
A Mutation in DNA Polymerase α Rescues WEE1KO Sensitivity to HU

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. A Mutation in the Catalytic Subunit of Polymerase α Rescues HU Sensitivity of WEE1KO Plants
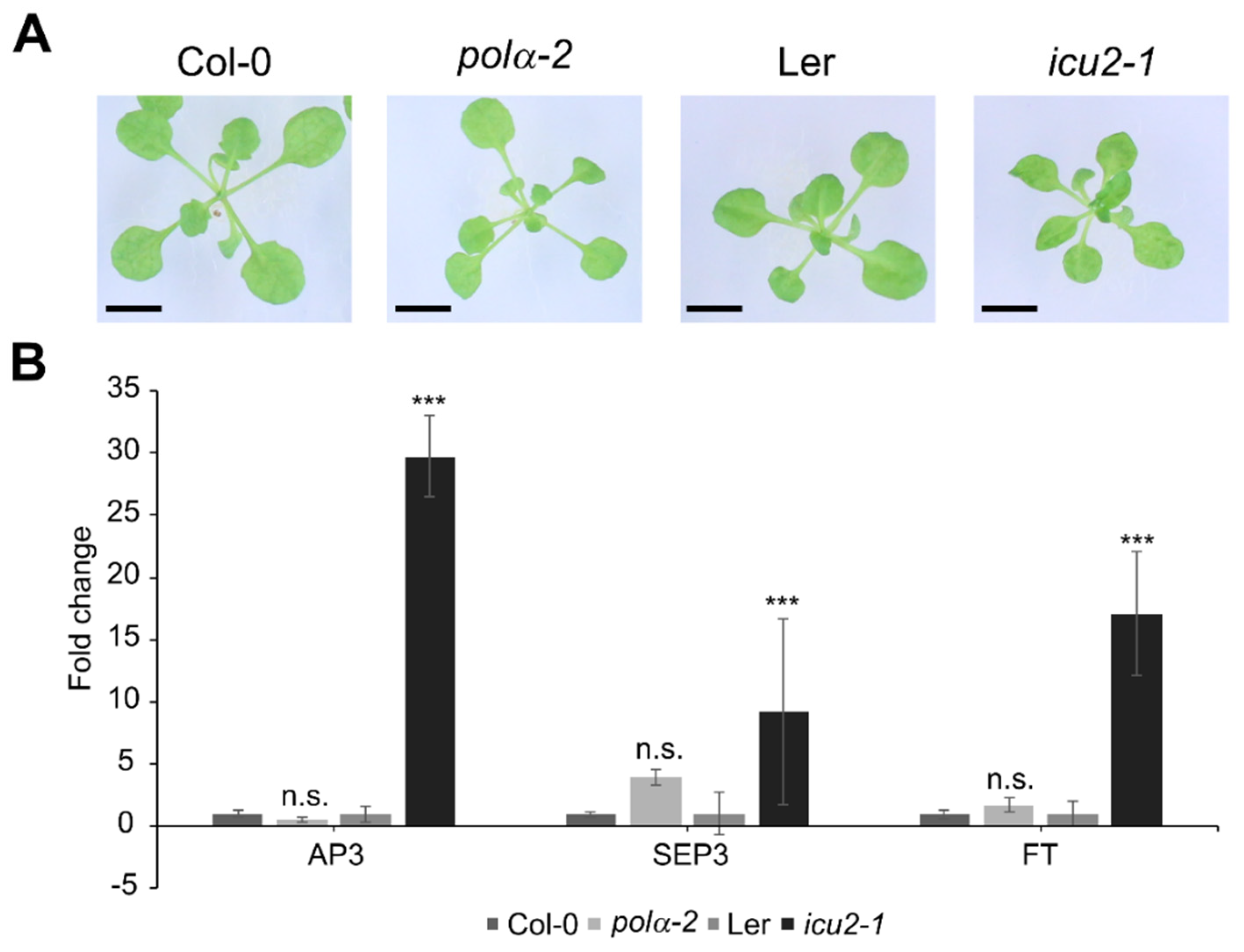
2.2. The Polα-2 Mutant Is Distinct from Earlier Described Polymerase Mutants
2.3. The Polα-2 Mutation Causes a Prolongation of the S-Phase and Activation of the DNA Damage Response
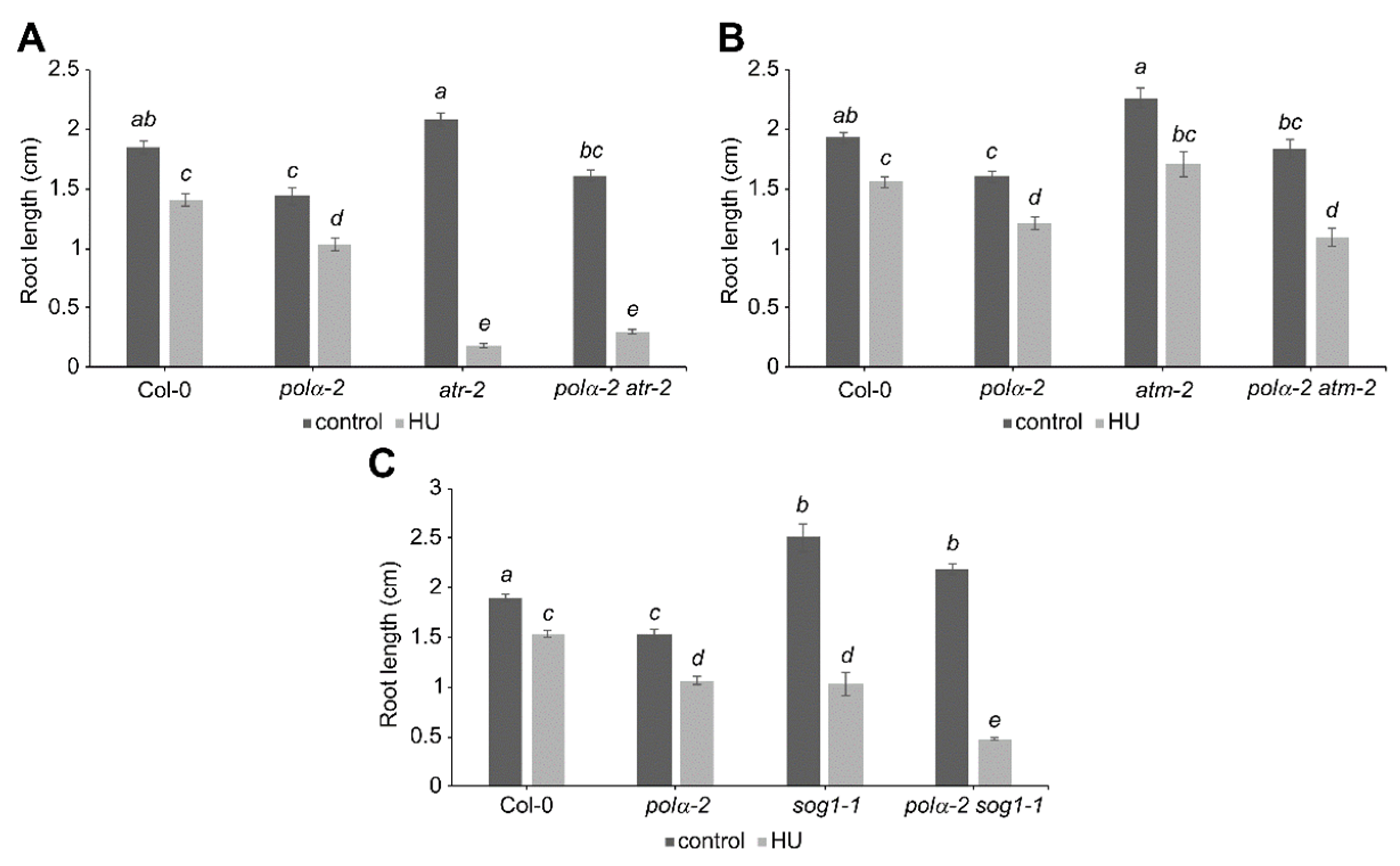
2.4. Polα-2 Cannot Rescues HU Sensitivity of ATRKO and SOG1KO Plants
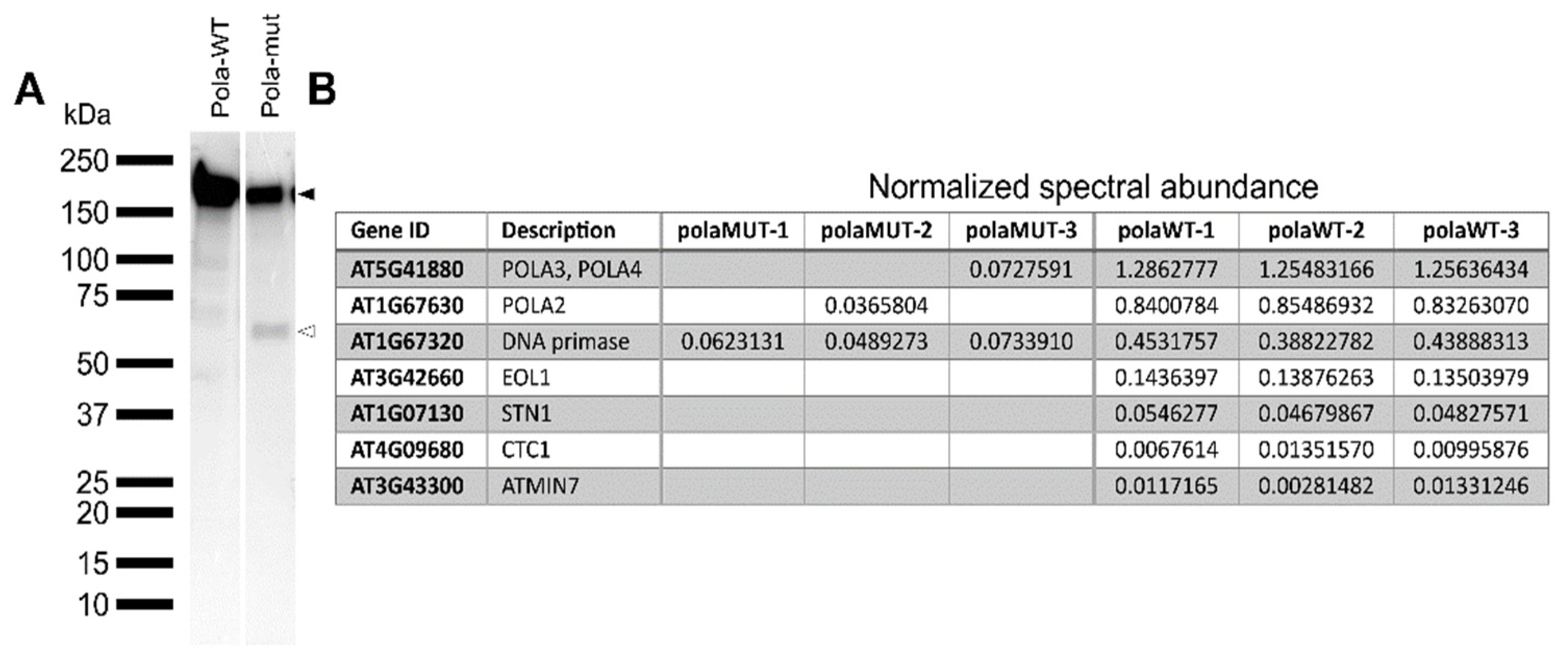
2.5. The Mutated Polymerase α Protein Is Unstable and Loses Interaction with the Other Subunits
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. EMS Mutagenesis and Mapping
4.3. RNA Extraction and qRT-PCR
4.4. Tandem Affinity Purification and Western Blot
4.5. EdU Labelling
4.6. Modelling
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costas, C.; de la Paz Sanchez, M.; Stroud, H.; Yu, Y.; Oliveros, J.C.; Feng, S.H.; Benguria, A.; Lopez-Vidriero, I.; Zhang, X.; Solano, R.; et al. Genome-wide mapping of Arabidopsis thaliana origins of DNA replication and their associated epigenetic marks. Nat. Struct. Mol. Biol. 2011, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Burgers, P.M.J.; Kunkel, T.A. Eukaryotic DNA Replication Fork. Annu. Rev. Biochem. 2017, 86, 417–438. [Google Scholar] [CrossRef] [PubMed]
- Arezi, B.; Kuchta, R.D. Eukaryotic DNA primase. Trends Biochem. Sci. 2000, 25, 572–576. [Google Scholar] [CrossRef]
- Pellegrini, L. The Pol alpha-primase complex. Sub-Cell. Biochem. 2012, 62, 157–169. [Google Scholar] [CrossRef]
- Stillman, B. DNA polymerases at the replication fork in eukaryotes. Mol. Cell 2008, 30, 259–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.E.; Klassen, R.; Prakash, L.; Prakash, S. A Major Role of DNA Polymerase delta in Replication of Both the Leading and Lagging DNA Strands. Mol. Cell 2015, 59, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Del Olmo, I.; Lopez-Gonzalez, L.; Martin-Trillo, M.M.; Martinez-Zapater, J.M.; Pineiro, M.; Jarillo, J.A. EARLY IN SHORT DAYS 7 (ESD7) encodes the catalytic subunit of DNA polymerase epsilon and is required for flowering repression through a mechanism involving epigenetic gene silencing. Plant, J. Cell Mol. Biol. 2010, 61, 623–636. [Google Scholar] [CrossRef]
- Liu, J.; Ren, X.; Yin, H.; Wang, Y.; Xia, R.; Wang, Y.; Gong, Z. Mutation in the catalytic subunit of DNA polymerase alpha influences transcriptional gene silencing and homologous recombination in Arabidopsis. Plant, J. Cell Mol. Biol. 2010, 61, 36–45. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, S.; Cheng, J.; Lai, J.; Zhu, J.K.; Gong, Z. The Second Subunit of DNA Polymerase Delta Is Required for Genomic Stability and Epigenetic Regulation. Plant Physiol. 2016, 171, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Pedroza-Garcia, J.A.; De Veylder, L.; Raynaud, C. Plant DNA Polymerases. Int. J. Mol. Sci. 2019, 20, 4814. [Google Scholar] [CrossRef] [Green Version]
- Barrero, J.M.; Gonzalez-Bayon, R.; del Pozo, J.C.; Ponce, M.R.; Micol, J.L. INCURVATA2 encodes the catalytic subunit of DNA Polymerase alpha and interacts with genes involved in chromatin-mediated cellular memory in Arabidopsis thaliana. Plant. Cell 2007, 19, 2822–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglesias, F.M.; Bruera, N.A.; Dergan-Dylon, S.; Marino-Buslje, C.; Lorenzi, H.; Mateos, J.L.; Turck, F.; Coupland, G.; Cerdan, P.D. The arabidopsis DNA polymerase delta has a role in the deposition of transcriptionally active epigenetic marks, development and flowering. PLoS Genet. 2015, 11, e1004975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Zhang, X.; Liu, J.; Wang, Y.; He, J.; Yang, T.; Hong, X.; Yang, Q.; Gong, Z. Epigenetic regulation, somatic homologous recombination, and abscisic acid signaling are influenced by DNA polymerase epsilon mutation in Arabidopsis. Plant. Cell 2009, 21, 386–402. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Saini, S.; Singh, J. Chromodomain protein Swi6-mediated role of DNA polymerase alpha in establishment of silencing in fission Yeast. J. Biol. Chem. 2001, 276, 47814–47821. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, J.; Allshire, R.C.; Klar, A.J.; Grewal, S.I. A role for DNA polymerase alpha in epigenetic control of transcriptional silencing in fission yeast. EMBO J. 2001, 20, 2857–2866. [Google Scholar] [CrossRef] [Green Version]
- Branzei, D.; Foiani, M. The checkpoint response to replication stress. DNA Repair 2009, 8, 1038–1046. [Google Scholar] [CrossRef]
- Hu, Z.; Cools, T.; De Veylder, L. Mechanisms Used by Plants to Cope with DNA Damage. Annu. Rev. Plant. Boil. 2016, 67, 439–462. [Google Scholar] [CrossRef] [PubMed]
- Byun, T.S.; Pacek, M.; Yee, M.C.; Walter, J.C.; Cimprich, K.A. Functional uncoupling of MCM helicase and DNA polymerase activities activates the ATR-dependent checkpoint. Genes Dev. 2005, 19, 1040–1052. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.; Cotta-Ramusino, C.; Pellicioli, A.; Liberi, G.; Plevani, P.; Muzi-Falconi, M.; Newlon, C.S.; Foiani, M. The DNA replication checkpoint response stabilizes stalled replication forks. Nature 2001, 412, 557–561. [Google Scholar] [CrossRef]
- Beck, H. Cyclin-dependent kinase suppression by WEE1 kinase protects the genome through control of replication initiation and nucleotide consumption. Mol. Cell. Biol. 2012, 32, 4226–4236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culligan, K.; Tissier, A.; Britt, A. ATR regulates a G2-phase cell-cycle checkpoint in Arabidopsis thaliana. Plant. Cell 2004, 16, 1091–1104. [Google Scholar] [CrossRef] [Green Version]
- De Schutter, K. Arabidopsis WEE1 kinase controls cell cycle arrest in response to activation of the DNA integrity checkpoint. Plant. Cell 2007, 19, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Schneeberger, K.; Ossowski, S.; Lanz, C.; Juul, T.; Petersen, A.H.; Nielsen, K.L.; Jorgensen, J.E.; Weigel, D.; Andersen, S.U. SHOREmap: Simultaneous mapping and mutation identification by deep sequencing. Nat. Methods 2009, 6, 550–551. [Google Scholar] [CrossRef]
- Coloma, J.; Johnson, R.E.; Prakash, L.; Prakash, S.; Aggarwal, A.K. Human DNA polymerase alpha in binary complex with a DNA:DNA template-primer. Sci. Rep. 2016, 6, 23784. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Hasegawa, J.; Matsunaga, S. The boundary of the meristematic and elongation zones in roots: Endoreduplication precedes rapid cell expansion. Sci. Rep. 2013, 3, 2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cools, T. The Arabidopsis thaliana checkpoint kinase WEE1 protects against premature vascular differentiation during replication stress. Plant. Cell 2011, 23, 1435–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eekhout, T. G2/M-checkpoint activation in fasciata1 rescues an aberrant S-phase checkpoint but causes genome instability. Plant. Physiol. 2021. [Google Scholar] [CrossRef]
- Yi, D.; Alvim Kamei, C.L.; Cools, T.; Vanderauwera, S.; Takahashi, N.; Okushima, Y.; Eekhout, T.; Yoshiyama, K.O.; Larkin, J.; Van den Daele, H.; et al. The Arabidopsis SIAMESE-RELATED cyclin-dependent kinase inhibitors SMR5 and SMR7 regulate the DNA damage checkpoint in response to reactive oxygen species. Plant. Cell 2014, 26, 296–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Cools, T.; Kalhorzadeh, P.; Heyman, J.; De Veylder, L. Deficiency of the Arabidopsis helicase RTEL1 triggers a SOG1-dependent replication checkpoint in response to DNA cross-links. Plant. Cell 2015, 27, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Van Leene, J.; Eeckhout, D.; Cannoot, B.; De Winne, N.; Persiau, G.; Van De Slijke, E.; Vercruysse, L.; Dedecker, M.; Verkest, A.; Vandepoele, K.; et al. An improved toolbox to unravel the plant cellular machinery by tandem affinity purification of Arabidopsis protein complexes. Nature protocols 2015, 10, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Derboven, E.; Ekker, H.; Kusenda, B.; Bulankova, P.; Riha, K. Role of STN1 and DNA polymerase alpha in telomere stability and genome-wide replication in Arabidopsis. PLoS Genet. 2014, 10, e1004682. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y. Ctf4-related protein recruits LHP1-PRC2 to maintain H3K27me3 levels in dividing cells in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, 4833–4838. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Xu, Y.J. The Cell Killing Mechanisms of Hydroxyurea. Genes 2016, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Michael, W.M.; Ott, R.; Fanning, E.; Newport, J. Activation of the DNA replication checkpoint through RNA synthesis by primase. Science 2000, 289, 2133–2137. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Nau, W.M. A conformational flexibility scale for amino acids in peptides. Angew. Chem. (Int. Ed. Engl.) 2003, 42, 2269–2272. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.C. A Ctf4 trimer couples the CMG helicase to DNA polymerase alpha in the eukaryotic replisome. Nature 2014, 510, 293–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Ines, O.; Degroote, F.; Goubely, C.; Amiard, S.; Gallego, M.E.; White, C.I. Meiotic recombination in Arabidopsis is catalysed by DMC1, with RAD51 playing a supporting role. PLoS Genet. 2013, 9, e1003787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reidt, W.; Wurz, R.; Wanieck, K.; Chu, H.H.; Puchta, H. A homologue of the breast cancer-associated gene BARD1 is involved in DNA repair in plants. EMBO J. 2006, 25, 4326–4337. [Google Scholar] [CrossRef]
- Kolinjivadi, A.M.; Sannino, V.; de Antoni, A.; Techer, H.; Baldi, G.; Costanzo, V. Moonlighting at replication forks—A new life for homologous recombination proteins BRCA1, BRCA2 and RAD51. FEBS Lett. 2017, 591, 1083–1100. [Google Scholar] [CrossRef]
- Pedroza-Garcia, J.A. Function of the Plant DNA Polymerase Epsilon in Replicative Stress Sensing, a Genetic Analysis. Plant. Physiol. 2017, 173, 1735–1749. [Google Scholar] [CrossRef] [Green Version]
- Eekhout, T.; Kalhorzadeh, P.; De Veylder, L. Lack of RNase H2 activity rescues HU-sensitivity of WEE1 deficient plants. Plant. Signal. Behav. 2015, 10, e1001226. [Google Scholar] [CrossRef] [PubMed]
- Kalhorzadeh, P.; Hu, Z.; Cools, T.; Amiard, S.; Willing, E.M.; De Winne, N.; Gevaert, K.; De Jaeger, G.; Schneeberger, K.; White, C.I.; et al. Arabidopsis thaliana RNase H2 deficiency counteracts the needs for the WEE1 checkpoint kinase but triggers genome instability. Plant. Cell 2014, 26, 3680–3692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, V.; Bruchet, H.; Camescasse, D.; Granier, F.; Bouchez, D.; Tissier, A. AtATM is essential for meiosis and the somatic response to DNA damage in plants. Plant. Cell 2003, 15, 119–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshiyama, K.; Conklin, P.A.; Huefner, N.D.; Britt, A.B. Suppressor of gamma response 1 (SOG1) encodes a putative transcription factor governing multiple responses to DNA damage. Proc. Natl. Acad. Sci. USA 2009, 106, 12843–12848. [Google Scholar] [CrossRef] [Green Version]
- Ossowski, S.; Schneeberger, K.; Clark, R.M.; Lanz, C.; Warthmann, N.; Weigel, D. Sequencing of natural strains of Arabidopsis thaliana with short reads. Genome Res. 2008, 18, 2024–2033. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eekhout, T.; Pedroza-Garcia, J.A.; Kalhorzadeh, P.; De Jaeger, G.; De Veylder, L. A Mutation in DNA Polymerase α Rescues WEE1KO Sensitivity to HU. Int. J. Mol. Sci. 2021, 22, 9409. https://doi.org/10.3390/ijms22179409
Eekhout T, Pedroza-Garcia JA, Kalhorzadeh P, De Jaeger G, De Veylder L. A Mutation in DNA Polymerase α Rescues WEE1KO Sensitivity to HU. International Journal of Molecular Sciences. 2021; 22(17):9409. https://doi.org/10.3390/ijms22179409
Chicago/Turabian StyleEekhout, Thomas, José Antonio Pedroza-Garcia, Pooneh Kalhorzadeh, Geert De Jaeger, and Lieven De Veylder. 2021. "A Mutation in DNA Polymerase α Rescues WEE1KO Sensitivity to HU" International Journal of Molecular Sciences 22, no. 17: 9409. https://doi.org/10.3390/ijms22179409
APA StyleEekhout, T., Pedroza-Garcia, J. A., Kalhorzadeh, P., De Jaeger, G., & De Veylder, L. (2021). A Mutation in DNA Polymerase α Rescues WEE1KO Sensitivity to HU. International Journal of Molecular Sciences, 22(17), 9409. https://doi.org/10.3390/ijms22179409