
Protective and Anti-Inflammatory Effects of Protegrin-1 on Citrobacter rodentium Intestinal Infection in Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. PG-1 Reduced C. rodentium—Induced Intestinal Infection
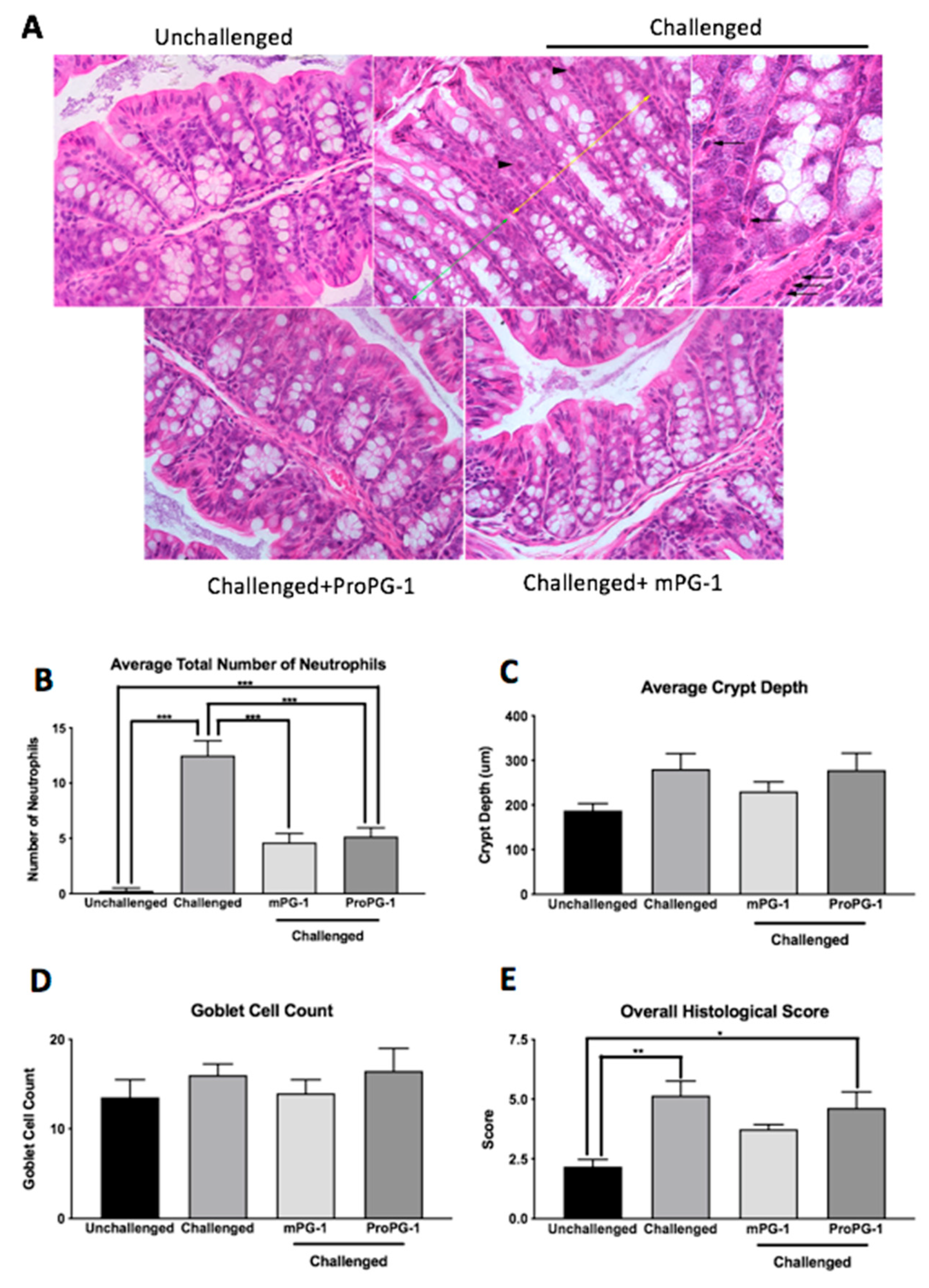
2.2. PG-1 Decreased C. rodentium—Induced Histologic Lesions in the Colon
2.3. PG-1 Modulates Intestinal Gene Expression during C. rodentium Infection
3. Discussion
4. Materials and Methods
4.1. Production of Recombinant ProPG-1 in Large-Scale Bioreactor Fermentations
4.2. Protein Sample Preparation
4.3. Ethics Statement
4.4. Bacterial Preparation
4.5. Animals and Induction of Colitis
4.6. Measurement of Bacterial Load and Fecal Water Content
4.7. Disease Activity Index Score
4.8. Tissue Collection
4.9. Histomorphology
4.10. Real-Time Quantitative PCR
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bhinder, G.; Sham, H.P.; Chan, J.M.; Morampudi, V.; Jacobson, K.; Vallance, B.A. The Citrobacter rodentium Mouse Model: Studying Pathogen and Host Contributions to Infectious Colitis. J. Vis. Exp. 2013, 72, e50222. [Google Scholar] [CrossRef]
- Papaconstantinou, H.T.; Thomas, J. Bacterial colitis. Clin. Colon Rectal. Surg. 2007, 20, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Loftus, E.V. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2004, 126, 1504–1517. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Aoki, W.; Ueda, M. Characterization of Antimicrobial Peptides toward the Development of Novel Antibiotics. Pharmaceuticals 2013, 6, 1055–1081. [Google Scholar] [CrossRef]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef]
- Li, J.; Post, M.; Volk, R.; Gao, Y.; Li, M.; Metais, C.; Sato, K.; Tsai, J.; Aird, W.; Rosenberg, R.D.; et al. PR39, a peptide regulator of angiogenesis. Nat. Med. 2000, 6, 49–55. [Google Scholar] [CrossRef]
- Carretero, M.; Escámez, M.J.; García, M.; Duarte, B.; Holguín, A.; Retamosa, L.; Jorcano, J.L.; Del Río, M.; Larcher, F. In vitro and in vivo wound healing-promoting activities of human cathelicidin LL-37. J. Investig. Dermatol. 2008, 128, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Büchau, A.S.; Morizane, S.; Trowbridge, J.; Schauber, J.; Kotol, P.; Bui, J.D.; Gallo, R.L. The host defense peptide cathelicidin is required for NK cell-mediated suppression of tumor growth. J. Immunol. 2010, 184, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Cunliffe, R.N.; Mahida, Y.R. Expression and regulation of antimicrobial peptides in the gastrointestinal tract. J. Leukoc. Biol. 2004, 75, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Pothoulakis, C.; Koon, H.W. Antimicrobial Peptides and Colitis. Curr. Pharm. Des. 2013, 19, 40–47. [Google Scholar] [PubMed]
- Tomasinsig, L.; Zanetti, M. The Cathelicidins—Structure, Function and Evolution. Curr. Protein Pept. Sci. 2005, 6, 23–24. [Google Scholar] [CrossRef]
- Zaiou, M.; Nizet, V.; Gallo, R.L. Antimicrobial and protease inhibitory functions of the human cathelicidin (hCAP18/LL-37) prosequence. J. Investig. Dermatol. 2003, 120, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of antimicrobial peptides. A review. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef]
- Zhao, C.; Ganz, T.; Lehre, R.I. The structure of porcine protegrin genes. FEBS Lett. 1995, 368, 197–202. [Google Scholar] [CrossRef]
- Steinberg, D.A.; Hurst, M.A.; Fujii, C.A.; Kung, A.H.; Ho, J.F.; Cheng, F.C.; Loury, D.J.; Fiddes, J.C. Protegrin-1: A Broad-Spectrum, Rapidly Microbicidal Peptide with In Vivo Activity. Antimicrob. Agents Chemother. 1997, 41, 1738–1742. [Google Scholar] [CrossRef]
- Harwig, S.S.; Waring, A.; Yang, H.J.; Cho, Y.; Tan, L.; Lehrer, R.I. Intramolecular disulfide bonds enhance the antimicrobial and lytic activities of protegrins at physiological sodium chloride concentrations. Eur. J. Biochem. 1996, 240, 352–357. [Google Scholar] [CrossRef]
- Cheung, Q.C.; Turner, P.V.; Song, C.; Wu, D.; Cai, H.Y.; MacInnes, J.I.; Li, J. Enhanced resistance to bacterial infection in protegrin-1 transgenic mice. Antimicrob. Agents Chemother. 2008, 52, 1812–1819. [Google Scholar] [CrossRef][Green Version]
- Huynh, E.; Penney, J.; Caswell, J.; Li, J. Protective effects of protegrin in dextran sodium sulfate induced murine colitis. Front. Pharmacol. 2019, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Saucedo, C.; Bernal-Reynaga, R.; Zayas-Jahuey, J.; Galindo-Gomez, S.; Shibayama, M.; Garcia-Galvez, C.; Estrada-Parra, S.; Estrada-Garcia, T. CD40 Ligand Deficient C57BL/6 Mouse Is a Potential Surrogate Model of Human X-Linked Hyper IgM (X-HIGM) Syndrome for Characterizing Immune Responses against Pathogens. BioMed Res. Int. 2015, 2015, 679850. [Google Scholar] [CrossRef]
- Symonds, E.L.; Riedel, C.U.; O’Mahony, D.; Lapthorne, S.; O’Mahony, L.; Shanahan, F. Involvement of T helper type 17 and regulatory T cell activity in Citrobacter rodentium invasion and inflammatory damage. Clin. Exp. Immunol. 2009, 157, 148–154. [Google Scholar] [CrossRef]
- Luperchio, S.A.; Newman, J.V.; Dangler, C.A.; Schrenzel, M.D.; Brenner, D.J.; Steigerwalt, A.G.; Schauer, D.B. Citrobacter rodentium, the Causative Agent of Transmissible Murine Colonic Hyperplasia, Exhibits Clonality: Synonymy of C. rodentium and Mouse-Pathogenic Escherichia coli. J. Clin. Microbiol. 2000, 38, 4343–4350. [Google Scholar] [CrossRef]
- Maaser, C.; Housley, M.P.; Iimura, M.; Smith, J.R.; Vallance, B.A.; Finlay, B.B.; Schreiber, J.R.; Varki, N.M.; Kagnoff, M.F.; Eckmann, L. Clearance of Citrobacter rodentium requires B cells but not secretory immunoglobulin A (IgA) or IgM antibodies. Infect. Immun. 2004, 72, 3315–3324. [Google Scholar] [CrossRef]
- Alipour, M.; Lou, Y.; Zimmerman, D.; Bording-Jorgensen, M.W.; Sergi, C.; Liu, J.J.; Wine, E. A balanced IL-1beta activity is required for host response to Citrobacter rodentium infection. PLoS ONE 2013, 8, e80656. [Google Scholar] [CrossRef] [PubMed]
- Crepin, V.F.; Collins, J.W.; Habibzay, M.; Frankel, G. Citrobacter rodentium mouse model of bacterial infection. Nat. Protoc. 2016, 11, 1851–1876. [Google Scholar] [CrossRef]
- van Ampting, M.T.; Loonen, L.M.; Schonewille, A.J.; Konings, I.; Vink, C.; Iovanna, J.; Chamaillard, M.; Dekker, J.; van der Meer, R.; Wells, J.M.; et al. Intestinally secreted C-type lectin Reg3b attenuates salmonellosis but not listeriosis in mice. Infect. Immun. 2012, 80, 1115–1120. [Google Scholar] [CrossRef]
- van Ampting, M.T.; Rodenburg, W.; Vink, C.; Kramer, E.; Schonewille, A.J.; Keijer, J.; van der Meer, R.; Bovee-Oudenhoven, I.M. Ileal mucosal and fecal pancreatitis associated protein levels reflect severity of salmonella infection in rats. Dig. Dis. Sci. 2009, 54, 2588–2597. [Google Scholar] [CrossRef] [PubMed]
- Schewe, M.; Franken, P.F.; Sacchetti, A.; Schmitt, M.; Joosten, R.; Böttcher, R.; van Royen, M.E.; Jeammet, L.; Payré, C.; Scott, P.M.; et al. Secreted Phospholipases A2 Are Intestinal Stem Cell Niche Factors with Distinct Roles in Homeostasis, Inflammation, and Cancer. Cell Stem Cell 2016, 19, 38–51. [Google Scholar] [CrossRef]
- Perez-Vilar, J.; Hill, R.L. The Structure and Assembly of Secreted Mucins. J. Biol. Chem. 1999, 274, 31751–31754. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.; Larsson, J.M.; Hansson, G.C. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host-microbial interactions. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4659–4665. [Google Scholar] [CrossRef]
- Nishida, A.; Lau, C.W.; Zhang, M.; Andoh, A.; Shi, H.N.; Mizoguchi, E.; Mizoguchi, A. The membrane-bound mucin Muc1 regulates T helper 17-cell responses and colitis in mice. Gastroenterology 2012, 142, 865–874. [Google Scholar] [CrossRef]
- Kato, K.; Lillehoj, E.P.; Lu, W.; Kim, K.C. Muc1: The first respiratory mucin with an anti-inflammatory function. J. Clin. Med. 2017, 6, 110. [Google Scholar] [CrossRef]
- Bergstrom, K.S.; Kissoon-Singh, V.; Gibson, D.L.; Ma, C.; Montero, M.; Sham, H.P.; Ryz, N.; Huang, T.; Velcich, A.; Finlay, B.B.; et al. Muc2 protects against lethal infectious colitis by disassociating pathogenic and commensal bacteria from the colonic mucosa. PLoS Pathog. 2010, 6, e1000902. [Google Scholar] [CrossRef]
- Hauber, H.P.; Foley, S.C.; Hamid, Q. Mucin overproduction in chronic inflammatory lung disease. Can. Respir. J. 2006, 13, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak-Jedrzejczyk, A.; Makosiej, R.; Kolejwa, M.; Glowacka, E.; Czkwianianc, E. The role of adhesion molecules in inflammatory bowel disease in children. Assessment of the possible risk of cardiovascular complications. Prz. Gastroenterol. 2017, 12, 181–185. [Google Scholar] [CrossRef]
- Sans, M.; Panés, J.; Ardite, E.; Elizalde, J.I.; Arce, Y.; Elena, M.; Palacín, A.; Fernández–Checa, J.C.; Anderson, D.C.; Lobb, R.; et al. VCAM-1 and ICAM-1 Mediate Leukocyte-Endothelial Cell Adhesion in Rat Experimental Colitis. Gastroenterology 1999, 116, 874–883. [Google Scholar] [CrossRef]
- Jones, S.C.; Banks, R.E.; Haidar, A.; Gearing, A.J.; Hemingway, I.K.; Ibbotson, S.H.; Dixon, M.F.; Axon, A.T. Adhesion molecules in inflammatory bowel disease. Gut 1995, 36, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yao, J.; Mercola, D.; Adamson, E. The transcription factor EGR-1 directly transactivates the fibronectin gene and enhances attachment of human glioblastoma cell line U251. J. Biol. Chem. 2000, 275, 20315–20323. [Google Scholar] [CrossRef] [PubMed]
- Min, I.M.; Pietramaggiori, G.; Kim, F.S.; Passegue, E.; Stevenson, K.E.; Wagers, A.J. The transcription factor EGR1 controls both the proliferation and localization of hematopoietic stem cells. Cell Stem Cell 2008, 2, 380–391. [Google Scholar] [CrossRef]
- Kim, J.M.; Eckmann, L.; Savidge, T.C.; Lowe, D.C.; Witthoft, T.; Kagnoff, M.F. Apoptosis of human intestinal epithelial cells after bacterial invasion. J. Clin. Investig. 1998, 102, 1815–1823. [Google Scholar] [CrossRef]
- Shah, Y.M. The role of hypoxia in intestinal inflammation. Mol. Cell Pediatr. 2016, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Garner, T.P.; Reyna, D.E.; Priyadarshi, A.; Chen, H.C.; Li, S.; Wu, Y.; Ganesan, Y.T.; Malashkevich, V.N.; Cheng, E.H.; Gavathiotis, E. An Autoinhibited Dimeric Form of BAX Regulates the BAX Activation Pathway. Mol. Cell. 2016, 63, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Bentires-Alj, M.; Dejardin, E.; Viatour, P.; Van Lint, C.; Froesch, B.; Reed, J.C.; Merville, M.P.; Bours, V. Inhibition of the NF-kB transcription factor increases Bax expression in cancer cell lines. Oncogene 2001, 20, 2805–2813. [Google Scholar] [CrossRef]
- Poveda, J.; Sanz, A.B.; Carrasco, S.; Ruiz-Ortega, M.; Cannata-Ortiz, P.; Sanchez-Niño, M.D.; Ortiz, A. Bcl3: A regulator of NF-kappaB inducible by TWEAK in acute kidney injury with anti-inflammatory and antiapoptotic properties in tubular cells. Exp. Mol. Med. 2017, 49, e352. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, B. Expression of Toll-like receptors in the mucosa of patients with ulcerative colitis. Exp. Ther. Med. 2015, 9, 1455–1459. [Google Scholar] [CrossRef]
- Morgan, M.E.; Koelink, P.J.; Zheng, B.; Den Brok, M.H.; Van De Kant, H.J.; Verspaget, H.W.; Folkerts, G.; Adema, G.J.; Kraneveld, A.D. Toll-like receptor 6 stimulation promotes T-helper 1 and 17 responses in gastrointestinal-associated lymphoid tissue and modulates murine experimental colitis. Mucosal Immunol. 2014, 7, 1266–1277. [Google Scholar] [CrossRef]
- Croker, B.A.; Kiu, H.; Nicholson, S.E. SOCS Regulation of the JAK/STAT Signalling Pathway. Semin. Cell. Dev. Biol. 2008, 19, 414–422. [Google Scholar] [CrossRef]
- Cole, A.M.; Shi, J.; Ceccarelli, A.; Kim, Y.H.; Park, A.; Ganz, T. Inhibition of neutrophil elastase prevents cathelicidin activation and impairs clearance of bacteria from wounds. Blood 2001, 97, 297–304. [Google Scholar] [CrossRef]
- Shahravan, S.H.; Qu, X.; Chan, I.S.; Shin, J.A. Enhancing the specificity of the enterokinase cleavage reaction to promote efficient cleavage of a fusion tag. Protein Expr. Purif. 2008, 59, 314–319. [Google Scholar] [CrossRef]
- Vergnolle, N. Protease inhibition as new therapeutic strategy for GI diseases. Gut 2016, 65, 1215–1224. [Google Scholar] [CrossRef]
- Collins, J.W.; Keeney, K.M.; Crepin, V.F.; Rathinam, V.A.; Fitzgerald, K.A.; Finlay, B.B.; Frankel, G. Citrobacter rodentium: Infection, inflammation and the microbiota. Nat. Rev. Microbiol. 2014, 12, 612–623. [Google Scholar] [CrossRef]
- Huynh, E.; Akhtar, N.; Li, J. Efficient Production of Recombinant Protegrin-1 from Pichia pastoris, and Its Antimicrobial and in vitro Cell Migration Activity. Front. Microbiol. 2018, 9, 2300. [Google Scholar] [CrossRef]
- Johnson-Henry, K.C.; Nadjafi, M.; Avitzur, Y.; Mitchell, D.J.; Ngan, B.Y.; Galindo-Mata, E.; Jones, N.L.; Sherman, P.M. Amelioration of the effects of Citrobacter rodentium infection in mice by pretreatment with probiotics. J. Infect. Dis. 2005, 191, 2106–2117. [Google Scholar] [CrossRef]
- MacDonald, T.T.; Frankel, G.; Dougan, G.; Goncalves, N.S.; Simmons, C. Host defences to Citrobacter rodentium. Int. J. Med. Microbiol. 2003, 293, 87–93. [Google Scholar] [CrossRef]
- Schauer, D.B.; Falkow, S. Attaching and effacing locus of a Citrobacter freundii biotype that causes transmissible murine colonic hyperplasia. Infect. Immun. 1993, 61, 2486–2492. [Google Scholar] [CrossRef]
- Spehlmann, M.E.; Dann, S.M.; Hruz, P.; Hanson, E.; McCole, D.F.; Eckmann, L. CXCR2-dependent mucosal neutrophil influx protects against colitis-associated diarrhea caused by an attaching/effacing lesion-forming bacterial pathogen. J. Immunol. 2009, 183, 3332–3343. [Google Scholar] [CrossRef]
- Maxwell, J.R.; Brown, W.A.; Smith, C.L.; Byrne, F.R.; Viney, J.L. Methods of inducing inflammatory bowel disease in mice. Curr. Protoc. Pharm. 2009, 72, 5–58. [Google Scholar] [CrossRef]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar] [PubMed]
- Buschor, S.; Cuenca, M.; Uster, S.S.; Schären, O.P.; Balmer, M.L.; Terrazos, M.A.; Schürch, C.M.; Hapfelmeier, S. Innate immunity restricts Citrobacter rodentium A/E pathogenesis initiation to an early window of opportunity. PLoS Pathog. 2017, 13, e1006476. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osakowicz, C.; Fletcher, L.; Caswell, J.L.; Li, J. Protective and Anti-Inflammatory Effects of Protegrin-1 on Citrobacter rodentium Intestinal Infection in Mice. Int. J. Mol. Sci. 2021, 22, 9494. https://doi.org/10.3390/ijms22179494
Osakowicz C, Fletcher L, Caswell JL, Li J. Protective and Anti-Inflammatory Effects of Protegrin-1 on Citrobacter rodentium Intestinal Infection in Mice. International Journal of Molecular Sciences. 2021; 22(17):9494. https://doi.org/10.3390/ijms22179494
Chicago/Turabian StyleOsakowicz, Celina, Lauren Fletcher, Jeff L. Caswell, and Julang Li. 2021. "Protective and Anti-Inflammatory Effects of Protegrin-1 on Citrobacter rodentium Intestinal Infection in Mice" International Journal of Molecular Sciences 22, no. 17: 9494. https://doi.org/10.3390/ijms22179494
APA StyleOsakowicz, C., Fletcher, L., Caswell, J. L., & Li, J. (2021). Protective and Anti-Inflammatory Effects of Protegrin-1 on Citrobacter rodentium Intestinal Infection in Mice. International Journal of Molecular Sciences, 22(17), 9494. https://doi.org/10.3390/ijms22179494