
Prognostic Impact and Functional Annotations of KIF11 and KIF14 Expression in Patients with Colorectal Cancer

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Immunohistochemical Expression of KIF11 and KIF14 Proteins: Association with Clinicopathological Parameters
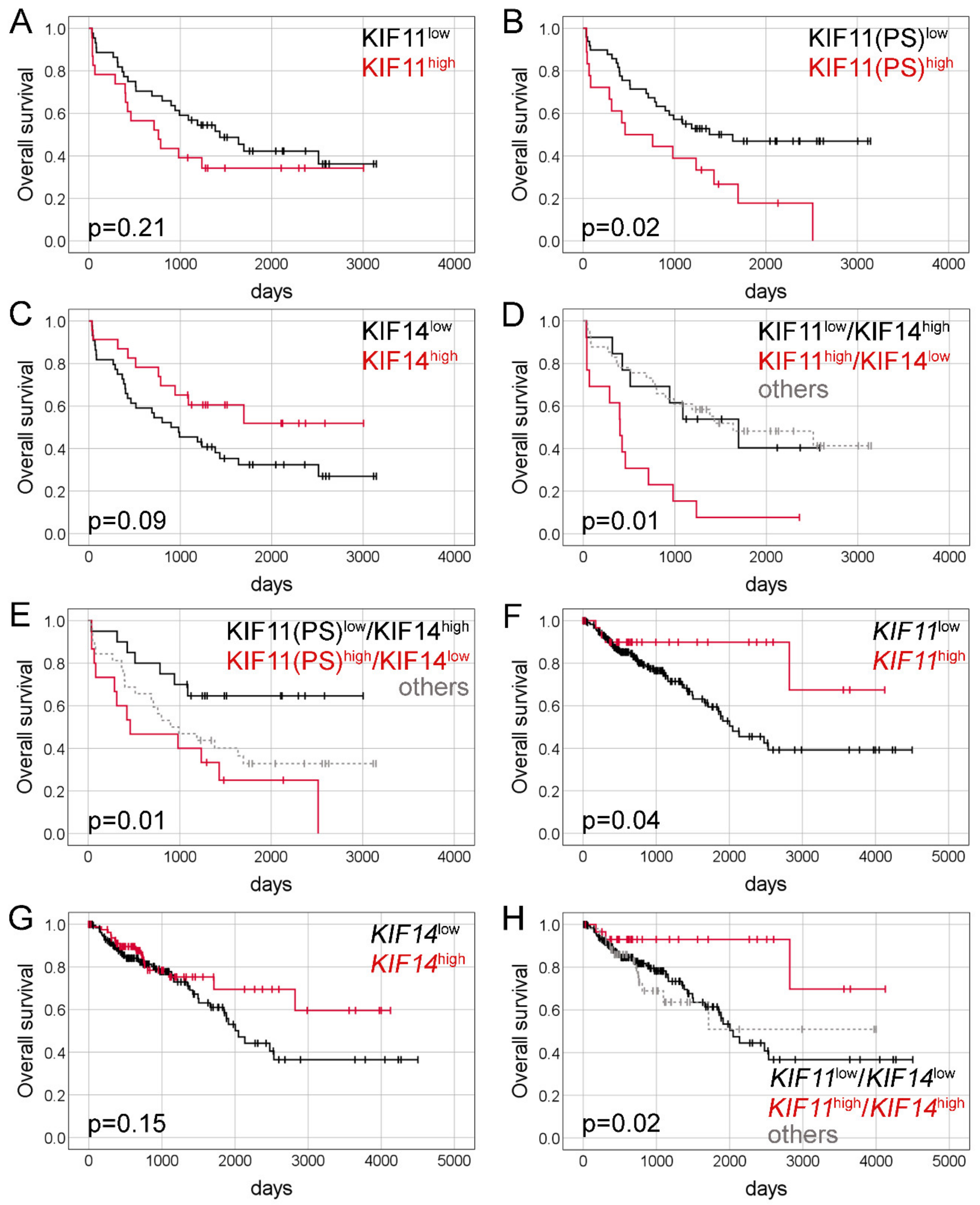
2.2. Immunohistochemical Expression of KIF11 and KIF14 Proteins: Association with Overall Survival
2.3. Expression of KIF11 and KIF14 Genes: Association with Clinicopathological Parameters
2.4. Expression of KIF11 and KIF14 Genes: Association with Overall Survival
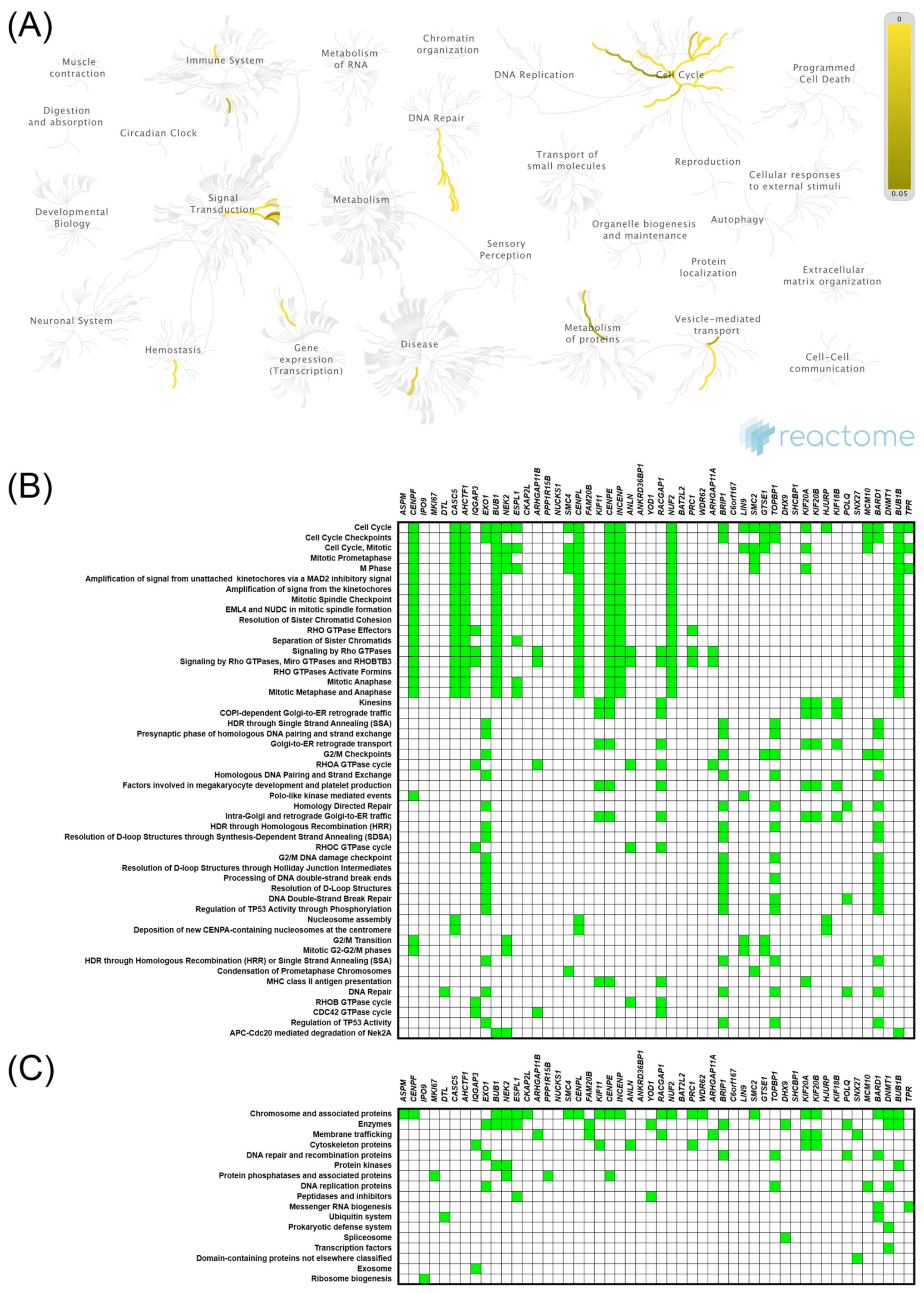
2.5. Expression of KIF11 and KIF14 Genes: Functional Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Tissue Material and Clinicopathological Data
4.2. Survival Data
4.3. Immunohistochemical Analysis
4.4. In Silico Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of Colorectal Cancer: Incidence, Mortality, Survival, and Risk Factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Goel, A.; Chung, D.C. Pathways of Colorectal Carcinogenesis. Gastroenterology 2020, 158, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Westdorp, H.; Fennemann, F.L.; Weren, R.D.A.; Bisseling, T.M.; Ligtenberg, M.J.L.; Figdor, C.G.; Schreibelt, G.; Hoogerbrugge, N.; Wimmers, F.; de Vries, I.J.M. Opportunities for Immunotherapy in Microsatellite Instable Colorectal Cancer. Cancer Immunol. Immunother. 2016, 65, 1249–1259. [Google Scholar] [CrossRef]
- Afrăsânie, V.-A.; Marinca, M.V.; Alexa-Stratulat, T.; Gafton, B.; Păduraru, M.; Adavidoaiei, A.M.; Miron, L.; Rusu, C. KRAS, NRAS, BRAF, HER2 and Microsatellite Instability in Metastatic Colorectal Cancer—Practical Implications for the Clinician. Radiol. Oncol. 2019, 53, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Chang, M.; Chang, H.M.; Chang, F. Expending Role of Microsatellite Instability in Diagnosis and Treatment of Colorectal Cancers. J. Gastrointest. Cancer 2017, 48, 305–313. [Google Scholar] [CrossRef]
- Ding, X.; Duan, H.; Luo, H. Identification of Core Gene Expression Signature and Key Pathways in Colorectal Cancer. Front. Genet. 2020, 11, 45. [Google Scholar] [CrossRef]
- Pirim, D. Integrative Analyses of Molecular Pathways and Key Candidate Biomarkers Associated with Colorectal Cancer. Cancer Biomark. 2020, 27, 555–568. [Google Scholar] [CrossRef]
- Cai, Z.; Wei, Y.; Chen, S.; Gong, Y.; Fu, Y.; Dai, X.; Zhou, Y.; Yang, H.; Tang, L.; Liu, H. Screening and Identification of Key Biomarkers in Alimentary Tract Cancers: A Bioinformatic Analysis. Cancer Biomark. 2020, 29, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.; Morse, H.C.; Godfrey, V.L.; Naeem, R.; Justice, M.J. Overexpression of Eg5 Causes Genomic Instability and Tumor Formation in Mice. Cancer Res. 2007, 67, 10138–10147. [Google Scholar] [CrossRef]
- Sishtla, K.; Pitt, N.; Shadmand, M.; O’Hare, M.N.; Sulaiman, R.S.; Sinn, A.L.; Condon, K.; Pollok, K.E.; Sandusky, G.E.; Corson, T.W. Observations on Spontaneous Tumor Formation in Mice Overexpressing Mitotic Kinesin Kif14. Sci. Rep. 2018, 8, 16152. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Zhao, J.; Bibikova, M.; Leverson, J.D.; Bossy-Wetzel, E.; Fan, J.-B.; Abraham, R.T.; Jiang, W. Functional Analysis of Human Microtubule-Based Motor Proteins, the Kinesins and Dyneins, in Mitosis/Cytokinesis Using RNA Interference. Mol. Biol. Cell 2005, 16, 3187–3199. [Google Scholar] [CrossRef] [PubMed]
- Rapley, J.; Nicolàs, M.; Groen, A.; Regué, L.; Bertran, M.T.; Caelles, C.; Avruch, J.; Roig, J. The NIMA-Family Kinase Nek6 Phosphorylates the Kinesin Eg5 at a Novel Site Necessary for Mitotic Spindle Formation. J. Cell Sci. 2008, 121, 3912–3921. [Google Scholar] [CrossRef] [PubMed]
- Daigo, K.; Takano, A.; Thang, P.M.; Yoshitake, Y.; Shinohara, M.; Tohnai, I.; Murakami, Y.; Maegawa, J.; Daigo, Y. Characterization of KIF11 as a Novel Prognostic Biomarker and Therapeutic Target for Oral Cancer. Int. J. Oncol. 2018, 52, 155–165. [Google Scholar] [CrossRef]
- Pei, Y.-Y.; Li, G.-C.; Ran, J.; Wei, F.-X. Kinesin Family Member 11 Contributes to the Progression and Prognosis of Human Breast Cancer. Oncol. Lett. 2017, 14, 6618–6626. [Google Scholar] [CrossRef][Green Version]
- Liu, J.; Tian, Y.; Yi, L.; Gao, Z.; Lou, M.; Yuan, K. High KIF11 Expression Is Associated with Poor Outcome of NSCLC. Tumori J. 2021, 300891620988342. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, N.; Li, J.; Kong, J.; Guan, X.; Wang, X. Eg5 Overexpression Is Predictive of Poor Prognosis in Hepatocellular Carcinoma Patients. Dis. Markers 2017, 2017, 2176460. [Google Scholar] [CrossRef]
- Jin, Q.; Dai, Y.; Wang, Y.; Zhang, S.; Liu, G. High Kinesin Family Member 11 Expression Predicts Poor Prognosis in Patients with Clear Cell Renal Cell Carcinoma. J. Clin. Pathol. 2019, 72, 354–362. [Google Scholar] [CrossRef]
- Imai, T.; Oue, N.; Sentani, K.; Sakamoto, N.; Uraoka, N.; Egi, H.; Hinoi, T.; Ohdan, H.; Yoshida, K.; Yasui, W. KIF11 Is Required for Spheroid Formation by Oesophageal and Colorectal Cancer Cells. Anticancer Res. 2017, 37, 47–55. [Google Scholar] [CrossRef]
- Klimaszewska-Wiśniewska, A.; Neska-Długosz, I.; Buchholz, K.; Durślewicz, J.; Grzanka, D.; Kasperska, A.; Antosik, P.; Zabrzyński, J.; Grzanka, A.; Gagat, M. Prognostic Significance of KIF11 and KIF14 Expression in Pancreatic Adenocarcinoma. Cancers 2021, 13, 3017. [Google Scholar] [CrossRef]
- Ogłuszka, M.; Orzechowska, M.; Jędroszka, D.; Witas, P.; Bednarek, A.K. Evaluate Cutpoints: Adaptable Continuous Data Distribution System for Determining Survival in Kaplan-Meier Estimator. Comput. Methods Programs Biomed. 2019, 177, 133–139. [Google Scholar] [CrossRef]
- Sun, P.; Cao, X.; Liu, Y.; Jia, L. Eg5 High Expression Predicts Dismal Prognosis in Epithelial Ovarian Cancer. Blood Genom. 2019, 3, 145–151. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, L.; Xiong, L.; Wang, K.; Hou, X.; Li, Q.; Kong, F.; Liu, X.; He, J. KIF11 Is Upregulated in Colorectal Cancer and Silencing of It Impairs Tumor Growth and Sensitizes Colorectal Cancer Cells to Oxaliplatin via P53/GSK3β Signaling. J. Cancer 2021, 12, 3741–3753. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, L.; Zhang, B.; Chen, L. Identification of Differentially Expressed Genes (DEGs) Relevant to Prognosis of Ovarian Cancer by Use of Integrated Bioinformatics Analysis and Validation by Immunohistochemistry Assay. Med. Sci. Monit. 2019, 25, 9902–9912. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-F.; Zeng, H.-J.; Shan, Z.; Ye, R.-Y.; Cheang, T.-Y.; Zhang, Y.-J.; Lu, S.-H.; Zhang, Q.; Shao, N.; Lin, Y. Overexpression of Kinesin Superfamily Members as Prognostic Biomarkers of Breast Cancer. Cancer Cell Int. 2020, 20, 123. [Google Scholar] [CrossRef]
- Schneider, M.A.; Christopoulos, P.; Muley, T.; Warth, A.; Klingmueller, U.; Thomas, M.; Herth, F.J.F.; Dienemann, H.; Mueller, N.S.; Theis, F.; et al. AURKA, DLGAP5, TPX2, KIF11 and CKAP5: Five Specific Mitosis-Associated Genes Correlate with Poor Prognosis for Non-Small Cell Lung Cancer Patients. Int. J. Oncol. 2017, 50, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing Protein Abundance and MRNA Expression Levels on a Genomic Scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef]
- Imai, T.; Oue, N.; Nishioka, M.; Mukai, S.; Oshima, T.; Sakamoto, N.; Sentani, K.; Matsusaki, K.; Yoshida, K.; Yasui, W. Overexpression of KIF11 in Gastric Cancer with Intestinal Mucin Phenotype. Pathobiology 2017, 84, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Singel, S.M.; Cornelius, C.; Zaganjor, E.; Batten, K.; Sarode, V.R.; Buckley, D.L.; Peng, Y.; John, G.B.; Li, H.C.; Sadeghi, N.; et al. KIF14 Promotes AKT Phosphorylation and Contributes to Chemoresistance in Triple-Negative Breast Cancer. Neoplasia 2014, 16, 247–256.e2. [Google Scholar] [CrossRef]
- Carleton, M.; Mao, M.; Biery, M.; Warrener, P.; Kim, S.; Buser, C.; Marshall, C.G.; Fernandes, C.; Annis, J.; Linsley, P.S. RNA Interference-Mediated Silencing of Mitotic Kinesin KIF14 Disrupts Cell Cycle Progression and Induces Cytokinesis Failure. Mol. Cell. Biol. 2006, 26, 3853–3863. [Google Scholar] [CrossRef]
- Nakagawa, T.; Tanaka, Y.; Matsuoka, E.; Kondo, S.; Okada, Y.; Noda, Y.; Kanai, Y.; Hirokawa, N. Identification and Classification of 16 New Kinesin Superfamily (KIF) Proteins in Mouse Genome. Proc. Natl. Acad. Sci. USA 1997, 94, 9654–9659. [Google Scholar] [CrossRef]
- Yang, Z.; Li, C.; Yan, C.; Li, J.; Yan, M.; Liu, B.; Zhu, Z.; Wu, Y.; Gu, Q. KIF14 Promotes Tumor Progression and Metastasis and Is an Independent Predictor of Poor Prognosis in Human Gastric Cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, Y.; Liang, P.; Zhang, Z.; Guo, X.; Xia, L.; Zhao, Y.; Shu, X.-S.; Sun, S.; Ying, Y.; et al. Overexpression of a Novel Candidate Oncogene KIF14 Correlates with Tumor Progression and Poor Prognosis in Prostate Cancer. Oncotarget 2017, 8, 45459–45469. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, L.; Li, D.; Deng, J.; Zhao, Z.; He, S.; Zhang, Y.; Tu, Y. Kinesin Family Member 14 Is a Candidate Prognostic Marker for Outcome of Glioma Patients. Cancer Epidemiol. 2013, 37, 79–84. [Google Scholar] [CrossRef]
- Qiu, H.-L.; Deng, S.-Z.; Li, C.; Tian, Z.-N.; Song, X.-Q.; Yao, G.-D.; Geng, J.-S. High Expression of KIF14 Is Associated with Poor Prognosis in Patients with Epithelial Ovarian Cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 239–245. [Google Scholar] [PubMed]
- Hung, P.-F.; Hong, T.-M.; Hsu, Y.-C.; Chen, H.-Y.; Chang, Y.-L.; Wu, C.-T.; Chang, G.-C.; Jou, Y.-S.; Pan, S.-H.; Yang, P.-C. The Motor Protein KIF14 Inhibits Tumor Growth and Cancer Metastasis in Lung Adenocarcinoma. PLoS ONE 2013, 8, e61664. [Google Scholar] [CrossRef]
- Corson, T.W.; Gallie, B.L. KIF14 MRNA Expression Is a Predictor of Grade and Outcome in Breast Cancer. Int. J. Cancer 2006, 119, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Corson, T.W.; Zhu, C.Q.; Lau, S.K.; Shepherd, F.A.; Tsao, M.-S.; Gallie, B.L. KIF14 Messenger RNA Expression Is Independently Prognostic for Outcome in Lung Cancer. Clin. Cancer Res. 2007, 13, 3229–3234. [Google Scholar] [CrossRef]
- Thériault, B.L.; Pajovic, S.; Bernardini, M.Q.; Shaw, P.A.; Gallie, B.L. Kinesin Family Member 14: An Independent Prognostic Marker and Potential Therapeutic Target for Ovarian Cancer. Int. J. Cancer 2012, 130, 1844–1854. [Google Scholar] [CrossRef]
- Li, X.; Huang, W.; Huang, W.; Wei, T.; Zhu, W.; Chen, G.; Zhang, J. Kinesin Family Members KIF2C/4A/10/11/14/18B/20A/23 Predict Poor Prognosis and Promote Cell Proliferation in Hepatocellular Carcinoma. Am. J. Transl. Res. 2020, 12, 1614–1639. [Google Scholar]
- Wang, Z.-Z.; Yang, J.; Jiang, B.-H.; Di, J.-B.; Gao, P.; Peng, L.; Su, X.-Q. KIF14 Promotes Cell Proliferation via Activation of Akt and Is Directly Targeted by MiR-200c in Colorectal Cancer. Int. J. Oncol. 2018, 53, 1939–1952. [Google Scholar] [CrossRef]
- Hanicinec, V.; Brynychova, V.; Rosendorf, J.; Palek, R.; Liska, V.; Oliverius, M.; Kala, Z.; Mohelnikova-Duchonova, B.; Krus, I.; Soucek, P. Gene Expression of Cytokinesis Regulators PRC1, KIF14 and CIT Has No Prognostic Role in Colorectal and Pancreatic Cancer. Oncol. Lett. 2021, 22, 598. [Google Scholar] [CrossRef]
- Kendrick, N. A Gene’s MRNA Level Does Not Usually Predict Its Protein Level. Available online: https://kendricklabs.com/wp-content/uploads/2016/08/WP1_mRNAvsProtein_KendrickLabs.pdf (accessed on 19 April 2021).
- Vogel, C.; Marcotte, E.M. Insights into the Regulation of Protein Abundance from Proteomic and Transcriptomic Analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Wang, D. Discrepancy between MRNA and Protein Abundance: Insight from Information Retrieval Process in Computers. Comput. Biol. Chem. 2008, 32, 462–468. [Google Scholar] [CrossRef]
- Aviner, R.; Shenoy, A.; Elroy-Stein, O.; Geiger, T. Uncovering Hidden Layers of Cell Cycle Regulation through Integrative Multi-Omic Analysis. PLoS Genet. 2015, 11, e1005554. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, J.; Wang, X.; Zhu, J.; Liu, Q.; Shi, Z.; Chambers, M.C.; Zimmerman, L.J.; Shaddox, K.F.; Kim, S.; et al. Proteogenomic Characterization of Human Colon and Rectal Cancer. Nature 2014, 513, 382–387. [Google Scholar] [CrossRef]
- Liu, M.; Wang, X.; Yang, Y.; Li, D.; Ren, H.; Zhu, Q.; Chen, Q.; Han, S.; Hao, J.; Zhou, J. Ectopic Expression of the Microtubule-Dependent Motor Protein Eg5 Promotes Pancreatic Tumourigenesis. J. Pathol. 2010, 221, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Asbaghi, Y.; Thompson, L.L.; Lichtensztejn, Z.; McManus, K.J. KIF11 Silencing and Inhibition Induces Chromosome Instability That May Contribute to Cancer. Genes Chromosomes Cancer 2017, 56, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Soca-Chafre, G.; Montiel-Dávalos, A.; Rosa-Velázquez, I.A.D.L.; Caro-Sánchez, C.H.S.; Peña-Nieves, A.; Arrieta, O. Multiple Molecular Targets Associated with Genomic Instability in Lung Cancer. Int. J. Genom. 2019, 2019, 9584504. [Google Scholar] [CrossRef]
- Kolinjivadi, A.M.; Chong, S.T.; Ngeow, J. Molecular Connections between Circadian Rhythm and Genome Maintenance Pathways. Endocr. Relat. Cancer 2021, 28, R55–R66. [Google Scholar] [CrossRef] [PubMed]
- Kautto, E.A.; Bonneville, R.; Miya, J.; Yu, L.; Krook, M.A.; Reeser, J.W.; Roychowdhury, S. Performance Evaluation for Rapid Detection of Pan-Cancer Microsatellite Instability with MANTIS. Oncotarget 2017, 8, 7452–7463. [Google Scholar] [CrossRef]
- Niu, B.; Ye, K.; Zhang, Q.; Lu, C.; Xie, M.; McLellan, M.D.; Wendl, M.C.; Ding, L. MSIsensor: Microsatellite Instability Detection Using Paired Tumor-Normal Sequence Data. Bioinformatics 2014, 30, 1015–1016. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, U.; Dinh, T.A.; Alani, E. Genomic Instability Promoted by Overexpression of Mismatch Repair Factors in Yeast: A Model for Understanding Cancer Progression. Genetics 2018, 209, 439–456. [Google Scholar] [CrossRef]
- Wilczak, W.; Rashed, S.; Hube-Magg, C.; Kluth, M.; Simon, R.; Büscheck, F.; Clauditz, T.S.; Grupp, K.; Minner, S.; Tsourlakis, M.C.; et al. Up-Regulation of Mismatch Repair Genes MSH6, PMS2 and MLH1 Parallels Development of Genetic Instability and Is Linked to Tumor Aggressiveness and Early PSA Recurrence in Prostate Cancer. Carcinogenesis 2017, 38, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Norris, A.M.; Woodruff, R.D.; D’Agostino, R.B.; Clodfelter, J.E.; Scarpinato, K.D. Elevated Levels of the Mismatch Repair Protein PMS2 Are Associated with Prostate Cancer. Prostate 2007, 67, 214–225. [Google Scholar] [CrossRef]
- Biermann, J.; Nemes, S.; Parris, T.Z.; Engqvist, H.; Werner Rönnerman, E.; Kovács, A.; Karlsson, P.; Helou, K. A 17-Marker Panel for Global Genomic Instability in Breast Cancer. Genomics 2020, 112, 1151–1161. [Google Scholar] [CrossRef]
- Madhavan, J.; Coral, K.; Mallikarjuna, K.; Corson, T.W.; Amit, N.; Khetan, V.; George, R.; Biswas, J.; Gallie, B.L.; Kumaramanickavel, G. High Expression of KIF14 in Retinoblastoma: Association with Older Age at Diagnosis. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4901–4906. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.J.; Lu, A.; Moon, C. Role of Genomic Instability in Human Carcinogenesis. Exp. Biol. Med. 2019, 244, 227–240. [Google Scholar] [CrossRef]
- Weaver, B.A.A.; Silk, A.D.; Montagna, C.; Verdier-Pinard, P.; Cleveland, D.W. Aneuploidy Acts Both Oncogenically and as a Tumor Suppressor. Cancer Cell 2007, 11, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.S.; Holland, A.J. The Impact of Mitotic Errors on Cell Proliferation and Tumorigenesis. Genes Dev. 2018, 32, 620–638. [Google Scholar] [CrossRef]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The Eighth Edition AJCC Cancer Staging Manual: Continuing to Build a Bridge from a Population-Based to a More “Personalized” Approach to Cancer Staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Gzil, A.; Zarębska, I.; Jaworski, D.; Antosik, P.; Durślewicz, J.; Maciejewska, J.; Domanowska, E.; Skoczylas-Makowska, N.; Ahmadi, N.; Grzanka, D.; et al. The Prognostic Value of Leucine-Rich Repeat-Containing G-Protein (Lgr5) and Its Impact on Clinicopathological Features of Colorectal Cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 2547–2557. [Google Scholar] [CrossRef]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and Interpreting Cancer Genomics Data via the Xena Platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING Database in 2017: Quality-Controlled Protein-Protein Association Networks, Made Broadly Accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Feature | n (%) n = 86 | KIF11 Expression | p Value | KIF14 Expression | p Value | ||
|---|---|---|---|---|---|---|---|
| Low n = 61 | High n = 25 | Low n = 57 | High n = 29 | ||||
| Age (years) | |||||||
| ≤65 | 38 (44.19) | 29 (76.32) | 9 (23.68) | 0.35 | 27 (71.05) | 11 (28.95) | 0.49 |
| >65 | 48 (55.81) | 32 (66.67) | 16 (33.33) | 30 (62.50) | 18 (37.50) | ||
| Gender | |||||||
| Male | 49 (56.98) | 33 (67.35) | 16 (32.65) | 0.48 | 35 (71.43) | 14 (28.57) | 0.26 |
| Female | 37 (43.02) | 28 (75.68) | 9 (24.32) | 22 (59.46) | 15 (40.54) | ||
| Grading | |||||||
| G2 | 76 (91.57) | 54 (71.05) | 22 (28.95) | >0.99 | 50 (65.79) | 26 (34.21) | >0.99 |
| G3 | 7 (8.43) | 5 (71.43) | 2 (28.57) | 5 (71.43) | 2 (28.57) | ||
| pT status | |||||||
| T2 | 13 (15.12) | 10 (76.92) | 3 (23.08) | 0.67 | 9 (69.23) | 4 (30.77) | 0.68 |
| T3 | 60 (69.77) | 42 (70.00) | 18 (30.00) | 38 (63.33) | 22 (36.67) | ||
| T4 | 13 (15.12) | 9 (69.23) | 4 (30.77) | 10 (76.92) | 3 (23.08) | ||
| pN status | |||||||
| N0 | 33 (40.74) | 25 (75.76) | 8 (24.24) | 0.62 | 20 (60.61) | 13 (39.39) | 0.48 |
| N1-N2 | 48 (59.26) | 33 (68.75) | 15 (31.25) | 33 (68.75) | 15 (31.25) | ||
| pM status | |||||||
| M0 | 42 (52.50) | 29 (69.05) | 13 (30.95) | >0.99 | 27 (64.29) | 15 (35.71) | >0.99 |
| M1 | 38 (47.50) | 27 (71.05) | 11 (28.95) | 25 (65.79) | 13 (34.21) | ||
| VI | |||||||
| Absent | 24 (60.00) | 16 (66.67) | 8 (33.33) | 0.53 | 9 (37.50) | 15 (62.50) | 0.01 |
| Present | 16 (40.00) | 9 (56.25) | 7 (43.75) | 13 (81.25) | 3 (18.75) | ||
| PNI | |||||||
| Absent | 25 (89.29) | 14 (56.00) | 11 (44.00) | 0.26 | 12 (48.00) | 13 (52.00) | >0.99 |
| Present | 3 (10.71) | 3 (100.00) | 0 (0.00) | 2 (66.67) | 1 (33.33) | ||
| Variable | Univariate Cox | Multivariate Cox: KIF11(PS) and KIF14 | Multivariate Cox: KIF11(IRS)/KIF14 | Multivariate Cox: KIF11(PS)/KIF14 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HR | 95%CI | p | HR | 95%CI | p | HR | 95%CI | p | HR | 95%CI | p | |||||
| Lower | Upper | Lower | Upper | lower | upper | Lower | Upper | |||||||||
| KIF11 (IRS) | 1.51 | 0.79 | 2.87 | 0.21 | - | - | - | - | - | - | - | - | - | - | - | - |
| KIF11 (PS) | 2.17 | 1.14 | 4.12 | 0.02 | 2.41 | 1.10 | 5.28 | 0.03 | - | - | - | - | - | - | - | - |
| KIF14 | 0.54 | 0.26 | 1.10 | 0.09 | 0.43 | 0.17 | 1.11 | 0.08 | - | - | - | - | - | - | - | - |
| KIF11(IRS)low/KIF14high | Ref. | - | - | - | - | Ref. | - | - | - | - | ||||||
| KIF11(IRS)high/KIF14low | 3.33 | 1.29 | 8.55 | 0.01 | - | - | - | - | 3.91 | 1.13 | 13.54 | 0.03 | - | - | - | - |
| Others | 0.91 | 0.39 | 2.15 | 0.84 | - | - | - | - | 1.28 | 0.43 | 3.82 | 0.66 | - | - | - | - |
| KIF11(PS)low/KIF14high | Ref. | - | - | - | - | - | - | - | - | Ref. | ||||||
| KIF11(PS)high/KIF14low | 3.29 | 1.29 | 8.37 | 0.01 | - | - | - | - | - | - | - | - | 5.72 | 1.74 | 18.83 | 0.004 |
| Others | 2.18 | 0.92 | 5.13 | 0.08 | - | - | - | - | - | - | - | - | 2.58 | 0.92 | 7.19 | 0.07 |
| age | 1.00 | 0.97 | 1.03 | 0.83 | 1.02 | 0.99 | 1.06 | 0.18 | 1.01 | 0.98 | 1.05 | 0.48 | 1.02 | 0.99 | 1.06 | 0.17 |
| gender | 1.06 | 0.57 | 1.99 | 0.85 | 1.85 | 0.84 | 4.06 | 0.13 | 1.81 | 0.77 | 4.23 | 0.17 | 1.86 | 0.85 | 4.08 | 0.12 |
| grade | 2.31 | 0.80 | 6.64 | 0.12 | 3.62 | 1.14 | 11.53 | 0.03 | 3.15 | 0.99 | 9.99 | 0.05 | 3.68 | 1.15 | 11.76 | 0.03 |
| pT | 2.14 | 0.84 | 5.47 | 0.11 | 0.83 | 0.26 | 2.65 | 0.75 | 0.95 | 0.30 | 3.02 | 0.92 | 0.85 | 0.27 | 2.70 | 0.78 |
| pN | 1.77 | 0.90 | 3.48 | 0.10 | 1.57 | 0.70 | 3.54 | 0.28 | 1.16 | 0.47 | 2.84 | 0.75 | 1.58 | 0.70 | 3.55 | 0.27 |
| pM | 3.27 | 1.64 | 6.53 | 0.001 | 3.09 | 1.37 | 6.99 | 0.007 | 2.80 | 1.22 | 6.40 | 0.02 | 3.03 | 1.35 | 6.80 | 0.007 |
| Clinicopathological Feature | n (%) n = 277 | KIF11 Expression | p Value | KIF14 Expression | p Value | ||
|---|---|---|---|---|---|---|---|
| Low n = 244 | High n = 33 | Low n = 194 | High n = 83 | ||||
| Age (years) | |||||||
| ≤65 | 129 (46.91) | 114 (88.37) | 15 (11.63) | >0.99 | 91 (70.54) | 38 (29.46) | >0.99 |
| >65 | 146 (53.09) | 128 (87.67) | 18 (12.33) | 102 (69.86) | 44 (30.14) | ||
| Gender | |||||||
| Male | 150 (54.55) | 136 (90.67) | 14 (9.33) | 0.14 | 103 (68.67) | 47 (31.33) | 0.60 |
| Female | 125 (45.45) | 106 (84.80) | 19 (15.20) | 90 (72.00) | 35 (28.00) | ||
| pT status | |||||||
| T1 | 6 (2.18) | 6 (100.00) | 0 (0.00) | 0.36 | 4 (66.67) | 2 (33.33) | 0.34 |
| T2 | 43 (15.64) | 38 (88.37) | 5 (11.63) | 34 (79.07) | 9 (20.93) | ||
| T3 | 188 (68.36) | 166 (88.30) | 22 (11.70) | 132 (70.21) | 56 (29.79) | ||
| T4 | 38 (13.82) | 32 (84.21) | 6 (15.79) | 23 (60.53) | 15 (39.47) | ||
| pN status | |||||||
| N0 | 160 (58.18) | 143 (89.38) | 17 (10.63) | 0.45 | 120 (75.00) | 40 (25.00) | 0.045 |
| N1-N2 | 115 (41.82) | 99 (86.09) | 16 (13.91) | 73 (63.48) | 42 (36.52) | ||
| pM status | |||||||
| M0 | 185 (83.33) | 162 (87.57) | 23 (12.43) | 0.27 | 130 (70.27) | 55 (29.73) | 0.84 |
| M1 | 37 (16.67) | 35 (94.59) | 2 (5.41) | 25 (67.57) | 12 (32.43) | ||
| TNM stage | |||||||
| I-II | 150 (55.97) | 133 (88.67) | 17 (11.33) | 0.58 | 115 (76.67) | 35 (23.33) | 0.01 |
| III-IV | 118 (44.03) | 102 (86.44) | 16 (13.56) | 73 (61.86) | 45 (38.14) | ||
| Variable | Univariate Analysis | Multivariate Analysis: KIF11 | Multivariate Analysis: KIF14 | Multivariate Analysis: KIF11/KIF14 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HR | 95% CI | p | HR | 95%CI | p | HR | 95%CI | p | HR | 95%CI | p | |||||
| Lower | Upper | Lower | Upper | Lower | Upper | Lower | Upper | |||||||||
| KIF11 | 0.36 | 0.13 | 0.996 | 0.049 | 0.32 | 0.11 | 0.89 | 0.03 | - | - | - | - | - | - | - | - |
| KIF14 | 0.66 | 0.38 | 1.17 | 0.16 | - | - | - | - | 0.47 | 0.26 | 0.86 | 0.02 | - | - | - | - |
| KIF11low/KIF14low | Ref. | - | - | - | - | - | - | - | - | Ref. | ||||||
| KIF11high/KIF14high | 0.27 | 0.08 | 0.88 | 0.03 | - | - | - | - | - | - | - | - | 0.22 | 0.07 | 0.71 | 0.01 |
| Others | 1.10 | 0.60 | 1.99 | 0.77 | - | - | - | - | - | - | - | - | 0.74 | 0.39 | 1.41 | 0.36 |
| age | 1.02 | 0.998 | 1.04 | 0.08 | 1.03 | 1.004 | 1.05 | 0.02 | 1.03 | 1.01 | 1.05 | 0.004 | 1.03 | 1.01 | 1.05 | 0.01 |
| gender | 1.43 | 0.87 | 2.35 | 0.16 | 1.30 | 0.76 | 2.20 | 0.34 | 1.38 | 0.81 | 2.35 | 0.24 | 1.36 | 0.80 | 2.33 | 0.26 |
| pT | 3.21 | 1.17 | 8.84 | 0.02 | - | - | - | - | - | - | - | - | - | - | - | - |
| pN | 2.46 | 1.50 | 4.04 | <0.0001 | - | - | - | - | - | - | - | - | - | - | - | - |
| pM | 4.21 | 2.34 | 7.56 | <0.0001 | - | - | - | - | - | - | - | - | - | - | - | - |
| stage | 2.60 | 1.55 | 4.35 | <0.0001 | 3.37 | 1.96 | 5.77 | <0.0001 | 3.83 | 2.18 | 6.73 | <0.0001 | 3.65 | 2.08 | 6.41 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neska-Długosz, I.; Buchholz, K.; Durślewicz, J.; Gagat, M.; Grzanka, D.; Tojek, K.; Klimaszewska-Wiśniewska, A. Prognostic Impact and Functional Annotations of KIF11 and KIF14 Expression in Patients with Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 9732. https://doi.org/10.3390/ijms22189732
Neska-Długosz I, Buchholz K, Durślewicz J, Gagat M, Grzanka D, Tojek K, Klimaszewska-Wiśniewska A. Prognostic Impact and Functional Annotations of KIF11 and KIF14 Expression in Patients with Colorectal Cancer. International Journal of Molecular Sciences. 2021; 22(18):9732. https://doi.org/10.3390/ijms22189732
Chicago/Turabian StyleNeska-Długosz, Izabela, Karolina Buchholz, Justyna Durślewicz, Maciej Gagat, Dariusz Grzanka, Krzysztof Tojek, and Anna Klimaszewska-Wiśniewska. 2021. "Prognostic Impact and Functional Annotations of KIF11 and KIF14 Expression in Patients with Colorectal Cancer" International Journal of Molecular Sciences 22, no. 18: 9732. https://doi.org/10.3390/ijms22189732
APA StyleNeska-Długosz, I., Buchholz, K., Durślewicz, J., Gagat, M., Grzanka, D., Tojek, K., & Klimaszewska-Wiśniewska, A. (2021). Prognostic Impact and Functional Annotations of KIF11 and KIF14 Expression in Patients with Colorectal Cancer. International Journal of Molecular Sciences, 22(18), 9732. https://doi.org/10.3390/ijms22189732