
Peptide-Assisted Nucleic Acid Delivery Systems on the Rise

 , and
, and
Abstract
1. Introduction
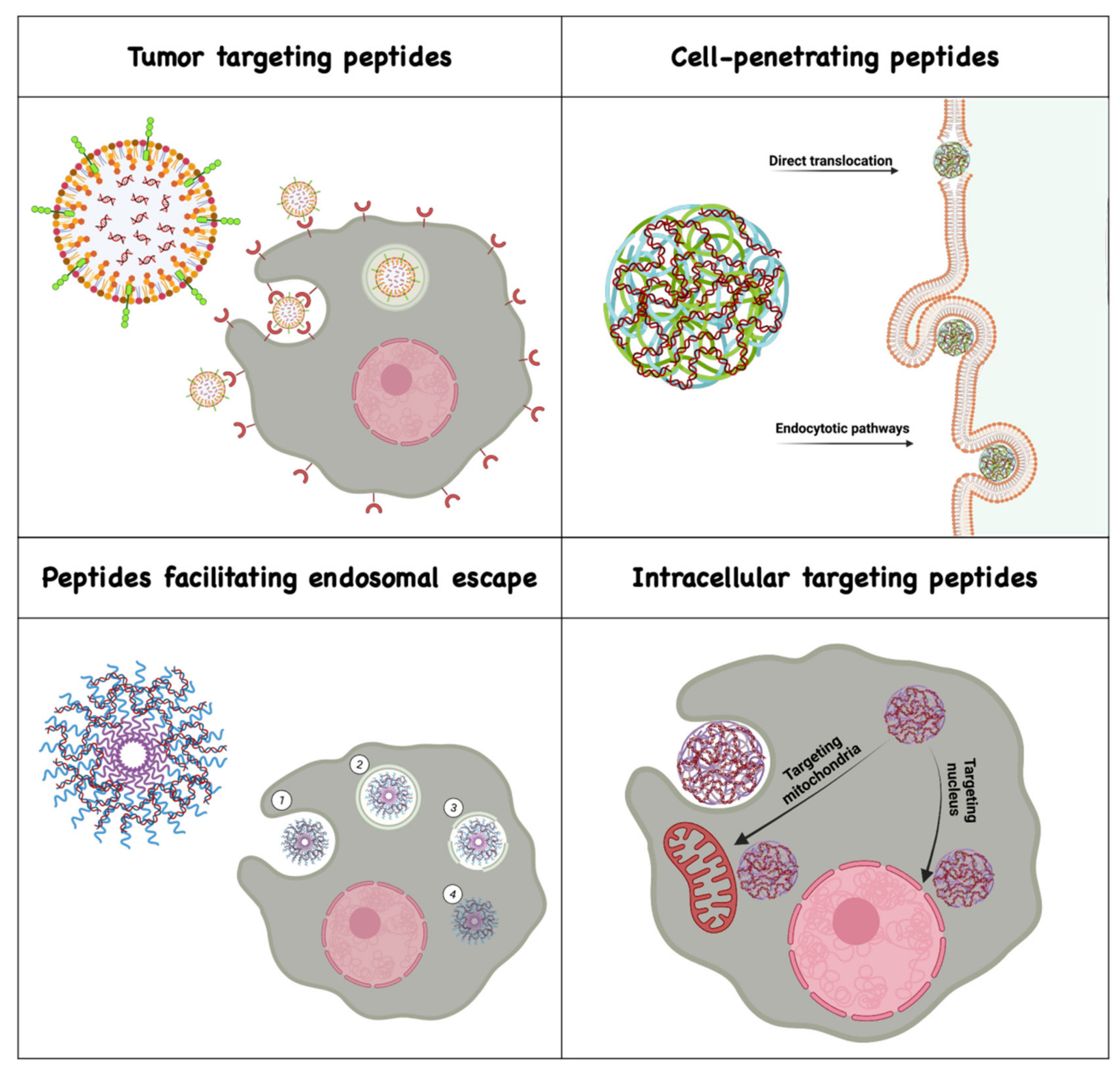
2. Peptide-Guided Delivery of Nucleic Acids across Biological Barriers
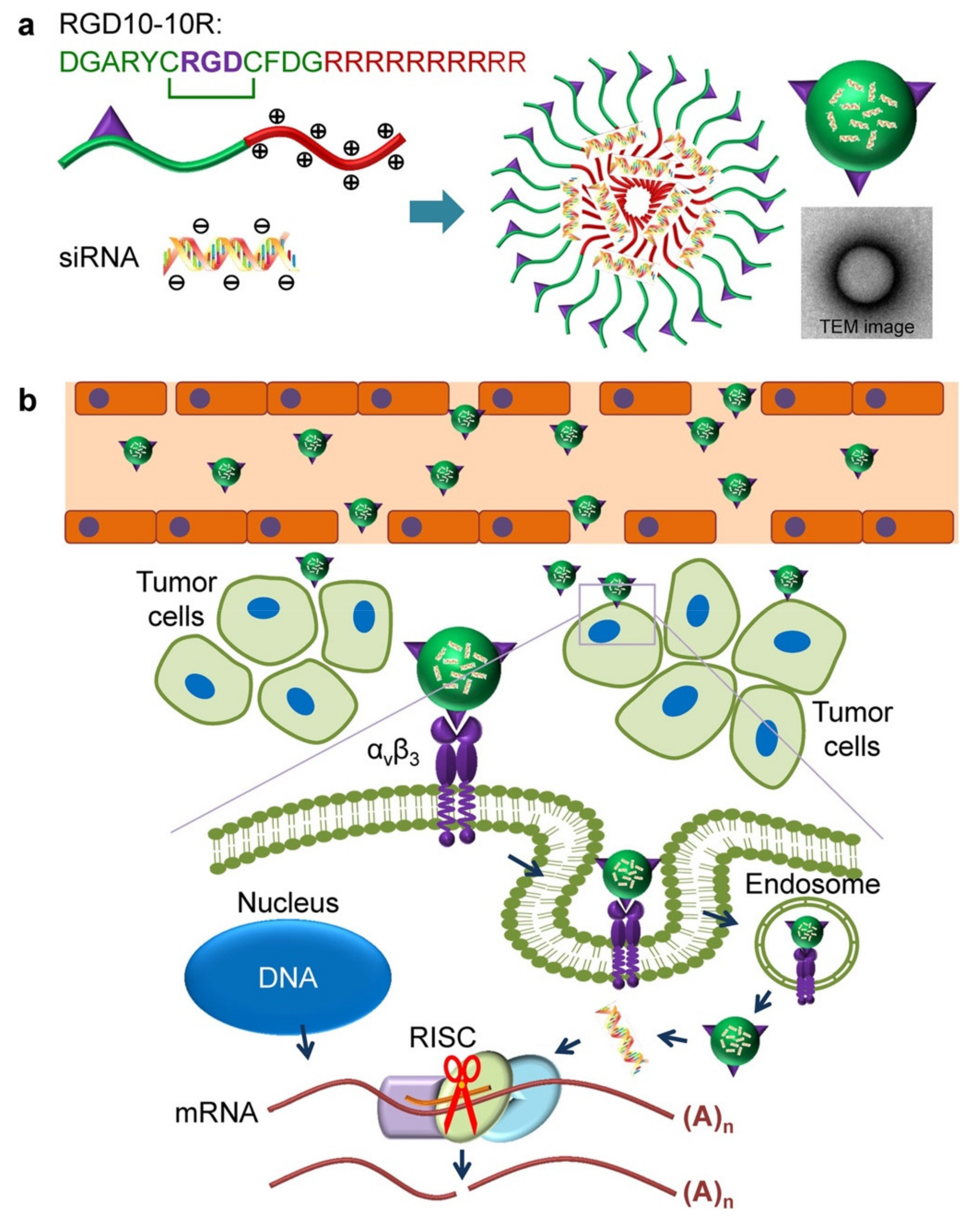
2.1. Tumor-Targeting Peptides
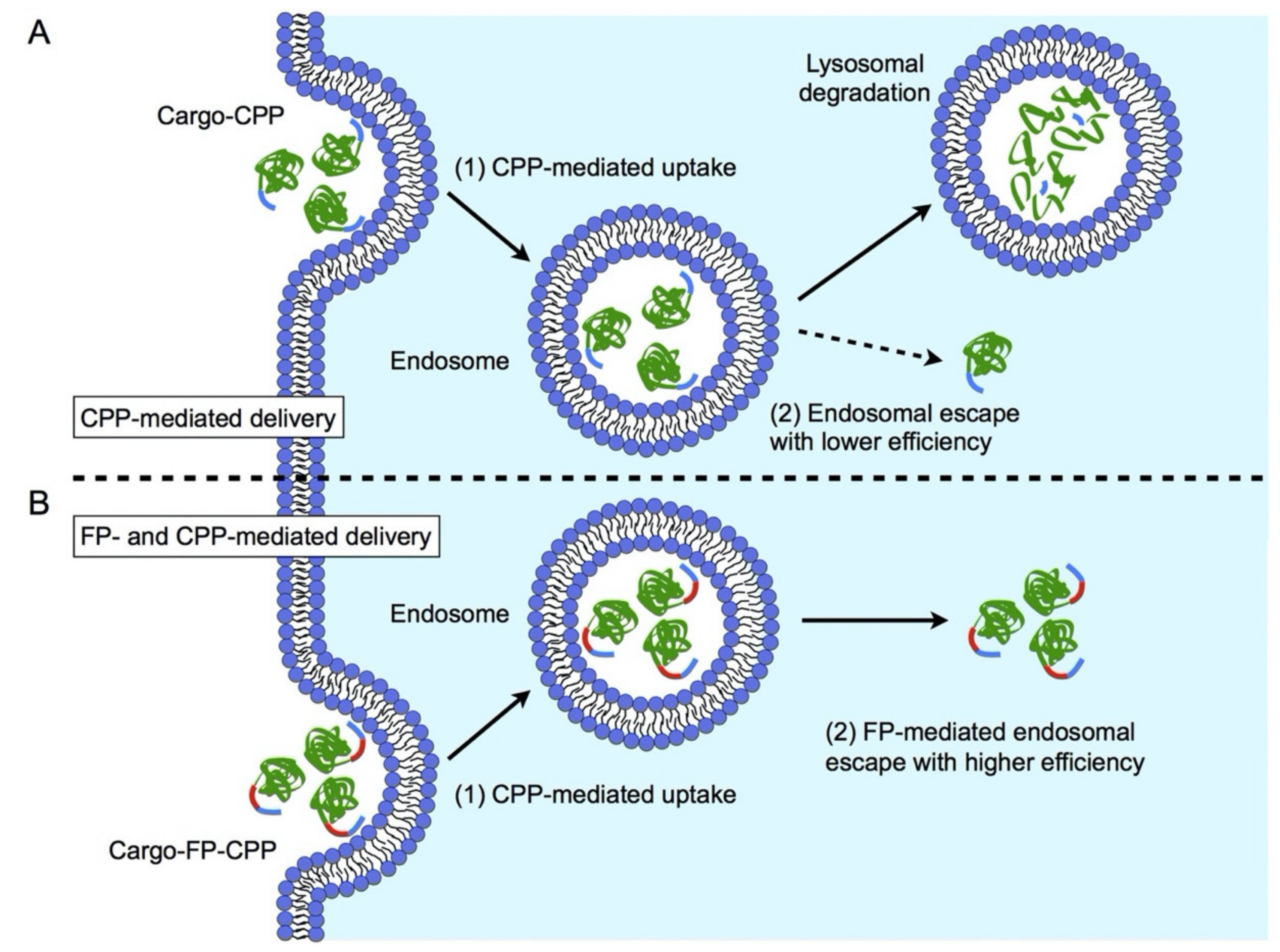
2.2. Cell-Penetrating Peptides
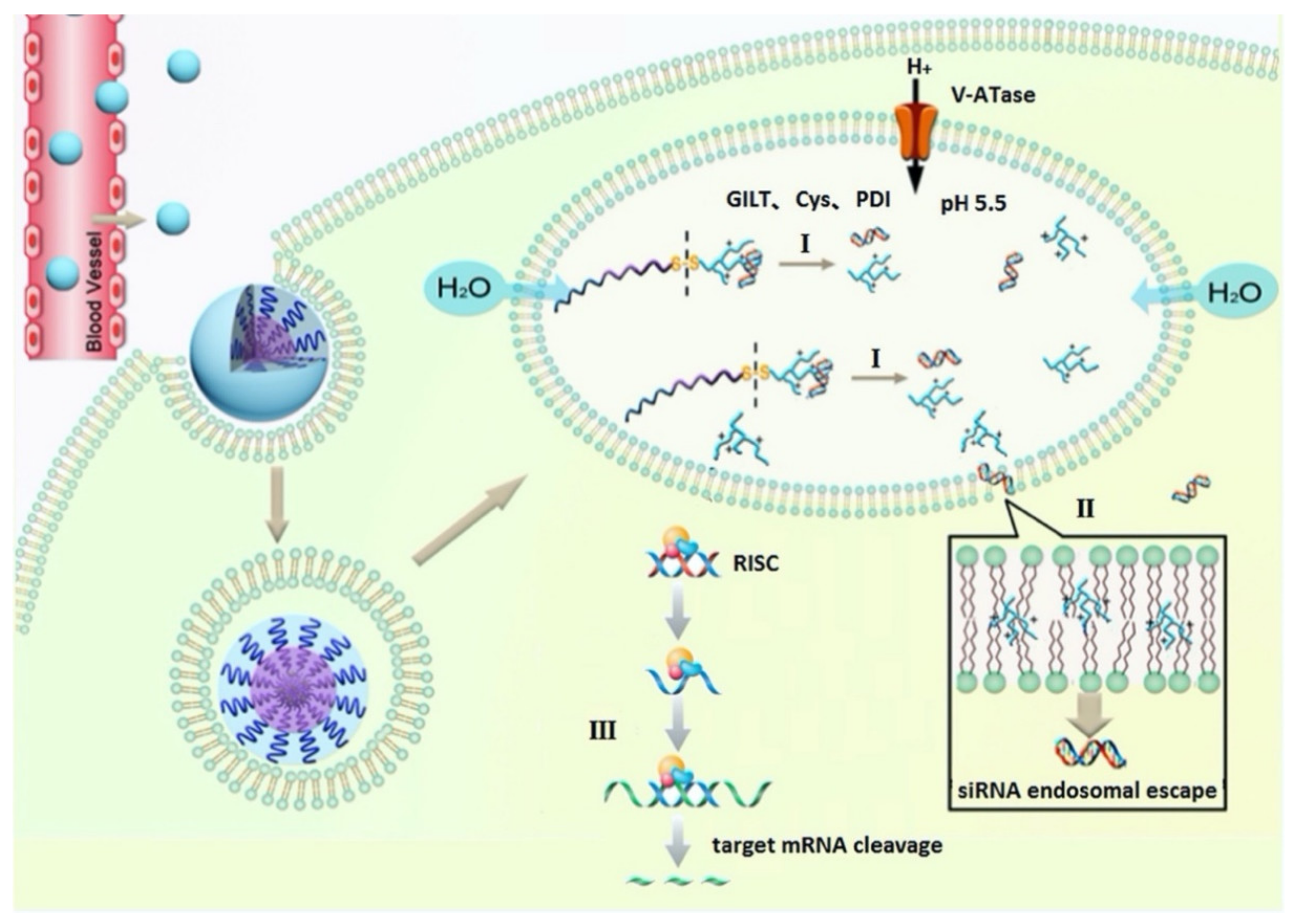
2.3. Peptides Facilitating Endosomal Escape
2.3.1. Fusogenic Peptides
2.3.2. Histidine-Rich Peptides
2.4. Peptides Assisting Delivery to Subcellular Organelles
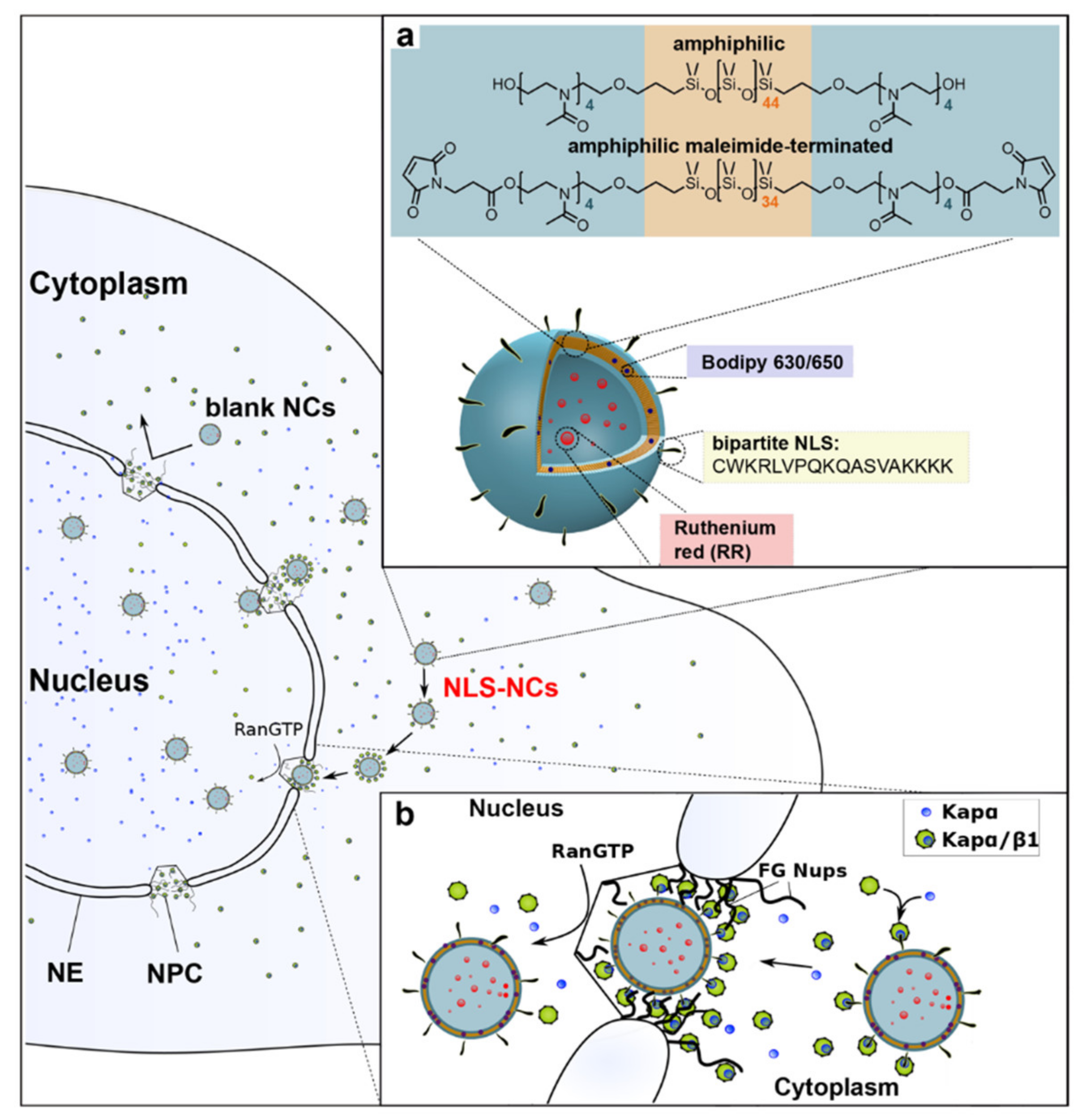
2.4.1. Nuclear Localization Signals
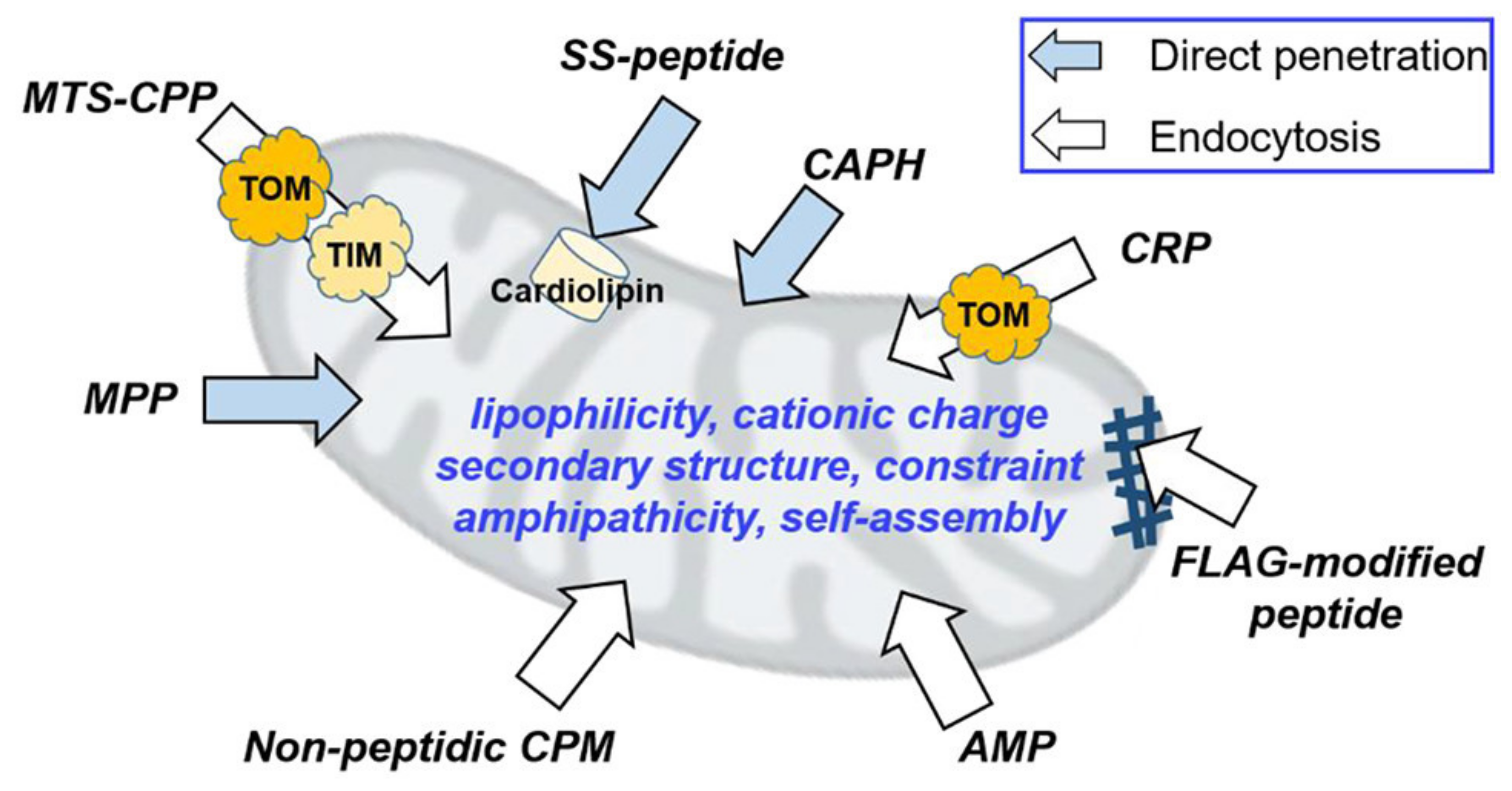
2.4.2. Mitochondrial Delivery
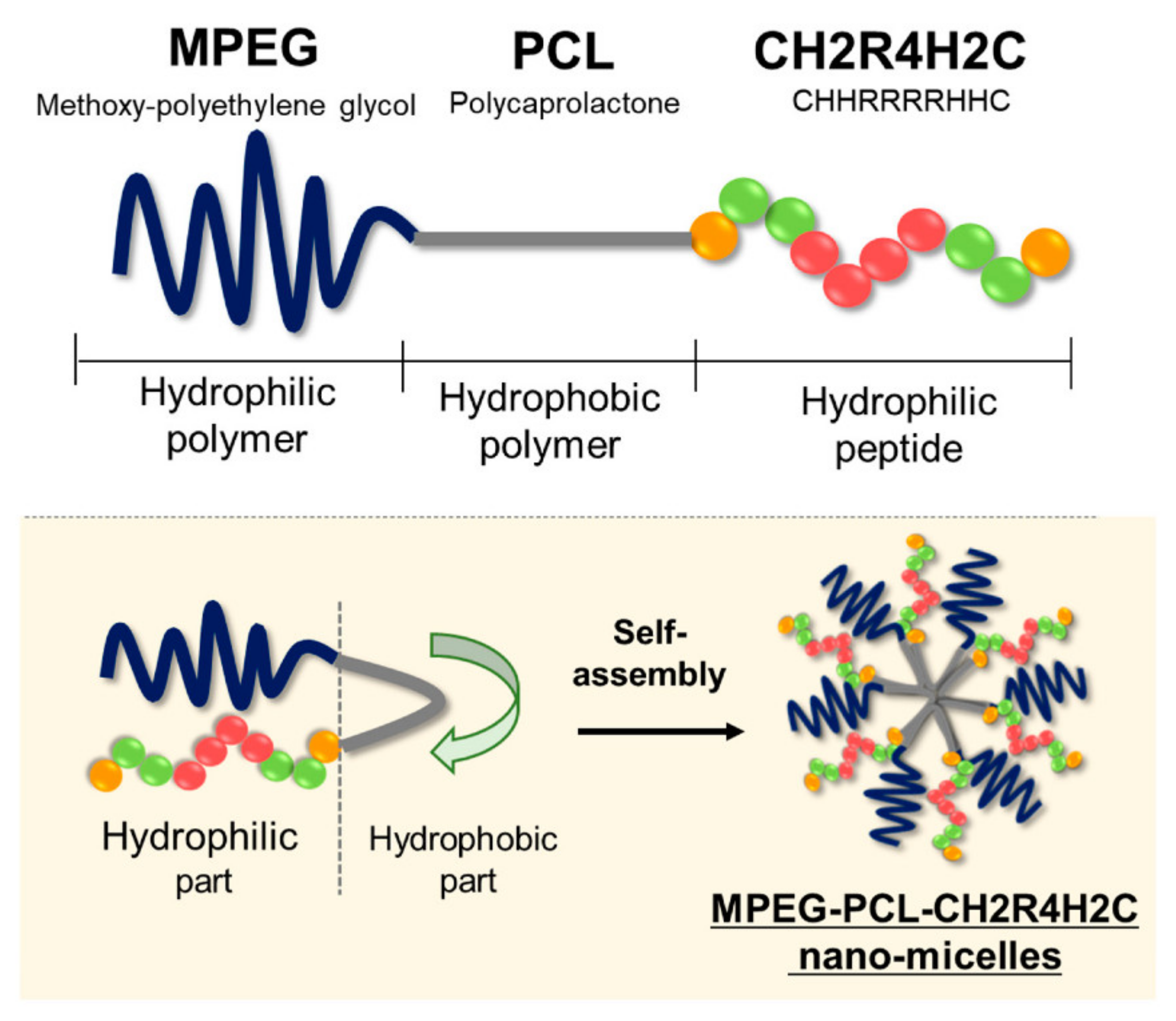
3. Peptide-Related Nano-Assemblies for Nucleic Acid Delivery
Stimuli-Responsive Gene Delivery Systems
4. Combinatorial Approach for Advanced Nucleic Acid Delivery
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gillmore, J.D.; Gane, E.; Taubel, J.; Kao, J.; Fontana, M.; Maitland, M.L.; Seitzer, J.; O’Connell, D.; Walsh, K.R.; Wood, K.; et al. CRISPR-Cas9 In Vivo Gene Editing for Transthyretin Amyloidosis. N. Engl. J. Med. 2021, 385, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, D.; Gutkin, A.; Kedmi, R.; Ramishetti, S.; Veiga, N.; Jacobi, A.M.; Schubert, M.S.; Friedmann-Morvinski, D.; Cohen, Z.R.; Behlke, M.A.; et al. CRISPR-Cas9 Genome Editing Using Targeted Lipid Nanoparticles for Cancer Therapy. Sci. Adv. 2020, 6, eabc9450. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines—A New Era in Vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An MRNA Vaccine against SARS-CoV-2 - Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Fogel, D.B. Factors Associated with Clinical Trials That Fail and Opportunities for Improving the Likelihood of Success: A Review. Contemp. Clin. Trials Commun. 2018, 11, 156–164. [Google Scholar] [CrossRef]
- Seyhan, A.A. Lost in Translation: The Valley of Death across Preclinical and Clinical Divide—Identification of Problems and Overcoming Obstacles. Transl. Med. Commun. 2019, 4, 18. [Google Scholar] [CrossRef]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of Nanoparticle Design for Overcoming Biological Barriers to Drug Delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Muttenthaler, M.; King, G.F.; Adams, D.J.; Alewood, P.F. Trends in Peptide Drug Discovery. Nat. Rev. Drug Discov. 2021, 20, 309–325. [Google Scholar] [CrossRef]
- Kumari, S.; Mg, S.; Mayor, S. Endocytosis Unplugged: Multiple Ways to Enter the Cell. Cell Res. 2010, 20, 256–275. [Google Scholar] [CrossRef] [PubMed]
- Parodi, A.; Corbo, C.; Cevenini, A.; Molinaro, R.; Palomba, R.; Pandolfi, L.; Agostini, M.; Salvatore, F.; Tasciotti, E. Enabling Cytoplasmic Delivery and Organelle Targeting by Surface Modification of Nanocarriers. Nanomedicine 2015, 10, 1923–1940. [Google Scholar] [CrossRef] [PubMed]
- Petros, R.A.; DeSimone, J.M. Strategies in the Design of Nanoparticles for Therapeutic Applications. Nat. Rev. Drug Discov. 2010, 9, 615–627. [Google Scholar] [CrossRef]
- Ruoslahti, E. Peptides as Targeting Elements and Tissue Penetration Devices for Nanoparticles. Adv. Mater. 2012, 24, 3747–3756. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Bu, J.; Kubiatowicz, L.J.; Chen, S.S.; Kim, Y.; Hong, S. Peptide–Nanoparticle Conjugates: A next Generation of Diagnostic and Therapeutic Platforms? Nano Converg. 2018, 5, 38. [Google Scholar] [CrossRef] [PubMed]
- Hossen, S.; Hossain, M.K.; Basher, M.K.; Mia, M.N.H.; Rahman, M.T.; Uddin, M.J. Smart Nanocarrier-Based Drug Delivery Systems for Cancer Therapy and Toxicity Studies: A Review. J. Adv. Res. 2019, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.-L.; Harris, J.L.; Khanna, K.K.; Hong, J.-H. A Comprehensive Review on Current Advances in Peptide Drug Development and Design. Int. J. Mol. Sci. 2019, 20, 2383. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.M.; Iegre, J.; Donovan, D.H.O.; Halvarsson, M.Ö.; Spring, D.R. Peptides as a Platform for Targeted Therapeutics for Cancer: Peptide–Drug Conjugates (PDCs). Chem. Soc. Rev. 2021, 50, 1480–1494. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering Precision Nanoparticles for Drug Delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef]
- Sandra, F.; Khaliq, N.U.; Sunna, A.; Care, A. Developing Protein-Based Nanoparticles as Versatile Delivery Systems for Cancer Therapy and Imaging. Nanomaterials 2019, 9, 1329. [Google Scholar] [CrossRef] [PubMed]
- Avci, F.G.; Sariyar Akbulut, B.; Ozkirimli, E. Membrane Active Peptides and Their Biophysical Characterization. Biomolecules 2018, 8, 77. [Google Scholar] [CrossRef]
- Tarvirdipour, S.; Schoenenberger, C.-A.; Benenson, Y.; Palivan, C.G. A Self-Assembling Amphiphilic Peptide Nanoparticle for the Efficient Entrapment of DNA Cargoes up to 100 Nucleotides in Length. Soft Matter. 2020, 16, 1678–1691. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Navarro, M.; Teixidó, M.; Giralt, E. Jumping Hurdles: Peptides Able To Overcome Biological Barriers. Acc. Chem. Res. 2017, 50, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Cabral, B.P.; da Graça Derengowski Fonseca, M.; Mota, F.B. The Recent Landscape of Cancer Research Worldwide: A Bibliometric and Network Analysis. Oncotarget 2018, 9, 30474–30484. [Google Scholar] [CrossRef]
- Ruoslahti, E. Specialization of Tumour Vasculature. Nat. Rev. Cancer 2002, 2, 83–90. [Google Scholar] [CrossRef]
- Marelli, U.K.; Rechenmacher, F.; Sobahi, T.R.A.; Mas-Moruno, C.; Kessler, H. Tumor Targeting via Integrin Ligands. Front. Oncol. 2013, 3. [Google Scholar] [CrossRef]
- Brown, K.C. Peptidic Tumor Targeting Agents: The Road from Phage Display Peptide Selections to Clinical Applications. Curr. Pharm. Des. 2010, 16, 1040–1054. [Google Scholar] [CrossRef]
- Zhao, N.; Qin, Y.; Liu, H.; Cheng, Z. Tumor-Targeting Peptides: Ligands for Molecular Imaging and Therapy. Anticancer Agents Med. Chem. 2018, 18, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.J.; Cho, C.H. Peptides as Targeting Probes against Tumor Vasculature for Diagnosis and Drug Delivery. J. Transl. Med. 2012, 10, S1. [Google Scholar] [CrossRef]
- Ko, J.K.; Auyeung, K.K. Identification of Functional Peptides from Natural and Synthetic Products on Their Anticancer Activities by Tumor Targeting. Curr. Med. Chem. 2014, 21, 2346–2356. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Santino, F.; Giacomini, D.; Gentilucci, L. Integrin-Targeting Peptides for the Design of Functional Cell-Responsive Biomaterials. Biomedicines 2020, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.-H.; Opadele, A.E.; Onodera, Y.; Nam, J.-M. Targeting Integrins in Cancer Nanomedicine: Applications in Cancer Diagnosis and Therapy. Cancers 2019, 11, 1783. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, X.; Huang, W.; Cheng, Q.; Zheng, S.; Guo, S.; Cao, H.; Liang, X.-J.; Du, Q.; Liang, Z. Systemic Administration of SiRNA via CRGD-Containing Peptide. Sci. Rep. 2015, 5, 12458. [Google Scholar] [CrossRef] [PubMed]
- Roth, L.; Agemy, L.; Kotamraju, V.R.; Braun, G.; Teesalu, T.; Sugahara, K.N.; Hamzah, J.; Ruoslahti, E. Transtumoral Targeting Enabled by a Novel Neuropilin-Binding Peptide. Oncogene 2012, 31, 3754–3763. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, S.; Zhang, Y.; Li, H.; He, H.; Dai, J.; Jiang, T.; Ji, W.; Geng, D.; Elzatahry, A.A.; et al. Magnetic Mesoporous Nanospheres Anchored with LyP-1 as an Efficient Pancreatic Cancer Probe. Biomaterials 2017, 115, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Zhao, L.; Zhu, M.; Zhao, J. Recent Progress in LyP-1-Based Strategies for Targeted Imaging and Therapy. Drug Deliv. 2019, 26, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.M.; Sun, Z.; Ayat, N.; Schilb, A.; Liu, X.; Jiang, H.; Sun, D.; Scheidt, J.; Qian, V.; He, S.; et al. Systemic Delivery of Tumor-Targeting SiRNA Nanoparticles against an Oncogenic LncRNA Facilitates Effective Triple-Negative Breast Cancer Therapy. Bioconjug. Chem. 2019, 30, 907–919. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Cen, B.; Liao, L.; Wang, Z.; Qin, Y.; Wu, Z.; Liao, W.; Zhang, Z.; Ji, A. A Tumor-Targeting CRGD-EGFR SiRNA Conjugate and Its Anti-Tumor Effect on Glioblastoma in Vitro and in Vivo. Drug Deliv. 2017, 24, 471–481. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Zhou, X.; Sun, J.; Fan, X.; Guan, Z.; Zhang, L.; Yang, Z. Construction of a Targeting Nanoparticle of 3′,3″-Bis-Peptide-SiRNA Conjugate/Mixed Lipid with Postinserted DSPE-PEG2000-CRGD. Mol. Pharm. 2019, 16. [Google Scholar] [CrossRef]
- Lo, J.H.; Hao, L.; Muzumdar, M.D.; Raghavan, S.; Kwon, E.J.; Pulver, E.M.; Hsu, F.; Aguirre, A.J.; Wolpin, B.M.; Fuchs, C.S.; et al. IRGD-Guided Tumor-Penetrating Nanocomplexes for Therapeutic SiRNA Delivery to Pancreatic Cancer. Mol. Cancer Ther. 2018, 17, 2377–2388. [Google Scholar] [CrossRef]
- Zhou, Y.; Yuan, Y.; Liu, M.; Hu, X.; Quan, Y.; Chen, X. Tumor-Specific Delivery of KRAS SiRNA with IRGD-Exosomes Efficiently Inhibits Tumor Growth. ExRNA 2019, 1, 28. [Google Scholar] [CrossRef]
- Xia, Y.; Xu, T.; Wang, C.; Li, Y.; Lin, Z.; Zhao, M.; Zhu, B. Novel Functionalized Nanoparticles for Tumor-Targeting Co-Delivery of Doxorubicin and SiRNA to Enhance Cancer Therapy. Int. J. Nanomed. 2017, 13, 143–159. [Google Scholar] [CrossRef]
- Bai, Y.; Li, Z.; Liu, L.; Sun, T.; Fan, X.; Wang, T.; Gou, Z.; Tan, S. Tumor-Targeting Peptide for Redox-Responsive Pt Prodrug and Gene Codelivery and Synergistic Cancer Chemotherapy. ACS Appl. Bio Mater. 2019, 2, 1420–1426. [Google Scholar] [CrossRef]
- Sharma, M.; El-Sayed, N.S.; Do, H.; Parang, K.; Tiwari, R.K.; Aliabadi, H.M. Tumor-Targeted Delivery of SiRNA Using Fatty Acyl-CGKRK Peptide Conjugates. Sci. Rep. 2017, 7, 6993. [Google Scholar] [CrossRef]
- Li, Y.; Wang, H.; Wang, K.; Hu, Q.; Yao, Q.; Shen, Y.; Yu, G.; Tang, G. Targeted Co-Delivery of PTX and TR3 SiRNA by PTP Peptide Modified Dendrimer for the Treatment of Pancreatic Cancer. Small 2017, 13, 1602697. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Qu, C.; Zhou, Y.; Zhang, L.; Chen, M.; Liu, Y.; You, B.; Li, F.; Wang, D.; Zhang, X. Doxorubicin and SiRNA-PD-L1 Co-Delivery with T7 Modified ROS-Sensitive Nanoparticles for Tumor Chemoimmunotherapy. Int. J. Pharm. 2019, 566, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Sun, S.; Varner, J.A.; Howell, S.B.; Ruoslahti, E.; Sailor, M.J. Securing the Payload, Finding the Cell, and Avoiding the Endosome: Peptide-Targeted, Fusogenic Porous Silicon Nanoparticles for Delivery of SiRNA. Adv. Mater. 2019, 31, 1902952. [Google Scholar] [CrossRef]
- Liang, Y.; Peng, J.; Li, N.; Yu-Wai-Man, C.; Wang, Q.; Xu, Y.; Wang, H.; Tagalakis, A.D.; Du, Z. Smart Nanoparticles Assembled by Endogenous Molecules for SiRNA Delivery and Cancer Therapy via CD44 and EGFR Dual-Targeting. Nanomed. Nanotechnol. Biol. Med. 2019, 15, 208–217. [Google Scholar] [CrossRef]
- Lu, Y.; Jiang, W.; Wu, X.; Huang, S.; Huang, Z.; Shi, Y.; Dai, Q.; Chen, J.; Ren, F.; Gao, S. Peptide T7-Modified Polypeptide with Disulfide Bonds for Targeted Delivery of Plasmid DNA for Gene Therapy of Prostate Cancer. Int. J. Nanomed. 2018, 13, 6913–6927. [Google Scholar] [CrossRef]
- Desale, K.; Kuche, K.; Jain, S. Cell-Penetrating Peptides (CPPs): An Overview of Applications for Improving the Potential of Nanotherapeutics. Biomater. Sci. 2021, 9, 1153–1188. [Google Scholar] [CrossRef]
- Torchilin, V.P. Cell Penetrating Peptide-Modified Pharmaceutical Nanocarriers for Intracellular Drug and Gene Delivery. Pept. Sci. 2008, 90, 604–610. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Y.; Zhang, X.; Zhang, W.; Guo, S.; Jin, F. Recent Progress of Cell-Penetrating Peptides as New Carriers for Intracellular Cargo Delivery. J. Control. Release 2014, 174, 126–136. [Google Scholar] [CrossRef]
- Khan, M.M.; Filipczak, N.; Torchilin, V.P. Cell Penetrating Peptides: A Versatile Vector for Co-Delivery of Drug and Genes in Cancer. J. Control. Release 2021, 330, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhou, M.; Shi, H.; Gao, S.; Xie, G.; Zhu, M.; Wu, M.; Chen, J.; Niu, Z. Integration of Cell-Penetrating Peptides with Rod-like Bionanoparticles: Virus-Inspired Gene-Silencing Technology. Nano Lett. 2018, 18, 5453–5460. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Ambrosone, A.; Hernandez, Y.; Tian, F.; McCully, M.; Berry, C.C.; Baptista, P.V.; Tortiglione, C.; de la Fuente, J.M. 15 Years on SiRNA Delivery: Beyond the State-of-the-Art on Inorganic Nanoparticles for RNAi Therapeutics. Nano Today 2015, 10, 421–450. [Google Scholar] [CrossRef]
- Kang, Z.; Meng, Q.; Liu, K. Peptide-Based Gene Delivery Vectors. J. Mater. Chem. B 2019, 7, 1824–1841. [Google Scholar] [CrossRef] [PubMed]
- Madani, F.; Lindberg, S.; Langel, Ü.; Futaki, S.; Gräslund, A. Mechanisms of Cellular Uptake of Cell-Penetrating Peptides. J. Biophys. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, M.; Langel, Ü. Classes and Prediction of Cell-Penetrating Peptides. In Cell-Penetrating Peptides: Methods and Protocols; Langel, Ü., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; pp. 3–19. ISBN 978-1-60761-919-2. [Google Scholar]
- Xie, J.; Bi, Y.; Zhang, H.; Dong, S.; Teng, L.; Lee, R.J.; Yang, Z. Cell-Penetrating Peptides in Diagnosis and Treatment of Human Diseases: From Preclinical Research to Clinical Application. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Trabulo, S.; Cardoso, A.L.; Mano, M.; De Lima, M.C.P. Cell-Penetrating Peptides—Mechanisms of Cellular Uptake and Generation of Delivery Systems. Pharmaceuticals 2010, 3, 961–993. [Google Scholar] [CrossRef]
- Derakhshankhah, H.; Jafari, S. Cell Penetrating Peptides: A Concise Review with Emphasis on Biomedical Applications. Biomed. Pharmacother. 2018, 108, 1090–1096. [Google Scholar] [CrossRef]
- Alluis, B.; Fruchart, J.-S. Cell Penetrating Peptide Conjugates for Delivering of Nucleic Acids into a Cell 2013. Patent Application No 13/659,395, 30 May 2013. [Google Scholar]
- Simeoni, F.; Morris, M.C.; Heitz, F.; Divita, G. Insight into the Mechanism of the Peptide-based Gene Delivery System MPG: Implications for Delivery of SiRNA into Mammalian Cells. Nucleic Acids Res. 2003, 31, 2717–2724. [Google Scholar] [CrossRef]
- Crombez, L.; Morris, M.C.; Dufort, S.; Aldrian-Herrada, G.; Nguyen, Q.; Mc Master, G.; Coll, J.-L.; Heitz, F.; Divita, G. Targeting Cyclin B1 through Peptide-Based Delivery of SiRNA Prevents Tumour Growth. Nucleic Acids Res. 2009, 37, 4559–4569. [Google Scholar] [CrossRef]
- Rhee, M.; Davis, P. Mechanism of Uptake of C105Y, a Novel Cell-Penetrating Peptide. J. Biol. Chem. 2006, 281, 1233–1240. [Google Scholar] [CrossRef]
- Sun, W.; Ziady, A.G. Real-Time Imaging of Gene Delivery and Expression with DNA Nanoparticle Technologies. In Micro and Nano Technologies in Bioanalysis: Methods and Protocols; Foote, R.S., Lee, J.W., Eds.; Methods in Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2009; pp. 525–546. ISBN 978-1-59745-483-4. [Google Scholar]
- Saamoah-Moffatt, S.; Wiehle, S.; Christina, R.J. Enhanced Gene Delivery by a Novel Tumor-Specific Vector Containing Peptide and Nucleic Acid Based Nuclear Translocation Signals. Mol. Ther. 2004, 9, S137–S138. [Google Scholar] [CrossRef]
- Lee, M.; Choi, J.S.; Ko, K.S. 202. DNA Delivery to the Mitochondria Sites Using Leader Peptide Conjugated Polyethylenimine. Mol. Ther. 2005, 11, S79–S80. [Google Scholar] [CrossRef]
- Rajala, A.; Wang, Y.; Zhu, Y.; Ranjo-Bishop, M.; Ma, J.-X.; Mao, C.; Rajala, R.V.S. Nanoparticle-Assisted Targeted Delivery of Eye-Specific Genes to Eyes Significantly Improves the Vision of Blind Mice In Vivo. Nano Lett. 2014, 14, 5257–5263. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhan, S.; Fan, W.; Ding, X.; Wu, X.; Zhang, W.; Fu, Y.; Huang, Y.; Huang, X.; Chen, R.; et al. Peptide-Mediated Tumor Targeting by a Degradable Nano Gene Delivery Vector Based on Pluronic-Modified Polyethylenimine. Nanoscale Res. Lett. 2016, 11, 122. [Google Scholar] [CrossRef]
- Dong, S.; Zhou, X.; Yang, J. TAT Modified and Lipid–PEI Hybrid Nanoparticles for Co-Delivery of Docetaxel and PDNA. Biomed. Pharmacother. 2016, 84, 954–961. [Google Scholar] [CrossRef]
- Ishiguro, S.; Alhakamy, N.A.; Uppalapati, D.; Delzeit, J.; Berkland, C.J.; Tamura, M. Combined Local Pulmonary and Systemic Delivery of AT2R Gene by Modified TAT Peptide Nanoparticles Attenuates Both Murine and Human Lung Carcinoma Xenografts in Mice. J. Pharm. Sci. 2017, 106, 385–394. [Google Scholar] [CrossRef]
- Bahadoran, A.; Ebrahimi, M.; Yeap, S.K.; Safi, N.; Moeini, H.; Hair-Bejo, M.; Hussein, M.Z.; Omar, A.R. Induction of a Robust Immune Response against Avian Influenza Virus Following Transdermal Inoculation with H5-DNA Vaccine Formulated in Modified Dendrimer-Based Delivery System in Mouse Model. Int. J. Nanomed. 2017, 12, 8573–8585. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hu, S.; Chen, X. Non-Viral Delivery Systems for CRISPR/Cas9-Based Genome Editing: Challenges and Opportunities. Biomaterials 2018, 171, 207–218. [Google Scholar] [CrossRef]
- Koizumi, K.; Nakamura, H.; Iijima, M.; Matsuzaki, T.; Somiya, M.; Kumasawa, K.; Kimura, T.; Kuroda, S. In Vivo Uterine Local Gene Delivery System Using TAT-Displaying Bionanocapsules. J. Gene Med. 2019, 21, e3140. [Google Scholar] [CrossRef]
- Yi, A.; Sim, D.; Lee, Y.-J.; Sarangthem, V.; Park, R.-W. Development of Elastin-like Polypeptide for Targeted Specific Gene Delivery in Vivo. J. Nanobiotechnol. 2020, 18, 15. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.Y.; Yang, K.-J.; Park, S.-N.; Kim, D.-K.; Kim, J.-D. The Effect of Dexamethasone/Cell-Penetrating Peptide Nanoparticles on Gene Delivery for Inner Ear Therapy. Int. J. Nanomed. 2016, 11, 6123–6134. [Google Scholar] [CrossRef]
- Khalil, I.A.; Harashima, H. An Efficient PEGylated Gene Delivery System with Improved Targeting: Synergism between Octaarginine and a Fusogenic Peptide. Int. J. Pharm. 2018, 538, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, J.; Kim, Y.-J.; Lee, E.; Choi, J.S. Gene Delivery of PAMAM Dendrimer Conjugated with the Nuclear Localization Signal Peptide Originated from Fibroblast Growth Factor 3. Int. J. Pharm. 2014, 459, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Wang, X.; Lee, R.J.; Teng, L. Fatty Acid Modified Octa-Arginine for Delivery of SiRNA. Int. J. Pharm. 2015, 495, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Van Rossenberg, S.; van Keulen, A.; Drijfhout, J.-W.; Vasto, S.; Koerten, H.K.; Spies, F.; van ’t Noordende, J.; van Berkel, T.; Biessen, E.a.L. Stable Polyplexes Based on Arginine-Containing Oligopeptides for in Vivo Gene Delivery. Gene Ther. 2004, 11, 457–464. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Br, L.; Md, L.; Hj, C.; Hj, L. Arginine-Rich Cell-Penetrating Peptides Deliver Gene into Living Human Cells. Gene 2012, 505, 37–45. [Google Scholar] [CrossRef]
- Liu, C.; Liu, X.; Rocchi, P.; Qu, F.; Iovanna, J.L.; Peng, L. Arginine-Terminated Generation 4 PAMAM Dendrimer as an Effective Nanovector for Functional SiRNA Delivery in Vitro and in Vivo. Bioconjug. Chem. 2014, 25, 521–532. [Google Scholar] [CrossRef]
- Kato, T.; Yamashita, H.; Misawa, T.; Nishida, K.; Kurihara, M.; Tanaka, M.; Demizu, Y.; Oba, M. Plasmid DNA Delivery by Arginine-Rich Cell-Penetrating Peptides Containing Unnatural Amino Acids. Bioorg. Med. Chem. 2016, 24, 2681–2687. [Google Scholar] [CrossRef]
- Arukuusk, P.; Pärnaste, L.; Hällbrink, M.; Langel, Ü. PepFects and NickFects for the Intracellular Delivery of Nucleic Acids. In Cell-Penetrating Peptides: Methods and Protocols; Langel, Ü., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; pp. 303–315. ISBN 978-1-4939-2806-4. [Google Scholar]
- Margus, H.; Arukuusk, P.; Langel, Ü.; Pooga, M. Characteristics of Cell-Penetrating Peptide/Nucleic Acid Nanoparticles. Mol. Pharm. 2016, 13, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Freimann, K.; Arukuusk, P.; Kurrikoff, K.; Pärnaste, L.; Raid, R.; Piirsoo, A.; Pooga, M.; Langel, Ü. Formulation of Stable and Homogeneous Cell-Penetrating Peptide NF55 Nanoparticles for Efficient Gene Delivery In Vivo. Mol. Ther. Nucleic Acids 2018, 10, 28–35. [Google Scholar] [CrossRef]
- Padari, K.; Porosk, L.; Arukuusk, P.; Pooga, M. Characterization of Peptide–Oligonucleotide Complexes Using Electron Microscopy, Dynamic Light Scattering, and Protease Resistance Assay. In Oligonucleotide-Based Therapies: Methods and Protocols; Gissberg, O., Zain, R., Lundin, K.E., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; pp. 127–139. ISBN 978-1-4939-9670-4. [Google Scholar]
- Minchin, R.F.; Yang, S. Endosomal Disruptors in Non-Viral Gene Delivery. Expert Opin. Drug Deliv. 2010, 7, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.-Y.; Lai, L.-H.; Shen, J.; Wang, Q.-Q.; Xu, F.-J.; Tang, G.-P. Gene Delivery to Tumor Cells by Cationic Polymeric Nanovectors Coupled to Folic Acid and the Cell-Penetrating Peptide Octaarginine. Biomaterials 2011, 32, 7253–7262. [Google Scholar] [CrossRef] [PubMed]
- Oba, M.; Demizu, Y.; Yamashita, H.; Kurihara, M.; Tanaka, M. Plasmid DNA Delivery Using Fluorescein-Labeled Arginine-Rich Peptides. Bioorg. Med. Chem. 2015, 23, 4911–4918. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Z.; Sun, F.; Xu, Y.; Du, Z. Effect of Inserted Spacer in Hepatic Cell-Penetrating Multifunctional Peptide Component on the DNA Intracellular Delivery of Quaternary Complexes Based on Modular Design. Int. J. Nanomed. 2016, 11, 6283–6295. [Google Scholar] [CrossRef] [PubMed]
- Van den Brand, D.; Gorris, M.A.J.; van Asbeck, A.H.; Palmen, E.; Ebisch, I.; Dolstra, H.; Hällbrink, M.; Massuger, L.F.A.G.; Brock, R. Peptide-Mediated Delivery of Therapeutic MRNA in Ovarian Cancer. Eur. J. Pharm. Biopharm. 2019, 141, 180–190. [Google Scholar] [CrossRef]
- Kurrikoff, K.; Veiman, K.-L.; Künnapuu, K.; Peets, E.M.; Lehto, T.; Pärnaste, L.; Arukuusk, P.; Langel, Ü. Effective in Vivo Gene Delivery with Reduced Toxicity, Achieved by Charge and Fatty Acid -Modified Cell Penetrating Peptide. Sci. Rep. 2017, 7, 17056. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.K.; Pan, H.; Lanza, G.M.; Wickline, S.A. Melittin Derived Peptides for Nanoparticle Based SiRNA Transfection. Biomaterials 2013, 34, 3110–3119. [Google Scholar] [CrossRef]
- Christiaens, B.; Dubruel, P.; Grooten, J.; Goethals, M.; Vandekerckhove, J.; Schacht, E.; Rosseneu, M. Enhancement of Polymethacrylate-Mediated Gene Delivery by Penetratin. Eur. J. Pharm. Sci. 2005, 24, 525–537. [Google Scholar] [CrossRef]
- Zorko, M.; Langel, Ü. Cell-Penetrating Peptides: Mechanism and Kinetics of Cargo Delivery. Adv. Drug Deliv. Rev. 2005, 57, 529–545. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, K.; Tai, L.; Liu, Y.; Wei, G.; Lu, W.; Pan, W. Facile Noninvasive Retinal Gene Delivery Enabled by Penetratin. ACS Appl. Mater. Interfaces 2016, 8, 19256–19267. [Google Scholar] [CrossRef]
- Mo, R.H.; Zaro, J.L.; Shen, W.-C. Comparison of Cationic and Amphipathic Cell Penetrating Peptides for SiRNA Delivery and Efficacy. Mol. Pharm. 2012, 9, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.R.; Huang, Y.-W.; Aronstam, R.S.; Lee, H.-J. Identification of a Short Cell-Penetrating Peptide from Bovine Lactoferricin for Intracellular Delivery of DNA in Human A549 Cells. PLoS ONE 2016, 11, e0150439. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.R.; Huang, Y.-W.; Korivi, M.; Lo, S.-Y.; Aronstam, R.S.; Lee, H.-J. The Primary Mechanism of Cellular Internalization for a Short Cell- Penetrating Peptide as a Nano-Scale Delivery System. Curr. Pharm. Biotechnol. 2017, 18, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Freitas, V.J.F.; Campelo, I.S.; Silva, M.M.A.S.; Cavalcanti, C.M.; Teixeira, D.I.A.; Camargo, L.S.A.; Melo, L.M.; Rádis-Baptista, G. Disulphide-Less Crotamine Is Effective for Formation of DNA–Peptide Complex but Is Unable to Improve Bovine Embryo Transfection. Zygote 2020, 28, 72–79. [Google Scholar] [CrossRef]
- Campeiro, J.D.; Dam, W.; Monte, G.G.; Porta, L.C.; de Oliveira, L.C.G.; Nering, M.B.; Viana, G.M.; Carapeto, F.C.; Oliveira, E.B.; van den Born, J.; et al. Long Term Safety of Targeted Internalization of Cell Penetrating Peptide Crotamine into Renal Proximal Tubular Epithelial Cells in Vivo. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Nascimento, F.D.; Hayashi, M.A.F.; Kerkis, A.; Oliveira, V.; Oliveira, E.B.; Rádis-Baptista, G.; Nader, H.B.; Yamane, T.; dos Santos Tersariol, I.L.; Kerkis, I. Crotamine Mediates Gene Delivery into Cells through the Binding to Heparan Sulfate Proteoglycans. J. Biol. Chem. 2007, 282, 21349–21360. [Google Scholar] [CrossRef]
- Zavaglia, D.; Favrot, M.-C.; Eymin, B.; Tenaud, C.; Coll, J.-L. Intercellular Trafficking and Enhanced in Vivo Antitumour Activity of a Non-Virally Delivered P27-VP22 Fusion Protein. Gene Ther. 2003, 10, 314–325. [Google Scholar] [CrossRef][Green Version]
- Chen, H.-C.G.; Chiou, S.-T.; Zheng, J.-Y.; Yang, S.-H.; Lai, S.-S.; Kuo, T.-Y. The Nuclear Localization Signal Sequence of Porcine Circovirus Type 2 ORF2 Enhances Intracellular Delivery of Plasmid DNA. Arch. Virol. 2011, 156, 803–815. [Google Scholar] [CrossRef]
- Kim, N.H.; Provoda, C.; Lee, K.-D. Design and Characterization of Novel Recombinant Listeriolysin O–Protamine Fusion Proteins for Enhanced Gene Delivery. Mol. Pharm. 2015, 12, 342–350. [Google Scholar] [CrossRef]
- Rezaee, M.; Oskuee, R.K.; Nassirli, H.; Malaekeh-Nikouei, B. Progress in the Development of Lipopolyplexes as Efficient Non-Viral Gene Delivery Systems. J. Control. Release 2016, 236, 1–14. [Google Scholar] [CrossRef]
- Men, K.; Zhang, R.; Zhang, X.; Huang, R.; Zhu, G.; Tong, R.; Yang, L.; Wei, Y.; Duan, X. Delivery of Modified MRNA Encoding Vesicular Stomatitis Virus Matrix Protein for Colon Cancer Gene Therapy. RSC Adv. 2018, 8, 12104–12115. [Google Scholar] [CrossRef]
- Limeres, M.J.; Suñé-Pou, M.; Prieto-Sánchez, S.; Moreno-Castro, C.; Nusblat, A.D.; Hernández-Munain, C.; Castro, G.R.; Suñé, C.; Suñé-Negre, J.M.; Cuestas, M.L. Development and Characterization of an Improved Formulation of Cholesteryl Oleate-Loaded Cationic Solid-Lipid Nanoparticles as an Efficient Non-Viral Gene Delivery System. Colloids Surf. B Biointerfaces 2019, 184, 110533. [Google Scholar] [CrossRef]
- Zhang, F.; Li, H.-Y. Preparation of Lipid–Peptide–DNA (LPD) Nanoparticles and Their Use for Gene Transfection. In Nanoparticles in Biology and Medicine: Methods and Protocols; Ferrari, E., Soloviev, M., Eds.; Methods in Molecular Biology; Springer US: New York, NY, USA, 2020; pp. 91–98. ISBN 978-1-07-160319-2. [Google Scholar]
- Astriab-Fisher, A.; Sergueev, D.; Fisher, M.; Ramsay Shaw, B.; Juliano, R.L. Conjugates of Antisense Oligonucleotides with the Tat and Antennapedia Cell-Penetrating Peptides: Effects on Cellular Uptake, Binding to Target Sequences, and Biologic Actions. Pharm. Res. 2002, 19, 744–754. [Google Scholar] [CrossRef]
- Davidson, T.J.; Harel, S.; Arboleda, V.A.; Prunell, G.F.; Shelanski, M.L.; Greene, L.A.; Troy, C.M. Highly Efficient Small Interfering RNA Delivery to Primary Mammalian Neurons Induces MicroRNA-Like Effects before MRNA Degradation. J. Neurosci. 2004, 24, 10040–10046. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Morris, M.A.; Heitz, F.; Divita, G.; Morris, M.C. The Peptide Carrier Pep-1 Forms Biologically Efficient Nanoparticle Complexes. Biochem. Biophys. Res. Commun. 2007, 355, 877–882. [Google Scholar] [CrossRef]
- Chang, J.-C.; Liu, K.-H.; Chuang, C.-S.; Su, H.-L.; Wei, Y.-H.; Kuo, S.-J.; Liu, C.-S. Treatment of Human Cells Derived from MERRF Syndrome by Peptide-Mediated Mitochondrial Delivery. Cytotherapy 2013, 15, 1580–1596. [Google Scholar] [CrossRef] [PubMed]
- Crowet, J.-M.; Lins, L.; Deshayes, S.; Divita, G.; Morris, M.; Brasseur, R.; Thomas, A. Modeling of Non-Covalent Complexes of the Cell-Penetrating Peptide CADY and Its SiRNA Cargo. Biochim. Biophys. Acta BBA Biomembr. 2013, 1828, 499–509. [Google Scholar] [CrossRef][Green Version]
- Rydström, A.; Deshayes, S.; Konate, K.; Crombez, L.; Padari, K.; Boukhaddaoui, H.; Aldrian, G.; Pooga, M.; Divita, G. Direct Translocation as Major Cellular Uptake for CADY Self-Assembling Peptide-Based Nanoparticles. PLoS ONE 2011, 6, e25924. [Google Scholar] [CrossRef]
- Konate, K.; Lindberg, M.F.; Vaissiere, A.; Jourdan, C.; Aldrian, G.; Margeat, E.; Deshayes, S.; Boisguerin, P. Optimisation of Vectorisation Property: A Comparative Study for a Secondary Amphipathic Peptide. Int. J. Pharm. 2016, 509, 71–84. [Google Scholar] [CrossRef]
- Elmquist, A.; Lindgren, M.; Bartfai, T.; Langel, Ü. VE-Cadherin-Derived Cell-Penetrating Peptide, PVEC, with Carrier Functions. Exp. Cell Res. 2001, 269, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Rajpal; Mann, A.; Khanduri, R.; Naik, R.J.; Ganguli, M. Structural Rearrangements and Chemical Modifications in Known Cell Penetrating Peptide Strongly Enhance DNA Delivery Efficiency. J. Control. Release 2012, 157, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.G.; Sayers, E.J.; He, L.; Narayan, R.; Williams, T.L.; Mills, E.M.; Allemann, R.K.; Luk, L.Y.P.; Jones, A.T.; Tsai, Y.-H. Cell-Penetrating Peptide Sequence and Modification Dependent Uptake and Subcellular Distribution of Green Florescent Protein in Different Cell Lines. Sci. Rep. 2019, 9, 6298. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, J.; Xu, D. Cell-Penetrating Peptides as Noninvasive Transmembrane Vectors for the Development of Novel Multifunctional Drug-Delivery Systems. J. Control. Release 2016, 229, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Gessner, I.; Neundorf, I. Nanoparticles Modified with Cell-Penetrating Peptides: Conjugation Mechanisms, Physicochemical Properties, and Application in Cancer Diagnosis and Therapy. Int. J. Mol. Sci. 2020, 21, 2536. [Google Scholar] [CrossRef]
- Fischer, R.; Fotin-Mleczek, M.; Hufnagel, H.; Brock, R. Break on through to the Other Side—Biophysics and Cell Biology Shed Light on Cell-Penetrating Peptides. ChemBioChem 2005, 6, 2126–2142. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, S.; Liang, Z.; Zhao, M.; Liu, G.; Wu, J. Progress in Arginine-Based Gene Delivery Systems. J. Mater. Chem. B 2020. [Google Scholar] [CrossRef]
- Nam, H.Y.; Nam, K.; Hahn, H.J.; Kim, B.H.; Lim, H.J.; Kim, H.J.; Choi, J.S.; Park, J.-S. Biodegradable PAMAM Ester for Enhanced Transfection Efficiency with Low Cytotoxicity. Biomaterials 2009, 30, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Tünnemann, G.; Ter-Avetisyan, G.; Martin, R.M.; Stöckl, M.; Herrmann, A.; Cardoso, M.C. Live-Cell Analysis of Cell Penetration Ability and Toxicity of Oligo-Arginines. J. Pept. Sci. 2008, 14, 469–476. [Google Scholar] [CrossRef]
- Singh, T.; Murthy, A.S.N.; Yang, H.-J.; Im, J. Versatility of Cell-Penetrating Peptides for Intracellular Delivery of SiRNA. Drug Deliv. 2018, 25, 2005–2015. [Google Scholar] [CrossRef]
- Ruseska, I.; Zimmer, A. Internalization Mechanisms of Cell-Penetrating Peptides. Beilstein J. Nanotechnol. 2020, 11, 101–123. [Google Scholar] [CrossRef]
- Guidotti, G.; Brambilla, L.; Rossi, D. Cell-Penetrating Peptides: From Basic Research to Clinics. Trends Pharmacol. Sci. 2017, 38, 406–424. [Google Scholar] [CrossRef]
- Munyendo, W.L.; Lv, H.; Benza-Ingoula, H.; Baraza, L.D.; Zhou, J. Cell Penetrating Peptides in the Delivery of Biopharmaceuticals. Biomolecules 2012, 2, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Veiman, K.-L.; Mäger, I.; Ezzat, K.; Margus, H.; Lehto, T.; Langel, K.; Kurrikoff, K.; Arukuusk, P.; Suhorutšenko, J.; Padari, K.; et al. PepFect14 Peptide Vector for Efficient Gene Delivery in Cell Cultures. Mol. Pharm. 2013, 10, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Pärnaste, L.; Arukuusk, P.; Langel, K.; Tenson, T.; Langel, Ü. The Formation of Nanoparticles between Small Interfering RNA and Amphipathic Cell-Penetrating Peptides. Mol. Ther. Nucleic Acids 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Carreras-Badosa, G.; Maslovskaja, J.; Periyasamy, K.; Urgard, E.; Padari, K.; Vaher, H.; Tserel, L.; Gestin, M.; Kisand, K.; Arukuusk, P.; et al. NickFect Type of Cell-Penetrating Peptides Present Enhanced Efficiency for MicroRNA-146a Delivery into Dendritic Cells and during Skin Inflammation. Biomaterials 2020, 262, 120316. [Google Scholar] [CrossRef] [PubMed]
- Varkouhi, A.K.; Scholte, M.; Storm, G.; Haisma, H.J. Endosomal Escape Pathways for Delivery of Biologicals. J. Control. Release 2011, 151, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, J.; García, I.; Liz-Marzán, L.M. Cellular Uptake of Nanoparticles versus Small Molecules: A Matter of Size. Acc. Chem. Res. 2018, 51, 2305–2313. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular Uptake of Nanoparticles: Journey Inside the Cell. Chem. Soc. Rev. 2017, 46, 4218–4244. [Google Scholar] [CrossRef]
- Lönn, P.; Kacsinta, A.D.; Cui, X.-S.; Hamil, A.S.; Kaulich, M.; Gogoi, K.; Dowdy, S.F. Enhancing Endosomal Escape for Intracellular Delivery of Macromolecular Biologic Therapeutics. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Ahmad, A.; Khan, J.M.; Haque, S. Strategies in the Design of Endosomolytic Agents for Facilitating Endosomal Escape in Nanoparticles. Biochimie 2019, 160, 61–75. [Google Scholar] [CrossRef]
- Erazo-Oliveras, A.; Muthukrishnan, N.; Baker, R.; Wang, T.-Y.; Pellois, J.-P. Improving the Endosomal Escape of Cell-Penetrating Peptides and Their Cargos: Strategies and Challenges. Pharmaceuticals 2012, 5, 1177–1209. [Google Scholar] [CrossRef]
- Daussy, C.F.; Wodrich, H. “Repair Me If You Can”: Membrane Damage, Response, and Control from the Viral Perspective. Cells 2020, 9, 2042. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Nigatu, A.S.; Berkland, C.J.; Ramsey, J.D. Noncovalently Associated -Penetrating Peptides for Gene Delivery Applications. Ther. Deliv. 2013, 4, 741–757. [Google Scholar] [CrossRef]
- Loughran, S.P.; McCrudden, C.M.; McCarthy, H.O. Designer Peptide Delivery Systems for Gene Therapy. Eur. J. Nanomed. 2015, 7, 85–96. [Google Scholar] [CrossRef]
- Sudo, K.; Niikura, K.; Iwaki, K.; Kohyama, S.; Fujiwara, K.; Doi, N. Human-Derived Fusogenic Peptides for the Intracellular Delivery of Proteins. J. Control. Release 2017, 255, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Miralles, N.; Vázquez, E.; Villaverde, A. Membrane-Active Peptides for Non-Viral Gene Therapy: Making the Safest Easier. Trends Biotechnol. 2008, 26, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Wadia, J.S.; Stan, R.V.; Dowdy, S.F. Transducible TAT-HA Fusogenic Peptide Enhances Escape of TAT-Fusion Proteins after Lipid Raft Macropinocytosis. Nat. Med. 2004, 10, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Tian, M.; Wang, T.; Ren, L.; Wang, D.; Shen, L.; Shang, T. Synergistic Effects of Cell-Penetrating Peptide Tat and Fusogenic Peptide HA2-Enhanced Cellular Internalization and Gene Transduction of Organosilica Nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Karjoo, Z.; McCarthy, H.O.; Patel, P.; Nouri, F.S.; Hatefi, A. Systematic Engineering of Uniform, Highly Efficient, Targeted and Shielded Viral-Mimetic Nanoparticles. Small 2013, 9, 2774–2783. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.; Tao, S.; Ye, T.; Kong, X.; Ren, L. In Vivo Bio-Distribution and Efficient Tumor Targeting of Gelatin/Silica Nanoparticles for Gene Delivery. Nanoscale Res. Lett. 2016, 11, 195. [Google Scholar] [CrossRef]
- Golan, M.; Feinshtein, V.; David, A. Conjugates of HA2 with Octaarginine-Grafted HPMA Copolymer Offer Effective SiRNA Delivery and Gene Silencing in Cancer Cells. Eur. J. Pharm. Biopharm. 2016, 109, 103–112. [Google Scholar] [CrossRef]
- Cantini, L.; Attaway, C.C.; Butler, B.; Andino, L.M.; Sokolosky, M.L.; Jakymiw, A. Fusogenic-Oligoarginine Peptide-Mediated Delivery of SiRNAs Targeting the CIP2A Oncogene into Oral Cancer Cells. PLoS ONE 2013, 8, e73348. [Google Scholar] [CrossRef]
- Funhoff, A.M.; van Nostrum, C.F.; Koning, G.A.; Schuurmans-Nieuwenbroek, N.M.E.; Crommelin, D.J.A.; Hennink, W.E. Endosomal Escape of Polymeric Gene Delivery Complexes Is Not Always Enhanced by Polymers Buffering at Low PH. Biomacromolecules 2004, 5, 32–39. [Google Scholar] [CrossRef]
- Wang, Y.; Mangipudi, S.S.; Canine, B.F.; Hatefi, A. A Designer Biomimetic Vector with a Chimeric Architecture for Targeted Gene Transfer. J. Control. Release Off. J. Control. Release Soc. 2009, 137, 46–53. [Google Scholar] [CrossRef]
- Hou, K.K.; Pan, H.; Schlesinger, P.H.; Wickline, S.A. A Role for Peptides in Overcoming Endosomal Entrapment in SiRNA Delivery—A Focus on Melittin. Biotechnol. Adv. 2015, 33, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Raghuraman, H.; Chattopadhyay, A. Melittin: A Membrane-Active Peptide with Diverse Functions. Biosci. Rep. 2007, 27, 189–223. [Google Scholar] [CrossRef]
- Paray, B.A.; Ahmad, A.; Khan, J.M.; Taufiq, F.; Pathan, A.; Malik, A.; Ahmed, M.Z. The Role of the Multifunctional Antimicrobial Peptide Melittin in Gene Delivery. Drug Discov. Today 2021, 26, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Schellinger, J.G.; Pahang, J.A.; Johnson, R.N.; Chu, D.S.H.; Sellers, D.L.; Maris, D.O.; Convertine, A.J.; Stayton, P.S.; Horner, P.J.; Pun, S.H. Melittin-Grafted HPMA-Oligolysine Based Copolymers for Gene Delivery. Biomaterials 2013, 34, 2318–2326. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, Z.; Wang, C.; Yang, T.; Cai, C.; Zhao, X.; Yang, L.; Ding, P. Exploring the Role of Peptides in Polymer-Based Gene Delivery. Acta Biomater. 2017, 60, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Nouri, F.S.; Wang, X.; Dorrani, M.; Karjoo, Z.; Hatefi, A. A Recombinant Biopolymeric Platform for Reliable Evaluation of the Activity of PH-Responsive Amphiphile Fusogenic Peptides. Biomacromolecules 2013, 14, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jeong, J.H.; Park, T.G. PEG Grafted Polylysine with Fusogenic Peptide for Gene Delivery: High Transfection Efficiency with Low Cytotoxicity. J. Control. Release 2002, 79, 283–291. [Google Scholar] [CrossRef]
- Bennett, R.; Yakkundi, A.; McKeen, H.D.; McClements, L.; McKeogh, T.J.; McCrudden, C.M.; Arthur, K.; Robson, T.; McCarthy, H.O. RALA-Mediated Delivery of FKBPL Nucleic Acid Therapeutics. Nanomedicine 2015, 10, 2989–3001. [Google Scholar] [CrossRef]
- Mulholland, E.J.; Ali, A.; Robson, T.; Dunne, N.J.; McCarthy, H.O. Delivery of RALA/SiFKBPL Nanoparticles via Electrospun Bilayer Nanofibres: An Innovative Angiogenic Therapy for Wound Repair. J. Control. Release 2019, 316, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.-P.; Castaño, I.M.; Sridharan, R.; Kelly, D.; Lemoine, M.; Cavanagh, B.L.; Dunne, N.J.; McCarthy, H.O.; O’Brien, F.J. Collagen/GAG Scaffolds Activated by RALA-SiMMP-9 Complexes with Potential for Improved Diabetic Foot Ulcer Healing. Mater. Sci. Eng. C 2020, 114, 111022. [Google Scholar] [CrossRef]
- Akita, H.; Masuda, T.; Nishio, T.; Niikura, K.; Ijiro, K.; Harashima, H. Improving in Vivo Hepatic Transfection Activity by Controlling Intracellular Trafficking: The Function of GALA and Maltotriose. Mol. Pharm. 2011, 8, 1436–1442. [Google Scholar] [CrossRef]
- Li, X.; Chen, Y.; Wang, M.; Ma, Y.; Xia, W.; Gu, H. A Mesoporous Silica Nanoparticle–PEI–Fusogenic Peptide System for SiRNA Delivery in Cancer Therapy. Biomaterials 2013, 34, 1391–1401. [Google Scholar] [CrossRef]
- Ali, A.A.; McCrudden, C.M.; McCaffrey, J.; McBride, J.W.; Cole, G.; Dunne, N.J.; Robson, T.; Kissenpfennig, A.; Donnelly, R.F.; McCarthy, H.O. DNA Vaccination for Cervical Cancer; a Novel Technology Platform of RALA Mediated Gene Delivery via Polymeric Microneedles. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 921–932. [Google Scholar] [CrossRef]
- McCrudden, C.M.; McBride, J.W.; McCaffrey, J.; McErlean, E.M.; Dunne, N.J.; Kett, V.L.; Coulter, J.A.; Robson, T.; McCarthy, H.O. Gene Therapy with RALA/INOS Composite Nanoparticles Significantly Enhances Survival in a Model of Metastatic Prostate Cancer. Cancer Nanotechnol. 2018, 9, 5. [Google Scholar] [CrossRef]
- Sousa, Â.; Almeida, A.M.; Faria, R.; Konate, K.; Boisguerin, P.; Queiroz, J.A.; Costa, D. Optimization of Peptide-Plasmid DNA Vectors Formulation for Gene Delivery in Cancer Therapy Exploring Design of Experiments. Colloids Surf. B Biointerfaces 2019, 183, 110417. [Google Scholar] [CrossRef]
- Gottschalk, S.; Sparrow, J.T.; Hauer, J.; Mims, M.P.; Leland, F.E.; Woo, S.L.-Y.; Smith, L.C. A Novel DNA-Peptide Complex for Efficient Gene Transfer and Expression in Mammalian Cells. Gene Ther. 1996, 3, 448–457. [Google Scholar]
- Guryanov, I.A.; Vlasov, G.P.; Lesina, E.A.; Kiselev, A.V.; Baranov, V.S.; Avdeeva, E.V.; Vorob’ev, V.I. Cationic Oligopeptides Modified with Lipophilic Fragments: Use for DNA Delivery to Cells. Russ. J. Bioorg. Chem. 2005, 31, 18–26. [Google Scholar] [CrossRef]
- Van Rossenberg, S.M.W.; Sliedregt-Bol, K.M.; Meeuwenoord, N.J.; van Berkel, T.J.C.; van Boom, J.H.; van der Marel, G.A.; Biessen, E.A.L. Targeted Lysosome Disruptive Elements for Improvement of Parenchymal Liver Cell-Specific Gene Delivery. J. Biol. Chem. 2002, 277, 45803–45810. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Luan, L.; Kang, Z.; Feng, S.; Meng, Q.; Liu, K. Histidine-Enriched Multifunctional Peptide Vectors with Enhanced Cellular Uptake and Endosomal Escape for Gene Delivery. J. Mater. Chem. B 2017, 5, 74–84. [Google Scholar] [CrossRef]
- Kichler, A.; Mason, A.J.; Bechinger, B. Cationic Amphipathic Histidine-Rich Peptides for Gene Delivery. Biochim. Biophys. Acta BBA Biomembr. 2006, 1758, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Midoux, P.; Pichon, C.; Yaouanc, J.-J.; Jaffrès, P.-A. Chemical Vectors for Gene Delivery: A Current Review on Polymers, Peptides and Lipids Containing Histidine or Imidazole as Nucleic Acids Carriers. Br. J. Pharmacol. 2009, 157, 166–178. [Google Scholar] [CrossRef]
- Chen, Q.-R.; Zhang, L.; Stass, S.A.; Mixson, A.J. Co-Polymer of Histidine and Lysine Markedly Enhances Transfection Efficiency of Liposomes. Gene Ther. 2000, 7, 1698–1705. [Google Scholar] [CrossRef]
- Pichon, C.; Gonçalves, C.; Midoux, P. Histidine-Rich Peptides and Polymers for Nucleic Acids Delivery. Adv. Drug Deliv. Rev. 2001, 53, 75–94. [Google Scholar] [CrossRef]
- Perche, F.; Gosset, D.; Mével, M.; Miramon, M.-L.; Yaouanc, J.-J.; Pichon, C.; Benvegnu, T.; Jaffrès, P.-A.; Midoux, P. Selective Gene Delivery in Dendritic Cells with Mannosylated and Histidylated Lipopolyplexes. J. Drug Target. 2011, 19, 315–325. [Google Scholar] [CrossRef]
- Perche, F.; Benvegnu, T.; Berchel, M.; Lebegue, L.; Pichon, C.; Jaffrès, P.-A.; Midoux, P. Enhancement of Dendritic Cells Transfection in Vivo and of Vaccination against B16F10 Melanoma with Mannosylated Histidylated Lipopolyplexes Loaded with Tumor Antigen Messenger RNA. Nanomed. Nanotechnol. Biol. Med. 2011, 7, 445–453. [Google Scholar] [CrossRef]
- Perche, F.; Lambert, O.; Berchel, M.; Jaffrès, P.-A.; Pichon, C.; Midoux, P. Gene Transfer by Histidylated Lipopolyplexes: A Dehydration Method Allowing Preservation of Their Physicochemical Parameters and Transfection Efficiency. Int. J. Pharm. 2012, 423, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Dong, C.; Dong, H.; Ren, T.; Wen, X.; Su, J.; Li, Y. Cleavable PEGylation and Hydrophobic Histidylation of Polylysine for SiRNA Delivery and Tumor Gene Therapy. ACS Appl. Mater. Interfaces 2014, 6, 10393–10407. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Qiao, M.; Wang, Q.; Ye, Y.; Ba, S.; Ma, J.; Hu, H.; Zhao, X.; Chen, D. Dual-Responsive Polyplexes with Enhanced Disassembly and Endosomal Escape for Efficient Delivery of SiRNA. Biomaterials 2018, 162, 47–59. [Google Scholar] [CrossRef]
- Sadeghian, F.; Hosseinkhani, S.; Alizadeh, A.; Hatefi, A. Design, Engineering and Preparation of a Multi-Domain Fusion Vector for Gene Delivery. Int. J. Pharm. 2012, 427, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Asseline, U.; Gonçalves, C.; Pichon, C.; Midoux, P. Improved Nuclear Delivery of Antisense 2’-Ome RNA by Conjugation with the Histidine-Rich Peptide H5WYG. J. Gene Med. 2014, 16, 157–165. [Google Scholar] [CrossRef]
- McErlean, E.M.; McCrudden, C.M.; McCarthy, H.O. Delivery of Nucleic Acids for Cancer Gene Therapy: Overcoming Extra- and Intra-Cellular Barriers. Ther. Deliv. 2016, 7, 619–637. [Google Scholar] [CrossRef]
- McBride, J.W.; Massey, A.S.; McCaffrey, J.; McCrudden, C.M.; Coulter, J.A.; Dunne, N.J.; Robson, T.; McCarthy, H.O. Development of TMTP-1 Targeted Designer Biopolymers for Gene Delivery to Prostate Cancer. Int. J. Pharm. 2016, 500, 144–153. [Google Scholar] [CrossRef]
- Alipour, M.; Hosseinkhani, S.; Sheikhnejad, R.; Cheraghi, R. Nano-Biomimetic Carriers Are Implicated in Mechanistic Evaluation of Intracellular Gene Delivery. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Alipour, M.; Majidi, A.; Molaabasi, F.; Sheikhnejad, R.; Hosseinkhani, S. In Vivo Tumor Gene Delivery Using Novel Peptideticles: PH-Responsive and Ligand Targeted Core–Shell Nanoassembly. Int. J. Cancer 2018, 143, 2017–2028. [Google Scholar] [CrossRef]
- Thapa, R.K.; Sullivan, M.O. Gene Delivery by Peptide-Assisted Transport. Curr. Opin. Biomed. Eng. 2018, 7, 71–82. [Google Scholar] [CrossRef]
- Liu, J.; Guo, N.; Gao, C.; Liu, N.; Zheng, X.; Tan, Y.; Lei, J.; Hao, Y.; Chen, L.; Zhang, X. Effective Gene Silencing Mediated by Polypeptide Nanoparticles LAH4-L1-SiMDR1 in Multi-Drug Resistant Human Breast Cancer. J. Biomed. Nanotechnol. 2019, 15, 531–543. [Google Scholar] [CrossRef]
- Liang, W.; Lam, J.K.W. Endosomal Escape Pathways for Non-Viral Nucleic Acid Delivery Systems. Intech Open: Rijeka, Croatia, 2012; ISBN 978-953-51-0662-3. [Google Scholar]
- Bechinger, B. Towards Membrane Protein Design: PH-Sensitive Topology of Histidine-Containing Polypeptides. J. Mol. Biol. 1996, 263, 768–775. [Google Scholar] [CrossRef]
- Perrone, B.; Miles, A.J.; Salnikov, E.S.; Wallace, B.A.; Bechinger, B. Lipid Interactions of LAH4, a Peptide with Antimicrobial and Nucleic Acid Transfection Activities. Eur. Biophys. J. 2014, 43, 499–507. [Google Scholar] [CrossRef]
- Marquette, A.; Mason, A.J.; Bechinger, B. Aggregation and Membrane Permeabilizing Properties of Designed Histidine-Containing Cationic Linear Peptide Antibiotics. J. Pept. Sci. Off. Publ. Eur. Pept. Soc. 2008, 14, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.; Aisenbrey, C.; Harmouche, N.; Raya, J.; Bertani, P.; Voievoda, N.; Süss, R.; Bechinger, B. PH-Dependent Membrane Interactions of the Histidine-Rich Cell-Penetrating Peptide LAH4-L1. Biophys. J. 2017, 113, 1290–1300. [Google Scholar] [CrossRef]
- Langlet-Bertin, B.; Leborgne, C.; Scherman, D.; Bechinger, B.; Mason, A.J.; Kichler, A. Design and Evaluation of Histidine-Rich Amphipathic Peptides for SiRNA Delivery. Pharm. Res. 2010, 27, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liang, W.; Qiu, Y.; Cespi, M.; Palmieri, G.F.; Mason, A.J.; Lam, J.K.W. Incorporation of a Nuclear Localization Signal in PH Responsive LAH4-L1 Peptide Enhances Transfection and Nuclear Uptake of Plasmid DNA. Mol. Pharm. 2016, 13, 3141–3152. [Google Scholar] [CrossRef] [PubMed]
- Moulay, G.; Leborgne, C.; Mason, A.J.; Aisenbrey, C.; Kichler, A.; Bechinger, B. Histidine-Rich Designer Peptides of the LAH4 Family Promote Cell Delivery of a Multitude of Cargo. J. Pept. Sci. 2017, 23, 320–328. [Google Scholar] [CrossRef]
- Liu, N.; Bechinger, B.; Süss, R. The Histidine-Rich Peptide LAH4-L1 Strongly Promotes PAMAM-Mediated Transfection at Low Nitrogen to Phosphorus Ratios in the Presence of Serum. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Kichler, A.; Mason, A.J.; Marquette, A.; Bechinger, B. Histidine-Rich Cationic Cell-Penetrating Peptides for Plasmid DNA and siRNA Delivery. In Nanotechnology for Nucleic Acid Delivery: Methods and Protocols; Ogris, M., Sami, H., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; pp. 39–59. ISBN 978-1-4939-9092-4. [Google Scholar]
- Marquette, A.; Leborgne, C.; Schartner, V.; Salnikov, E.; Bechinger, B.; Kichler, A. Peptides Derived from the C-Terminal Domain of HIV-1 Viral Protein R in Lipid Bilayers: Structure, Membrane Positioning and Gene Delivery. Biochim. Biophys. Acta BBA Biomembr. 2020, 1862, 183149. [Google Scholar] [CrossRef]
- Chamarthy, S.P.; Kovacs, J.R.; McClelland, E.; Gattens, D.; Meng, W.S. A Cationic Peptide Consists of Ornithine and Histidine Repeats Augments Gene Transfer in Dendritic Cells. Mol. Immunol. 2003, 40, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, J.R.; Zheng, Y.; Shen, H.; Meng, W.S. Polymeric Microspheres as Stabilizing Anchors for Oligonucleotide Delivery to Dendritic Cells. Biomaterials 2005, 26, 6754–6761. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Kovacs, J.R.; Zheng, Y.; Gawalt, E.S.; Shen, H.; Meng, W.S. Attenuated Alloreactivity of Dendritic Cells Engineered with Surface-Modified Microspheres Carrying a Plasmid Encoding Interleukin-10. Biomaterials 2006, 27, 2076–2082. [Google Scholar] [CrossRef]
- Lo, S.L.; Wang, S. An Endosomolytic Tat Peptide Produced by Incorporation of Histidine and Cysteine Residues as a Nonviral Vector for DNA Transfection. Biomaterials 2008, 29, 2408–2414. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Li, Q.; Ali, H.; Zaidi, S.S.A.; Guo, J.; Ren, X.; Shi, C.; Xia, S.; Zhang, W.; Feng, Y. POSS-Cored and Peptide Functionalized Ternary Gene Delivery Systems with Enhanced Endosomal Escape Ability for Efficient Intracellular Delivery of Plasmid DNA. J. Mater. Chem. B 2018, 6, 4251–4263. [Google Scholar] [CrossRef]
- Li, Q.; Hao, X.; Zaidi, S.S.A.; Guo, J.; Ren, X.; Shi, C.; Zhang, W.; Feng, Y. Oligohistidine and Targeting Peptide Functionalized TAT-NLS for Enhancing Cellular Uptake and Promoting Angiogenesis in Vivo. J. Nanobiotechnol. 2018, 16, 29. [Google Scholar] [CrossRef] [PubMed]
- Read, M.L.; Singh, S.; Ahmed, Z.; Stevenson, M.; Briggs, S.S.; Oupicky, D.; Barrett, L.B.; Spice, R.; Kendall, M.; Berry, M.; et al. A Versatile Reducible Polycation-Based System for Efficient Delivery of a Broad Range of Nucleic Acids. Nucleic Acids Res. 2005, 33, e86. [Google Scholar] [CrossRef]
- Stevenson, M.; Ramos-Perez, V.; Singh, S.; Soliman, M.; Preece, J.A.; Briggs, S.S.; Read, M.L.; Seymour, L.W. Delivery of SiRNA Mediated by Histidine-Containing Reducible Polycations. J. Control. Release Off. J. Control. Release Soc. 2008, 130, 46–56. [Google Scholar] [CrossRef]
- Durymanov, M.; Reineke, J. Non-Viral Delivery of Nucleic Acids: Insight Into Mechanisms of Overcoming Intracellular Barriers. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Lin, G.; Li, L.; Panwar, N.; Wang, J.; Tjin, S.C.; Wang, X.; Yong, K.-T. Non-Viral Gene Therapy Using Multifunctional Nanoparticles: Status, Challenges, and Opportunities. Coord. Chem. Rev. 2018, 374, 133–152. [Google Scholar] [CrossRef]
- Uludag, H.; Ubeda, A.; Ansari, A. At the Intersection of Biomaterials and Gene Therapy: Progress in Non-Viral Delivery of Nucleic Acids. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef]
- Vermeulen, L.M.P.; Brans, T.; De Smedt, S.C.; Remaut, K.; Braeckmans, K. Methodologies to Investigate Intracellular Barriers for Nucleic Acid Delivery in Non-Viral Gene Therapy. Nano Today 2018, 21, 74–90. [Google Scholar] [CrossRef]
- Shi, B.; Zheng, M.; Tao, W.; Chung, R.; Jin, D.; Ghaffari, D.; Farokhzad, O.C. Challenges in DNA Delivery and Recent Advances in Multifunctional Polymeric DNA Delivery Systems. Biomacromolecules 2017, 18, 2231–2246. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Fan, Y.; Li, Y.; Huang, L. Strategies on the Nuclear-Targeted Delivery of Genes. J. Drug Target. 2013, 21, 926–939. [Google Scholar] [CrossRef]
- Zelmer, C.; Zweifel, L.P.; Kapinos, L.E.; Craciun, I.; Güven, Z.P.; Palivan, C.G.; Lim, R.Y.H. Organelle-Specific Targeting of Polymersomes into the Cell Nucleus. Proc. Natl. Acad. Sci. USA 2020, 117, 2770–2778. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six Classes of Nuclear Localization Signals Specific to Different Binding Grooves of Importin α. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef]
- Pan, L.; He, Q.; Liu, J.; Chen, Y.; Ma, M.; Zhang, L.; Shi, J. Nuclear-Targeted Drug Delivery of TAT Peptide-Conjugated Monodisperse Mesoporous Silica Nanoparticles. J. Am. Chem. Soc. 2012, 134, 5722–5725. [Google Scholar] [CrossRef] [PubMed]
- Van der Aa, M.A.E.M.; Mastrobattista, E.; Oosting, R.S.; Hennink, W.E.; Koning, G.A.; Crommelin, D.J.A. The Nuclear Pore Complex: The Gateway to Successful Nonviral Gene Delivery. Pharm. Res. 2006, 23, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Cartier, R.; Reszka, R. Utilization of Synthetic Peptides Containing Nuclear Localization Signals for Nonviral Gene Transfer Systems. Gene Ther. 2002, 9, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Escriou, V.; Carrière, M.; Scherman, D.; Wils, P. NLS Bioconjugates for Targeting Therapeutic Genes to the Nucleus. Adv. Drug Deliv. Rev. 2003, 55, 295–306. [Google Scholar] [CrossRef]
- Zhao, M.; Li, J.; Ji, H.; Chen, D.; Hu, H. A Versatile Endosome Acidity-Induced Sheddable Gene Delivery System: Increased Tumor Targeting and Enhanced Transfection Efficiency. Int. J. Nanomedicine 2019, 14, 6519–6538. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.; Strong, D.; Zimmer, W. Nuclear Entry of Nonviral Vectors. Gene Ther. 2005, 12, 881–890. [Google Scholar] [CrossRef]
- Morille, M.; Passirani, C.; Vonarbourg, A.; Clavreul, A.; Benoit, J.-P. Progress in Developing Cationic Vectors for Non-Viral Systemic Gene Therapy against Cancer. Biomaterials 2008, 29, 3477–3496. [Google Scholar] [CrossRef] [PubMed]
- Bogacheva, M.; Egorova, A.; Slita, A.; Maretina, M.; Baranov, V.; Kiselev, A. Arginine-Rich Cross-Linking Peptides with Different SV40 Nuclear Localization Signal Content as Vectors for Intranuclear DNA Delivery. Bioorg. Med. Chem. Lett. 2017, 27, 4781–4785. [Google Scholar] [CrossRef] [PubMed]
- Siomi, H.; Dreyfuss, G. A Nuclear Localization Domain in the HnRNP A1 Protein. J. Cell Biol. 1995, 129, 551–560. [Google Scholar] [CrossRef]
- Bremner, K.H.; Seymour, L.W.; Logan, A.; Read, M.L. Factors Influencing the Ability of Nuclear Localization Sequence Peptides To Enhance Nonviral Gene Delivery. Bioconjug. Chem. 2004, 15, 152–161. [Google Scholar] [CrossRef]
- Nicolson, G.L. Mitochondrial Dysfunction and Chronic Disease: Treatment With Natural Supplements. Integr. Med. Encinitas Calif 2014, 13, 35–43. [Google Scholar]
- Tiku, V.; Tan, M.-W.; Dikic, I. Mitochondrial Functions in Infection and Immunity. Trends Cell Biol. 2020, 30, 263–275. [Google Scholar] [CrossRef]
- Gallay, P.; Stitt, V.; Mundy, C.; Oettinger, M.; Trono, D. Role of the Karyopherin Pathway in Human Immunodeficiency Virus Type 1 Nuclear Import. J. Virol. 1996, 70, 1027–1032. [Google Scholar] [CrossRef]
- Robbins, J.; Dilwortht, S.M.; Laskey, R.A.; Dingwall, C. Two Interdependent Basic Domains in Nucleoplasmin Nuclear Targeting Sequence: Identification of a Class of Bipartite Nuclear Targeting Sequence. Cell 1991, 64, 615–623. [Google Scholar] [CrossRef]
- Donkuru, M.; Badea, I.; Wettig, S.; Verrall, R.; Elsabahy, M.; Foldvari, M. Advancing Nonviral Gene Delivery: Lipid- and Surfactant-Based Nanoparticle Design Strategies. Nanomedicine 2010, 5, 1103–1127. [Google Scholar] [CrossRef]
- Preuss, M.; Tecle, M.; Shah, I.; Matthews, D.A.; D. Miller, A. Comparison between the Interactions of Adenovirus-Derived Peptides with Plasmid DNA and Their Role in Gene Delivery Mediated by Liposome–Peptide–DNA Virus-like Nanoparticles. Org. Biomol. Chem. 2003, 1, 2430–2438. [Google Scholar] [CrossRef]
- Hébert, E. Improvement of Exogenous DNA Nuclear Importation by Nuclear Localization Signal-Bearing Vectors: A Promising Way for Non-Viral Gene Therapy? Biol. Cell 2003, 95, 59–68. [Google Scholar] [CrossRef]
- Li, Q.; Huang, Y. Mitochondrial Targeted Strategies and Theirapplication for Cancer and Other Diseases Treatment. J. Pharm. Investig. 2020, 50, 271–293. [Google Scholar] [CrossRef]
- Kim, S.; Nam, H.Y.; Lee, J.; Seo, J. Mitochondrion-Targeting Peptides and Peptidomimetics: Recent Progress and Design Principles. Biochemistry 2020, 59, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Lim, K. Recent Advances in Mitochondria-Targeted Gene Delivery. Mol. J. Synth. Chem. Nat. Prod. Chem. 2018, 23, 2316. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fang, B.; Peng, B.; Wang, L.; Xue, Y.; Bai, H.; Lu, S.; Voelcker, N.H.; Li, L.; Fu, L.; et al. Recent Advances in Chemical Biology of Mitochondria Targeting. Front. Chem. 2021, 9, 321. [Google Scholar] [CrossRef]
- Kang, Y.C.; Son, M.; Kang, S.; Im, S.; Piao, Y.; Lim, K.S.; Song, M.-Y.; Park, K.-S.; Kim, Y.-H.; Pak, Y.K. Cell-Penetrating Artificial Mitochondria-Targeting Peptide-Conjugated Metallothionein 1A Alleviates Mitochondrial Damage in Parkinson’s Disease Models. Exp. Mol. Med. 2018, 50, 1–13. [Google Scholar] [CrossRef]
- Horton, K.L.; Stewart, K.M.; Fonseca, S.B.; Guo, Q.; Kelley, S.O. Mitochondria-Penetrating Peptides. Chem. Biol. 2008, 15, 375–382. [Google Scholar] [CrossRef]
- Horne, W.S.; Stout, C.D.; Ghadiri, M.R. A Heterocyclic Peptide Nanotube. J. Am. Chem. Soc. 2003, 125, 9372–9376. [Google Scholar] [CrossRef]
- Wu, J.; Li, J.; Wang, H.; Liu, C.-B. Mitochondrial-Targeted Penetrating Peptide Delivery for Cancer Therapy. Expert Opin. Drug Deliv. 2018, 15, 951–964. [Google Scholar] [CrossRef]
- Lin, R.; Zhang, P.; Cheetham, A.G.; Walston, J.; Abadir, P.; Cui, H. Dual Peptide Conjugation Strategy for Improved Cellular Uptake and Mitochondria Targeting. Bioconjug. Chem. 2015, 26, 71–77. [Google Scholar] [CrossRef]
- Ezzat, K.; EL Andaloussi, S.; Zaghloul, E.M.; Lehto, T.; Lindberg, S.; Moreno, P.M.D.; Viola, J.R.; Magdy, T.; Abdo, R.; Guterstam, P.; et al. PepFect 14, a Novel Cell-Penetrating Peptide for Oligonucleotide Delivery in Solution and as Solid Formulation. Nucleic Acids Res. 2011, 39, 5284–5298. [Google Scholar] [CrossRef] [PubMed]
- Cerrato, C.P.; Kivijärvi, T.; Tozzi, R.; Lehto, T.; Gestin, M.; Langel, Ü. Intracellular Delivery of Therapeutic Antisense Oligonucleotides Targeting MRNA Coding Mitochondrial Proteins by Cell-Penetrating Peptides. J. Mater. Chem. B 2020, 8, 10825–10836. [Google Scholar] [CrossRef] [PubMed]
- Deshayes, S.; Konate, K.; Dussot, M.; Chavey, B.; Vaissière, A.; Van, T.N.N.; Aldrian, G.; Padari, K.; Pooga, M.; Vivès, E.; et al. Deciphering the Internalization Mechanism of WRAP:SiRNA Nanoparticles. Biochim. Biophys. Acta BBA Biomembr. 2020, 1862, 183252. [Google Scholar] [CrossRef] [PubMed]
- Faria, R.; Vivés, E.; Boisguerin, P.; Sousa, A.; Costa, D. Development of Peptide-Based Nanoparticles for Mitochondrial Plasmid DNA Delivery. Polymers 2021, 13, 1836. [Google Scholar] [CrossRef]
- Jain, A.; Chugh, A. Mitochondrial Transit Peptide Exhibits Cell Penetration Ability and Efficiently Delivers Macromolecules to Mitochondria. FEBS Lett. 2016, 590, 2896–2905. [Google Scholar] [CrossRef] [PubMed]
- Fatouros, D.G.; Lamprou, D.A.; Urquhart, A.J.; Yannopoulos, S.N.; Vizirianakis, I.S.; Zhang, S.; Koutsopoulos, S. Lipid-like Self-Assembling Peptide Nanovesicles for Drug Delivery. ACS Appl. Mater. Interfaces 2014, 6, 8184–8189. [Google Scholar] [CrossRef]
- Liang, J.; Wu, W.-L.; Xu, X.-D.; Zhuo, R.-X.; Zhang, X.-Z. PH Responsive Micelle Self-Assembled from a New Amphiphilic Peptide as Anti-Tumor Drug Carrier. Colloids Surf. B Biointerfaces 2014, 114, 398–403. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, X.; Zheng, J.; Liu, D. Self-Assembled Peptide Nanotubes as Potential Nanocarriers for Drug Delivery. RSC Adv. 2014, 4, 25461. [Google Scholar] [CrossRef]
- Zhang, C.; Xue, X.; Luo, Q.; Li, Y.; Yang, K.; Zhuang, X.; Jiang, Y.; Zhang, J.; Liu, J.; Zou, G.; et al. Self-Assembled Peptide Nanofibers Designed as Biological Enzymes for Catalyzing Ester Hydrolysis. ACS Nano 2014, 8, 11715–11723. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Zhou, P.; Deng, J.; Zhao, Y.; Sun, Y.; Yang, W.; Wang, D.; Li, Z.; Hu, X.; et al. Nanoribbons Self-Assembled from Short Peptides Demonstrate the Formation of Polar Zippers between β-Sheets. Nat. Commun. 2018, 9, 5118. [Google Scholar] [CrossRef]
- Pereira-Silva, M.; Jarak, I.; Alvarez-Lorenzo, C.; Concheiro, A.; Santos, A.C.; Veiga, F.; Figueiras, A. Micelleplexes as Nucleic Acid Delivery Systems for Cancer-Targeted Therapies. J. Control. Release 2020, 323, 442–462. [Google Scholar] [CrossRef]
- Ibaraki, H.; Kanazawa, T.; Owada, M.; Iwaya, K.; Takashima, Y.; Seta, Y. Anti-Metastatic Effects on Melanoma via Intravenous Administration of Anti-NF-ΚB SiRNA Complexed with Functional Peptide-Modified Nano-Micelles. Pharmaceutics 2020, 12, 64. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Meng, L.; Huang, Q.; Zhu, Y.; Cui, W.; Cheng, Y.; Liu, R. Targeted Micelles with Chemotherapeutics and Gene Drugs to Inhibit the G1/S and G2/M Mitotic Cycle of Prostate Cancer. J. Nanobiotechnol. 2021, 19, 17. [Google Scholar] [CrossRef]
- Mazza, M.; Hadjidemetriou, M.; de Lázaro, I.; Bussy, C.; Kostarelos, K. Peptide Nanofiber Complexes with SiRNA for Deep Brain Gene Silencing by Stereotactic Neurosurgery. ACS Nano 2015, 9, 1137–1149. [Google Scholar] [CrossRef]
- Avila, L.A.; Aps, L.R.M.M.; Sukthankar, P.; Ploscariu, N.; Gudlur, S.; Šimo, L.; Szoszkiewicz, R.; Park, Y.; Lee, S.Y.; Iwamoto, T.; et al. Branched Amphiphilic Cationic Oligopeptides Form Peptiplexes with DNA: A Study of Their Biophysical Properties and Transfection Efficiency. Mol. Pharm. 2015, 12, 706–715. [Google Scholar] [CrossRef]
- Abedi-Gaballu, F.; Dehghan, G.; Ghaffari, M.; Yekta, R.; Abbaspour-Ravasjani, S.; Baradaran, B.; Dolatabadi, J.E.N.; Hamblin, M.R. PAMAM Dendrimers as Efficient Drug and Gene Delivery Nanosystems for Cancer Therapy. Appl. Mater. Today 2018, 12, 177–190. [Google Scholar] [CrossRef]
- Gorzkiewicz, M.; Konopka, M.; Janaszewska, A.; Tarasenko, I.I.; Sheveleva, N.N.; Gajek, A.; Neelov, I.M.; Klajnert-Maculewicz, B. Application of New Lysine-Based Peptide Dendrimers D3K2 and D3G2 for Gene Delivery: Specific Cytotoxicity to Cancer Cells and Transfection in Vitro. Bioorg. Chem. 2020, 95, 103504. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, A.; Moeini, H.; Bejo, M.H.; Hussein, M.Z.; Omar, A.R. Development of Tat-Conjugated Dendrimer for Transdermal DNA Vaccine Delivery. J. Pharm. Pharm. Sci. 2016, 19, 325. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tang, H.; Sun, H. Progress in Photo-Responsive Polypeptide Derived Nano-Assemblies. Micromachines 2018, 9, 296. [Google Scholar] [CrossRef]
- Mura, S.; Nicolas, J.; Couvreur, P. Stimuli-Responsive Nanocarriers for Drug Delivery. Nat. Mater. 2013, 12, 991–1003. [Google Scholar] [CrossRef]
- Lee, D.; Rejinold, N.S.; Jeong, S.D.; Kim, Y.-C. Stimuli-Responsive Polypeptides for Biomedical Applications. Polymers 2018, 10, 830. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Chen, S.; Chen, W.-H.; Lei, Q.; Liu, Y.; Zhuo, R.-X.; Zhang, X.-Z. Synergistic Gene and Drug Tumor Therapy Using a Chimeric Peptide. Biomaterials 2013, 34, 4680–4689. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.N.; Raines, R.T. Trimethyl Lock: A Trigger for Molecular Release in Chemistry, Biology, and Pharmacology. Chem. Sci. 2012, 3, 2412. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, P.; Li, H.; Ilangovan, G.; Cardounel, A.J.; Zweier, J.L.; Yamada, K.; Krishna, M.C.; Mitchell, J.B. Noninvasive Imaging of Tumor Redox Status and Its Modification by Tissue Glutathione Levels. Cancer Res. 2002, 62, 307–312. [Google Scholar] [PubMed]
- Cao, C.-Y.; Shen, Y.-Y.; Wang, J.-D.; Li, L.; Liang, G.-L. Controlled Intracellular Self-Assembly of Gadolinium Nanoparticles as Smart Molecular MR Contrast Agents. Sci. Rep. 2013, 3, 1024. [Google Scholar] [CrossRef]
- Löwik, D.W.P.M.; Leunissen, E.H.P.; van den Heuvel, M.; Hansen, M.B.; van Hest, J.C.M. Stimulus Responsive Peptide Based Materials. Chem. Soc. Rev. 2010, 39, 3394–3412. [Google Scholar] [CrossRef]
- Mackay, J.A.; Chilkoti, A. Temperature Sensitive Peptides: Engineering Hyperthermia-Directed Therapeutics. Int. J. Hyperthermia 2008, 24, 483–495. [Google Scholar] [CrossRef]
- Sigg, S.J.; Postupalenko, V.; Duskey, J.T.; Palivan, C.G.; Meier, W. Stimuli-Responsive Codelivery of Oligonucleotides and Drugs by Self-Assembled Peptide Nanoparticles. Biomacromolecules 2016, 17, 935–945. [Google Scholar] [CrossRef]
- Ouboter, D.d.B.; Schuster, T.; Shanker, V.; Heim, M.; Meier, W. Multicompartment Micelle-Structured Peptide Nanoparticles: A New Biocompatible Gene- and Drug-Delivery Tool. J. Biomed. Mater. Res. A 2014, 102, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Zou, Q.; Li, S.; Yan, X. Self-Assembled Peptide- and Protein-Based Nanomaterials for Antitumor Photodynamic and Photothermal Therapy. Adv. Mater. 2017, 29, 1605021. [Google Scholar] [CrossRef]
- Ding, J.; Zhuang, X.; Xiao, C.; Cheng, Y.; Zhao, L.; He, C.; Tang, Z.; Chen, X. Preparation of Photo-Cross-Linked PH-Responsive Polypeptide Nanogels as Potential Carriers for Controlled Drug Delivery. J. Mater. Chem. 2011, 21, 11383. [Google Scholar] [CrossRef]
- Liu, G.; Dong, C.-M. Photoresponsive Poly( S -( o -Nitrobenzyl)- l -Cysteine)- b -PEO from a l -Cysteine N -Carboxyanhydride Monomer: Synthesis, Self-Assembly, and Phototriggered Drug Release. Biomacromolecules 2012, 13, 1573–1583. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xia, X.; Dong, W.; Wang, H.; Li, L.; Ma, P.; Sheng, W.; Xu, X.; Liu, Y. Acid Sensitive Polymeric Micelles Combining Folate and Bioreducible Conjugate for Specific Intracellular SiRNA Delivery. Macromol. Biosci. 2016, 16, 759–773. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Wang, K.; Sun, X.; Li, Y.; Fu, Q.; Liang, T.; Tang, G. A Redox-Sensitive, Oligopeptide-Guided, Self-Assembling, and Efficiency-Enhanced (ROSE) System for Functional Delivery of MicroRNA Therapeutics for Treatment of Hepatocellular Carcinoma. Biomaterials 2016, 104, 192–200. [Google Scholar] [CrossRef]
- Wang, H.-X.; Song, Z.; Lao, Y.-H.; Xu, X.; Gong, J.; Cheng, D.; Chakraborty, S.; Park, J.S.; Li, M.; Huang, D.; et al. Nonviral Gene Editing via CRISPR/Cas9 Delivery by Membrane-Disruptive and Endosomolytic Helical Polypeptide. Proc. Natl. Acad. Sci. USA 2018, 115, 4903–4908. [Google Scholar] [CrossRef]
- Reissmann, S.; Filatova, M.P. New Generation of Cell-Penetrating Peptides: Functionality and Potential Clinical Application. J. Pept. Sci. 2021, 27, e3300. [Google Scholar] [CrossRef]
- Hallan, S.S.; Sguizzato, M.; Esposito, E.; Cortesi, R. Challenges in the Physical Characterization of Lipid Nanoparticles. Pharmaceutics 2021, 13, 549. [Google Scholar] [CrossRef]
- Aldosari, B.N.; Alfagih, I.M.; Almurshedi, A.S. Lipid Nanoparticles as Delivery Systems for RNA-Based Vaccines. Pharmaceutics 2021, 13, 206. [Google Scholar] [CrossRef] [PubMed]
- Reese, H.R.; Shanahan, C.C.; Proulx, C.; Menegatti, S. Peptide Science: A “Rule Model” for New Generations of Peptidomimetics. Acta Biomater. 2020, 102, 35–74. [Google Scholar] [CrossRef] [PubMed]
- Qvit, N.; Rubin, S.J.S.; Urban, T.J.; Mochly-Rosen, D.; Gross, E.R. Peptidomimetic Therapeutics: Scientific Approaches and Opportunities. Drug Discov. Today 2017, 22, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, H.J. Racemization in Reverse: Evidence That D-Amino Acid Toxicity on Earth Is Controlled by Bacteria with Racemases. PLoS ONE 2014, 9, e92101. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Cargo | Cancer Type | Ref. |
|---|---|---|---|
| RGD | siRNA | breast | [35] |
| cRGD | siRNA | brain | [36] |
| siRNA | skin | [37] | |
| iRGD | siRNA | pancreatic | [38] |
| siRNA | lung | [39] | |
| RGDfC | siRNA and doxorubicin | liver | [40] |
| CRGDK | siRNA and BAplatin | breast | [41] |
| CGKRK | siRNA | breast and brain | [42] |
| KTLLPTP | siRNA and paclitaxel | pancreatic | [43] |
| HAIYPRH | siRNA and doxorubicin | breast | [44] |
| LyP-1 and iRGD | siRNA | ovarian | [45] |
| YHWYGYTPQNVI | siRNA | liver | [46] |
| T7 | pDNA | bone | [47] |
| Cell-Penetrating Peptides | ||||||||
|---|---|---|---|---|---|---|---|---|
| Cationic | Amphipathic | Hydrophobic | ||||||
| Name(origin) | Cargo | Ref. | Name(origin) | Cargo | Ref. | Name(origin) | Cargo | Ref. |
| Diatos Peptide Vectors (DPV) | siRNA | [60] | MPG | pDNA siRNA | [61,62] | C105Y | pDNA | [63,64,65,66] |
| HIV-1 twinarginine translocation (TAT) | pDNA siRNA | [67,68,69,70,71,72,73,74] | Transportan | pDNA siRNA | [72,75,76] | K-FGF | pDNA | [77] |
| arginine-rich peptides | pDNA siRNA | [78,79,80,81,82] | NickFect (NF) | pDNA siRNA | [83,84,85,86] | Bip | pDNA | [87] |
| Polyarginine | pDNA | [75,76,88,89,90] | PepFect (PF) | pDNA mRNA | [91,92] | Melittin-derived peptides | siRNA | [93] |
| Penetratin | pDNA | [94,95,96] | MAP | siRNA | [97] | |||
| L5a | pDNA | [98,99] | Crotamine | pDNA | [100,101,102] | |||
| VP22 | pDNA | [103,104] | ||||||
| Protamine | pDNA mRNA | [105,106,107,108,109] | Antennapedia (Antp) | AON siRNA | [110,111] | |||
| Pep-1 | pDNA | [112,113] | ||||||
| CADY | siRNA | [114,115,116] | ||||||
| FGF | pDNA | [77] | ||||||
| pVEC | pDNA | [117,118] | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarvirdipour, S.; Skowicki, M.; Schoenenberger, C.-A.; Palivan, C.G. Peptide-Assisted Nucleic Acid Delivery Systems on the Rise. Int. J. Mol. Sci. 2021, 22, 9092. https://doi.org/10.3390/ijms22169092
Tarvirdipour S, Skowicki M, Schoenenberger C-A, Palivan CG. Peptide-Assisted Nucleic Acid Delivery Systems on the Rise. International Journal of Molecular Sciences. 2021; 22(16):9092. https://doi.org/10.3390/ijms22169092
Chicago/Turabian StyleTarvirdipour, Shabnam, Michal Skowicki, Cora-Ann Schoenenberger, and Cornelia G. Palivan. 2021. "Peptide-Assisted Nucleic Acid Delivery Systems on the Rise" International Journal of Molecular Sciences 22, no. 16: 9092. https://doi.org/10.3390/ijms22169092
APA StyleTarvirdipour, S., Skowicki, M., Schoenenberger, C.-A., & Palivan, C. G. (2021). Peptide-Assisted Nucleic Acid Delivery Systems on the Rise. International Journal of Molecular Sciences, 22(16), 9092. https://doi.org/10.3390/ijms22169092