cells
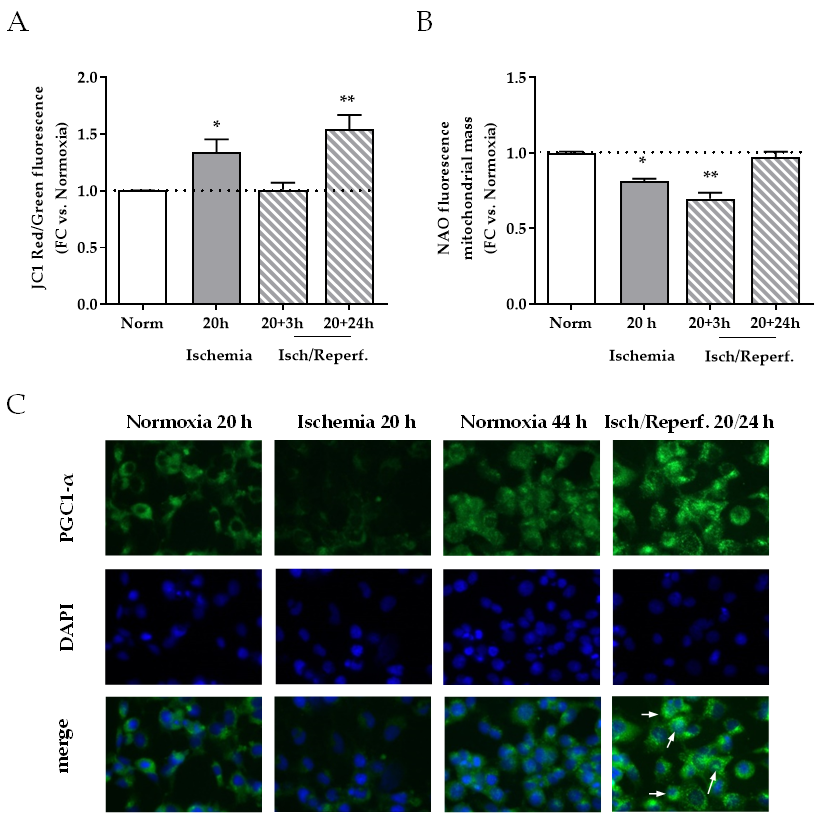
2.5. Effects of I/R on Mitochondria in HK-2 Renal Tubular Cells
2.6. Effects of I/R on Mstn and PCSK9 Expression in HK-2 Renal Tubular Cells
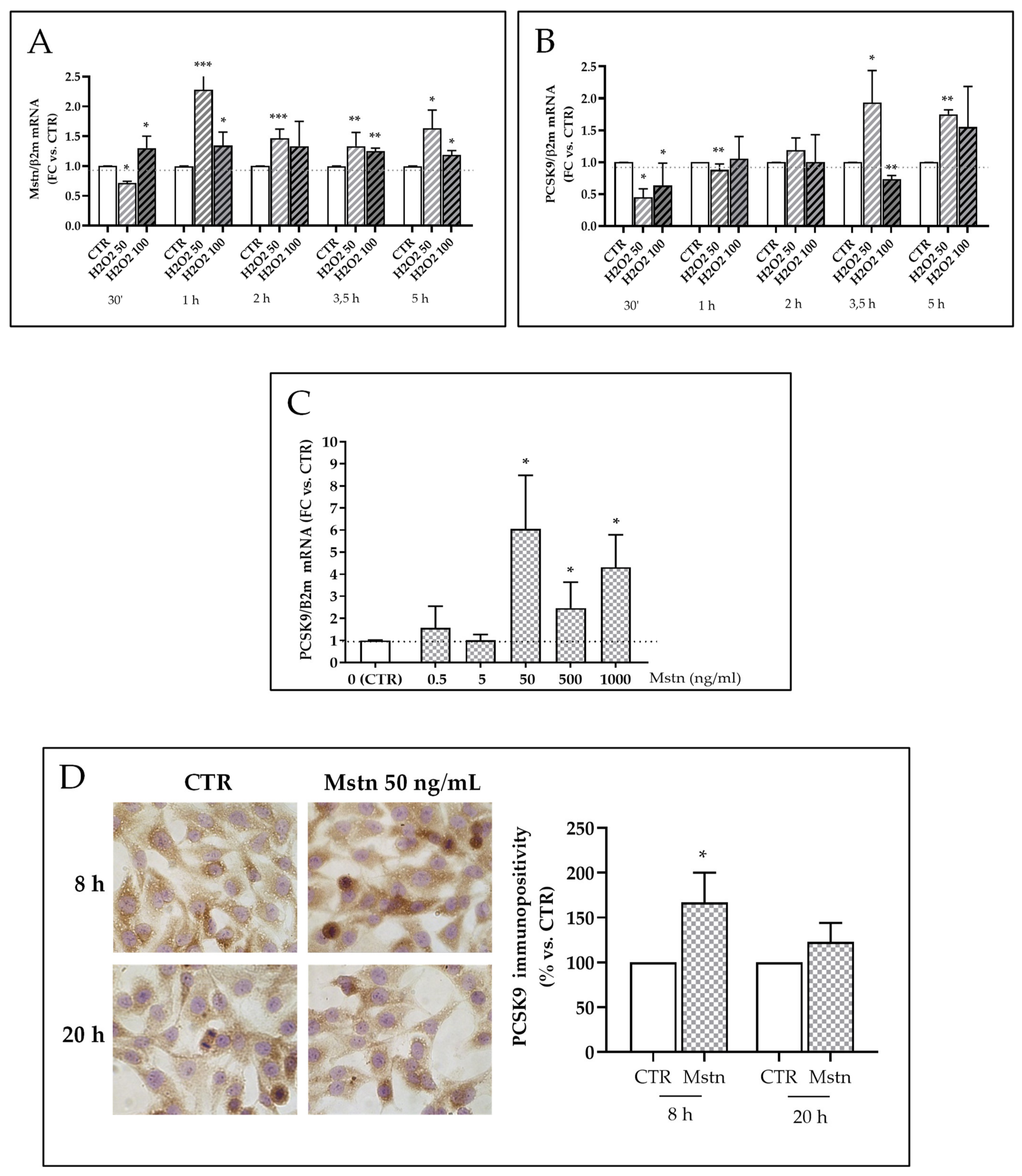
2.7. Mstn Induces PCSK9 Expression in HK-2 Renal Tubular Cells
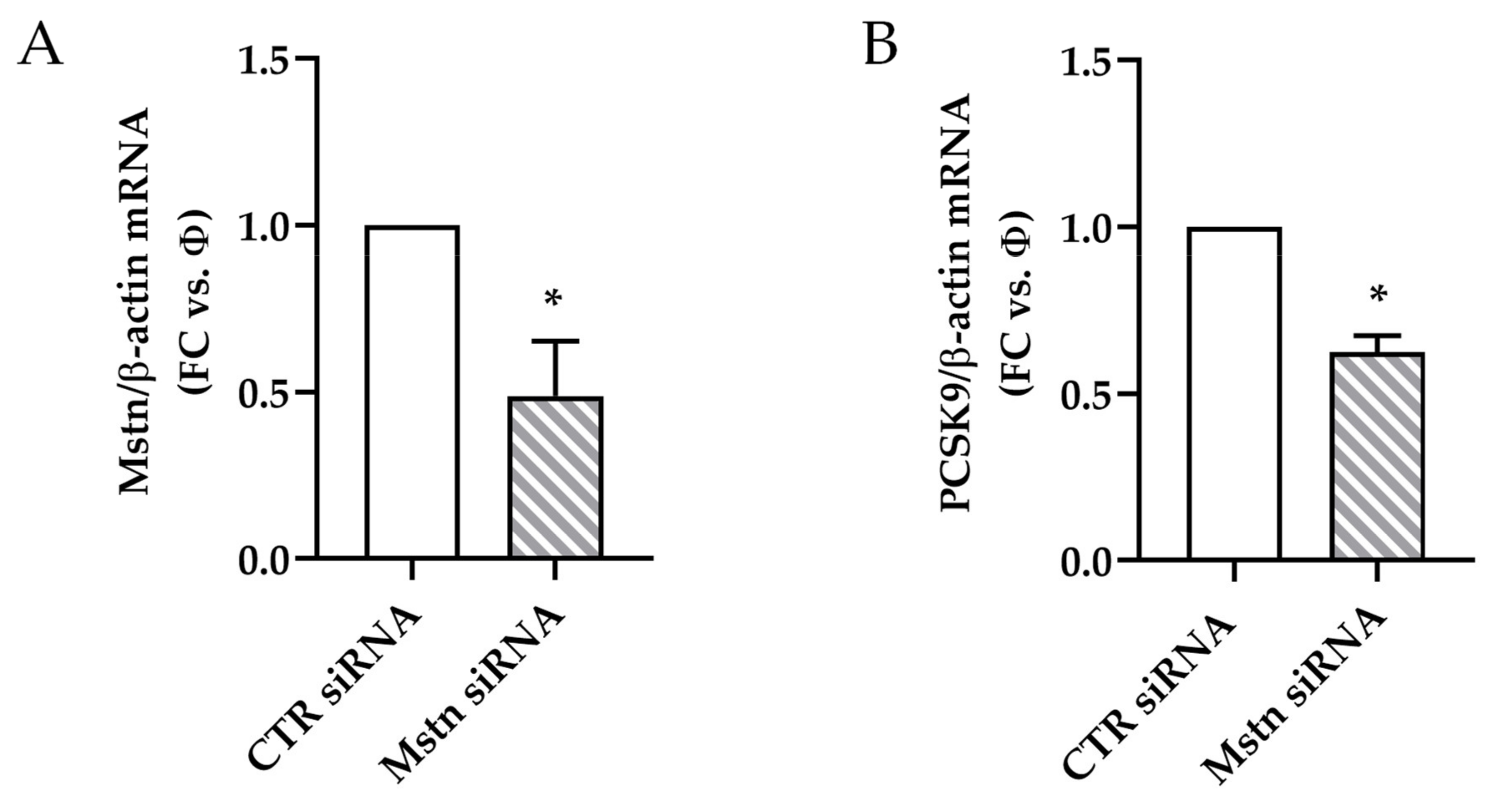
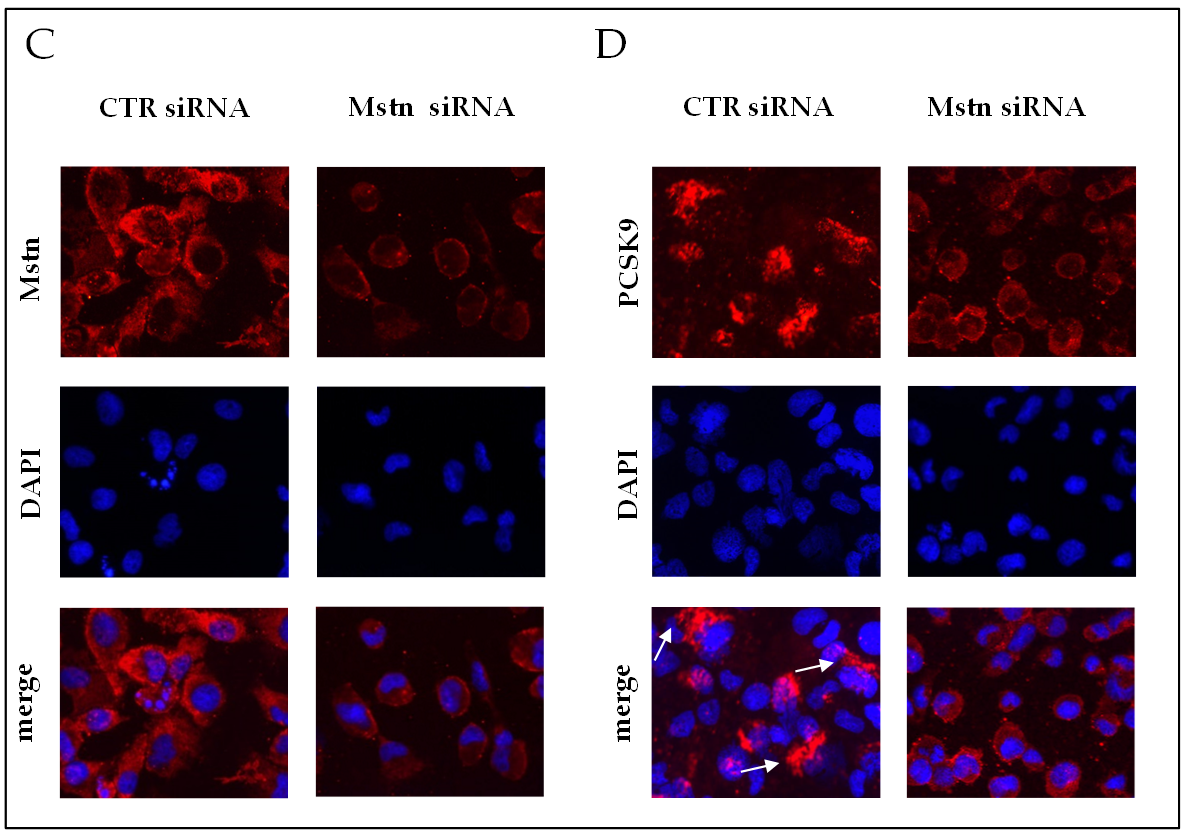
2.8. Effects of Mstn Silencing on PCSK9 Expression Induced by Ischemia in HK-2 Renal Tubular Cells
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. Histopathological Examination
4.3. Immunohistochemistry
4.4. PCSK9 and Creatinine Quantification in Plasma
4.5. Cell Cultures and Treatments
- ischemia-like conditions were reproduced by replacing complete medium with DMEM/F12 alone and incubating cells in a modular incubator chamber (Billups-Rothenberg, CA, USA), filled with an anoxic gas mixture (95% N2 and 5% CO2), at 37 °C for at most 20 h;
- reperfusion-like conditions were reproduced by exposing cells to fresh complete medium in standard conditions (atmospheric air supplemented with 5% CO2) up to 24 h;
- H2O2-dependent effects on Mstn and PCSK9 expression were evaluated by treating cells with two different concentrations of H2O2 (50 and 100 μM) (Carlo Erba, Milan, Italy) for different times (30 min, 1, 2, 3,5, and 5 h) in standard culture conditions;
- Mstn-dependent effects on PCSK9 expression were evaluated by adding recombinant Mstn (0.5–1000 ng/mL) (Peprotech, LiStarFish, Cernusco sul Naviglio, Italy) to complete culture medium for 5, 8, and 20 h in standard culture conditions;
- Mstn silencing was obtained by the transfection of HK-2 cells with 30 nM Mstn specific siRNA (Mstn siRNA) or negative control siRNA (CTR siRNA), also named “Φ”, using lipofectamine (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s protocol. Cells were then incubated at 37 °C in a CO2 incubator for 24 h until they were ready for assay. The efficacy of knockdown was determined by real-time PCR and immunofluorescence.
4.6. Flow Cytometry
4.7. Immunocytochemistry
4.8. Immunofluorescence
4.9. Image Analysis
4.10. mRNA Analysis
4.11. cDNA Reverse Transcription and Quantitative Real-Time PCR
4.12. Western Blotting
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Jassem, W.; Heaton, N.D. The role of mitochondria in ischemia/reperfusion injury in organ transplantation. Kidney Int. 2004, 66, 514–517. [Google Scholar] [CrossRef]
- Zhao, Z.; Tang, Z.; Zhang, W.; Liu, J.; Li, B. Magnesium isoglycyrrhizinate protects against renal-ischemia-reperfusion injury in a rat model via anti-inflammation, anti-oxidation and anti-apoptosis. Mol. Med. Rep. 2017, 16, 3627–3633. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Murphy, M.P.; Xing, W.; Wu, H.; Zhang, R.; Sun, H. Mitochondria-targeted antioxidant MitoQ reduced renal damage caused by ischemia-reperfusion injury in rodent kidneys: Longitudinal observations of T2-weighted imaging and dynamic contrast-enhanced MRI. Magn. Reson. Med. 2018, 79, 1559–1567. [Google Scholar] [CrossRef]
- Yamamoto, S.; Hagiwara, S.; Hidaka, S.; Shingu, C.; Goto, K.; Kashima, K.; Noguchi, T. The antioxidant EPC-K1 attenuates renal ischemia-reperfusion injury in a rat model. Am. J. Nephrol. 2011, 33, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rong, S.; Feng, Y.; Zhao, L.; Hong, J.; Wang, R.; Yuan, W. Simvastatin attenuates renal ischemia/reperfusion injury from oxidative stress via targeting Nrf2/HO-1 pathway. Exp. Ther. Med. 2017, 14, 4460–4466. [Google Scholar] [CrossRef]
- Nezu, M.; Souma, T.; Yu, L.; Suzuki, T.; Saigusa, D.; Ito, S.; Suzuki, N.; Yamamoto, M. Transcription factor Nrf2 hyperactivation in early-phase renal ischemia-reperfusion injury prevents tubular damage progression. Kidney Int. 2017, 91, 387–401. [Google Scholar] [CrossRef] [PubMed]
- De Vries, D.K.; Kortekaas, K.A.; Tsikas, D.; Wijermars, L.G.; van Noorden, C.J.; Suchy, M.T.; Cobbaert, C.M.; Klautz, R.J.; Schaapherder, A.F.; Lindeman, J.H. Oxidative damage in clinical ischemia/reperfusion injury: A reappraisal. Antioxid. Redox Signal. 2013, 19, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Pane, B.; Gazzola, V.; Spinella, G.; Bagnato, P.; Grillo, F.; Vellone, V.G.; Palombo, D. Inflammatory response modulation through a PPARγ agonist during surgically induced visceral ischemia in an animal model. Ann. Vasc. Surg. 2018, 48, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Wang, X.; Liu, S.; Shahanawaz, J.; Theus, S.; Fan, Y.; Deng, X.; Zhou, S.; Mehta, J.L. PCSK9 expression in the ischaemic heart and its relationship to infarct size, cardiac function, and development of autophagy. Cardiovasc. Res. 2018, 114, 1738–1751. [Google Scholar] [CrossRef]
- Palee, S.; McSweeney, C.M.; Maneechote, C.; Moisescu, D.M.; Jaiwongkam, T.; Kerdphoo, S.; Chattipakorn, S.C.; Chattipakorn, N. PCSK9 inhibitor improves cardiac function and reduces infarct size in rats with ischaemia/reperfusion injury: Benefits beyond lipid-lowering effects. J. Cell. Mol. Med. 2019, 11, 7310–7319. [Google Scholar] [CrossRef]
- Pomiès, P.; Blaquière, M.; Maury, J.; Mercier, J.; Gouzi, F.; Hayot, M. Involvement of the FoxO1/MuRF1/Atrogin-1 signaling pathway in the oxidative stress-induced atrophy of cultured chronic obstructive pulmonary disease myotubes. PLoS ONE 2016, 11, e0160092. [Google Scholar] [CrossRef]
- Castillero, E.; Akashi, H.; Wang, C.; Najjar, M.; Ji, R.; Kennel, P.J.; Sweeney, H.L.; Schulze, P.C.; George, I. Cardiac myostatin upregulation occurs immediately after myocardial ischemia and is involved in skeletal muscle activation of atrophy. Biochem. Biophys. Res. Commun. 2015, 30, 106–111. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Verzola, D.; Milanesi, S.; Bertolotto, M.; Garibaldi, S.; Villaggio, B.; Brunelli, C.; Balbi, M.; Ameri, P.; Montecucco, F.; Palombo, D.; et al. Myostatin mediates abdominal aortic atherosclerosis progression by inducing vascular smooth muscle cell dysfunction and monocyte recruitment. Sci. Rep. 2017, 7, 46362. [Google Scholar] [CrossRef] [PubMed]
- Verzola, D.; Milanesi, S.; Viazzi, F.; Ansaldo, F.; Saio, M.; Garibaldi, S.; Carta, A.; Costigliolo, F.; Salvidio, G.; Barisione, C.; et al. Enhanced myostatin expression and signalling promote tubulointerstitial inflammation in diabetic nephropathy. Sci. Rep. 2020, 10, 6343. [Google Scholar] [CrossRef] [PubMed]
- Shanley, P.F.; Rosen, M.D.; Brezis, M.; Silva, P.; Epstein, F.H.; Rosen, S. Topography of focal proximal tubular necrosis after ischemia with reflow in the rat kidney. Am. J. Pathol. 1986, 122, 462–468. [Google Scholar]
- Knott, A.W.; Kalra, M.; Duncan, A.A.; Reed, N.R.; Bower, T.C.; Hoskin, T.L.; Oderich, G.S.; Gloviczki, P. Open repair of juxtarenal aortic aneurysms (JAA) remains a safe option in the era of fenestrated endografts. J. Vasc. Surg. 2008, 47, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Jean-Claude, J.M.; Reilly, L.M.; Stoney, R.J.; Messina, L.M. Pararenal aortic aneurysms: The future of open aortic aneurysm repair. J. Vasc. Surg. 1999, 29, 902–912. [Google Scholar] [CrossRef][Green Version]
- Sarac, T.P.; Clair, D.G.; Hertzer, N.R.; Greenberg, R.K.; Krajewski, L.P.; O’Hara, P.J.; Ouriel, K. Contemporary results of juxtarenal aneurysm repair. J. Vasc. Surg. 2002, 36, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Jongkind, V.; Yeung, K.K.; Akkersdijk, G.J.M.; Heidsieck, D.; Reitsma, J.B.; Tangelder, G.J.; Wisselink, W. Juxtarenal aortic aneurysm repair. J. Vasc. Surg. 2010, 52, 760–767. [Google Scholar] [CrossRef]
- Bicknell, C.D.; Cowan, A.R.; Kerle, M.I.; Mansfield, A.O.; Cheshire, N.J.W.; Wolfe, J.H.N. Renal dysfunction and prolonged visceral ischaemia increase mortality rate after suprarenal aneurysm repair. Br. J. Surg. 2003, 90, 1142–1146. [Google Scholar] [CrossRef]
- Norwood, M.G.A.; Polimenovi, N.M.; Sutton, A.J.; Bown, M.J.; Sayers, R.D. Abdominal aortic aneurysm repair in patients with chronic renal disease. Eur. J. Vasc. Endovasc. Surg. 2004, 27, 287–291. [Google Scholar] [CrossRef][Green Version]
- Wahlberg, E.; DiMuzio, P.J.; Stoney, R.J. Aortic clamping during elective operations for infrarenal disease: The influence of clamping time on renal function. J. Vasc. Surg. 2002, 36, 13–18. [Google Scholar] [CrossRef]
- Bartekova, M.; Barancik, M.; Ferenczyova, K.; Dhalla, N.S. Beneficial effects of N-acetylcysteine and N-mercaptopropionylglycine on ischemia reperfusion injury in the heart. Curr. Med. Chem. 2018, 25, 355–366. [Google Scholar] [CrossRef]
- Demetrius, L. Of mice and men. When it comes to studying ageing and the means to slow it down, mice are not just small humans. EMBO Rep. 2005, 6, S39–S44. [Google Scholar]
- Ralto, K.M.; Rhee, E.P.; Parikh, S.M. NAD+ homeostasis in renal health and disease. Nat. Rev. Nephrol. 2020, 16, 99–111. [Google Scholar] [CrossRef]
- Pefanis, A.; Ierino, F.L.; Murphy, J.M.; Cowan, P.J. Regulated necrosis in kidney ischemia-reperfusion injury. Kidney Int. 2019, 96, 291–301. [Google Scholar] [CrossRef]
- Lee, H.; Abe, Y.; Lee, I.; Shrivastav, S.; Crusan, A.P.; Hüttemann, M.; Hopfer, U.; Feld, R.A. Increased mitochondrial activity in renal proximal tubule cells from young spontaneously hypertensive rats. Kidney Int. 2014, 85, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Enoki, Y.; Watanabe, H.; Arake, R.; Sugimoto, R.; Imafuku, T.; Tominaga, Y.; Ishima, Y.; Kotani, S.; Nakajima, M.; Tanaka, M.; et al. Indoxyl sulfate potentiates skeletal muscle atrophy by inducing the oxidative stress-mediated expression of myostatin and atrogin-1. Sci. Rep. 2016, 23, 32084. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, H.; Zhou, Y.; Zhu, Y.; Bian, R.; Chen, Y.; Li, C.; Ma, Q.; Zheng, Q.; Zhang, Y.; et al. Myostatin induces mitochondrial metabolic alteration and typical apoptosis in cancer cells. Cell Death Dis. 2013, 4, e494. [Google Scholar] [CrossRef] [PubMed]
- Sriram, S.; Subramanian, S.; Sathiakumar, D.; Venkatesh, R.; Salerno, M.S.; McFarlane, C.D.; Kambadur, R.; Sharma, M. Modulation of reactive oxygen species in skeletal muscle by myostatin is mediated through NF-κB. Aging Cell 2011, 10, 931–948. [Google Scholar] [CrossRef]
- Lim, S.; McMahon, C.D.; Matthews, K.G.; Devlin, G.P.; Elston, M.S.; Conaglen, J.V. Absence of myostatin improves cardiac function following myocardial infarction. Heart Lung Circ. 2018, 27, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Castillero, E.; Akashi, H.; Najjar, M.; Ji, R.; Brandstetter, L.M.; Wang, C.; Liao, X.; Zhang, X.; Sperry, A.; Gailes, M.; et al. Activin type II receptor ligand signaling inhibition after experimental ischemic heart failure attenuates cardiac remodeling and prevents fibrosis. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H378–H390. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Zeng, Y.D.; Hu, Z.X.; Linag, H. PCSK9 promotes the secretion of pro-inflammatory cytokines by macrophages to aggravate H/R-induced cardiomyocyte injury via activating NF-κB signalling. Gen. Physiol. Biophys. 2020, 39, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Pothineni, N.V.K.; Goel, A.; Lüscher, T.F.; Mehta, J.L. PCSK9 and inflammation: Role of shear stress, pro-inflammatory cytokines, and LOX-1. Cardiovasc. Res. 2020, 116, 908–915. [Google Scholar] [CrossRef]
- Andreadou, I.; Tsoumani, M.; Vilahur, G.; Ikonomidis, I.; Badimon, L.; Varga, Z.V.; Ferdinandy, P.; Schulz, R. PCSK9 in Myocardial infarction and cardioprotection: Importance of lipid metabolism and inflammation. Front. Physiol. 2020, 11, 602497. [Google Scholar] [CrossRef]
- Gencer, B.; Mach, F.; Murphy, S.A.; De Ferrari, G.M.; Huber, K.; Lewis, B.S.; Ferreira, J.; Kurtz, C.E.; Wang, H.; Honarpour, N.; et al. Efficacy of evolocumab on cardiovascular outcomes in patients with recent myocardial infarction: A prespecified secondary analysis from the FOURIER trial. JAMA Cardiol. 2020, 5, 952–957. [Google Scholar] [CrossRef]
- White, H.D.; Steg, P.G.; Szarek, M.; Bhatt, D.L.; Bittner, V.A.; Diaz, R.; Edelberg, J.M.; Erglis, A.; Goodman, S.G.; Hanotin, C.; et al. Effects of alirocumab on types of myocardial infarction: Insights from the OYSSEY OUTCOMES trial. Eur. Heart J. 2019, 40, 2801–2809. [Google Scholar] [CrossRef]
- Stoekenbroek, R.M.; Lambert, G.; Cariou, B.; Hovingh, G.K. Inhibiting PCSK9—Biology beyond LDL control. Nat. Rev. Endocrinol. 2018, 1, 52–62. [Google Scholar] [CrossRef]
- Miura, T.; Kishioka, Y.; Wakamatsu, J.-I.; Hattori, A.; Nishimura, T. Interaction between myostatin and extracellular matrix components. Anim. Sci. J. 2010, 81, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Speir, R.W.; Stallings, J.D.; Andrews, J.M.; Gelnett, M.S.; Brand, T.C.; Salgar, S.K. Effects of valproic acid and dexamethasone administration on early bio-markers and gene expression profile in acute kidney ischemia-reperfusion injury in the rat. PLoS ONE 2015, 13, e0126622. [Google Scholar] [CrossRef]
- Verzola, D.; Cappuccino, L.; D’Amato, E.; Villaggio, B.; Gianiorio, F.; Mij, M.; Simonato, A.; Viazzi, F.; Salvidio, G.; Garibotto, G. Enhanced glomerular Toll-like receptor 4 expression and signaling in patients with type 2 diabetic nephropathy and microalbuminuria. Kidney Int. 2014, 86, 1229–1243. [Google Scholar] [CrossRef] [PubMed]
- Spallarossa, P.; Garibaldi, S.; Barisione, C.; Ghigliotti, G.; Altieri, P.; Tracchi, I.; Fabbi, P.; Barsotti, A.; Brunelli, C. Postprandial serum induces apoptosis in endothelial cells: Role of polymorphonuclear-derived myeloperoxidase and metalloproteinase-9 activity. Atherosclerosis 2008, 198, 458–467. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Species | Accession number | Forward | Reverse |
|---|---|---|---|---|
| Proprotein convertase subtilisin/kexin type 9 (PCSK9) | rat | NM_199253 | 5′ TCT.AAG AGA TAC ACC TCC ACC TG 3′ | 5′ ACT CAT TAG TCT TCG CCC AGA 3′ |
| Myostatin (Mstn) | rat | NM_019151.1 | 5′GGCACTGGTATTTGGCAGAGT 3′ | 5′AGGGATTCAGCCCATCTTCTC 3′ |
| Monocyte chemoattractant protein-1 (MCP-1) | rat | NM_031530.1 | 5′TGACAAATACTACAGCTTCTTTGGG 3′ | 5′CAGTTAATGCCCCACTCACCT 3′ |
| β2-microglobulin (β2-m) | rat | NM_012512 | 5′ TAG CAG TTG AGG AAG TTG GG 3′ | 5′ TGA TCT TTC TGG TGC TTG TCT C 3′ |
| Proprotein convertase subtilisin/kexin type 9 (PCSK-9) | human | NM_174936 | 5′ AGG GGA GGA CAT CAT TGG TG 3′ | 5′-CAG GTT GGG GGT CAG TAC C 3′ |
| Myostatin (Mstn) | human | NM_005259. | 5′CCAAAGCTCCTCCACTCCG 3′ | 5′GGGAGTACAGCAAGGGCC 3′ |
| Monocyte chemoattractant protein-1 (MCP-1) | human | NM_002982 | 5′ACCGAGAGGCTGAGACTAAC 3′ | 5′AATGAAGGTGGCTGCTATGAG 3′ |
| β2-microglobulin (β2-m) | human | NM_004048 | 5′ CCA GCG TAC TCC AAA GAT TCA 3′ | 5′ TGC TCC ACT TTT TCA ATT CTC TC 3′ |
| β-actin | human | NM_001101.2 | 5′CCTCGCCTTTGCCGATCC 3′ | 5′CTCGTCGCCCACATAGGAAT 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barisione, C.; Verzola, D.; Garibaldi, S.; Ferrari, P.F.; Garibotto, G.; Ameri, P.; Pane, B.; Spinella, G.; Pratesi, G.; Palombo, D. Renal Ischemia/Reperfusion Early Induces Myostatin and PCSK9 Expression in Rat Kidneys and HK-2 Cells. Int. J. Mol. Sci. 2021, 22, 9884. https://doi.org/10.3390/ijms22189884
Barisione C, Verzola D, Garibaldi S, Ferrari PF, Garibotto G, Ameri P, Pane B, Spinella G, Pratesi G, Palombo D. Renal Ischemia/Reperfusion Early Induces Myostatin and PCSK9 Expression in Rat Kidneys and HK-2 Cells. International Journal of Molecular Sciences. 2021; 22(18):9884. https://doi.org/10.3390/ijms22189884
Chicago/Turabian StyleBarisione, Chiara, Daniela Verzola, Silvano Garibaldi, Pier Francesco Ferrari, Giacomo Garibotto, Pietro Ameri, Bianca Pane, Giovanni Spinella, Giovanni Pratesi, and Domenico Palombo. 2021. "Renal Ischemia/Reperfusion Early Induces Myostatin and PCSK9 Expression in Rat Kidneys and HK-2 Cells" International Journal of Molecular Sciences 22, no. 18: 9884. https://doi.org/10.3390/ijms22189884
APA StyleBarisione, C., Verzola, D., Garibaldi, S., Ferrari, P. F., Garibotto, G., Ameri, P., Pane, B., Spinella, G., Pratesi, G., & Palombo, D. (2021). Renal Ischemia/Reperfusion Early Induces Myostatin and PCSK9 Expression in Rat Kidneys and HK-2 Cells. International Journal of Molecular Sciences, 22(18), 9884. https://doi.org/10.3390/ijms22189884