
Does Coenzyme Q10 Supplementation Improve Human Oocyte Quality?



Abstract
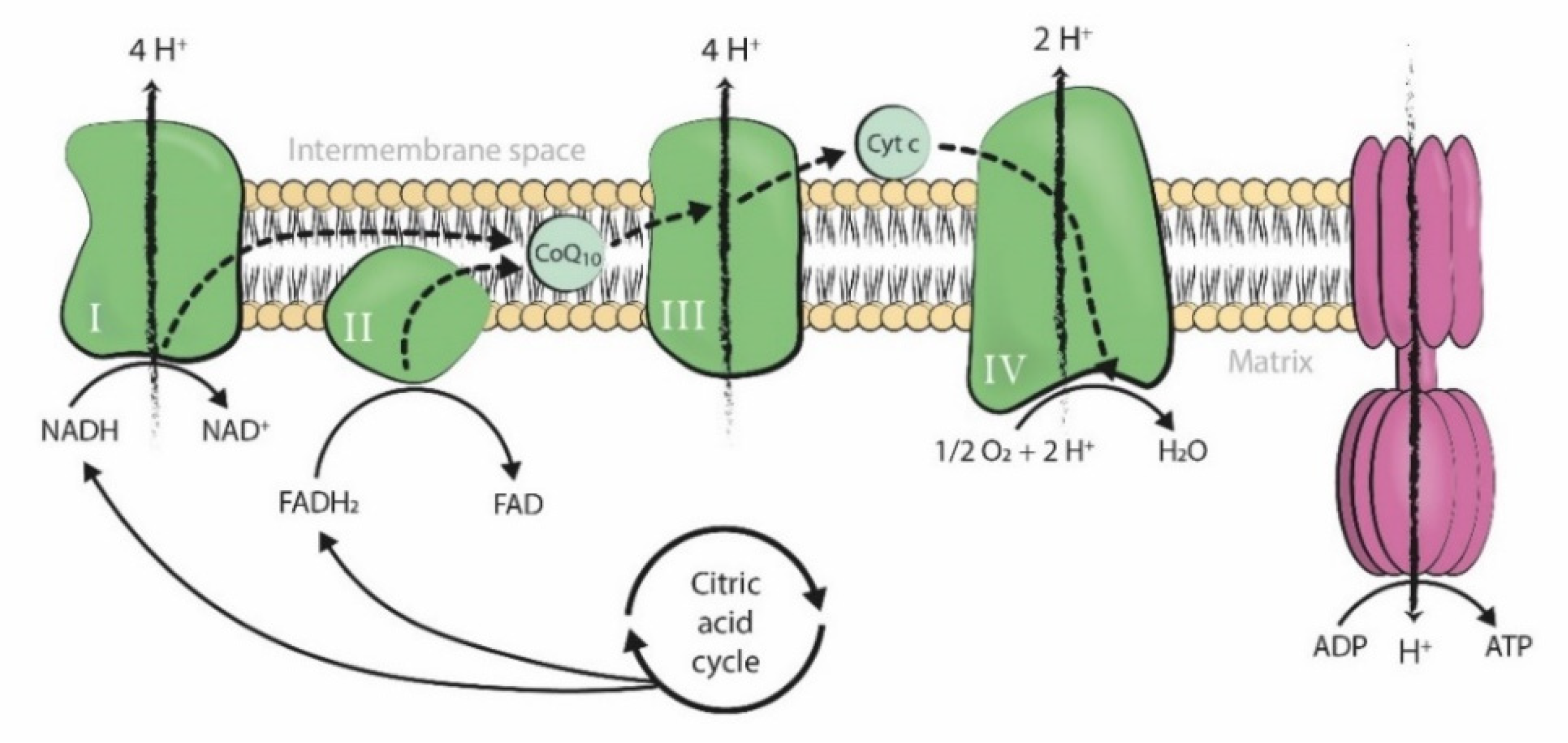
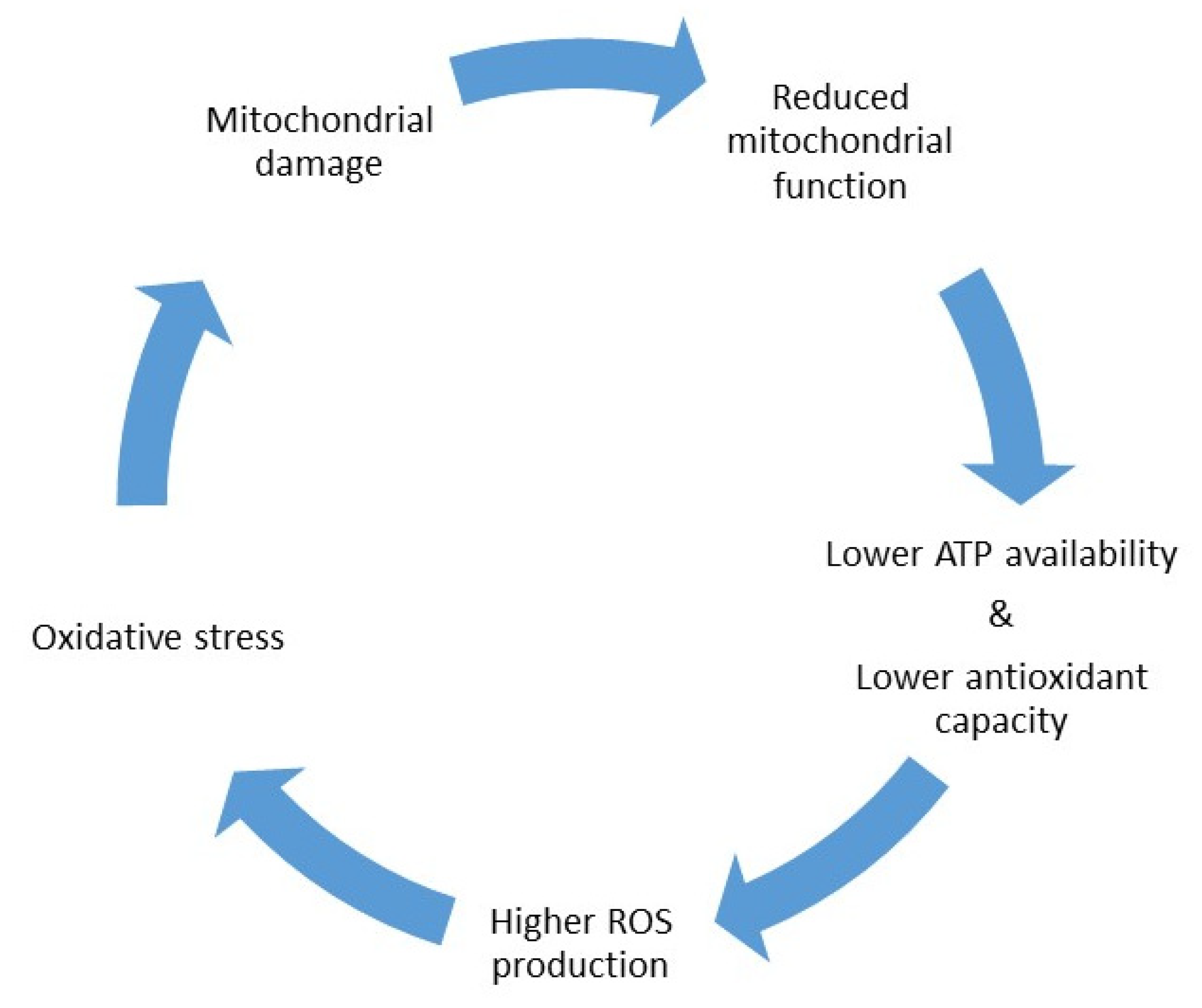
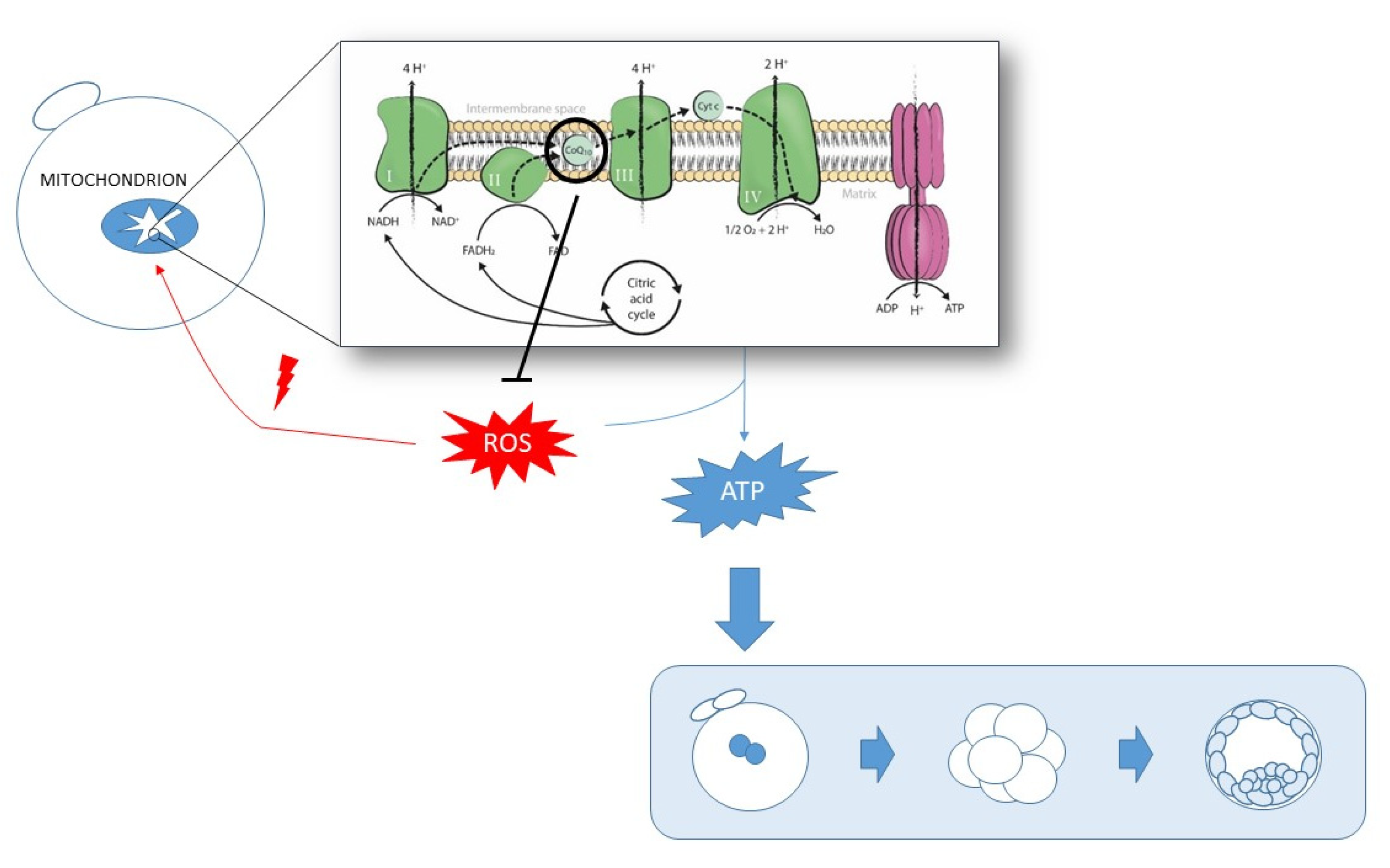
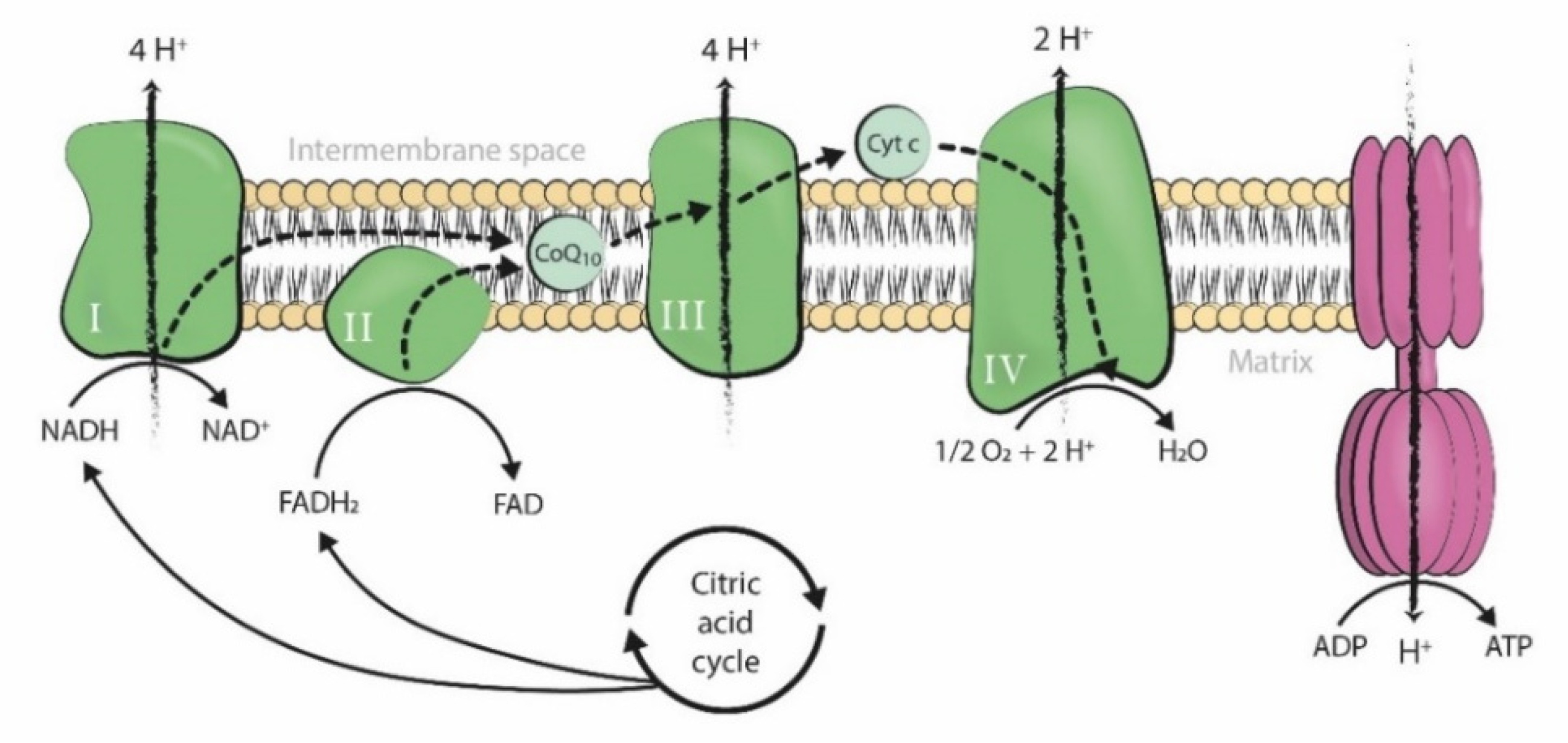
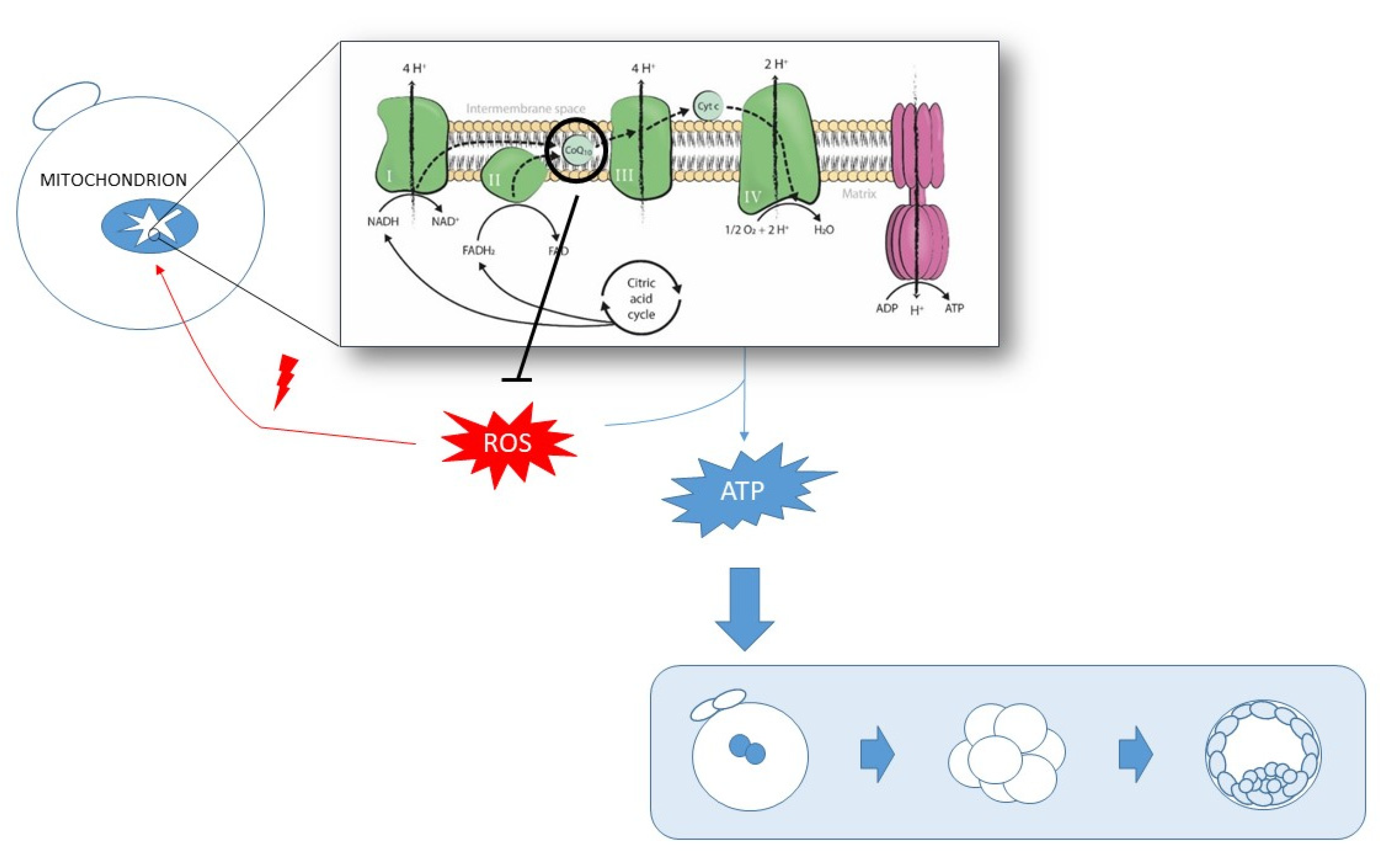
:1. Introduction
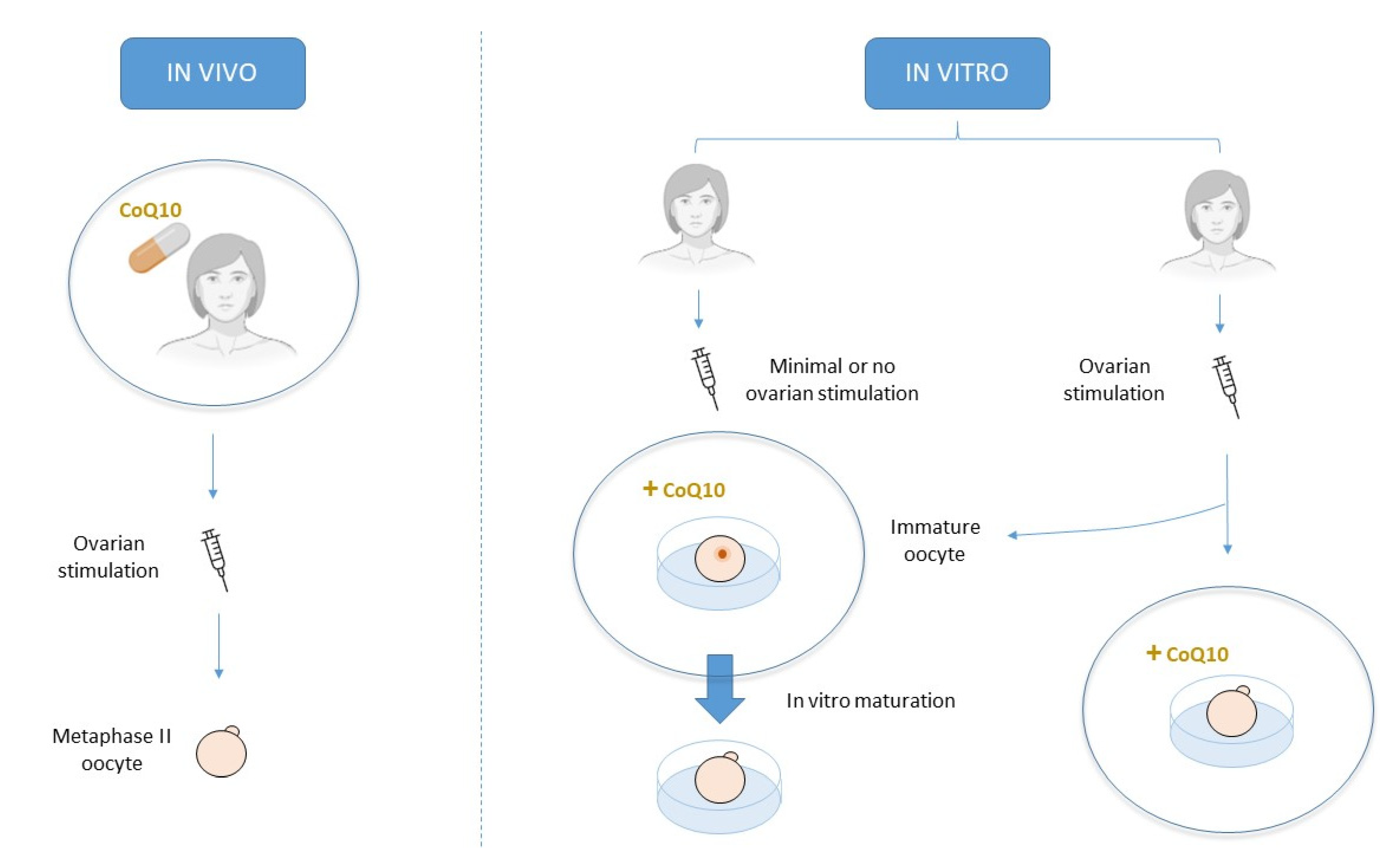
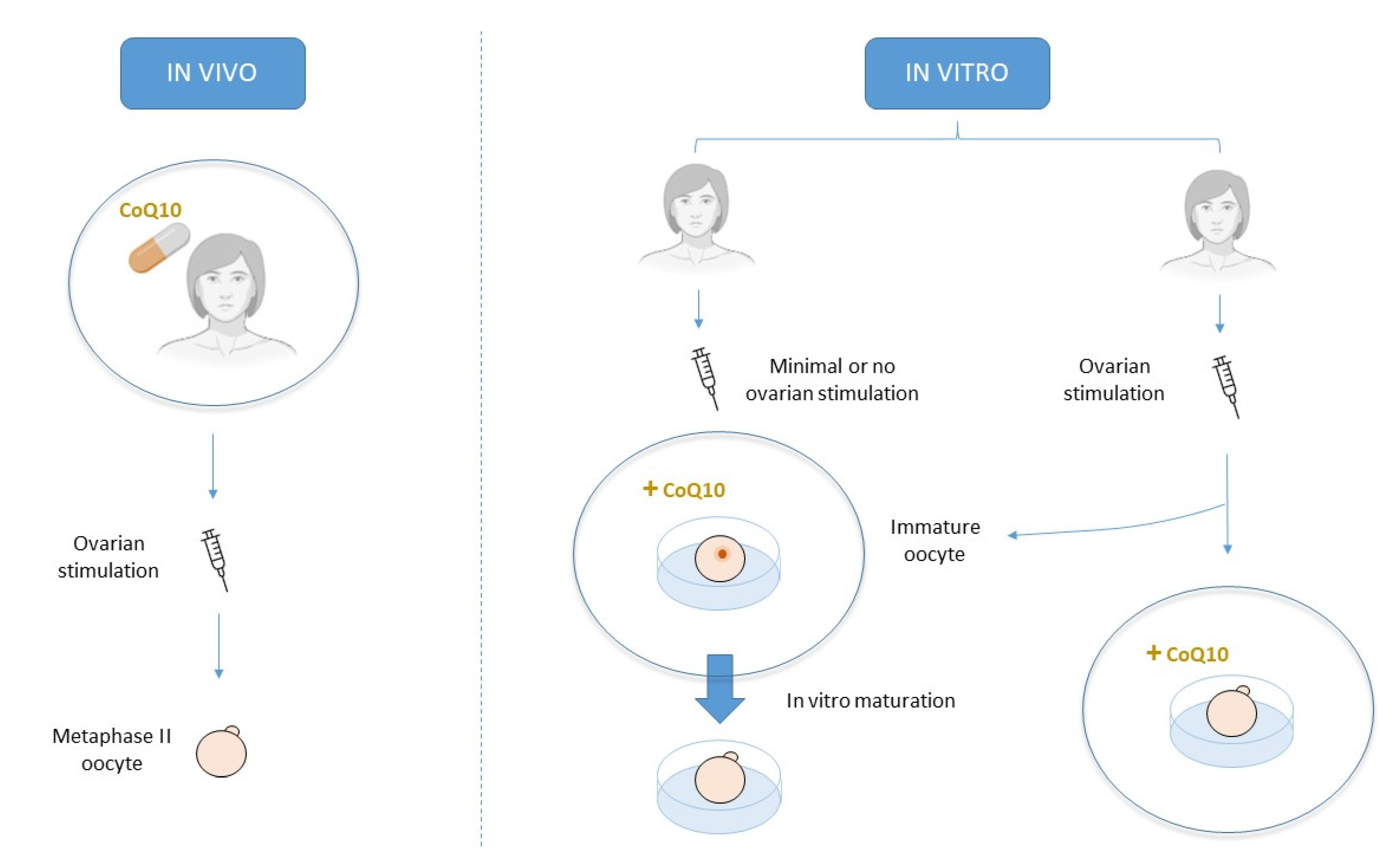
2. CoQ10 Supplementation in IVF Treatments
2.1. Oral Supplementation
2.2. Culture Media Supplementation
2.2.1. Standard Culture
2.2.2. In Vitro Maturation Culture
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Peixoto de Barcelos, I.; Haas, R.H. Coq10 and aging. Biology 2019, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Varela, C.; Labarta, E. Clinical application of antioxidants to improve human oocyte mitochondrial function: A review. Antioxidants 2020, 9, 1197. [Google Scholar] [CrossRef]
- Tran, U.P.C.; Clarke, C.F. Endogenous synthesis of coenzyme Q in eukaryotes. Mitochondrion 2007, 7, S62–S71. [Google Scholar] [CrossRef] [Green Version]
- Marcoff, L.; Thompson, P.D. The Role of Coenzyme Q10 in Statin-Associated Myopathy. A Systematic Review. J. Am. Coll. Cardiol. 2007, 49, 2231–2237. [Google Scholar] [CrossRef] [Green Version]
- Quinzii, C.M.; Hirano, M. Coenzyme Q and mitochondrial disease. Dev. Disabil. Res. Rev. 2010, 16, 183–188. [Google Scholar] [CrossRef]
- Bianchi, S.; Macchiarelli, G.; Micara, G.; Aragona, C.; Maione, M.; Nottola, S.A. Ultrastructural and morphometric evaluation of aged cumulus-oocyte-complexes. Ital. J. Anat. Embryol. 2013, 118, 28. [Google Scholar]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of oxidative stress on age-associated decline in oocyte developmental competence. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-Y.; Wang, D.H.; Zou, X.Y.; Xu, C.M. Mitochondrial functions on oocytes and preimplantation embryos. J. Zhejiang Univ. Sci. B 2009, 10, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, S.; Nottola, S.A.; Torge, D.; Palmerini, M.G.; Necozione, S.; Macchiarelli, G. Association between female reproductive health and mancozeb: Systematic review of experimental models. Int. J. Environ. Res. Public Health 2020, 17, 2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Song, Z.; Pei, S.; Yu, X.; Miao, J.; Liang, H.; Miao, Y.; Du, Z. Single cell RNA-seq reveals molecular pathways altered by 7, 12-dimethylbenz[a]anthracene treatment on pig oocytes. Theriogenology 2020, 157, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Revelli, A.; Piane, L.D.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7. [Google Scholar] [CrossRef] [Green Version]
- Turi, A.; Giannubilo, S.R.; Brugè, F.; Principi, F.; Battistoni, S.; Santoni, F.; Tranquilli, A.L.; Littarru, G.; Tiano, L. Coenzyme Q10 content in follicular fluid and its relationship with oocyte fertilization and embryo grading. Arch. Gynecol. Obstet. 2011, 285, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Akarsu, S.; Gode, F.; Isik, A.Z.; Dikmen, Z.G.; Tekindal, M.A. The association between coenzyme Q10 concentrations in follicular fluid with embryo morphokinetics and pregnancy rate in assisted reproductive techniques. J. Assist. Reprod. Genet. 2017, 34, 599–605. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Camacho, J.D.; Bernier, M.; López-Lluch, G.; Navas, P. Coenzyme Q10 supplementation in aging and disease. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Ben-Meir, A.; Burstein, E.; Borrego-Alvarez, A.; Chong, J.; Wong, E.; Yavorska, T.; Naranian, T.; Chi, M.; Wang, Y.; Bentov, Y.; et al. Coenzyme Q10 restores oocyte mitochondrial function and fertility during reproductive aging. Aging Cell 2015, 14, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Ben-Meir, A.; Kim, K.; McQuaid, R.; Esfandiari, N.; Bentov, Y.; Casper, R.F.; Jurisicova, A. Co-enzyme q10 supplementation rescues cumulus cells dysfunction in a maternal aging model. Antioxidants 2019, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; ShiYang, X.; Zhang, Y.; Miao, Y.; Chen, Y.; Cui, Z.; Xiong, B. Coenzyme Q10 ameliorates the quality of postovulatory aged oocytes by suppressing DNA damage and apoptosis. Free Radic. Biol. Med. 2019, 143, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.J.; Zhou, W.; Nie, Z.W.; Zhou, D.; Xu, Y.N.; Ock, S.A.; Yan, C.G.; Cui, X.S. Ubiquinol-10 delays postovulatory oocyte aging by improving mitochondrial renewal in pigs. Aging 2020, 12, 1256–1271. [Google Scholar] [CrossRef] [PubMed]
- Boots, C.E.; Boudoures, A.; Zhang, W.; Drury, A.; Moley, K.H. Obesity-induced oocyte mitochondrial defects are partially prevented and rescued by supplementation with co-enzyme Q10 in a mouse model. Hum. Reprod. 2016, 31, 2090–2097. [Google Scholar] [CrossRef] [Green Version]
- Özcan, P.; Fıçıcıoğlu, C.; Kizilkale, O.; Yesiladali, M.; Tok, O.E.; Ozkan, F.; Esrefoglu, M. Can Coenzyme Q10 supplementation protect the ovarian reserve against oxidative damage? J. Assist. Reprod. Genet. 2016, 33, 1223–1230. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, M.F.H.; Shin, N.; Karthikraj, R.; Barbosa, F.; Kannan, K.; Colaiácovo, M.P. Antioxidant CoQ10 restores fertility by rescuing bisphenol a-induced oxidative DNA damage in the caenorhabditis elegans germline. Genetics 2020, 214, 381–395. [Google Scholar] [CrossRef]
- Maside, C.; Martinez, C.A.; Cambra, J.M.; Lucas, X.; Martinez, E.A.; Gil, M.A.; Rodriguez-Martinez, H.; Parrilla, I.; Cuello, C. Supplementation with exogenous coenzyme Q10 to media for in vitro maturation and embryo culture fails to promote the developmental competence of porcine embryos. Reprod. Domest. Anim. 2019, 54, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Abdulhasan, M.K.; Li, Q.; Dai, J.; Abu-Soud, H.M.; Puscheck, E.E.; Rappolee, D.A. CoQ10 increases mitochondrial mass and polarization, ATP and Oct4 potency levels, and bovine oocyte MII during IVM while decreasing AMPK activity and oocyte death. J. Assist. Reprod. Genet. 2017, 34, 1595–1607. [Google Scholar] [CrossRef] [PubMed]
- Romero, S.; Pella, R.; Zorrilla, I.; Berrío, P.; Escudero, F.; Pérez, Y.; García, M.; Gonzalez, C.; Orihuela, P. Coenzyme q10 improves the in vitro maturation of oocytes exposed to the intrafollicular environment of patients on fertility treatment. JBRA Assist. Reprod. 2020, 24, 283–288. [Google Scholar] [CrossRef]
- Bentov, Y.; Hannam, T.; Jurisicova, A.; Esfandiari, N.; Casper, R.F. Coenzyme Q10 supplementation and oocyte aneuploidy in women undergoing IVF-ICSI treatment. Clin. Med. Insights Reprod. Health 2014, 8, CMRH.S14681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, T.; Fiameni, F.; Valcarcel, A.; Buzzi, J. Dietary supplementation with coenzyme Q10 in poor responder patients undergoing IVF-ICSI Treatment. Fertil. Steril. 2016, 106, e58. [Google Scholar] [CrossRef]
- Gat, I.; Mejia, S.B.; Balakier, H.; Librach, C.L.; Claessens, A.; Ryan, E.A.J. The use of coenzyme Q10 and DHEA during IUI and IVF cycles in patients with decreased ovarian reserve. Gynecol. Endocrinol. 2016, 32, 534–537. [Google Scholar] [CrossRef]
- Xu, Y.; Nisenblat, V.; Lu, C.; Li, R.; Qiao, J.; Zhen, X.; Wang, S. Pretreatment with coenzyme Q10 improves ovarian response and embryo quality in low-prognosis young women with decreased ovarian reserve: A randomized controlled trial. Reprod. Biol. Endocrinol. 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Giannubilo, S.R.; Orlando, P.; Silvestri, S.; Cirilli, I.; Marcheggiani, F.; Ciavattini, A.; Tiano, L. CoQ10 supplementation in patients undergoing IVF-ET: The relationship with follicular fluid content and oocyte maturity. Antioxidants 2018, 7, 141. [Google Scholar] [CrossRef] [Green Version]
- El Refaeey, A.; Selem, A.; Badawy, A. Combined coenzyme Q10 and clomiphene citrate for ovulation induction in clomiphene-citrate-resistant polycystic ovary syndrome. Reprod. Biomed. Online 2014, 29, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Sen Sharma, D. Co-enzyme Q10-A mitochondrial antioxidant—A new hope for success in infertility in clomiphene-citrate-resistant polycystic ovary syndrome. Int. J. Obstet. Gynaecol. Oral Commun. 2017. [Google Scholar] [CrossRef] [Green Version]
- Kile, R.; Logsdon, D.M.; Nathanson, C.; McCormick, S.; Schoolcraft, W.B.; Krisher, R.L. Mitochondrial support of embryos from women of advanced maternal age during ART. Fertil. Steril. 2020, 114, e122. [Google Scholar] [CrossRef]
- Ma, L.; Hu, M.; Ma, X.; Cui, Y.; Liu, J. CoQ10 decreases aneuploidy rate and increases mitochondrial mass during in vitro maturation of human immature oocytes. Fertil. Steril. ASRM Congr. 2018; P-510 2018, 110, e312. [Google Scholar] [CrossRef]
- Ma, L.; Cai, L.; Hu, M.; Wang, J.; Xie, J.; Xing, Y.; Shen, J.; Cui, Y.; Liu, X.J.; Liu, J. Coenzyme Q10 supplementation of human oocyte in vitro maturation reduces postmeiotic aneuploidies. Fertil. Steril. 2020, 114, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Al-Zubaidi, U.; Adhikari, D.; Cinar, O.; Zhang, Q.H.; Yuen, W.S.; Murphy, M.P.; Rombauts, L.; Robker, R.L.; Carroll, J. Mitochondria-targeted therapeutics, MitoQ and BGP-15, reverse aging-associated meiotic spindle defects in mouse and human oocytes. Hum. Reprod. 2020, 36, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Florou, P.; Anagnostis, P.; Theocharis, P.; Chourdakis, M.; Goulis, D.G. Does coenzyme Q10 supplementation improve fertility outcomes in women undergoing assisted reproductive technology procedures? A systematic review and meta-analysis of randomized-controlled trials. J. Assist. Reprod. Genet. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chiang, T.; Schultz, R.M.; Lampson, M.A. Meiotic origins of maternal age-related aneuploidy. Biol. Reprod. 2012, 86. [Google Scholar] [CrossRef]
- Du, K.; Ramachandran, A.; Weemhoff, J.L.; Woolbright, B.L.; Jaeschke, A.H.; Chao, X.; Ding, W.X.; Jaeschke, H. Mito-tempo protects against acute liver injury but induces limited secondary apoptosis during the late phase of acetaminophen hepatotoxicity. Arch. Toxicol. 2019, 93, 163–178. [Google Scholar] [CrossRef]
- Murphy, M.P.; Smith, R.A.J. Targeting antioxidants to mitochondria by conjugation to lipophilic cations. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 629–656. [Google Scholar] [CrossRef]
- Kelso, G.F.; Porteous, C.M.; Coulter, C.V.; Hughes, G.; Porteous, W.K.; Ledgerwood, E.C.; Smith, R.A.J.; Murphy, M.P. Selective targeting of a redox-active ubiquinone to mitochondria within cells: Antioxidant and antiapoptotic properties. J. Biol. Chem. 2001, 276, 4588–4596. [Google Scholar] [CrossRef] [Green Version]
- Jie, H.; Zhao, M.; Alqawasmeh, O.A.M.; Chan, C.P.S.; Lee, T.L.; Li, T.; Chan, D.Y.L. In vitro rescue immature oocytes—A literature review. Hum. Fertil. 2021. [Google Scholar] [CrossRef]
- Yeung, C.K.; Billings, F.T.; Claessens, A.J.; Roshanravan, B.; Linke, L.; Sundell, M.B.; Ahmad, S.; Shao, B.; Shen, D.D.; Ikizler, T.A.; et al. Coenzyme Q10 dose-escalation study in hemodialysis patients: Safety, tolerability, and effect on oxidative stress Dialysis and Transplantation. BMC Nephrol. 2015, 16, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, S.L.; Venkateswaran, S. Nutraceuticals in the prophylaxis of pediatric migraine: Evidence-based review and recommendations. Cephalalgia 2014, 34, 568–583. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, G.; Kanou, K.; Takamatsu, T.; Takeshita, M.; Morichi, S.; Suzuki, S.; Ishida, Y.; Watanabe, Y.; Go, S.; Oana, S.; et al. Complementary and integrative medicines as prophylactic agents for pediatric migraine: A narrative literature review. J. Clin. Med. 2021, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Richards, J.S. Ontogeny of the ovary in polycystic ovary syndrome. Fertil. Steril. 2013, 100, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Esteves, S.C.; Roque, M.; Bedoschi, G.M.; Conforti, A.; Humaidan, P.; Alviggi, C. Defining low prognosis patients undergoing assisted reproductive technology: POSEIDON criteria—The why. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xie, Y.; Yang, D.; Ren, D. Oxidative stress-induced apoptosis in granulosa cells involves JNK, p53 and Puma. Oncotarget 2017, 8, 25310–25322. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.; Zhang, J.; Zhang, J.; Wang, Y.; Wang, J.; Kang, J.; Quan, F.; Su, J.; Zhang, Y. Coenzyme Q10 supplement rescues postovulatory oocyte aging by regulating SIRT4 expression. Curr. Mol. Pharmacol. 2021. [Google Scholar] [CrossRef]
- Qiao, J.; Wang, Z.B.; Feng, H.L.; Miao, Y.L.; Wang, Q.; Yu, Y.; Wei, Y.C.; Yan, J.; Wang, W.H.; Shen, W.; et al. The root of reduced fertility in aged women and possible therapentic options: Current status and future perspects. Mol. Asp. Med. 2014, 38, 54–85. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Design | Treatment Groups (n) | Population | Outcomes | Results | ||
|---|---|---|---|---|---|---|---|
| In vivo | Bentov et al., 2014 [25] | RCT | CoQ10 gr.: 17 Control gr: 22 | IVF patients 35–43 y.o. | % Top quality embryos | X | |
| % Aneuploidy | X | ||||||
| Caballero et al., 2016 [26] | RCT | CoQ10 gr.: 39 Control gr.: 39 | Poor ovarian responders 36–40 y.o. | No. mature oocytes | X | ||
| Pregnancy outcomes | X | ||||||
| Gat et al., 2016 [27] | Retrospective | IUI cycles CoQ10 gr.: 330 Control gr.: 467 | Poor ovarian reserve IUI patients | Antral follicle count | ☺ | ||
| No. follicles >16 mm | ☺ | ||||||
| Pregnancy outcome | X | ||||||
| IVF cycles CoQ10 gr.: 78 Control gr.: 175 | Poor ovarian reserve IVF patients | Antral follicle count | ☺ | ||||
| No. follicles >16 mm | X | ||||||
| No. oocytes | X | ||||||
| No. zygotes | X | ||||||
| No. blastocysts | X | ||||||
| Pregnancy outcome | X | ||||||
| Xu et al., 2018 [28] | RCT | CoQ10 gr.: 76 Control gr.: 93 | Poor ovarian responders <35 y.o. | No. oocytes | ☺ | ||
| % Fertilization | ☺ | ||||||
| No. Top quality embryos | ☺ | ||||||
| Pregnancy outcome | X | ||||||
| Giannubilo et al., 2018 [29] | Prospective | 15 | IVF patients 31-46 y.o. with CoQ10 treatment | FF CoQ10 levels in follicles with an oocyte | X | ||
| El Refaeey et al., 2014 [30] | RCT | CoQ10 gr.: 51 Control gr.: 50 | CC-resistant PCOS | No. follicles >14 mm and ≥18 mm | ☺ | ||
| % Ovulation | ☺ | ||||||
| E2 and P4 levels | ☺ | ||||||
| Sen Sharma et al., 2017 [31] | RCT | CoQ10 gr.: 32 Control gr.: 30 | CC-resistant PCOS | Mature follicle size | ☺ | ||
| In vitro | Standard Culture | Kile et al., 2020 [32] | RCT | CoQ10 gr.: 66 Control gr.: 143 zygotes | IVF patients 35–46 y.o. | % Fertilization | X |
| % Top quality embryos | X | ||||||
| % Euploidy | X | ||||||
| IVM | Ma et al., 2018 [33] | RCT | CoQ10 gr.: 32 Control gr.: 32 immature oocytes | IVM patients 35–46 y.o. | Mitochondrial mass | ☺ | |
| Ma et al., 2020 [34] | RCT | Women ≤30 y.o. CoQ10 gr.: 37 Control gr.: 37 immature oocytes from stimulated cycles | IVF patients ≤30 y.o. | % Maturation | X | ||
| % Post-meiotic oocyte aneuploidy | X | ||||||
| Women ≥38 y.o. CoQ10 gr.: 46 Control gr.: 46 immature oocytes from stimulated cycles | IVF patients ≥38 y.o. | % Maturation | ☺ | ||||
| % Post-meiotic oocyte aneuploidy | ☺ | ||||||
| Al-Zubaidi et al., 2021 [35] | RCT | MitoQ gr.: 44 Control gr.: 45 immature oocytes from stimulated cycles | IVF patients 29–45 y.o. | % Maturation | ☺ | ||
| Mitochondrial membrane potential | ☺ | ||||||
| % Oocytes with misaligned chromosomes | ☺ | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Varela, C.; Labarta, E. Does Coenzyme Q10 Supplementation Improve Human Oocyte Quality? Int. J. Mol. Sci. 2021, 22, 9541. https://doi.org/10.3390/ijms22179541
Rodríguez-Varela C, Labarta E. Does Coenzyme Q10 Supplementation Improve Human Oocyte Quality? International Journal of Molecular Sciences. 2021; 22(17):9541. https://doi.org/10.3390/ijms22179541
Chicago/Turabian StyleRodríguez-Varela, Cristina, and Elena Labarta. 2021. "Does Coenzyme Q10 Supplementation Improve Human Oocyte Quality?" International Journal of Molecular Sciences 22, no. 17: 9541. https://doi.org/10.3390/ijms22179541
APA StyleRodríguez-Varela, C., & Labarta, E. (2021). Does Coenzyme Q10 Supplementation Improve Human Oocyte Quality? International Journal of Molecular Sciences, 22(17), 9541. https://doi.org/10.3390/ijms22179541