
RNA Granules in the Mitochondria and Their Organization under Mitochondrial Stresses


Abstract
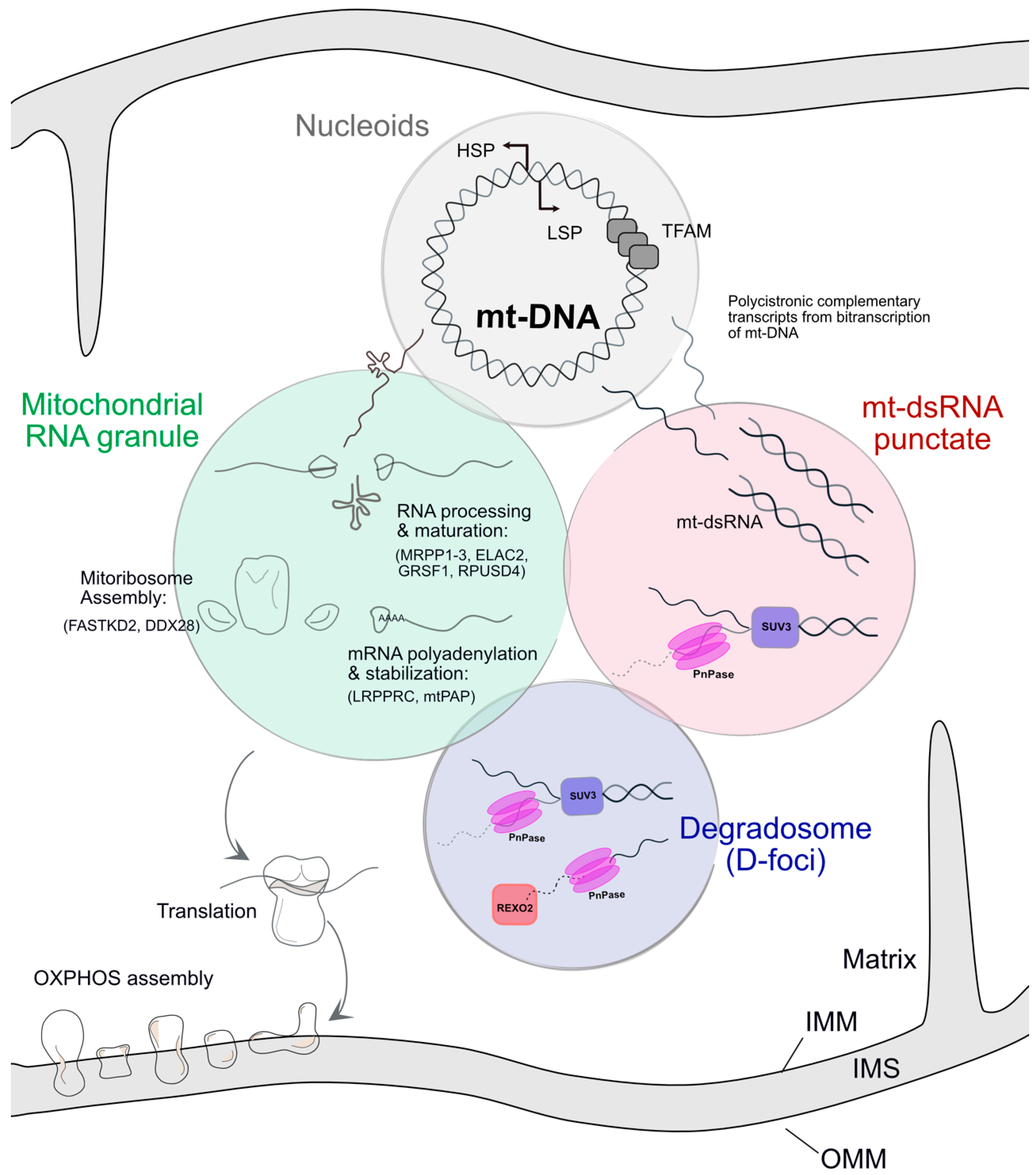
:1. Mitochondrial Gene Expression Is Compartmentalised
2. Mitochondrial RNA Granules Are Formed of Large Ribonucleoprotein Complexes
2.1. RNA Composition of MRGs
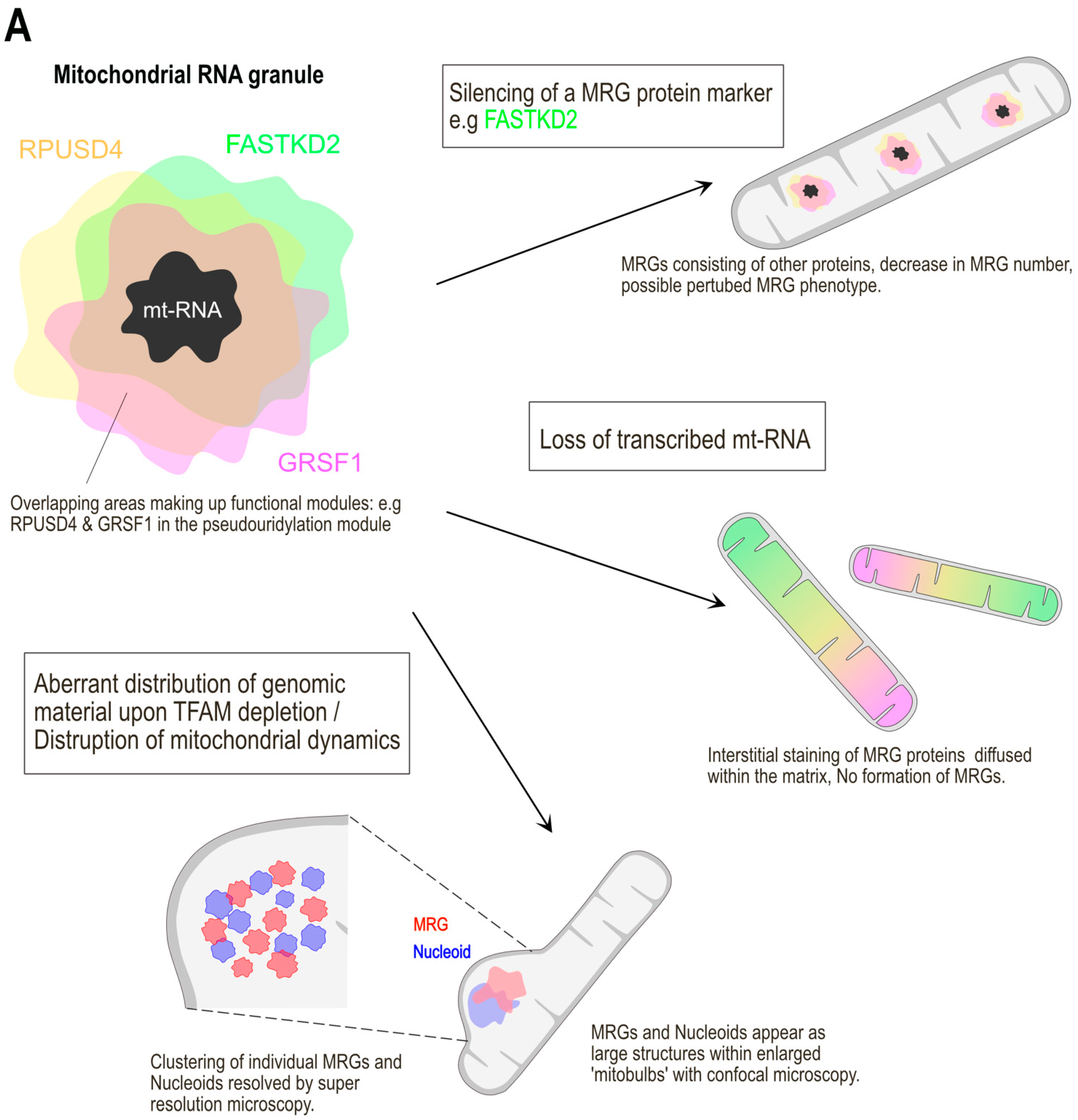
2.2. Protein Composition of MRGs

{kind=link}
{kind=link}
{kind=link}
| Gene Name | Function | Experimental Evidence |
|---|---|---|
| ALKBH1 | Histone H2A dioxygenase | Punctate foci with endogenous immunostaining [36]. |
| C17orf80c 1 | Uncharacterised Protein | RNA processing module of MRGs [35]. |
| CLPX | Mitochondrial Protease subunit | |
| DBT | Branched-chain alpha-keto acid dehydrogenase complex subunit | |
| DDX28 | Mitoribosome assembly | Punctate foci with overexpression of DDX28 tagged with HA [37]; endogenous immunostaining [25]. |
| DHX30 | Mitoribosome assembly | Punctate foci with endogenous immunostaining [25,38]. |
| ELAC2 2 | RNAse Z: 3′ tRNA processing | Proximal punctate foci with overexpression of ELAC2 fused with Eos [39]. |
| ERAL1 | Mitoribosome assembly | Punctate foci with overexpression of ERAL1 tagged with FLAG [33]. |
| FASTK | RNA processing | Punctate foci with overexpression of mitochondrial isoform of FASTK tagged with HA [21]. |
| FASTKD1 | RNA processing | Punctate foci with overexpression of FASTKD1 tagged with FLAG [30]. |
| FASTKD2 | RNA processing; mitoribosome assembly | Punctate foci with overexpression of FASTKD2 tagged with HA [21]. and endogenous immunostaining [25]. |
| FASTKD5 | RNA processing; mitoribosome assembly | Punctate foci with endogenous immunostaining [25] |
| FTSJ2 (MRM2) 2 | rRNA methylation | Proximal punctate foci with overexpression of MRM2 fused with Eos [40]. |
| GRSF1 | RNA processing; mitoribosome assembly | Punctate foci with endogenous immunostaining [15,16]. Overexpression of mitochondrial isoform of GRSF1 [15]. |
| GTPBP10 | Mitoribosome assembly | Member of the protein interactome co-purified with DDX28 [41]. |
| HARS2 | Mitochondrial histidyl tRNA synthetase | mtLSU assembly module of MRGs [35]. |
| HNRNPUL1 1 | Nuclear Ribonucleoprotein | Pseudouridylation module of MRGs [35]. |
| HNRNPA2B1 1 | Nuclear Ribonucleoprotein | |
| HNRNPA3 1 | Nuclear Ribonucleoprotein | |
| HNRNPK 1 | Nuclear Ribonucleoprotein | |
| HSD17B10 2 (MRPP2) | RNAse P subunit: 5′ tRNA processing | Putative partner of GRSF1 [15]. Proximal punctate foci with overexpression of MRPP2 fused with Eos [39]. |
| IBA57 | Iron-sulfur cluster assembly factor | RNA processing module of MRGs [35]. |
| LRPPRC | mRNA stability | Pseudouridylation module of MRGs [35]. |
| METTL15 | rRNA methylation | Punctate foci with overexpression of METTL15 tagged with FLAG [34]. |
| MRM1 2 | rRNA methylation | Proximal punctate foci with overexpression of MRM1 fused with Eos [40]. |
| MRPL12 | Mitoribosome subunit | RNA processing module of MRGs [35]. |
| MRPL14 | Mitoribosome subunit | |
| MRPL47 | Mitoribosome subunit | Punctate foci with overexpression of MRPL47 tagged with FLAG [33]. |
| MRPL58 | Mitoribosome subunit | RNA processing module of MRGs [35]. |
| MRPS7 | Mitoribosome subunit | Punctate foci with overexpression of MRPS7 tagged with FLAG [33]. |
| MRPS9 | Mitoribosome subunit | Punctate foci with overexpression of MRPS9 tagged with FLAG [33]. |
| MTERF3 (MTERFD1) | mt-LSU assembly | Punctate foci with overexpression of MTERFD1 tagged with FLAG [34]. |
| mtPAP | Poly(A) polymerase | Punctate foci with endogenous immunostaining and overexpression of mtPAP tagged with FLAG [32]. |
| NFS1 | Cysteine desulfurase | RNA processing module of MRGs [35]. |
| NGRN | RNA pseudouridylation | Punctate foci with overexpression of NGRN tagged with FLAG [34]. |
| NLRX1 | Regulator of mitochondrial antiviral immunity | Punctate foci with overexpression of NLRX1 fused with GFP [42]. |
| NOA1 | Involved in mitochondrial protein translation | Punctate foci with overexpression of NOA1 tagged with FLAG [33] |
| NT5DC2 | 5′-Nucleotidase | RNA processing module of MRGs [35]. |
| PNPT1 (PNPase) | 3′-5′-exoribonuclease activity | Bimolecular fluorescence complementation of SUV3-PNPase complex partially colocalizing with MRGs [43]. |
| PRORP (MRPP3) | RNAse P subunit: 5′ tRNA processing | Punctate foci with overexpression of MRPP3 tagged with MYC [15]. |
| PTCD1 | 3′ tRNA processing | Punctate foci with overexpression of PTCD1 tagged with FLAG [33]. |
| PTCD2 | RNA processing | Punctate foci with overexpression of PTCD2 tagged with FLAG [33]. |
| RBMX 1 | Nuclear Ribonucleoprotein | mtLSU assembly module of MRGs [35]. |
| RCC1L (WBSCR16) | RNA pseudouridylation | Punctate foci with overexpression of WBSCR16 tagged with FLAG [34]. |
| RNMTL1 2 (MRM3) | rRNA methylation | Proximal punctate foci with overexpression of MRM3 fused with Eos and endogenous immunostaining [40]. |
| RPUSD3 | RNA pseudouridylation | Punctate foci with overexpression of RPUSD3 tagged with FLAG [33,34]. |
| RPUSD4 | RNA pseudouridylation | Punctate foci with overexpression of RPUSD4 tagged with FLAG [33,34]. |
| SUPV3L1 (hSUV3p) | RNA helicase | Bimolecular fluorescence complementation of SUV3-PNPase complex partially colocalizing with MRGs [43]. |
| TFB1M | RNA modification | Punctate foci with overexpression of TFB1M tagged with FLAG [33]. |
| TRMT10C (MRPP1) | RNAse P subunit: 5′ tRNA processing; tRNA methyltransferase | Punctate foci with overexpression of MRPPI tagged with FLAG [15]. |
| TRUB2 | RNA pseudouridylation | Punctate foci with overexpression of TRUB2 tagged with FLAG [33,34]. |
| ZC3H4 1 | Zinc Finger domain protein | Pseudouridylation module of MRGs [35]. |
| ZFR 1 | Zinc Finger RNA Binding Protein |
2.3. MRGs Are Sub-Compartmentalised
2.4. Physical Properties of MRGs: The Role of RNA
2.5. Clustering of MRGs upon Impairment of Mitochondrial Dynamics
3. Granules Containing Mitochondrial Double-Stranded RNA (mt-dsRNA)
4. The Mitochondrial Degradosome or D-Foci
5. Dysregulation of the Degradosome and Its Effects on mt-dsRNA
6. Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BiFC | Bimolecular fluorescence complementation |
| BioID | Proximity-dependent biotinylation identification |
| BrU | 5-BromoUridine |
| cGAS–STING | Cyclic GMP/AMP Synthase–Stimulator of Interferon Genes |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| Dm | Drosophila melanogaster |
| dsRNA | Double-stranded RNA |
| ETC | Electron transport chain |
| FASTK | Fas-activated serine/threonine kinase |
| G-rich | Guanine-rich |
| G4 | G-quadruplexes |
| GRSF1 | G-rich sequence factor 1 |
| H-strand | Heavy strand |
| IMM | Inner mitochondrial membrane |
| IMS | Intermembrane space |
| L-strand | Light strand |
| LETM1 | Leucine zipper EF-hand-containing transmembrane protein 1 |
| LLPS | Liquid–liquid phase separation |
| LRPPRC | Leucine-rich PPR motif-containing protein |
| MELAS | Mitochondrial Encephylomyopathy, Lactic Acidosis and Stroke-like episodes |
| Mfn2 | Mitochondrial fusion factor Mitofusin 2 |
| MOMP | Mitochondrial outer membrane permeabilization |
| MRGs | Mitochondrial RNA granules |
| mt-DNA | Mitochondrial DNA |
| mt-dsRNA | Mitochondrial double-stranded RNA |
| mt-RNA | Mitochondrial RNA |
| mtLSU | Mitochondrial large ribosomal subunit |
| mtPAP | Poly(A) polymerase |
| OMM | Outer mitochondrial membrane |
| OXPHOS | Oxidative Phosphorylation |
| RBPs | RNA binding proteins |
| RIG-I-MDA5 | Retinoic Acid Inducible Protein I- Melanoma Differentiation Associated Gene 5 |
| RNAi | RNA interference |
| SLIRP | Stemloop interacting RNA-binding protein |
| STED | Stimulated emission depletion |
| TFAM | Transcription factor A, mitochondrial |
References
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z. Bin Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.L.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F. Sequence and organization of the human mitochondrial genome. Nature 1981, 290, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Aloni, Y.; Attardi, G. Symmetrical in vivo transcription of mitochondrial DNA in HeLa cells. Proc. Natl. Acad. Sci. USA 1971, 68, 1757–1761. [Google Scholar] [CrossRef] [Green Version]
- Aloni, Y.; Attardi, G. Expression of the Mitochondrial Genome in Hela Cells. J. Cell Biol. 1971, 56, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, C.M.; Falkenberg, M.; Larsson, N.G. Maintenance and Expression of Mammalian Mitochondrial DNA. Annu. Rev. Biochem. 2016, 85, 133–160. [Google Scholar] [CrossRef]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef]
- Rackham, O.; Busch, J.D.; Matic, S.; Siira, S.J.; Kuznetsova, I.; Atanassov, I.; Ermer, J.A.; Shearwood, A.M.J.; Richman, T.R.; Stewart, J.B.; et al. Hierarchical RNA Processing Is Required for Mitochondrial Ribosome Assembly. Cell Rep. 2016, 16, 1874–1890. [Google Scholar] [CrossRef] [Green Version]
- Siira, S.J.; Rossetti, G.; Richman, T.R.; Perks, K.; Ermer, J.A.; Kuznetsova, I.; Hughes, L.; Shearwood, A.J.; Viola, H.M.; Hool, L.C.; et al. Concerted regulation of mitochondrial and nuclear non-coding RNA s by a dual-targeted RN ase Z. EMBO Rep. 2018, 19, 1–18. [Google Scholar] [CrossRef]
- Kukat, C.; Davies, K.M.; Wurm, C.A.; Spåhr, H.; Bonekamp, N.A.; Kühl, I.; Joos, F.; Polosa, P.L.; Park, C.B.; Posse, V.; et al. Cross-strand binding of TFAM to a single mtDNA molecule forms the mitochondrial nucleoid. Proc. Natl. Acad. Sci. USA 2015, 112, 11288–11293. [Google Scholar] [CrossRef] [Green Version]
- Kukat, C.; Wurm, C.A.; Spåhr, H.; Falkenberg, M.; Larsson, N.G.; Jakobs, S. Super-resolution microscopy reveals that mammalian mitochondrial nucleoids have a uniform size and frequently contain a single copy of mtDNA. Proc. Natl. Acad. Sci. USA 2011, 108, 13534–13539. [Google Scholar] [CrossRef] [Green Version]
- Bogenhagen, D.F.; Rousseau, D.; Burke, S. The layered structure of human mitochondrial DNA nucleoids. J. Biol. Chem. 2008, 283, 3665–3675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajala, N.; Hensen, F.; Wessels, H.J.C.T.; Ives, D.; Gloerich, J.; Spelbrink, J.N. Whole cell formaldehyde cross-linking simplifies purification of mitochondrial nucleoids and associated proteins involved in mitochondrial gene expression. PLoS ONE 2015, 10, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Udeshi, N.D.; Deerinck, T.J.; Svinkina, T.; Ellisman, M.H.; Carr, S.A.; Ting, A.Y. Proximity Biotinylation as a Method for Mapping Proteins Associated with mtDNA in Living Cells. Cell Chem. Biol. 2017, 24, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iborra, F.J.; Kimura, H.; Cook, P.R. The functional organization of mitochondrial genomes in human cells. BMC Biol. 2004, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, A.A.; Koppen, M.; Wydro, M.; Rodley, C.D.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.; Martinou, J.-C. GRSF1 Regulates RNA Processing in Mitochondrial RNA Granules. Cell Metab. 2013, 17, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Antonicka, H.; Sasarman, F.; Nishimura, T.; Paupe, V.; Shoubridge, E.A. The mitochondrial RNA-binding protein GRSF1 localizes to RNA granules and is required for posttranscriptional mitochondrial gene expression. Cell Metab. 2013, 17, 386–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorkau, M.; Albus, C.A.; Berlinguer-Palmini, R.; Chrzanowska-Lightowlers, Z.M.A.; Lightowlers, R.N. High-resolution imaging reveals compartmentalization of mitochondrial protein synthesis in cultured human cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2008778118. [Google Scholar] [CrossRef]
- Schonborn, J.; Oberstraß, J.; Breyel, E.; Tittgen, J.; Schumacher, J.; Lukacs, N. Monoclonal antibodies to double-stranded RNA as probes of RNA structure in crude nucleic acid extracts. Nucleic Acids Res. 1991, 19, 2993–3000. [Google Scholar] [CrossRef]
- Dhir, A.; Dhir, S.; Borowski, L.S.; Jimenez, L.; Teitell, M.; Rötig, A.; Crow, Y.J.; Rice, G.I.; Duffy, D.; Tamby, C.; et al. Mitochondrial double-stranded RNA triggers antiviral signalling in humans. Nature 2018, 560, 238–242. [Google Scholar] [CrossRef]
- Qian, Z.; Wilusz, J. GRSF-1: A poly(A)+ mRNA binding protein which interacts with a conserved G-rich element. Nucleic Acids Res. 1994, 22, 2334–2343. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, A.A.; Koppen, M.; Rodley, C.D.; Maundrell, K.; Gueguen, N.; Reynier, P.; Guaras, A.M.; Enriquez, J.A.; Anderson, P.; Simarro, M.; et al. A Mitochondria-Specific Isoform of FASTK Is Present In Mitochondrial RNA Granules and Regulates Gene Expression and Function. Cell Rep. 2015, 10, 1110–1121. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, D.; Saada, A.; D’Adamo, P.; Fernandez-Vizarra, E.; Gasparini, P.; Tiranti, V.; Elpeleg, O.; Zeviani, M. FASTKD2 Nonsense Mutation in an Infantile Mitochondrial Encephalomyopathy Associated with Cytochrome C Oxidase Deficiency. Am. J. Hum. Genet. 2008, 83, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, D.H.; Choi, Y.C.; Nam, D.E.; Choi, S.S.; Kim, J.W.; Choi, B.O.; Chung, K.W. Identification of FASTKD2 compound heterozygous mutations as the underlying cause of autosomal recessive MELAS-like syndrome. Mitochondrion 2017, 35, 54–58. [Google Scholar] [CrossRef]
- Popow, J.; Alleaume, A.M.; Curk, T.; Schwarzl, T.; Sauer, S.; Hentze, M.W. FASTKD2 is an RNA-binding protein required for mitochondrial RNA processing and translation. Rna 2015, 21, 1873–1884. [Google Scholar] [CrossRef] [Green Version]
- Antonicka, H.; Shoubridge, E.A. Mitochondrial RNA Granules Are Centers for Posttranscriptional RNA Processing and Ribosome Biogenesis. Cell Rep. 2015, 10, 920–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perks, K.L.; Rossetti, G.; Kuznetsova, I.; Hughes, L.A.; Ermer, J.A.; Ferreira, N.; Busch, J.D.; Rudler, D.L.; Spahr, H.; Schöndorf, T.; et al. PTCD1 Is Required for 16S rRNA Maturation Complex Stability and Mitochondrial Ribosome Assembly. Cell Rep. 2018, 23, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, J.D.; Jourdain, A.A.; Calvo, S.E.; Ballarano, C.A.; Doench, J.G.; Root, D.E.; Mootha, V.K. A Genome-wide CRISPR Death Screen Identifies Genes Essential for Oxidative Phosphorylation. Cell Metab. 2016, 24, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, A.A.; Popow, J.; De La Fuente, M.A.; Martinou, J.C.; Anderson, P.; Simarro, M. Survey and summary: The FASTK family of proteins: Emerging regulators of mitochondrial RNA biology. Nucleic Acids Res. 2017, 45, 10941–10947. [Google Scholar] [CrossRef] [Green Version]
- Boehm, E.; Zornoza, M.; Jourdain, A.A.; Magdalena, A.D.; García-Consuegra, I.; Merino, R.T.; Orduña, A.; Martín, M.A.; Martinou, J.C.; De La Fuente, M.A.; et al. Role of FAST kinase domains 3 (FASTKD3) in post-transcriptional regulation of mitochondrial gene expression. J. Biol. Chem. 2016, 291, 25877–25887. [Google Scholar] [CrossRef] [Green Version]
- Boehm, E.; Zaganelli, S.; Maundrell, K.; Jourdain, A.A.; Thore, S.; Martinou, J.C. FASTKD1 and FASTKD4 have opposite effects on expression of specific mitochondrial RNAs, depending upon their endonuclease-like RAP domain. Nucleic Acids Res. 2017, 45, 6135–6146. [Google Scholar] [CrossRef]
- Wolf, A.R.; Mootha, V.K. Functional Genomic Analysis of Human Mitochondrial RNA Processing. Cell Rep. 2014, 7, 918–931. [Google Scholar] [CrossRef] [Green Version]
- Wilson, W.C.; Hornig-Do, H.T.; Bruni, F.; Chang, J.H. o.; Jourdain, A.A.; Martinou, J.C.; Falkenberg, M.; Spåhr, H.; Larsson, N.G.; Lewis, R.J.; et al. A human mitochondrial poly(A) polymerase mutation reveals the complexities of post-transcriptional mitochondrial gene expression. Hum. Mol. Genet. 2014, 23, 6345–6355. [Google Scholar] [CrossRef] [Green Version]
- Zaganelli, S.; Rebelo-Guiomar, P.; Maundrell, K.; Rozanska, A.; Pierredon, S.; Powell, C.A.; Jourdain, A.A.; Hulo, N.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.; et al. The pseudouridine synthase RPUSD4 is an essential component of mitochondrial RNA granules. J. Biol. Chem. 2017, 292, 4519–4532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonicka, H.; Choquet, K.; Lin, Z.; Gingras, A.; Kleinman, C.L.; Shoubridge, E.A. A pseudouridine synthase module is essential for mitochondrial protein synthesis and cell viability. EMBO Rep. 2017, 18, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Antonicka, H.; Lin, Z.Y.; Janer, A.; Aaltonen, M.J.; Weraarpachai, W.; Gingras, A.C.; Shoubridge, E.A. A High-Density Human Mitochondrial Proximity Interaction Network. Cell Metab. 2020, 32, 479–497.e9. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Hofmeister, O.; Rolland, S.G.; Maiser, A.; Aasumets, K.; Schmitt, S.; Schorpp, K.; Feuchtinger, A.; Hadian, K.; Schneider, S.; et al. Mitochondrial Alkbh1 localizes to mtRNA granules and its knockdown induces the mitochondrial UPR in humans and C. elegans. J. Cell Sci. 2019, 132, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tu, Y.-T.; Barrientos, A. The Human Mitochondrial DEAD-Box Protein DDX28 Resides in RNA Granules and Functions in Mitoribosome Assembly. Cell Rep. 2015, 10, 854–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Bogenhagen, D.F. Human mitochondrial DNA nucleoids are linked to protein folding machinery and metabolic enzymes at the mitochondrial inner membrane. J. Biol. Chem. 2006, 281, 25791–25802. [Google Scholar] [CrossRef] [Green Version]
- Bogenhagen, D.F.; Martin, D.W.; Koller, A. Initial steps in RNA processing and ribosome assembly occur at mitochondrial DNA nucleoids. Cell Metab. 2014, 19, 618–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.; Okot-Kotber, C.; La Comb, J.F.; Bogenhagen, D.F. Mitochondrial ribosomal RNA (rRNA) methyltransferase family members are positioned to modify nascent rRNA in foci near the mitochondrial DNA nucleoid. J. Biol. Chem. 2013, 288, 31386–31399. [Google Scholar] [CrossRef] [Green Version]
- Maiti, P.; Kim, H.J.; Tu, Y.T.; Barrientos, A. Human GTPBP10 is required for mitoribosome maturation. Nucleic Acids Res. 2018, 46, 11423–11437. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Sripada, L.; Lipatova, A.; Roy, M.; Prajapati, P.; Gohel, D.; Bhatelia, K.; Chumakov, P.M.; Singh, R. NLRX1 resides in mitochondrial RNA granules and regulates mitochondrial RNA processing and bioenergetic adaptation. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1260–1276. [Google Scholar] [CrossRef]
- Borowski, L.S.; Dziembowski, A.; Hejnowicz, M.S.; Stepien, P.P.; Szczesny, R.J. Human mitochondrial RNA decay mediated by PNPase-hSuv3 complex takes place in distinct foci. Nucleic Acids Res. 2013, 41, 1223–1240. [Google Scholar] [CrossRef]
- Rey, T.; Zaganelli, S.; Cuillery, E.; Vartholomaiou, E.; Croisier, M.; Martinou, J.C.; Manley, S. Mitochondrial RNA granules are fluid condensates positioned by membrane dynamics. Nat. Cell Biol. 2020, 22, 1180–1186. [Google Scholar] [CrossRef]
- Pietras, Z.; Wojcik, M.A.; Borowski, L.S.; Szewczyk, M.; Kulinski, T.M.; Cysewski, D.; Stepien, P.P.; Dziembowski, A.; Szczesny, R.J. Dedicated surveillance mechanism controls G-quadruplex forming non-coding RNAs in human mitochondria. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Na, Z.; Slavoff, S.A. P-Bodies: Composition, Properties, and Functions. Biochemistry 2018, 57, 2424–2431. [Google Scholar] [CrossRef]
- Buchan, J.R.; Parker, R. Eukaryotic Stress Granules: The Ins and Outs of Translation. Mol. Cell 2009, 36, 932–941. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, T.; Souquere, S.; Chujo, T.; Kobelke, S.; Chong, Y.S.; Fox, A.H.; Bond, C.S.; Nakagawa, S.; Pierron, G.; Hirose, T. Functional Domains of NEAT1 Architectural lncRNA Induce Paraspeckle Assembly through Phase Separation. Mol. Cell 2018, 70, 1038–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesterov, S.V.; Ilyinsky, N.S.; Uversky, V.N. Liquid-liquid phase separation as a common organizing principle of intracellular space and biomembranes providing dynamic adaptive responses. BBA Mol. Cell Res. 2021, 1868. [Google Scholar] [CrossRef]
- Maharana, S.; Wang, J.; Papadopoulos, D.K.; Richter, D.; Pozniakovsky, A.; Poser, I.; Bickle, M.; Rizk, S.; Guillén-boixet, J.; Franzmann, T.M.; et al. RNA buffers the phase separation behavior of prion-like RNA binding proteins. Science 2018, 360, 918–921. [Google Scholar] [CrossRef] [Green Version]
- Protter, D.S.W.; Rao, B.S.; Van Treeck, B.; Lin, Y.; Mizoue, L.; Rosen, M.K.; Parker, R. Intrinsically Disordered Regions Can Contribute Promiscuous Interactions to RNP Granule Assembly. Cell Rep. 2018, 22, 1401–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feric, M.; Demarest, T.G.; Tian, J.; Croteau, D.L.; Bohr, V.A.; Misteli, T. Self-assembly of multi-component mitochondrial nucleoids via phase separation. EMBO J. 2021, 40, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Kleele, T.; Rey, T.; Winter, J.; Zaganelli, S.; Mahecic, D.; Perreten Lambert, H.; Ruberto, F.P.; Nemir, M.; Wai, T.; Pedrazzini, T.; et al. Distinct fission signatures predict mitochondrial degradation or biogenesis. Nature 2021, 593, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Hensen, F.; Potter, A.; Van Esveld, S.L.; Tarrés-Solé, A.; Chakraborty, A.; Solà, M.; Spelbrink, J.N. Mitochondrial RNA granules are critically dependent on mtDNA replication factors Twinkle and mtSSB. Nucleic Acids Res. 2019, 47, 3680–3698. [Google Scholar] [CrossRef] [Green Version]
- Spelbrink, J.N.; Li, F.Y.; Tiranti, V.; Nikali, K.; Yuan, Q.P.; Tariq, M.; Wanrooij, S.; Garrido, N.; Comi, G.; Morandi, L.; et al. Correction: Human mitochondrial DNA deletions associated with mutations in the gene encoding Twinkle, a phage T7 gene 4-like protein localized in mitochondria. Nat. Genet. 2001, 29, 100. [Google Scholar] [CrossRef] [Green Version]
- Rajala, N.; Gerhold, J.M.; Martinsson, P.; Klymov, A.; Spelbrink, J.N. Replication factors transiently associate with mtDNA at the mitochondrial inner membrane to facilitate replication. Nucleic Acids Res. 2014, 42, 952–967. [Google Scholar] [CrossRef] [Green Version]
- Durigon, R.; Mitchell, A.L.; Jones, A.W.; Manole, A.; Mennuni, M.; Hirst, E.M.; Houlden, H.; Maragni, G.; Lattante, S.; Doronzio, P.N.; et al. LETM 1 couples mitochondrial DNA metabolism and nutrient preference. EMBO Mol. Med. 2018, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Tamai, S.; Iida, H.; Yokota, S.; Sayano, T.; Kiguchiya, S.; Ishihara, N.; Hayashi, J.I.; Mihara, K.; Oka, T. Characterization of the mitochondrial protein LETM1, which maintains the mitochondrial tubular shapes and interacts with the AAA-ATPase BCS1L. J. Cell Sci. 2008, 121, 2588–2600. [Google Scholar] [CrossRef] [Green Version]
- Zollino, M.; Lecce, R.; Fischetto, R.; Murdolo, M.; Faravelli, F.; Selicorni, A.; Buttè, C.; Memo, L.; Capovilla, G.; Neri, G. Mapping the Wolf-Hirschhorn syndrome phenotype outside the currently accepted WHS critical region and defining a new critical region, WHSCR-2. Am. J. Hum. Genet. 2003, 72, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lepage, P.; Miller, K.; Bunkenborg, J.; Reich, M.; Hjerrild, M.; Delmonte, T.; Villeneuve, A.; Sladek, R.; Xu, F.; et al. Identification of a gene causing human cytochrome c oxidase deficiency by integrative genomics. Proc. Natl. Acad. Sci. USA 2003, 100, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Oláhová, M.; Hardy, S.A.; Hall, J.; Yarham, J.W.; Haack, T.B.; Wilson, W.C.; Alston, C.L.; He, L.; Aznauryan, E.; Brown, R.M.; et al. LRPPRC mutations cause early-onset multisystem mitochondrial disease outside of the French-Canadian population. Brain 2015, 138, 3503–3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasarman, F.; Brunel-Guitton, C.; Antonicka, H.; Wai, T.; Shoubridge, E.A.; Allen, B.; Burelle, Y.; Charron, G.; Coderre, L.; DesRosiers, C.; et al. LRPPRC and SLIRP interact in a ribonucleoprotein complex that regulates posttranscriptional gene expression in mitochondria. Mol. Biol. Cell 2010, 21, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Chujo, T.; Ohira, T.; Sakaguchi, Y.; Goshima, N.; Nomura, N.; Nagao, A.; Suzuki, T. LRPPRC/SLIRP suppresses PNPase-mediated mRNA decay and promotes polyadenylation in human mitochondria. Nucleic Acids Res. 2012, 40, 8033–8047. [Google Scholar] [CrossRef] [Green Version]
- Ruzzenente, B.; Metodiev, M.D.; Wredenberg, A.; Bratic, A.; Park, C.B.; Cámara, Y.; Milenkovic, D.; Zickermann, V.; Wibom, R.; Hultenby, K.; et al. LRPPRC is necessary for polyadenylation and coordination of translation of mitochondrial mRNAs. EMBO J. 2012, 31, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Pajak, A.; Laine, I.; Clemente, P.; El-Fissi, N.; Schober, F.A.; Maffezzini, C.; Calvo-Garrido, J.; Wibom, R.; Filograna, R.; Dhir, A.; et al. Defects of mitochondrial RNA turnover lead to the accumulation of double-stranded RNA in vivo. PLoS Genet. 2019, 15, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Szczesny, R.J.; Borowski, L.S.; Malecki, M.; Wojcik, M.A.; Stepien, P.P.; Golik, P. RNA Degradation in Yeast and Human Mitochondria. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 1027–1034. [Google Scholar] [CrossRef]
- Dziembowski, A.; Piwowarski, J.; Hoser, R.; Minczuk, M.; Dmochowska, A.; Siep, M.; Van der Spek, H.; Grivell, L.; Stepien, P.P. The yeast mitochondrial degradosome: Its composition, interplay between RNA helicase and RNase activities and the role in mitochondrial RNA metabolism. J. Biol. Chem. 2003, 278, 1603–1611. [Google Scholar] [CrossRef] [Green Version]
- Kolesnikova, S.; Curtis, E.A. Structure and function of multimeric G-Quadruplexes. Molecules 2019, 24, 3074. [Google Scholar] [CrossRef] [Green Version]
- Bruni, F.; Gramegna, P.; Oliveira, J.M.A.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.A. REXO2 Is an Oligoribonuclease Active in Human Mitochondria. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, T.J.; Spåhr, H.; Jiang, S.; Siira, S.J.; Koolmeister, C.; Sharma, S.; Kauppila, J.H.K.; Jiang, M.; Kaever, V.; Rackham, O.; et al. Dinucleotide Degradation by REXO2 Maintains Promoter Specificity in Mammalian Mitochondria. Mol. Cell 2019, 76, 784–796.e6. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, M.; Malik, D.; Borowski, L.S.; Czarnomska, S.D.; Kotrys, A.V.; Klosowska-Kosicka, K.; Nowotny, M.; Szczesny, R.J. Human REXO2 controls short mitochondrial RNAs generated by mtRNA processing and decay machinery to prevent accumulation of double-stranded RNA. Nucleic Acids Res. 2020, 48, 5572–5590. [Google Scholar] [CrossRef]
- Barchiesi, A.; Vascotto, C. Transcription, processing, and decay of mitochondrial RNA in health and disease. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Camino, L.P.; Aguilera, A. Human mitochondrial degradosome prevents harmful mitochondrial R loops and mitochondrial genome instability. Proc. Natl. Acad. Sci. USA 2018, 115, 11024–11029. [Google Scholar] [CrossRef] [Green Version]
- Riley, J.S.; Quarato, G.; Cloix, C.; Lopez, J.; O’Prey, J.; Pearson, M.; Chapman, J.; Sesaki, H.; Carlin, L.M.; Passos, J.F.; et al. Mitochondrial inner membrane permeabilisation enables mt DNA release during apoptosis. EMBO J. 2018, 37, 1–16. [Google Scholar] [CrossRef] [PubMed]
- McArthur, K.; Whitehead, L.W.; Heddleston, J.M.; Li, L.; Padman, B.S.; Oorschot, V.; Geoghegan, N.D.; Chappaz, S.; Davidson, S.; Chin, H.S.; et al. BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science 2018, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tigano, M.; Vargas, D.C.; Fu, Y.; Tremblay-Belzile, S.; Sfeir, A. Nuclear sensing of mitochondrial DNA breaks enhances immune surveillance. Nature 2020, 591. [Google Scholar] [CrossRef]
- West, A.P.; Khoury-Hanold, W.; Staron, M.; Tal, M.C.; Pineda, C.M.; Lang, S.M.; Bestwick, M.; Duguay, B.A.; Raimundo, N.; MacDuff, D.A.; et al. Mitochondrial DNA stress primes the antiviral innate immune response. Nature 2015, 520, 553–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.J.; McArthur, K.; Metcalf, D.; Lane, R.M.; Cambier, J.C.; Herold, M.J.; Van Delft, M.F.; Bedoui, S.; Lessene, G.; Ritchie, M.E.; et al. Apoptotic caspases suppress mtDNA-induced STING-mediated type i IFN production. Cell 2014, 159, 1549–1562. [Google Scholar] [CrossRef] [Green Version]
- Rongvaux, A.; Jackson, R.; Harman, C.C.D.; Li, T.; West, A.P.; De Zoete, M.R.; Wu, Y.; Yordy, B.; Lakhani, S.A.; Kuan, C.Y.; et al. Apoptotic caspases prevent the induction of type i interferons by mitochondrial DNA. Cell 2014, 159, 1563–1577. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-W.; Rainey, R.N.; Balatoni, C.E.; Dawson, D.W.; Troke, J.J.; Wasiak, S.; Hong, J.S.; McBride, H.M.; Koehler, C.M.; Teitell, M.A.; et al. Mammalian Polynucleotide Phosphorylase Is an Intermembrane Space RNase That Maintains Mitochondrial Homeostasis. Mol. Cell. Biol. 2006, 26, 8475–8487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Ameln, S.; Wang, G.; Boulouiz, R.; Rutherford, M.A.; Smith, G.M.; Li, Y.; Pogoda, H.M.; Nürnberg, G.; Stiller, B.; Volk, A.E.; et al. A mutation in PNPT1, encoding mitochondrial-RNA-import protein PNPase, causes hereditary hearing loss. Am. J. Hum. Genet. 2012, 91, 919–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alodaib, A.; Sobreira, N.; Gold, W.A.; Riley, L.G.; Van Bergen, N.J.; Wilson, M.J.; Bennetts, B.; Thorburn, D.R.; Boehm, C.; Christodoulou, J. Whole-exome sequencing identifies novel variants in PNPT1 causing oxidative phosphorylation defects and severe multisystem disease. Eur. J. Hum. Genet. 2016, 25, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matilainen, S.; Carroll, C.J.; Richter, U.; Euro, L.; Pohjanpelto, M.; Paetau, A.; Isohanni, P.; Suomalainen, A. Defective mitochondrial RNA processing due to PNPT1 variants causes Leigh syndrome. Hum. Mol. Genet. 2017, 26, 3352–3361. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xavier, V.J.; Martinou, J.-C. RNA Granules in the Mitochondria and Their Organization under Mitochondrial Stresses. Int. J. Mol. Sci. 2021, 22, 9502. https://doi.org/10.3390/ijms22179502
Xavier VJ, Martinou J-C. RNA Granules in the Mitochondria and Their Organization under Mitochondrial Stresses. International Journal of Molecular Sciences. 2021; 22(17):9502. https://doi.org/10.3390/ijms22179502
Chicago/Turabian StyleXavier, Vanessa Joanne, and Jean-Claude Martinou. 2021. "RNA Granules in the Mitochondria and Their Organization under Mitochondrial Stresses" International Journal of Molecular Sciences 22, no. 17: 9502. https://doi.org/10.3390/ijms22179502
APA StyleXavier, V. J., & Martinou, J.-C. (2021). RNA Granules in the Mitochondria and Their Organization under Mitochondrial Stresses. International Journal of Molecular Sciences, 22(17), 9502. https://doi.org/10.3390/ijms22179502