
Applications of Adductomics in Chemically Induced Adverse Outcomes and Major Emphasis on DNA Adductomics: A Pathbreaking Tool in Biomedical Research



 ,
, 
 , , ,
, , ,  , ,
, , 
Abstract
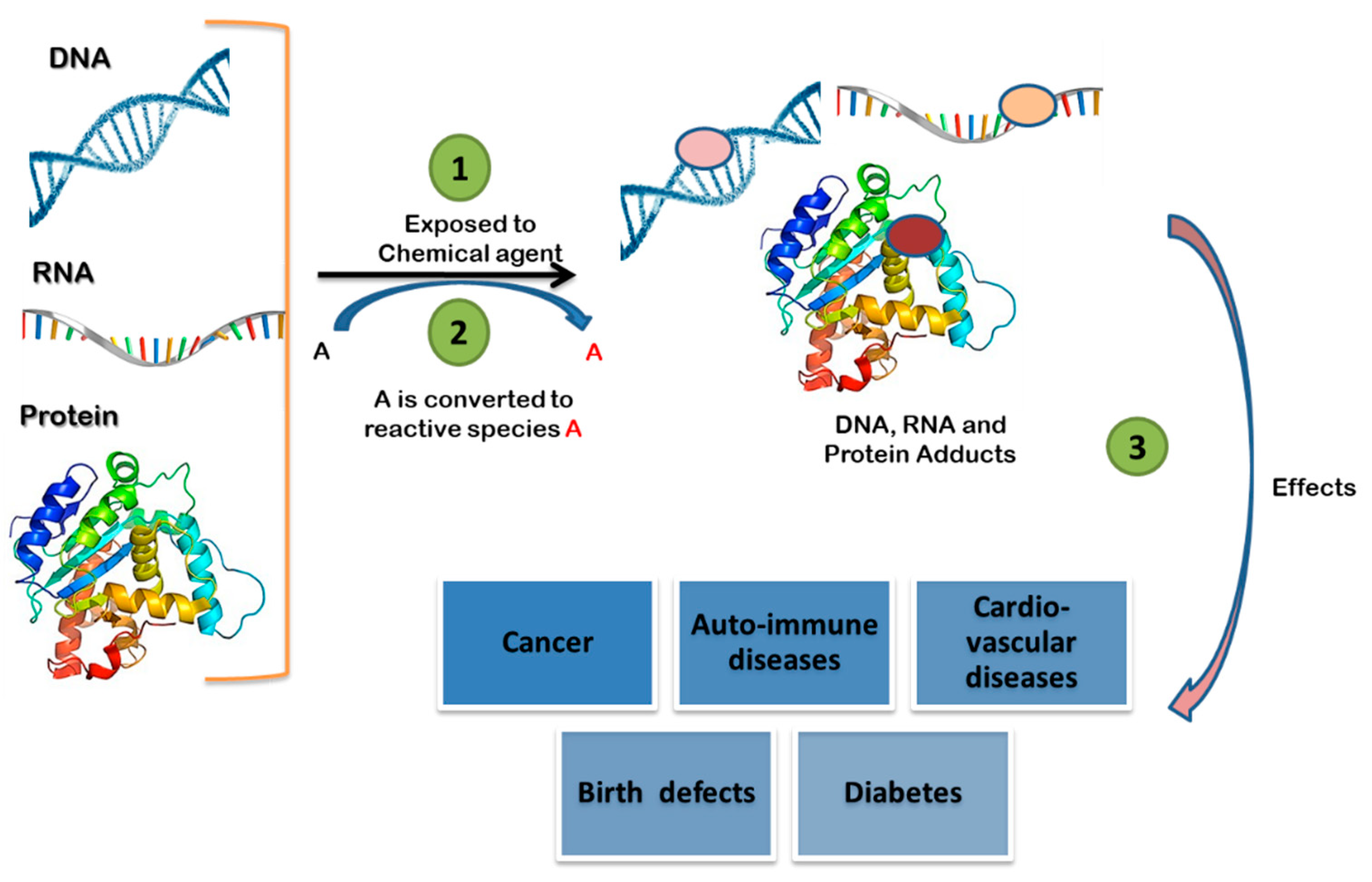
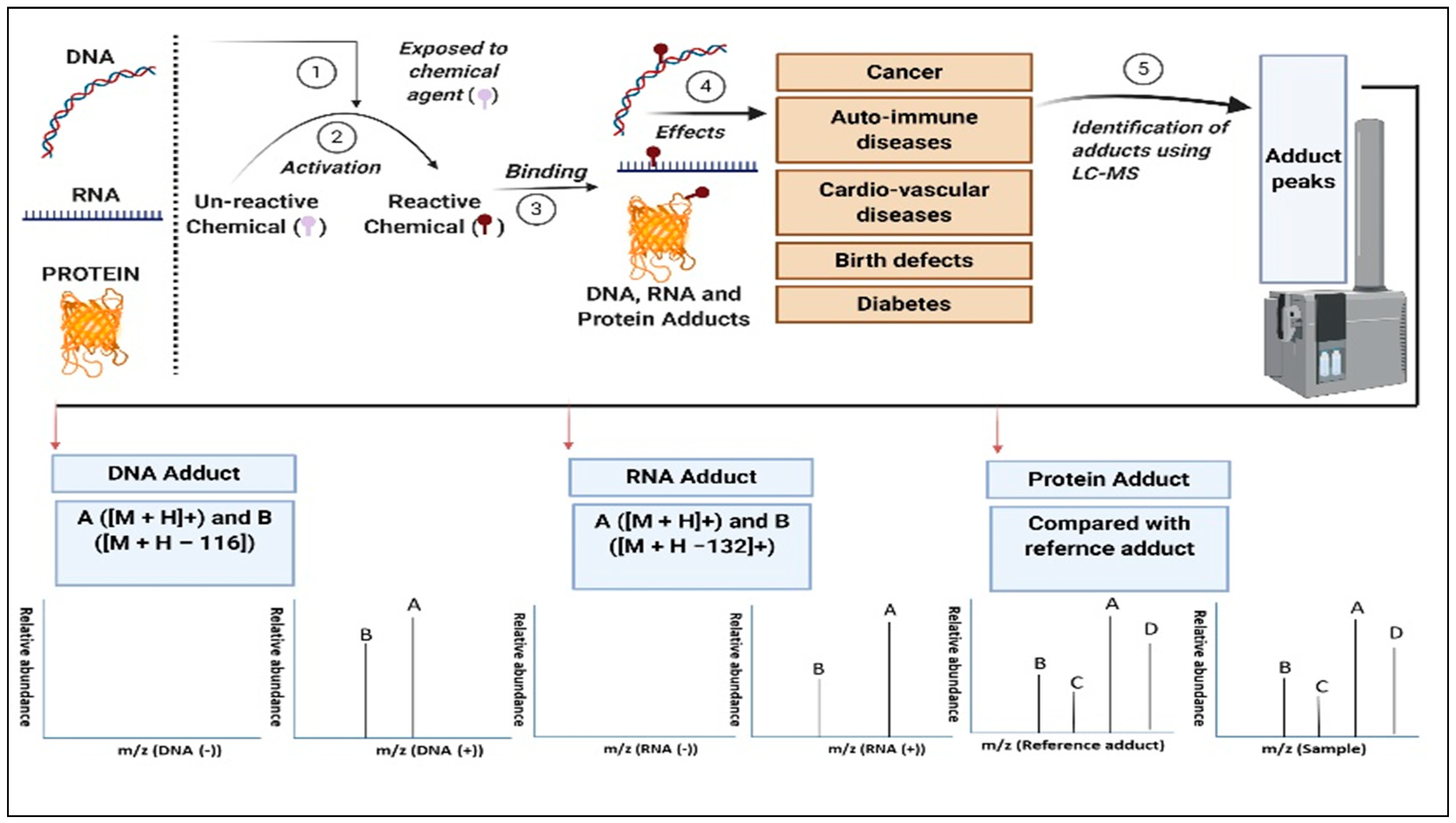
1. Introduction
2. Application of Adductomics
2.1. Adductomics and Disease
2.2. Adductomics and Pollution
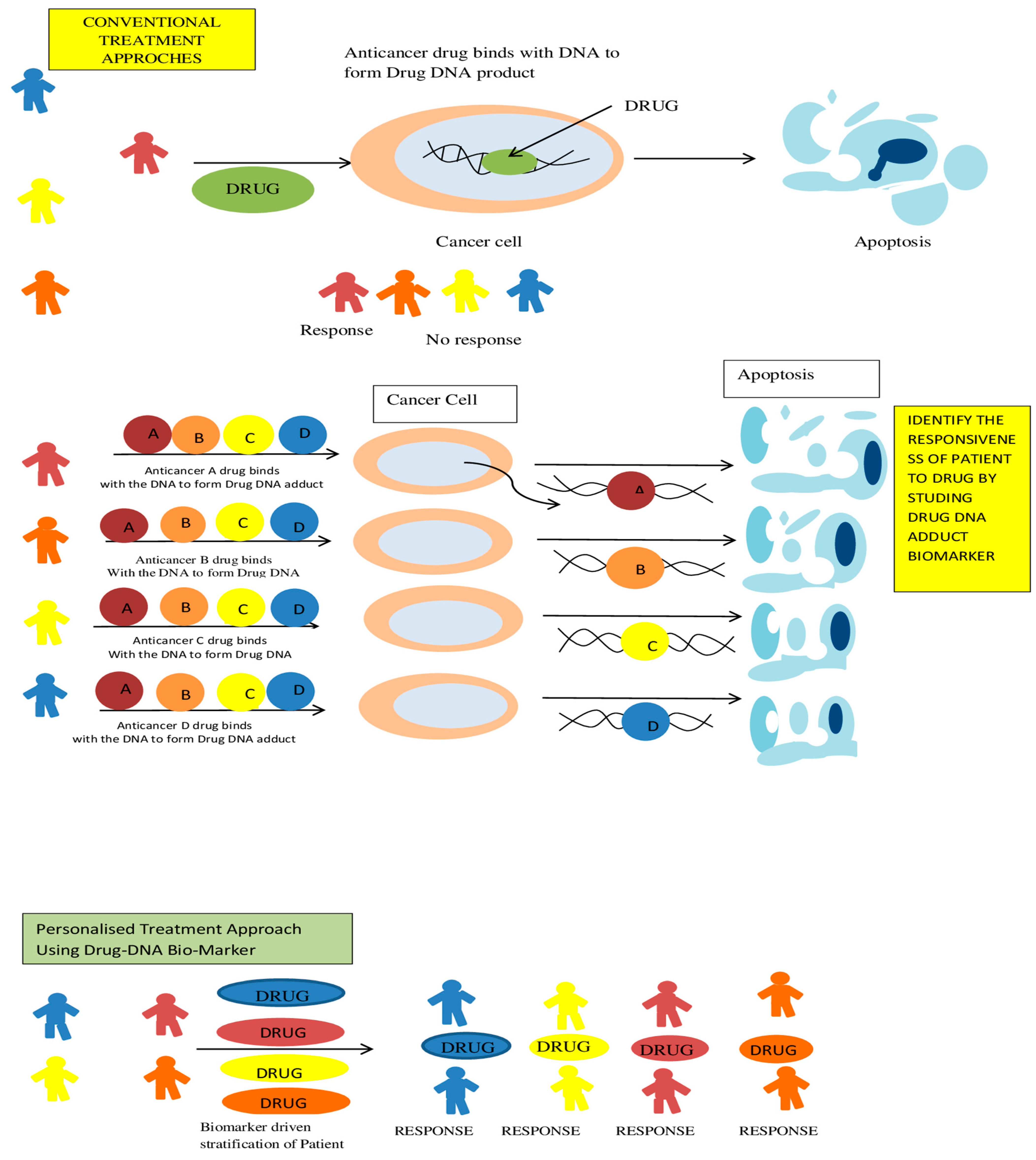
2.3. Adductomics in Precision Medicine in Cancer
2.3.1. Detecting DNA Adducts in Oral Cells as a Potential Biomarkers for Detecting Lung Cancer Progression in Smokers
2.3.2. Glycated Hemoglobin, Protein Adduct—A Diagnostic Tool for Diabetes and a Classic Example of the Application of Adductomics
2.4. Adductomics in Preventive Healthcare or Adductomics in Predictive and Prognosis of Diseases
2.5. Furan Toxicants and Adductomics
2.6. Adductomics in Designing Tools for Food Safety Assessment
2.6.1. Applications of Adductomics in the Verification of Chemical Warfare Agents in Biological Specimens
2.6.2. Environmental Adductomics—Linking DNA Adducts with Embryo Aberration in Baltic Amphipods
2.7. DNA Adductomics—A Confirmatory Tool in the Assessment of DNA Damage
2.8. Adductomics: Role in the Human Exposome Project (OR) EXPOSOMICS Project
3. Other Applications
4. Current Challenges and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Kanaly, R.A.; Hanaoka, T.; Sugimura, H.; Toda, H.; Matsui, S.; Matsuda, T. Development of the Adductome Approach to Detect DNA Damage in Humans. Antioxidants Redox Signal. 2006, 8, 993–1001. [Google Scholar] [CrossRef]
- Carlsson, H.; Rappaport, S.M.; Törnqvist, M. Protein Adductomics: Methodologies for Untargeted Screening of Adducts to Serum Albumin and Hemoglobin in Human Blood Samples. High-Throughput 2019, 8, 6. [Google Scholar] [CrossRef]
- Preston, G.W.; Phillips, D.H. Protein Adductomics: Analytical Developments and Applications in Human Biomonitoring. Toxics 2019, 7, 29. [Google Scholar] [CrossRef]
- Guengerich, F.P. Common and Uncommon Cytochrome P450 Reactions Related to Metabolism and Chemical Toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar] [CrossRef]
- Rendic, S.; Guengerich, F.P. Contributions of Human Enzymes in Carcinogen Metabolism. Chem. Res. Toxicol. 2012, 25, 1316–1383. [Google Scholar] [CrossRef] [PubMed]
- Beland, F.A.; Beranek, D.T.; Dooley, K.L.; Heflich, R.H.; Kadlubar, F.F. Arylamine-DNA adducts in vitro and in vivo: Their role in bacterial mutagenesis and urinary bladder carcinogenesis. Environ. Health Perspect. 1983, 49, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Shrivastav, N.; Li, D.; Essigmann, J.M. Chemical biology of mutagenesis and DNA repair: Cellular responses to DNA alkylation. Carcinogenesis 2009, 31, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Rajski, S.R.; Williams, R.M. DNA Cross-Linking Agents as Antitumor Drugs. Chem. Rev. 1998, 98, 2723–2796. [Google Scholar] [CrossRef]
- Marnett, L.J. Lipid peroxidation—DNA damage by malondialdehyde. Mutat. Res. Mol. Mech. Mutagen. 1999, 424, 83–95. [Google Scholar] [CrossRef]
- Tudek, B.; Zdżalik-Bielecka, D.; Tudek, A.; Kosicki, K.; Fabisiewicz, A.; Speina, E. Lipid peroxidation in face of DNA damage, DNA repair and other cellular processes. Free. Radic. Biol. Med. 2017, 107, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Marnett, L.J. Oxy radicals, lipid peroxidation and DNA damage. Toxicology 2002, 181, 219–222. [Google Scholar] [CrossRef]
- Guo, J.; Turesky, R.J. Emerging Technologies in Mass Spectrometry-Based DNA Adductomics. High-Throughput 2019, 8, 13. [Google Scholar] [CrossRef]
- Colombo, G.; Clerici, M.; Giustarini, D.; Rossi, R.; Milzani, A.D.G.; Dalle-Donne, I. Redox Albuminomics: Oxidized Albumin in Human Diseases. Antioxid. Redox Signal. 2012, 17, 1515–1527. [Google Scholar] [CrossRef]
- Gorokhova, E.; Martella, G.; Motwani, N.H.; Tretyakova, N.Y.; Sundelin, B.; Motwani, H.V. DNA epigenetic marks are linked to embryo aberrations in amphipods. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Zhang, H.; Ge, Y. Comprehensive Analysis of Protein Modifications by Top-Down Mass Spectrometry. Circ. Cardiovasc. Genet. 2011, 4, 711. [Google Scholar] [CrossRef]
- Stornetta, A.; Zimmermann, M.; Cimino, G.D.; Henderson, P.T.; Sturla, S.J. DNA Adducts from Anticancer Drugs as Candidate Predictive Markers for Precision Medicine. Chem. Res. Toxicol. 2017, 30, 388–409. [Google Scholar] [CrossRef]
- Harris, C.C. Future directions in the use of DNA adducts as internal dosimeters for monitoring human exposure to environmental mutagens and carcinogens. Environ. Heal. Perspect. 1985, 62, 185–191. [Google Scholar] [CrossRef] [PubMed]
- La, D.K.; Swenberg, J.A. DNA adducts: Biological markers of exposure and potential applications to risk assessment. Mutat. Res. Genet. Toxicol. 1996, 365, 129–146. [Google Scholar] [CrossRef]
- Jarabek, A.M.; Pottenger, L.H.; Andrews, L.S.; Casciano, D.; Embry, M.R.; Kim, J.H.; Preston, R.J.; Reddy, M.V.; Schoeny, R.; Shuker, D.; et al. Creating context for the use of DNA adduct data in cancer risk assessment: I. Data organization. Crit. Rev. Toxicol. 2009, 39, 659–678. [Google Scholar] [CrossRef] [PubMed]
- Balbo, S.; Turesky, R.J.; Villalta, P.W. DNA Adductomics. Chem. Res. Toxicol. 2014, 27, 356–366. [Google Scholar] [CrossRef]
- Takeshita, T.; Kanaly, R.A. In vitro DNA/RNA Adductomics to Confirm DNA Damage Caused by Benzo[a]pyrene in the Hep G2 Cell Line. Front. Chem. 2019, 7, 7. [Google Scholar] [CrossRef]
- Rappaport, S.M. Genetic Factors Are Not the Major Causes of Chronic Diseases. PLOS ONE 2016, 11, e0154387. [Google Scholar] [CrossRef]
- Lim, S.S.; Vos, T.; Flaxman, A.D.; Danaei, G.; Shibuya, K.; Adair-Rohani, H.; AlMazroa, M.A.; Amann, M.; Anderson, H.R.; Andrews, K.G.; et al. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2224–2260. [Google Scholar] [CrossRef]
- Hemminki, K.; Bermejo, J.L.; Försti, A. The balance between heritable and environmental aetiology of human disease. Nat. Rev. Genet. 2006, 7, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Powers, S.; Zhu, W.; Hannun, Y.A. Substantial contribution of extrinsic risk factors to cancer development. Nature 2016, 529, 43–47. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut Flora Metabolism of Phosphatidylcholine Promotes Cardiovascular Disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Ulrich, C.M.; Neuhouser, M.L.; Malysheva, O.; Bailey, L.B.; Xiao, L.; Brown, E.C.; Cushing-Haugen, K.L.; Zheng, Y.; Cheng, T.-Y.D.; et al. Plasma Choline Metabolites and Colorectal Cancer Risk in the Women’s Health Initiative Observational Study. Cancer Res. 2014, 74, 7442–7452. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, H.; Li, H.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Cys34 Adducts of Reactive Oxygen Species in Human Serum Albumin. Chem. Res. Toxicol. 2012, 25, 1633–1642. [Google Scholar] [CrossRef] [PubMed]
- Grimmer, V.G. Environmental Carcinogens: Polycyclic Aromatic Hydrocarbons; CRC Press: Boca Raton, FL, USA, 1983. [Google Scholar]
- Singh, R.; Teichert, F.; Seidel, A.; Roach, J.; Cordell, R.; Cheng, M.-K.; Frank, H.; Steward, W.P.; Manson, M.M.; Farmer, P.B. Development of a targeted adductomic method for the determination of polycyclic aromatic hydrocarbon DNA adducts using online column-switching liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2329–2340. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Marshall, W.D.; Frazer, D.G.; Law, B.; Lewis, D.M. Characterization of DNA adducts from lung tissue of asphalt fume-exposed mice by nanoflow liquid chromatography quadrupole time-of-flight mass spectrometry. Anal. Biochem. 2003, 322, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ewa, B.; Danuta, M.-. Švach Polycyclic aromatic hydrocarbons and PAH-related DNA adducts. J. Appl. Genet. 2017, 58, 321–330. [Google Scholar] [CrossRef]
- Whyatt, R.M.; Santella, R.M.; Jedrychowski, W.; Garte, S.J.; Bell, D.; Ottman, R.; Gladek-Yarborough, A.; Cosma, G.; Young, T.L.; Cooper, T.B.; et al. Relationship between ambient air pollution and DNA damage in Polish mothers and newborns. Environ. Heal. Perspect. 1998, 106, 821–826. [Google Scholar] [CrossRef]
- Palli, D.; Saieva, C.; Grechi, D.; Masala, G.; Zanna, I.; Barbaro, A.; Decarli, A.; Munnia, A.; Peluso, M. DNA bulky adducts in a Mediterranean population correlate with environmental ozone concentration, an indicator of photochemical smog. Int. J. Cancer 2004, 109, 17–23. [Google Scholar] [CrossRef]
- Hylland, K.; Skei, B.B.; Brunborg, G.; Lang, T.; Gubbins, M.J.; le Goff, J.; Burgeot, T. DNA damage in dab (Limanda limanda) and haddock (Melanogrammus aeglefinus) from European seas. Mar. Environ. Res. 2017, 124, 54–60. [Google Scholar] [CrossRef]
- A Hagger, J.; Jones, M.B.; Leonard, P.; Owen, R.; Galloway, T.S. Biomarkers and integrated environmental risk assessment: Are there more questions than answers? Integr. Environ. Assess. Manag. 2006, 2, 312–329. [Google Scholar] [CrossRef]
- Pampanin, D.M.; Brooks, S.J.; Grøsvik, B.E.; Le Goff, J.; Meier, S.; Sydnes, M.O. DNA adducts in marine fish as biological marker of genotoxicity in environmental monitoring: The way forward. Mar. Environ. Res. 2017, 125, 49–62. [Google Scholar] [CrossRef]
- Lyons, B.; Stewart, C.; Kirby, M. The detection of biomarkers of genotoxin exposure in the European flounder (Platichthys flesus) collected from the River Tyne Estuary. Mutat. Res. Toxicol. Environ. Mutagen. 1999, 446, 111–119. [Google Scholar] [CrossRef]
- Aas, E.; Baussant, T.; Balk, L.; Liewenborg, B.; Andersen, O.K. PAH metabolites in bile, cytochrome P4501A and DNA adducts as environmental risk parameters for chronic oil exposure: A laboratory experiment with Atlantic cod. Aquat. Toxicol. 2000, 51, 241–258. [Google Scholar] [CrossRef]
- Harvey, J.; Lyons, B.; Page, T.; Stewart, C.; Parry, J. An assessment of the genotoxic impact of the Sea Empress oil spill by the measurement of DNA adduct levels in selected invertebrate and vertebrate species. Mutat. Res. Toxicol. Environ. Mutagen. 1999, 441, 103–114. [Google Scholar] [CrossRef]
- Arlt, V.M.; Stiborova, M.; Henderson, C.J.; Thiemann, M.; Frei, E.; Svášková, D.; Singh, R.; Da Costa, G.G.; Schmitz, O.J.; Farmer, P.B.; et al. Metabolic activation of benzo[a]pyrene in vitro by hepatic cytochrome P450 contrasts with detoxification in vivo: Experiments with hepatic cytochrome P450 reductase null mice. Carcinogenesis 2007, 29, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.; Jarvis, I.W.; Phillips, D.H.; Arlt, V.M. Enhanced DNA adduct formation by benzo[a]pyrene in human liver cells lacking cytochrome P450 oxidoreductase. Mutat. Res. Toxicol. Environ. Mutagen. 2020, 852, 503162. [Google Scholar] [CrossRef]
- Park, J.W.; Kerbel, R.S.; Kelloff, G.J.; Barrett, J.C.; Chabner, B.A.; Parkinson, D.R.; Peck, J.; Ruddon, R.W.; Sigman, C.C.; Slamon, D.J. Rationale for Biomarkers and Surrogate End Points in Mechanism-Driven Oncology Drug Development. Clin. Cancer Res. 2004, 10, 3885–3896. [Google Scholar] [CrossRef][Green Version]
- Cimino, G.D.; Pan, C.-X.; Henderson, P.T. Personalized medicine for targeted and platinum-based chemotherapy of lung and bladder cancer. Bioanalysis 2013, 5, 369–391. [Google Scholar] [CrossRef]
- Poirier, M.C.; Lippard, S.J.; Zwelling, L.A.; Ushay, H.M.; Kerrigan, D.; Thill, C.C.; Santella, R.M.; Grunberger, D.; Yuspa, S.H. Antibodies elicited against cis-diamminedichloroplatinum(II)-modified DNA are specific for cis-diamminedichloroplatinum(II)-DNA adducts formed in vivo and in vitro. Proc. Natl. Acad. Sci. 1982, 79, 6443–6447. [Google Scholar] [CrossRef] [PubMed]
- Poirier, M.C.; Reed, E.; A Zwelling, L.; Ozols, R.F.; Litterst, C.L.; Yuspa, S.H. Polyclonal antibodies to quantitate cis-diamminedichloroplatinum(II)--DNA adducts in cancer patients and animal models. Environ. Heal. Perspect. 1985, 62, 89–94. [Google Scholar] [CrossRef]
- Hoebers, F.J.P.; Pluim, D.; Hart, A.A.M.; Verheij, M.; Balm, A.J.M.; Fons, G.; Rasch, C.R.N.; Schellens, J.H.M.; Stalpers, L.J.A.; Bartelink, H.; et al. Cisplatin-DNA adduct formation in patients treated with cisplatin-based chemoradiation: Lack of correlation between normal tissues and primary tumor. Cancer Chemother. Pharmacol. 2008, 61, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Reed, E.; Ozols, R.F.; Tarone, R.; Yuspa, S.H.; Poirier, M.C. Platinum-DNA adducts in leukocyte DNA correlate with disease response in ovarian cancer patients receiving platinum-based chemotherapy. Proc. Natl. Acad. Sci. 1987, 84, 5024–5028. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-J.; Kuo, C.-C.; Pan, H.-Y.; Tsou, T.-C.; Yeh, S.-C.; Chang, J.-Y. Mechanistic basis of a combination d-penicillamine and platinum drugs synergistically inhibits tumor growth in oxaliplatin-resistant human cervical cancer cells in vitro and in vivo. Biochem. Pharmacol. 2015, 95, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Li, T.; Semrad, T.J.; Wu, C.-Y.; Yu, A.; Cimino, G.; Malfatti, M.; Haack, K.; Turteltaub, K.W.; Pan, C.-X.; et al. Oxaliplatin–DNA Adducts as Predictive Biomarkers of FOLFOX Response in Colorectal Cancer: A Potential Treatment Optimization Strategy. Mol. Cancer Ther. 2020, 19, 1070–1079. [Google Scholar] [CrossRef] [PubMed]
- Hengstler, J.G.; Fuchs, J.; Oesch, F. DNA strand breaks and DNA cross-links in peripheral mononuclear blood cells of ovarian cancer patients during chemotherapy with cyclophosphamide/carboplatin. Cancer Res. 1992, 52, 52. [Google Scholar]
- Johnson, L.A.; Malayappan, B.; Tretyakova, N.; Campbell, C.; MacMillan, M.; Wagner, J.E.; Jacobson, P.A. Formation of cyclophosphamide specific DNA adducts in hematological diseases. Pediatr. Blood Cancer 2012, 58, 708–714. [Google Scholar] [CrossRef]
- Stornetta, A.; Villalta, P.W.; Gossner, F.; Wilson, W.R.; Balbo, S.; Sturla, S.J. DNA Adduct Profiles Predict in Vitro Cell Viability after Treatment with the Experimental Anticancer Prodrug PR104A. Chem. Res. Toxicol. 2017, 30, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Hecht, S.S. Oral Cell DNA Adducts as Potential Biomarkers for Lung Cancer Susceptibility in Cigarette Smokers. Chem. Res. Toxicol. 2017, 30, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Hukkanen, J.; Jacob, P., III; Benowitz, N.L. Metabolism and Disposition Kinetics of Nicotine. Pharmacol. Rev. 2005, 57, 79–115. [Google Scholar] [CrossRef] [PubMed]
- Carmella, S.G.; Ming, X.; Olvera, N.; Brookmeyer, C.; Yoder, A.; Hecht, S. High Throughput Liquid and Gas Chromatography–Tandem Mass Spectrometry Assays for Tobacco-Specific Nitrosamine and Polycyclic Aromatic Hydrocarbon Metabolites Associated with Lung Cancer in Smokers. Chem. Res. Toxicol. 2013, 26, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Carmella, S.G.; Chen, M.; Yagi, H.; Jerina, D.M.; Hecht, S.S. Analysis of phenanthrols in human urine by gas chromatography-mass spectrometry: Potential use in carcinogen metabolite phenotyping. Cancer Epidemiol. Biomarker. Prev. 2004, 13, 2167–2174. [Google Scholar]
- Carmella, S.G.; Chen, M.; Han, S.; Briggs, A.; Jensen, J.; Hatsukami, D.K.; Hecht, S.S. Effects of Smoking Cessation on Eight Urinary Tobacco Carcinogen and Toxicant Biomarkers. Chem. Res. Toxicol. 2009, 22, 734–741. [Google Scholar] [CrossRef]
- Carmella, S.G.; Chen, M.; Zarth, A.; Hecht, S.S. High throughput liquid chromatography–tandem mass spectrometry assay for mercapturic acids of acrolein and crotonaldehyde in cigarette smokers’ urine. J. Chromatogr. B 2013, 935, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Milne, G.L.; Yin, H.; Hardy, K.D.; Davies, S.S.; Roberts, I.L.J. Isoprostane Generation and Function. Chem. Rev. 2011, 111, 5973–5996. [Google Scholar] [CrossRef]
- Wang, D.; Dubois, R.N. Urinary PGE-M: A Promising Cancer Biomarker. Cancer Prev. Res. 2013, 6, 507–510. [Google Scholar] [CrossRef]
- John, H.; Van Der Schans, M.J.; Koller, M.; Spruit, H.E.T.; Worek, F.; Thiermann, H.; Noort, D. Fatal sarin poisoning in Syria 2013: Forensic verification within an international laboratory network. Forensic Toxicol. 2018, 36, 61–71. [Google Scholar] [CrossRef]
- Golime, R.; Palit, M.; Acharya, J.; Dubey, D.K. Neuroprotective Effects of Galantamine on Nerve Agent-Induced Neuroglial and Biochemical Changes. Neurotox. Res. 2017, 33, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Chai, P.R.; Boyer, E.W.; Al-Nahhas, H.; Erickson, T.B. Toxic chemical weapons of assassination and warfare: Nerve agents VX and sarin. Toxicol. Commun. 2017, 1, 21–23. [Google Scholar] [CrossRef]
- Golime, R.; Chandra, B.; Palit, M.; Dubey, D.K. Adductomics: A promising tool for the verification of chemical warfare agents’ exposures in biological samples. Arch. Toxicol. 2019, 93, 1473–1484. [Google Scholar] [CrossRef]
- Phillips, D.H.; Venitt, S. DNA and protein adducts in human tissues resulting from exposure to tobacco smoke. Int. J. Cancer 2012, 131, 2733–2753. [Google Scholar] [CrossRef]
- Nunes, J.; Charneira, C.; Morello, J.; Rodrigues, J.; Pereira, S.A.; Antunes, A.M.M. Mass Spectrometry-Based Methodologies for Targeted and Untargeted Identification of Protein Covalent Adducts (Adductomics): Current Status and Challenges. High-Throughput 2019, 8, 9. [Google Scholar] [CrossRef]
- Rahbar, S. The Discovery of Glycated Hemoglobin: A Major Event in the Study of Nonenzymatic Chemistry in Biological Systems. Ann. New York Acad. Sci. 2005, 1043, 9–19. [Google Scholar] [CrossRef]
- National Toxicology Program (NTP). Toxicology and Carcinogenesis Studies of Furan (CAS NO. 110-00-9) in F344/N Rats and B6C3F1 Mice (Gavage Studies); NTP TR 402 (NIH Publication No. 93-2857); U.S. Department of Health and Human Services, Public Health: Washington, DC, USA, 1993; pp. 1–287. [Google Scholar]
- Mariotti, M.S.; Granby, K.; Rozowski, J.; Pedreschi, F. Furan: A critical heat induced dietary contaminant. Food Funct. 2013, 4, 1001–1015. [Google Scholar] [CrossRef]
- Nunes, J.; Martins, I.L.; Charneira, C.; Pogribny, I.P.; de Conti, A.; Beland, F.; Marques, M.M.; Jacob, C.; Antunes, A.M. New insights into the molecular mechanisms of chemical carcinogenesis: In vivo adduction of histone H2B by a reactive metabolite of the chemical carcinogen furan. Toxicol. Lett. 2016, 264, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Gates, L.A.; Phillips, M.B.; Matter, B.A.; Peterson, L.A. Comparative Metabolism of Furan in Rodent and Human Cryopreserved Hepatocytes. Drug Metab. Dispos. 2014, 42, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Peterson, L.A. Identification of furan metabolites derived from Cysteine− cis-2-Butene-1, 4-dial—Lysine cross-links. Chem. Res. Toxicol. 2010, 23, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Kedderis, G.; Carfagna, M.; Held, S.; Batra, R.; Murphy, J.; Gargas, M. Kinetic Analysis of Furan Biotransformation by F-344 Rats in Vivo and in Vitro. Toxicol. Appl. Pharmacol. 1993, 123, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Schembri, F.; Zeskind, J.; Shah, V.; Gustafson, A.M.; Steiling, K.; Liu, G.; Dumas, Y.-M.; Zhang, X.; Brody, J.S.; et al. Smoking-induced gene expression changes in the bronchial airway are reflected in nasal and buccal epithelium. BMC Genom. 2008, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.O.; Gümüş, Z.; Kacker, A.; Choksi, V.L.; Bocker, J.M.; Zhou, X.K.; Yantiss, R.K.; Hughes, D.B.; Du, B.; Judson, B.L.; et al. Effects of Cigarette Smoke on the Human Oral Mucosal Transcriptome. Cancer Prev. Res. 2010, 3, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Spira, A. Upper airway gene expression in smokers: The mouth as a “window to the soul” of lung carcinogenesis? Cancer Prev. Res. 2010, 3, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Tung, N.T.; Tue, P.T.; Lien, T.T.N.; Ohno, Y.; Maehashi, K.; Matsumoto, K.; Nishigaki, K.; Biyani, M.; Takamura, Y. Peptide aptamer-modified single-walled carbon nanotube-based transistors for high-performance biosensors. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Minami, T.; Esipenko, N.A.; Zhang, B.; Isaacs, L.; Anzenbacher, P. “Turn-on” fluorescent sensor array for basic amino acids in water. Chem. Commun. 2014, 50, 61–63. [Google Scholar] [CrossRef]
- Amat-Bronnert, A.; Burgeot, T.; Castegnaro, M.; Pfohl-Leszkowicz, A. DNA adducts in fish following an oil spill exposure. Environ. Chem. Lett. 2006, 4, 93–99. [Google Scholar] [CrossRef]
- Azizi, M.H.; Bahadori, M.; Azizi, F. Breakthrough discovery of HbA1c by Professor Samuel Rahbar in 1968. Arch. Iran. Med. 2013, 16, 743. [Google Scholar]
- Lauby-Secretan, B.; Straif, K.; Baan, R.; Grosse, Y.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens—Part E: Tobacco, areca nut, alcohol, coal smoke, and salted fish. Lancet Oncol. 2009, 10, 1033–1034. [Google Scholar] [CrossRef]
- Ngongang, A.D.; Duy, S.V.; Sauvé, S. Analysis of nine N-nitrosamines using liquid chromatography-accurate mass high resolution-mass spectrometry on a Q-Exactive instrument. Anal. Methods 2015, 7, 5748–5759. [Google Scholar] [CrossRef]
- Sam, M.S.; Lintang, H.O.; Sanagi, M.M.; Lee, S.L.; Yuliati, L. Mesoporous carbon nitride for adsorption and fluorescence sensor of N-nitrosopyrrolidine. Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 2014, 124, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Pesch, B.; Kendzia, B.; Gustavsson, P.; Jöckel, K.-H.; Johnen, G.; Pohlabeln, H.; Olsson, A.; Ahrens, W.; Gross, I.M.; Brüske, I.; et al. Cigarette smoking and lung cancer-relative risk estimates for the major histological types from a pooled analysis of case-control studies. Int. J. Cancer 2011, 131, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Agudo, A.; Bonet, C.; Travier, N.; González, C.A.; Vineis, P.; Bueno-De-Mesquita, H.B.; Trichopoulos, D.; Boffetta, P.; Clavel-Chapelon, F.; Boutron-Ruault, M.-C.; et al. Impact of Cigarette Smoking on Cancer Risk in the European Prospective Investigation into Cancer and Nutrition Study. J. Clin. Oncol. 2012, 30, 4550–4557. [Google Scholar] [CrossRef] [PubMed]
- Hecht, S.S. Tobacco carcinogens, their biomarkers and tobacco-induced cancer. Nat. Rev. Cancer 2003, 3, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Shin, H.-S. Determination of tobacco-specific nitrosamines in replacement liquids of electronic cigarettes by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2013, 1291, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Choi, N.R.; Kim, Y.P.; Ji, W.H.; Hwang, G.-S.; Ahn, Y.G. Identification and quantification of seven volatile n-nitrosamines in cosmetics using gas chromatography/chemical ionization–mass spectrometry coupled with head space-solid phase microextraction. Talanta 2016, 148, 69–74. [Google Scholar] [CrossRef]
- Aas, P. The Threat of Mid-Spectrum Chemical Warfare Agents. Prehospital Disaster Med. 2003, 18, 306–312. [Google Scholar] [CrossRef]
- Organisation for the Prohibition of Chemical Weapons. Available online: https://www.opcw.org/ (accessed on 17 September 2021).
- Chai, P.R.; Hayes, B.D.; Erickson, T.B.; Boyer, E.W. Novichok agents: A historical, current, and toxicological perspective. Toxicol. Commun. 2018, 2, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Vale, J.A.; Marrs, T.C.; Maynard, R.L. Novichok: A murderous nerve agent attack in the UK. Clin. Toxicol. 2018, 56, 1093–1097. [Google Scholar] [CrossRef]
- Sundelin, B.; Eriksson, A. Malformations in embryos of the deposit-feeding amphipod Monoporeia affinis in the Baltic Sea. Mar. Ecol. Prog. Ser. 1998, 171, 165–180. [Google Scholar] [CrossRef]
- Venkatesan, A.K.; Pycke, B.F.G.; Halden, R.U. Detection and Occurrence of N-Nitrosamines in Archived Biosolids from the Targeted National Sewage Sludge Survey of the U.S. Environmental Protection Agency. Environ. Sci. Technol. 2014, 48, 5085–5092. [Google Scholar] [CrossRef]
- Jacobson, T.; Sundelin, B.; Yang, G.; Ford, A.T. Low dose TBT exposure decreases amphipod immunocompetence and reproductive fitness. Aquat. Toxicol. 2011, 101, 72–77. [Google Scholar] [CrossRef]
- Löf, M.; Sundelin, B.; Liewenborg, B.; Bandh, C.; Broeg, K.; Schatz, S.; Gorokhova, E. Biomarker-enhanced assessment of reproductive disorders in Monoporeia affinis exposed to contaminated sediment in the Baltic Sea. Ecol. Indic. 2016, 63, 187–195. [Google Scholar] [CrossRef]
- Löf, M.F.; Sundelin, B.; Bandh, C.; Gorokhova, E. Embryo aberrations in the amphipod Monoporeia affinis as indicators of toxic pollutants in sediments: A field evaluation. Ecol. Indic. 2016, 60, 18–30. [Google Scholar] [CrossRef]
- Cimino, M.C. Comparative overview of current international strategies and guidelines for genetic toxicology testing for regulatory purposes. Environ. Mol. Mutagen. 2006, 47, 362–390. [Google Scholar] [CrossRef] [PubMed]
- Thybaud, V.; Lorge, E.; Levy, D.; van Benthem, J.; Douglas, G.R.; Marchetti, F.; Moore, M.M.; Schoeny, R. Main issues addressed in the 2014-2015 revisions to the OECD Genetic Toxicology Test Guidelines. Environ. Mol. Mutagen. 2017, 58, 284–295. [Google Scholar] [CrossRef]
- Kirkland, D.; Aardema, M.; Henderson, L.; Müller, L. Evaluation of the ability of a battery of three in vitro genotoxicity tests to discriminate rodent carcinogens and non-carcinogens: I. Sensitivity, specificity and relative predictivity. Mutat. Res. Toxicol. Environ. Mutagen. 2005, 584, 1–256. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Honma, M.; Takahashi, M.; Horibe, A.; Tanaka, J.; Tsuchiya, M.; Morita, T. Identification and Evaluation of Potentially Genotoxic Agricultural and Food-related Chemicals. Food Saf. 2013, 1, 2013003. [Google Scholar] [CrossRef][Green Version]
- Kirkland, D.; Pfuhler, S.; Tweats, D.; Aardema, M.; Corvi, R.; Darroudi, F.; Elhajouji, A.; Glatt, H.; Hastwell, P.; Hayashi, M.; et al. How to reduce false positive results when undertaking in vitro genotoxicity testing and thus avoid unnecessary follow-up animal tests: Report of an ECVAM Workshop. Mutat. Res. Toxicol. Environ. Mutagen. 2007, 628, 31–55. [Google Scholar] [CrossRef]
- Sobus, J.R.; Wambaugh, J.; Isaacs, K.; Williams, A.; McEachran, A.; Richard, A.M.; Grulke, C.; Ulrich, E.; Rager, J.E.; Strynar, M.; et al. Integrating tools for non-targeted analysis research and chemical safety evaluations at the US EPA. J. Expo. Sci. Environ. Epidemiology 2018, 28, 411–426. [Google Scholar] [CrossRef]
- Kanaly, R.A.; Matsui, S.; Hanaoka, T.; Matsuda, T. Application of the adductome approach to assess intertissue DNA damage variations in human lung and esophagus. Mutat. Res. Mol. Mech. Mutagen. 2007, 625, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Yamamura, E.; Kawanishi, M.; Yagi, T.; Matsuda, T.; Sugiyama, A.; Uno, Y. Application of the DNA adductome approach to assess the DNA-damaging capability of in vitro micronucleus test-positive compounds. Mutat. Res. Toxicol. Environ. Mutagen. 2011, 721, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, S.M.; Smith, M.T. Environment and Disease Risks. Science 2010, 330, 460–461. [Google Scholar] [CrossRef] [PubMed]
- HEP. The Human Exposome Project: A Toolbox for Assessing and Addressing the Impact of Environment on Health. Available online: https://cordis.europa.eu/programme/id/H2020_SC1-BHC-28-2019 (accessed on 10 July 2021).
- Vineis, P.; Chadeau-Hyam, M.; Gmuender, H.; Gulliver, J.; Herceg, Z.; Kleinjans, J.; Kogevinas, M.; Kyrtopoulos, S.; Nieuwenhuijsen, M.; Phillips, D.; et al. The exposome in practice: Design of the EXPOsOMICS project. Int. J. Hyg. Environ. Heal. 2017, 220, 142–151. [Google Scholar] [CrossRef]
- The Human Exposome Project. Available online: https://humanexposomeproject.com/ (accessed on 17 September 2021).
- Marsillach, J.; Costa, L.G.; Furlong, C.E. Paraoxonase-1 and Early-Life Environmental Exposures. Ann. Glob. Health 2016, 82, 100–110. [Google Scholar] [CrossRef]
- Rappaport, S.M.; Li, H.; Grigoryan, H.; Funk, W.E.; Williams, E.R. Adductomics: Characterizing exposures to reactive electrophiles. Toxicol. Lett. 2012, 213, 83–90. [Google Scholar] [CrossRef]
- Aldini, G.; Vistoli, G.; Regazzoni, L.; Gamberoni, L.; Facino, R.M.; Yamaguchi, S.; Uchida, K.; Carini, M. Albumin Is the Main Nucleophilic Target of Human Plasma: A Protective Role Against Pro-atherogenic Electrophilic Reactive Carbonyl Species? Chem. Res. Toxicol. 2008, 21, 824–835. [Google Scholar] [CrossRef]
- Grigoryan, H.; Edmands, W.M.B.; Lan, Q.; Carlsson, H.; Vermeulen, R.; Zhang, L.; Yin, S.-N.; Li, G.-L.; Smith, M.T.; Rothman, N.; et al. Adductomic signatures of benzene exposure provide insights into cancer induction. Carcinogenesis 2018, 39, 661–668. [Google Scholar] [CrossRef]
- Lu, S.S.; Grigoryan, H.; Edmands, W.M.B.; Hu, W.; Iavarone, A.T.; Hubbard, A.; Rothman, N.; Vermeulen, R.; Lan, Q.; Rappaport, S.M. Profiling the Serum Albumin Cys34 Adductome of Solid Fuel Users in Xuanwei and Fuyuan, China. Environ. Sci. Technol. 2016, 51, 46–57. [Google Scholar] [CrossRef]
- Marsillach, J.; Costa, L.G.; Furlong, C.E. Protein adducts as biomarkers of exposure to organophosphorus compounds. Toxicol. 2013, 307, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Kanaly, R.A.; Micheletto, R.; Matsuda, T.; Utsuno, Y.; Ozeki, Y.; Hamamura, N. Application of DNA adductomics to soil bacterium Sphingobium sp. strain KK 22. Microbiologyopen 2015, 4, 841–856. [Google Scholar] [CrossRef]
- Cunliffe, M.; Kertesz, M.A. Autecological properties of soil sphingomonads involved in the degradation of polycyclic aromatic hydrocarbons. Appl. Microbiol. Biotechnol. 2006, 72, 1083–1089. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Kawasaki, A. Hydrocarbon-Degrading Sphingomonads: Sphingomonas, Sphingobium, Novosphingobium, and Sphingopyxis. Handbook of Hydrocarbon and Lipid Microbiology 2010, 1693–1705. [Google Scholar] [CrossRef]
- Gaskell, M.; McLuckie, K.I.; Farmer, P.B. Comparison of the mutagenic activity of the benzene metabolites, hydroquinone and para-benzoquinone in the supF forward mutation assay: A role for minor DNA adducts formed from hydroquinone in benzene mutagenicity. Mutat. Res. Mol. Mech. Mutagen. 2004, 554, 387–398. [Google Scholar] [CrossRef]
- Goldstein, B.D. Benzene toxicity: A critical evaluation: Hematotoxicity in humans. J. Toxicol. Environ. Health 1977, 2, 69–105. [Google Scholar]
- Huff, J.E.; Haseman, J.K.; DeMarini, D.M.; Eustis, S.; Maronpot, R.R.; Peters, A.C.; Persing, R.L.; Chrisp, C.E.; Jacobs, A.C. Multiple-site carcinogenicity of benzene in Fischer 344 rats and B6C3F1 mice. Environ. Health Perspect. 1989, 82, 125–163. [Google Scholar] [CrossRef]
- Snyder, R.; Witz, G.; Goldstein, B.D. The toxicology of benzene. Environ. Heal. Perspect. 1993, 100, 293–306. [Google Scholar] [CrossRef]
- Greenlee, W.F.; Sun, J.D.; Bus, J.S. A proposed mechanism of benzene toxicity: Formation of reactive intermediates from polyphenol metabolites. Toxicol. Appl. Pharmacol. 1981, 59, 187–195. [Google Scholar] [CrossRef]
- Wilson, M.R.; Jiang, Y.; Villalta, P.W.; Stornetta, A.; Boudreau, P.D.; Carrá, A.; Brennan, C.A.; Chun, E.; Ngo, L.; Samson, L.D.; et al. The human gut bacterial genotoxin colibactin alkylates DNA. Science 2019, 363, eaar7785. [Google Scholar] [CrossRef]
- Cooke, M.S.; Hu, C.-W.; Chang, Y.-J.; Chao, M.-R. Urinary DNA adductomics—A novel approach for exposomics. Environ. Int. 2018, 121, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Guina, T.; Biasi, F.; Calfapietra, S.; Nano, M.; Poli, G. Inflammatory and redox reactions in colorectal carcinogenesis. Ann. New York Acad. Sci. 2015, 1340, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, H.; Von Stedingk, H.; Nilsson, U.; Törnqvist, M. LC–MS/MS Screening Strategy for Unknown Adducts to N-Terminal Valine in Hemoglobin Applied to Smokers and Nonsmokers. Chem. Res. Toxicol. 2014, 27, 2062–2070. [Google Scholar] [CrossRef]
- Grigoryan, H.; Edmands, W.; Luca, R.; Yano, Y.; Regazzoni, L.; Iavarone, A.T.; Williams, E.R.; Rappaport, S.M. Adductomics Pipeline for Untargeted Analysis of Modifications to Cys34 of Human Serum Albumin. Anal. Chem. 2016, 88, 10504–10512. [Google Scholar] [CrossRef]
- Sabbioni, G.; Turesky, R.J. Biomonitoring Human Albumin Adducts: The Past, the Present, and the Future. Chem. Res. Toxicol. 2017, 30, 332–366. [Google Scholar] [CrossRef]
- Grigoryan, H.; Schiffman, C.; Gunter, M.J.; Naccarati, A.; Polidoro, S.; Dagnino, S.; Dudoit, S.; Vineis, P.; Rappaport, S.M. Cys34 Adductomics Links Colorectal Cancer with the Gut Microbiota and Redox Biology. Cancer Res. 2019, 79, 6024–6031. [Google Scholar] [CrossRef]
- Li, D.; Wang, M.; Dhingra, K.; Hittelman, W.N. Aromatic DNA adducts in adjacent tissues of breast cancer patients: Clues to breast cancer etiology. Cancer Res. 1996, 56, 287–293. [Google Scholar]
- Wang, M.; Abbruzzese, J.L.; Friess, H.; Hittelman, W.N.; Evans, D.B.; Abbruzzese, M.C.; Chiao, P.; Li, D. DNA adducts in human pancreatic tissues and their potential role in carcinogenesis. Cancer Res. 1998, 58, 38–41. [Google Scholar]
- Schut, H.A.; Snyderwine, E.G. DNA adducts of heterocyclic amine food mutagens: Implications for mutagenesis and carcinogenesis. Carcinogenesis 1999, 20, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, G.; Mattioli, F.; Martelli, A. Genotoxic and carcinogenic effects of gastrointestinal drugs. Mutagen. 2010, 25, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Vrijheid, M.; Slama, R.; Robinson, O.; Chatzi, L.; Coen, M.; Hazel, P.V.D.; Thomsen, C.; Wright, J.; Athersuch, T.J.; Avellana, N.; et al. The Human Early-Life Exposome (HELIX): Project Rationale and Design. Environ. Heal. Perspect. 2014, 122, 535–544. [Google Scholar] [CrossRef]
- Törnqvist, M.; Fred, C.; Haglund, J.; Helleberg, H.; Paulsson, B.; Rydberg, P. Protein adducts: Quantitative and qualitative aspects of their formation, analysis and applications. J. Chromatogr. B 2002, 778, 279–308. [Google Scholar] [CrossRef]
- Liebler, D.C. Proteomic approaches to characterize protein modifications: New tools to study the effects of environmental exposures. Environ. Heal. Perspect. 2002, 110, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Stepan, A.F.; Walker, D.P.; Bauman, J.; Price, D.A.; Baillie, T.A.; Kalgutkar, A.S.; Aleo, M.D. Structural Alert/Reactive Metabolite Concept as Applied in Medicinal Chemistry to Mitigate the Risk of Idiosyncratic Drug Toxicity: A Perspective Based on the Critical Examination of Trends in the Top 200 Drugs Marketed in the United States. Chem. Res. Toxicol. 2011, 24, 1345–1410. [Google Scholar] [CrossRef]
- Uetrecht, J.; Naisbitt, D.J. Idiosyncratic Adverse Drug Reactions: Current Concepts. Pharmacol. Rev. 2013, 65, 779–808. [Google Scholar] [CrossRef]
- Walmsley, S.; Guo, J.; Wang, J.; Villalta, P.W.; Turesky, R.J. Methods and Challenges for Computational Data Analysis for DNA Adductomics. Chem. Res. Toxicol. 2019, 32, 2156–2168. [Google Scholar] [CrossRef] [PubMed]
- Human Metabolome Database. Available online: https://hmdb.ca/ (accessed on 17 September 2021).
- Toxic Exposome Database, T3DB. Available online: http://www.t3db.ca/ (accessed on 10 July 2021).
- Neveu, V.; Moussy, A.; Rouaix, H.; Wedekind, R.; Pon, A.; Knox, C.; Wishart, D.S.; Scalbert, A. Exposome-Explorer: A manually-curated database on biomarkers of exposure to dietary and environmental factors. Nucleic Acids Res. 2017, 45, D979–D984. [Google Scholar] [CrossRef] [PubMed]
- Rattray, N.J.W.; DeZiel, N.C.; Wallach, J.D.; Khan, S.A.; Vasiliou, V.; Ioannidis, J.P.A.; Johnson, C.H. Beyond genomics: Understanding exposotypes through metabolomics. Hum. Genom. 2018, 12, 4. [Google Scholar] [CrossRef]
- Molecular Weight Search. Available online: http://webbook.nist.gov/chemistry/mw-ser.html (accessed on 10 July 2021).
- Unimod. Available online: http://www.unimod.org/modifications_list.php (accessed on 10 July 2021).




{kind=link}
{kind=link}
{kind=link}
| Research Areas and Application of Adductomics | Significant Outcomes of Adductomics Technology Application in Various Studies |
|---|---|
| Paraoxonase-1 and Early-Life Environmental Exposures—Application of adductomics in biomonitoring of organophosphates exposure using OP-adducted biomarker proteins [114] | Children and infants are vulnerable to Organophosphates, which are widely used pesticides in agriculture, because detoxification mechanisms are not developed to their fullest potential, that only happens several years after birth. Paraoxonase-1 (PON1) is a detoxification enzyme that neutralizes the toxic effects of organophosphates, and the status of PON1 in the mother determines the level of protection in the child against the adverse effects of organophosphates. Though PON1 status gives an account of the susceptibility to the organophosphates, analyzing OP-adducted biomarker proteins becomes handy in biomonitoring of the exposure trends. Moreover, using adductomics instead of enzymatic activity assays (cholinesterase enzymatic inhibition assay) also offers multiple fold of advantages, and the use of dried blood spots would suffice to monitor the pesticides in infants and mothers. |
| Cys34 blood adductomics method [115] | Cys 34 residue in Human Serum Albumin is highly reactive to several reactive electrophiles [116], and this non targeted approach is used to distinguish: smokers from non-smokers [117] and users of smoky coal from smokeless coal [118]. Cys 34 residues are also implicated in several adducts that form after exposure to multitude of hazardous exogenous agents. |
| Organophosphate-adducted serine esterase—Classical Biomarker of pesticide exposure [119] | Organophosphate-adducted serine esterases are among the most widely studied and known protein adducts employed in biomonitoring of pesticide exposure in humans. There is a growing interest protein biomarker-based identification of pesticide exposure because protein adducts have longer half-lives, and proteins are only modified by the active parent molecule but not by their breakdown products or metabolites. The cited reasons above ensure a realistic assessment of pesticide exposure with adductomics when compared with other methods of assessment [119]. |
| DNA adductomics application to soil bacterium Sphingobium sp. strain KK2 [120] | This was the first study to report DNA damage in bacteria and paved the path for the use of microbes in toxicity studies using biomarker adducts. Sphingomonad bacteria are renowned for their existence in polluted ambiance, especially in the areas contaminated with hazardous hydrocarbon pollutants [121,122]. Acrolein is a biocide used in the hydraulic fracturing process of hydrocarbon extraction, and bacteria are exposed to this pollutant. Later the DNA was extracted and analyzed for DNA adducts using positive ionization electron spray LC-MS in pre-set reaction monitoring mode transmitting the [M + H] +> [M + H−16] + transition over 100 transitions. Two exposure-specific adducts: 3-(2′-deoxyribosyl)-5,6,7,8-tetrahydro-6-hydroxyl PdG and 8-hydroxy-pyrimido [1,2-a]-purine-(3H)-one PdG (6-and 8-hydroxy-PdG) were selected. These 6-and 8-hydroxy-PdG putative adducts were specific to Acrolein, and they serve as potential biomarkers of Acrolein contamination in the environment. This study demonstrated the application of DNA adductomics using soil microbes to assess environmental contamination and develop compound-specific biomarkers for prognosis of pollutants [120]. |
| Comparing mutagenic activity of the benzene metabolites: hydroquinone (HQ) and para-benzoquinone (p-BQ) using supF forward mutation assay [123] | Benzene is a human leukemogenic and rodent carcinogen that is omnipresent in the atmosphere as a hazardous occupational chemical. Though the precise mechanism underneath the mutagenicity of benzene is not deciphered, it was assumed that benzene-DNA adducts play a crucial role in mutagenesis. Benzene is a neutral molecule, and it is catalyzed in the body to generate reactive electrophiles, and these reactive metabolites intercalate with DNA and proteins invoking detrimental effects. In this in-vitro study Hydroquinone [HQ] and Para-benzoquinone [p-BQ], from among the pool of electrophiles generated from benzene, react with DNA forming carcinogenic adducts (dGp adducts); p-BQ yielded four adducts while HQ yielded only one DNA adduct. Mutagenicity of these reactive metabolites is investigated using the supF forward mutation assay and screened using an indicator bacterium. Upon comparing the mutagenic potential of both the metabolites, it was revealed that 5–20 mM p-BQ treatment resulted in a 12–40-fold increase in mutation rate, whereas 5–20 mM HQ treatment resulted in an 8–26-fold increase in mutation rate, which signifies the greater mutagenic potential of the former that the later. The study revealed that dGp adducts formed by the benzene metabolites are instrumental in mutagenicity and myelotoxicity of benzene [123,124,125,126]. |
| Adenine-Colibactin adduct—Biomarker for Colonorectal Cancer [127] | Colibactin, polyketide/nonribosomal peptide produced, is produced by Escherichia coli that encompass biosynthetic gene islands called pks. Colibactin is a genotoxic secondary metabolite implicated in Colorectal cancer (CRC), which induces mutagenesis by forming, adducts culminating in the breaking of double-stranded DNA. Colibactin contains a reactive cyclopropane ring that reacts with DNA (adenine) to generate adenine-Colibactin adducts, and these adducts serve as biomarkers for pks + E. coli exposure, providing a forecast of the genesis of colorectal cancer [127]. |
| Adductomics Research Programme [128] | It is a flagship program of the National Cancer Institute (Division of Cancer Prevention) that aims at applying the principles of the adductomics to design strategies for cancer screening and prevention by identifying molecular signatures/biomarkers that bear a testament for risk exposure. Moreover, the program also laid emphasis on amalgamating adducts signatures with other molecular signatures (DNA repair defects and capacity to repair, genetic aberrations and alterations, imaging, role of non-coding RNAs in cancer risk) to present the true situation in the cell, which facilitates early cancer identification and risk prevention. The program also aids projects investigating the potential roles of adducts in gene-environment interaction research (GxE) and cancer etiology. |
| Urinary DNA adductomics [129] | DNA adductomics is an inalienable part of the exposome approach, and invasive sample collection methods (blood or tissue samples) are generally adopted to isolate the required DNA for sensitive analysis of DNA adducts. However, invasive tissue sampling to obtain sufficient DNA poses IRB and logistical challenges, particularly when investigating vulnerable populations. This study provided a novel approach of adductomics in biomonitoring environmental exposures in a non-invasive manner using a urine sample, easing the exposome’s evaluation across the life-course, which reflects the totality of body burden of adducts amassed overtime. |
| Cys34 Adductomics Links Colorectal Cancer with the Gut Microbiota and Redox Biology [130] | It is well very well known that chronic colonic inflammation is implicated in inflammatory bowel disease and colorectal cancer mediated through reactive oxygen species (ROS) and reactive carbon species (RCS) that alters biomolecules (proteins and DNA) and modulates redox signaling pathways. ROS and RCS, despite holding huge potential of predicting the risk of colorectal cancer, cannot be measured in-vivo and to overwhelm this barrier, untargeted adductomics approach was used to identify reactive electrophile products of human or microbial metabolism by monitoring adducts of those species that react with hemoglobin and Human Serum Albumin (HAS). In this study, the adductomics pipeline was developed to investigate amendments at the highly nucleophilic Cys34 residue in HAS and the rationale for the selection of Cys34 in HAS was for its involvement in scavenging reactive electrophiles. Using untargeted adductomics, the study was performed to diagnose adducts in pre-diagnostic serum collected from the EPIC Italy cohort. For the study, seven Cys34 adducts which are associated with progression of colorectal cancer, and body mass indexes (BMI) are carefully chosen as risk factors. 5 out of 7 adducts were identified predominant in colorectal cancer cases when compared with controls. In those five adducts, two of them were modified by methanethiol (a microbial–human cometabolite), and crotonaldehyde (a product of lipid peroxidation). This manifests inflammation, a precursor step to colorectal cancer, caused by gut microbiome infiltration into intestinal mucosa. It also demonstrated that selected adducts and BMI would serve as potential casual factors when compared to other variables such as processed meat consumption and smoking that were associated with progression of colorectal cancer previously. The study exposed the role of Cys34 adducts of HSA taking the role as biomarker for calculating the chances of onset of colorectal cancer [131,132,133,134,135]. |
| Biomarkers in epidemiology studies [17] | Several biomarkers (carcinogen-DNA adducts) pertinent to tumor initiation and progression are also currently under development. In addition to DNA, albumin also forms adduct aflatoxin B1 which is human carcinogen which were successfully explored as biomarkers in molecular epidemiology studies. Other examples such as Benzo(a)pyrene (BP)-like DNA adduct biomarker gives clues regarding breast cancer etiology and Putative lipid peroxidation-related DNA adducts role in forecasting pancreatic cancer is also explored. The DNA adducts formed from specific chemicals are used as biomarkers in molecular epidemiology and cancer prevention studies. Benzene exposure is resulting in the induction of oxygen and carbonyl species adducts and Cys34 disulfides of small thiols in HSA, which serve as adductomics signatures offering significant insights into cancer [135,136,137,138,139,140,141]. |
| HELIX project and adductomics [140] | The HELIX project investigates the impact of early life exposure of chemical and physical agents in existing birth cohorts, while EXPOSOMICS project pays emphasis on the assessment of the effect of air pollution studied in an adult and child populations [109]. Both these projects involve two components: exposure monitoring using technology support and investigating biomarkers associated with exposures using a multiomics approach, and the second component include metabolome, proteome, transcriptome, epigenome and adductome profiling [110]. |
| Protein adductomics and its applications [135] | Biomonitoring of protein-adducts formed due to carcinogens is one of the superior and effective substitutes for the detection of DNA adducts for risk assessment owing to the abundance of proteins, easy detection, and stable adducts for longer duration [142,143,144]. Several proteins, such as albumin, hemoglobin, glutathione, and histones in the body, form adducts with reactive chemicals. Studying protein adducts also facilitates evaluating the quality of the environment as proteins in several biological systems possess nucleophilic reactive sites that easily form adducts with toxicants, and the extent and type of adduct provide us vital information pertinent to the quantity and category of pollutants prevalent in the environment. This is substantiated by the fact that some of the common industrial pollutants such as benzene, isocyanates, naphthalene, aromatic amines form adducts with albumin. Albumin, being the most abundant protein, forms adduct with so many very well-established therapeutic drugs or their reactive metabolites such as acetylsalicylic acid, non-steroidal anti-inflammatory drugs, acetaminophen, β-lactam antibiotics, antiretroviral therapy drugs and chemotherapeutic agents. Identifying and characterizing adduct structures formed by Alb with drug metabolites play a key role in understanding the generation of reactive metabolites and assisting in predicting idiosyncratic drug reactions toxicities [134,135]. The Alb-drug metabolite adducts are included in a series of screening tools and technologies in assessing potential toxicities of drugs, and it also aids in calculating the biologically effective dosage of drugs and developing personalized, and precision treatment approaches, which minimizes adverse effects of drugs. Several proteins adduct are becoming beneficial in predicting early and late biological effects of several chemicals and toxicants [145]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behl, T.; Rachamalla, M.; Najda, A.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Al-Harrasi, A.; Chigurupati, S.; Vargas-De-La-Cruz, C.; et al. Applications of Adductomics in Chemically Induced Adverse Outcomes and Major Emphasis on DNA Adductomics: A Pathbreaking Tool in Biomedical Research. Int. J. Mol. Sci. 2021, 22, 10141. https://doi.org/10.3390/ijms221810141
Behl T, Rachamalla M, Najda A, Sehgal A, Singh S, Sharma N, Bhatia S, Al-Harrasi A, Chigurupati S, Vargas-De-La-Cruz C, et al. Applications of Adductomics in Chemically Induced Adverse Outcomes and Major Emphasis on DNA Adductomics: A Pathbreaking Tool in Biomedical Research. International Journal of Molecular Sciences. 2021; 22(18):10141. https://doi.org/10.3390/ijms221810141
Chicago/Turabian StyleBehl, Tapan, Mahesh Rachamalla, Agnieszka Najda, Aayush Sehgal, Sukhbir Singh, Neelam Sharma, Saurabh Bhatia, Ahmed Al-Harrasi, Sridevi Chigurupati, Celia Vargas-De-La-Cruz, and et al. 2021. "Applications of Adductomics in Chemically Induced Adverse Outcomes and Major Emphasis on DNA Adductomics: A Pathbreaking Tool in Biomedical Research" International Journal of Molecular Sciences 22, no. 18: 10141. https://doi.org/10.3390/ijms221810141
APA StyleBehl, T., Rachamalla, M., Najda, A., Sehgal, A., Singh, S., Sharma, N., Bhatia, S., Al-Harrasi, A., Chigurupati, S., Vargas-De-La-Cruz, C., Hobani, Y. H., Mohan, S., Goyal, A., Katyal, T., Solarska, E., & Bungau, S. (2021). Applications of Adductomics in Chemically Induced Adverse Outcomes and Major Emphasis on DNA Adductomics: A Pathbreaking Tool in Biomedical Research. International Journal of Molecular Sciences, 22(18), 10141. https://doi.org/10.3390/ijms221810141