
Combined Transcriptomic Analysis and RNA Interference Reveal the Effects of Methoxyfenozide on Ecdysone Signaling Pathway of Spodoptera exigua

Abstract
:Highlights
1. Introduction
2. Results
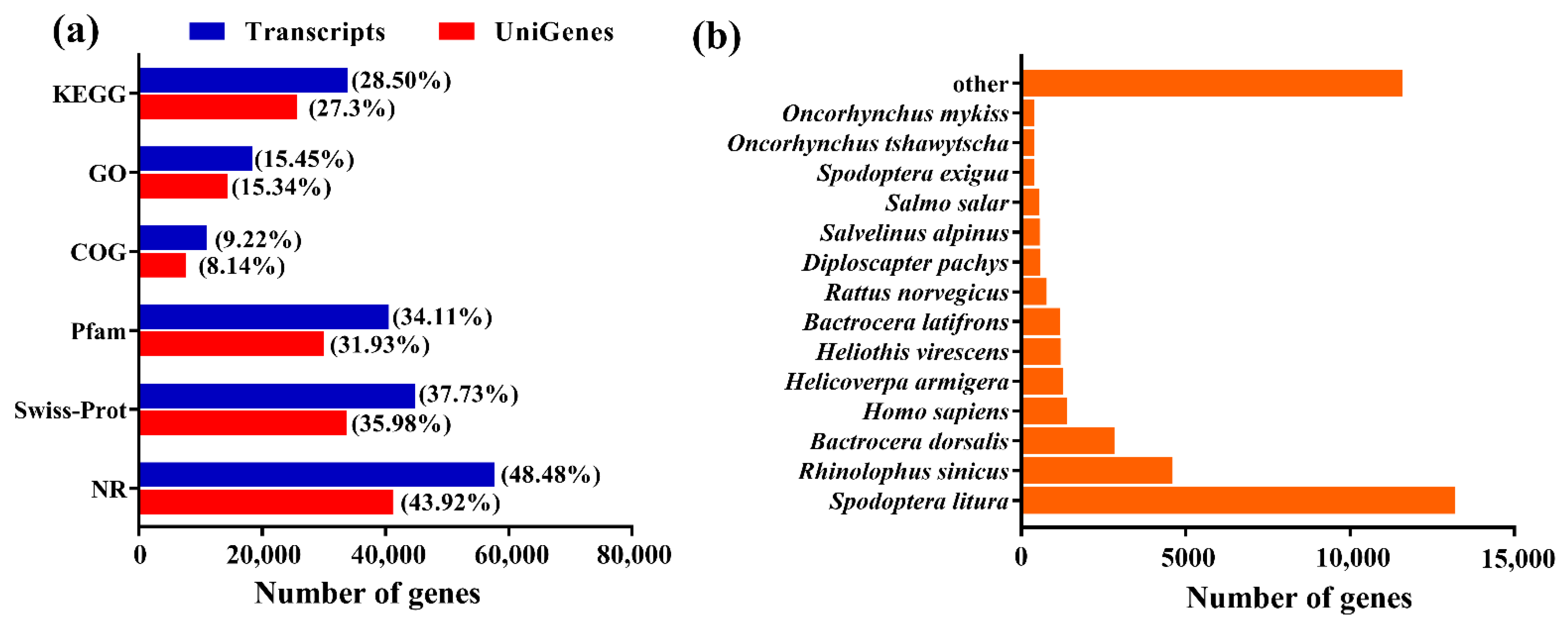
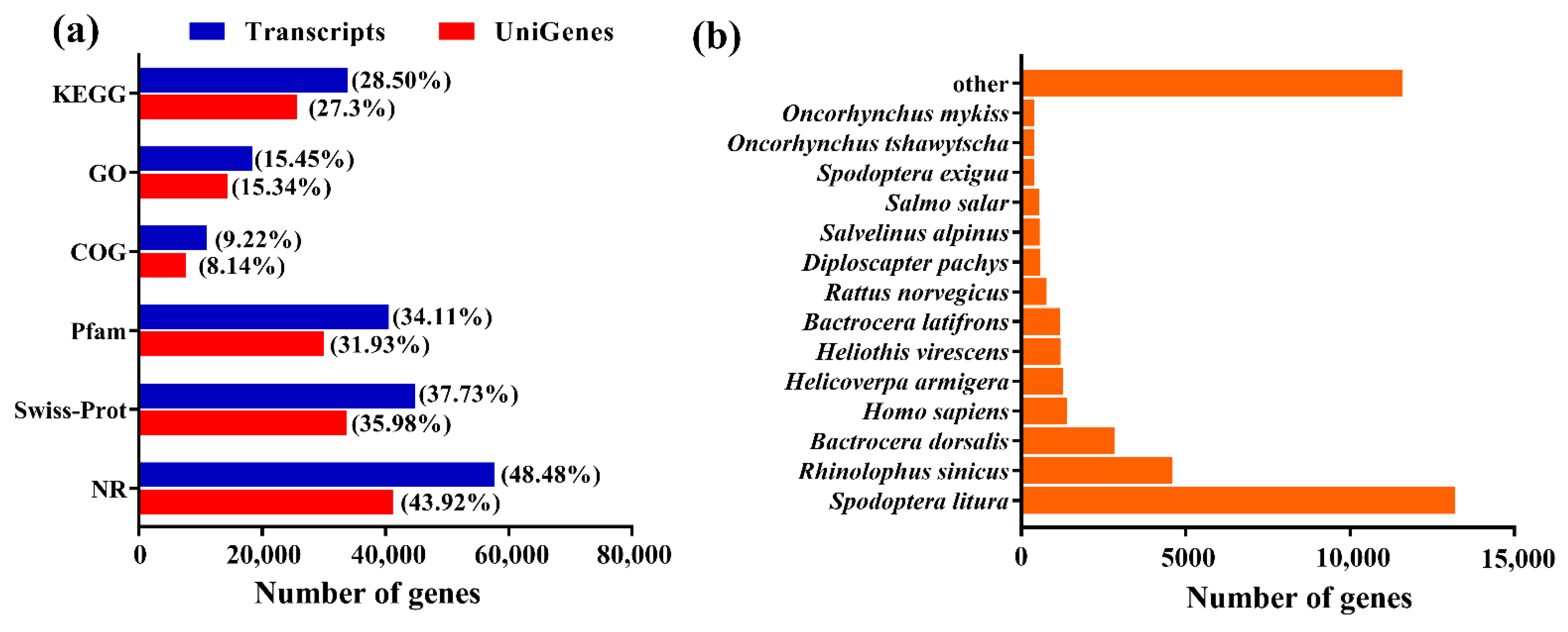
2.1. Transcriptome Data Analysis
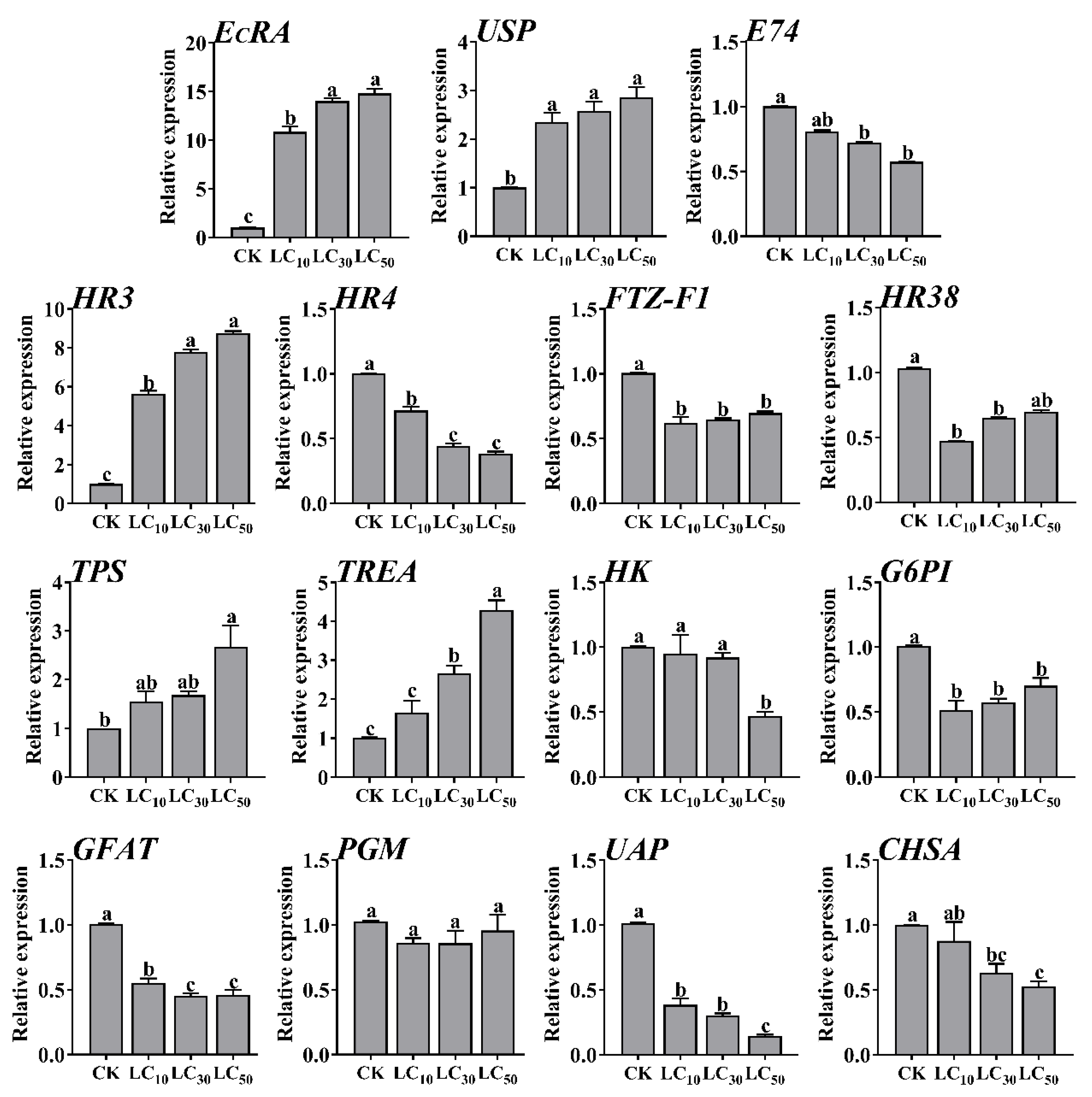
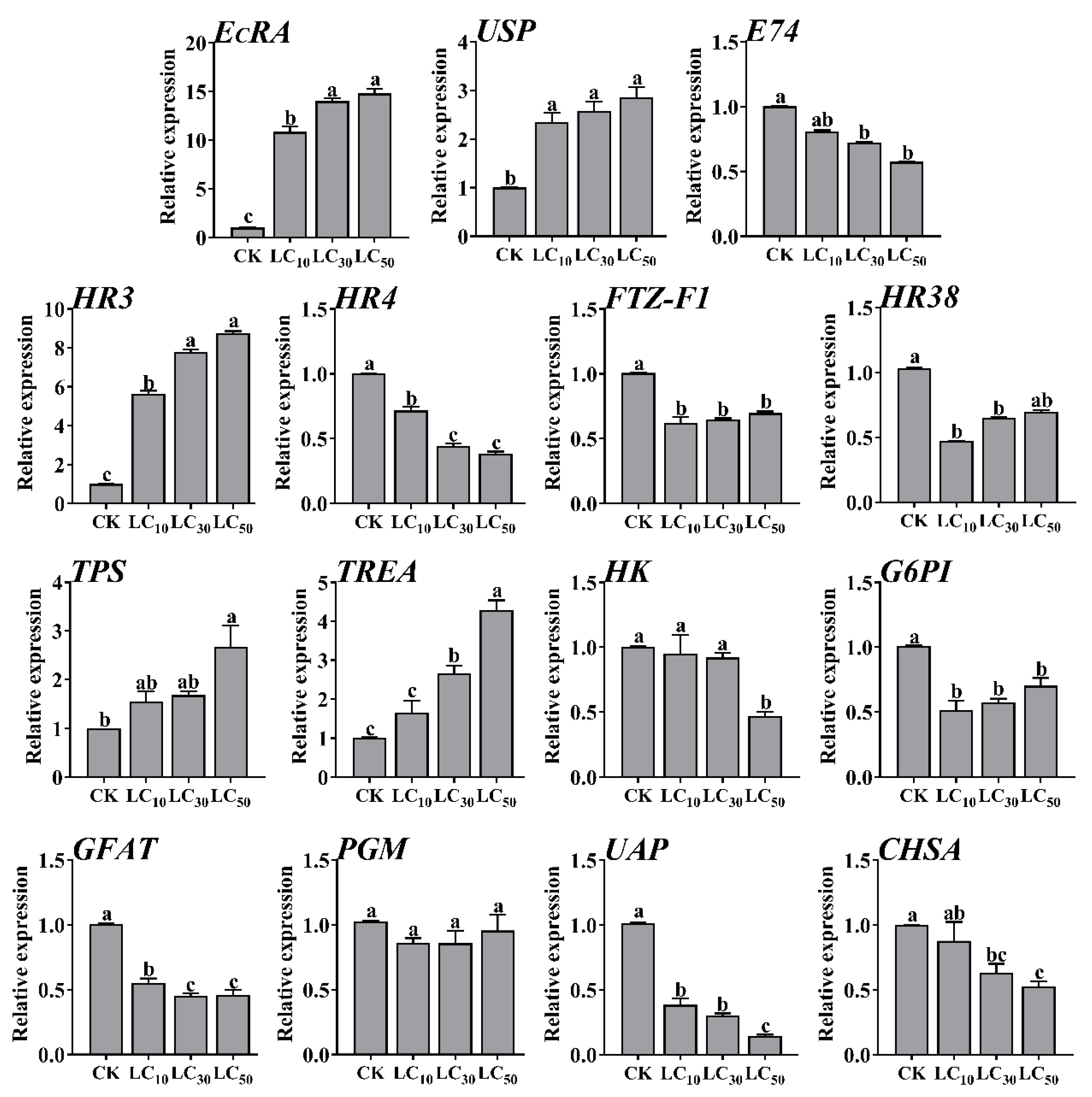
2.2. Sublethal Doses of MET Induction on Ecdysone Signaling Pathway
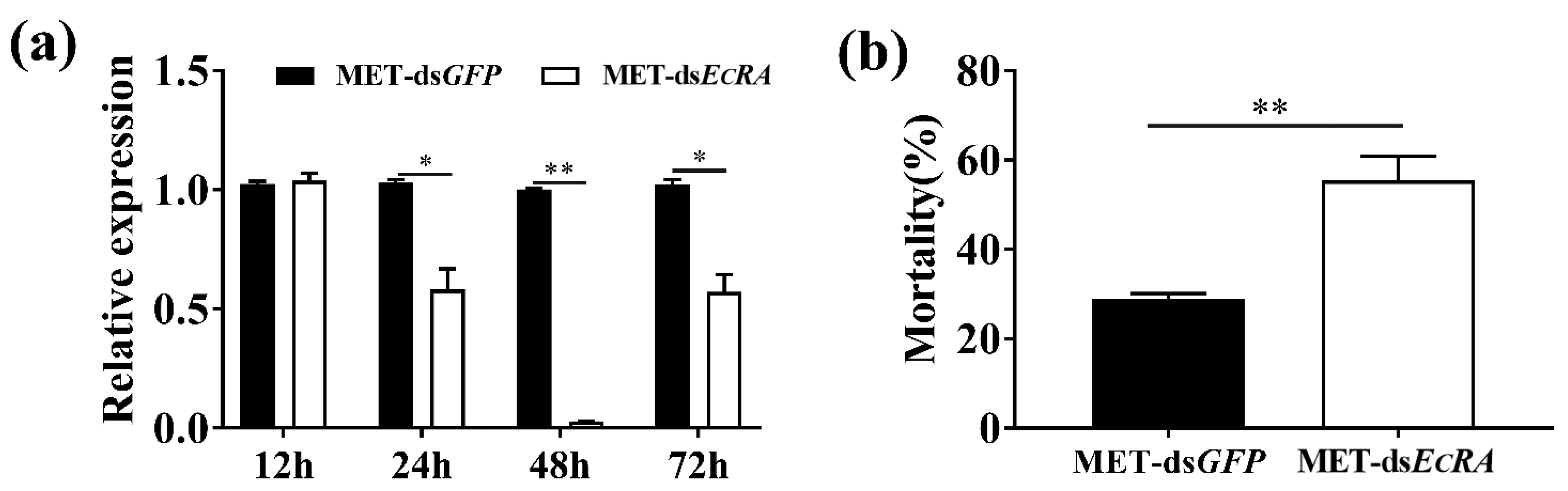
2.3. Down-Regulation of EcRA Increase the Mortality of MET to S. exigua
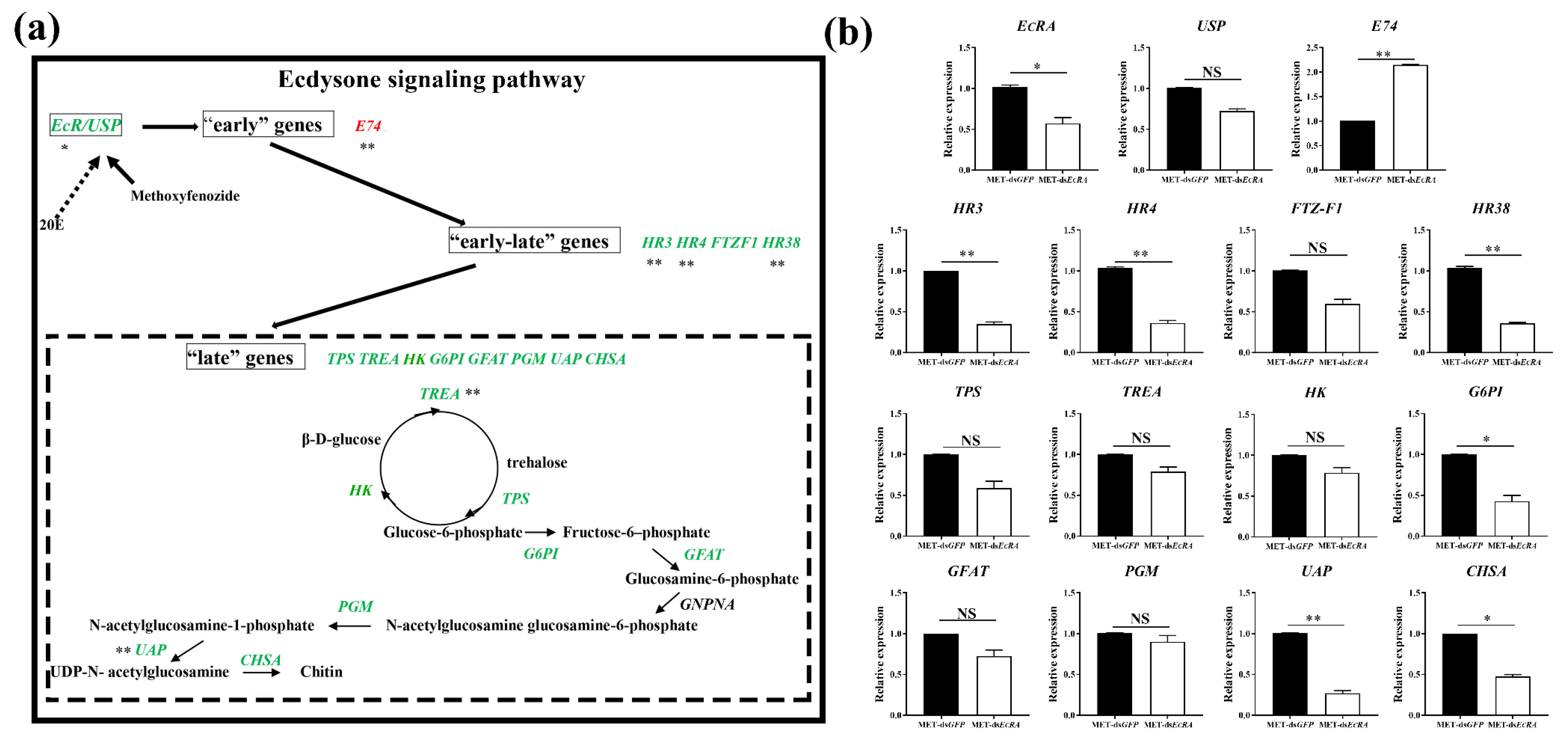
2.4. The Expression of Ecdysone Signaling Pathway Genes Are Regulated by MET-dsEcRA
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Insect Rearing
4.3. Sample Collection and RNA Isolation
4.4. Transcriptome Data Analysis
4.5. Sublethal Doses of MET Induction
4.6. Double-Stranded RNA (dsRNA) Synthesis
4.7. RNA Interference (RNAi) and Bioassays
4.8. Expression of Ecdysone Signaling Pathway Genes Regulated by MET-dsEcRA
4.9. qPCR Assays
4.10. Statistical Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lai, T.; Li, J.; Su, J. Monitoring of beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae) resistance to chlorantraniliprole in China. Pestic. Biochem. Physiol. 2011, 101, 198–205. [Google Scholar] [CrossRef]
- Ishtiaq, M.; Saleem, M.A.; Razaq, M. Monitoring of resistance in Spodoptera exigua (Lepidoptera: Noctuidae) from four districts of the Southern Punjab, Pakistan to four conventional and six new chemistry insecticides. Crop. Prot. 2012, 33, 13–20. [Google Scholar] [CrossRef]
- Su, J.; Sun, X.-X. High level of metaflumizone resistance and multiple insecticide resistance in field populations of Spodoptera exigua (Lepidoptera: Noctuidae) in Guangdong Province, China. Crop. Prot. 2014, 61, 58–63. [Google Scholar] [CrossRef]
- Ahmad, M.; Arif, M.I. Resistance of beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae) to endosulfan, organophosphorus and pyrethroid insecticides in Pakistan. Crop. Prot. 2010, 29, 1428–1433. [Google Scholar] [CrossRef]
- Kumar, M.S.; Kabra, A.N.; Min, B.; El-Dalatony, M.M.; Xiong, J.; Thajuddin, N.; Lee, D.S.; Jeon, B.H. Insecticides induced biochemical changes in freshwater microalga Chlamydomonas mexicana. Environ. Sci. Pollut. Res. Int. 2016, 23, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Osorio, A.; Martinez, A.M.; Schneider, M.I.; Diaz, O.; Corrales, J.L.; Aviles, M.C.; Smagghe, G.; Pineda, S. Monitoring of beet armyworm resistance to spinosad and methoxyfenozide in Mexico. Pest. Manag. Sci. 2008, 64, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Guo, B.; Cui, J.; Dong, Y.; Cui, L.; Zhang, L.; Yang, Q.; Yang, X. New lead discovery of insect growth regulators based on the scaffold hopping strategy. Bioorg. Med. Chem. Lett. 2020, 30, 127500. [Google Scholar] [CrossRef]
- Petryk, A.; Warren, J.T.; Marqués, G.; Jarcho, M.P.; Gilbert, L.I.; Kahler, J.; Parvy, J.P.; Li, Y.; Dauphin-Villemant, C.; O’Connor, M.B. Shade is the Drosophila P450 enzyme that mediates the hydroxylation of ecdysone to the steroid insect molting hormone 20-hydroxyecdysone. Proc. Natl. Acad. Sci. USA 2003, 100, 13773–13778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.-C.; Liu, X.-P.; Fu, K.-Y.; Shi, J.-F.; Lü, F.-G.; Li, G.Q. Functions of nuclear receptor HR3 during larval-pupal molting in Leptinotarsa decemlineata (Say) revealed by in vivo RNA interference. Insect Biochem. Mol. Biol. 2015, 63, 22–33. [Google Scholar] [CrossRef]
- Xu, Q.-Y.; Meng, Q.; Deng, P.; Guo, W.-C.; Li, G.Q. Leptinotarsa hormone receptor 4 (HR4) tunes ecdysteroidogenesis and mediates 20-hydroxyecdysone signaling during larval-pupal metamorphosis. Insect Biochem. Mol. Biol. 2017, 94, 50–60. [Google Scholar] [CrossRef]
- Liu, X.-P.; Fu, K.-Y.; Lü, F.-G.; Meng, Q.; Guo, W.-C.; Li, G.Q. Involvement of FTZ-F1 in the regulation of pupation in Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 2014, 55, 51–60. [Google Scholar] [CrossRef]
- Yogindran, S.; Rajam, M.V. Artificial miRNA-mediated silencing of ecdysone receptor (EcR) affects larval development and oogenesis in Helicoverpa armigera. Insect Biochem. Mol. Biol. 2016, 77, 21–30. [Google Scholar] [CrossRef]
- Chen, J.; Tang, B.; Chen, H.; Yao, Q.; Huang, X.; Chen, J.; Zhang, D.; Zhang, W. Different functions of the insect soluble and membrane-bound trehalase genes in chitin biosynthesis revealed by RNA interference. PLoS ONE 2010, 5, e10133. [Google Scholar] [CrossRef]
- Yao, Q.; Zhang, D.; Tang, B.; Chen, J.; Chen, J.; Lu, L.; Zhang, W. Identification of 20-hydroxyecdysone late-response genes in the chitin biosynthesis pathway. PLoS ONE 2010, 5, e14058. [Google Scholar] [CrossRef]
- Chen, J.; Li, T.; Pang, R. miR-2703 regulates the chitin biosynthesis pathway by targeting chitin synthase 1a in Nilaparvata lugens. Insect Mol. Biol. 2020, 29, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhao, L.; Shen, Q.; Xie, G.; Wang, S.; Tang, B. Knockdown of two trehalose-6-phosphate synthases severely affects chitin metabolism gene expression in the brown planthopper Nilaparvata lugens. Pest. Manag. Sci. 2017, 73, 206–216. [Google Scholar] [CrossRef]
- Gu, S.H.; Lin, J.L.; Lin, P.L. PTTH-stimulated ERK phosphorylation in prothoracic glands of the silkworm, Bombyx mori: Role of Ca(2+)/calmodulin and receptor tyrosine kinase. J. Insect Physiol. 2010, 56, 93–101. [Google Scholar] [CrossRef]
- Scieuzo, C.; Nardiello, M.; Salvia, R.; Pezzi, M.; Chicca, M.; Leis, M.; Bufo, S.A.; Vinson, S.B.; Rao, A.; Vogel, H.; et al. Ecdysteroidogenesis and development in Heliothis virescens (Lepidoptera: Noctuidae): Focus on PTTH-stimulated pathways. J. Insect Physiol. 2018, 107, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Rybczynski, R.; Bell Sc Fau-Gilbert, L.I.; Gilbert, L.I. Activation of an extracellular signal-regulated kinase (ERK) by the insect prothoracicotropic hormone. Mol. Cell. Endocrinol. 2001, 184, 1–11. [Google Scholar] [CrossRef]
- Yao, Q.; Xu, S.; Dong, Y.; Quan, L.; Chen, B. Ecdysone Receptor (EcR) and Ultraspiracle Protein (USP) Genes From Conopomorpha sinensis Bradley Eggs: Identification and Expression in Response to Insecticides. Front. Physiol. 2020, 11, 851. [Google Scholar] [CrossRef]
- Smagghe, G.; Böhm, G.-A.; Richter, K.; Degheele, D. Effect of Nonsteroidal Ecdysteroid Agonists on Ecdysteroid Titer in Spodoptera exigua and Leptinotarsa decemlineata. J. Insect Physiol. 1995, 41, 971–974. [Google Scholar] [CrossRef]
- Sun, X.; Song, Q.; Barrett, B. Effects of ecdysone agonists on the expression of EcR, USP and other specific proteins in the ovaries of the codling moth (Cydia pomonella L.). Insect Biochem. Mol. Biol. 2003, 33, 829–840. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, W.; Hu, H.; Ma, X.; Li, Q.; Song, X.; Ren, X.; Ma, Y. Joint toxicity of methoxyfenozide and lufenuron on larvae of Spodoptera exigua Hübner (Lepidoptera: Noctuidae). J. Asia-Pac. Entomol. 2019, 22, 795–801. [Google Scholar] [CrossRef]
- Aguirre, O.U.; Martinez, A.M.; Campos-Garcia, J.; Hernandez, L.A.; Figueroa, J.I.; Lobit, P.; Vinuela, E.; Chavarrieta, J.M.; Smagghe, G.; Pineda, S. Foliar persistence and residual activity of methoxyfenozide against beet armyworm (Lepidoptera: Noctuidae). Insect Sci. 2013, 20, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Mosallanejad, H.; Soin, T.; Smagghe, G. Selection for resistance to methoxyfenozide and 20-hydroxyecdysone in cells of the beet armyworm, Spodoptera exigua. Arch. Insect Biochem. Physiol. 2008, 67, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, A.R.; Fresia, P.; Consoli, F.L.; Omoto, C. Comparative transcriptome analysis of lufenuron-resistant and susceptible strains of Spodoptera frugiperda (Lepidoptera: Noctuidae). BMC Genom. 2015, 16, 985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraudo, M.; Audant, P.; Feyereisen, R.; Le Goff, G. Nuclear receptors HR96 and ultraspiracle from the fall armyworm (Spodoptera frugiperda), developmental expression and induction by xenobiotics. J. Insect Physiol. 2013, 59, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Q.; Wang, N.M.; Li, J.J.; Jin, M.H.; Xue, C.B. Reduced fecundity and regulation of reproductive factors in flubendiamide-resistant strains of Plutella xylostella. Pestic. Biochem. Physiol. 2020, 169, 104668. [Google Scholar] [CrossRef]
- Tang, B.; Yang, M.; Shen, Q.; Xu, Y.; Wang, H.; Wang, S. Suppressing the activity of trehalase with validamycin disrupts the trehalose and chitin biosynthesis pathways in the rice brown planthopper, Nilaparvata lugens. Pestic. Biochem. Physiol. 2017, 137, 81–90. [Google Scholar] [CrossRef]
- Chen, J.; Lu, Z.; Li, M.; Mao, T.; Wang, H.; Li, F.; Sun, H.; Dai, M.; Ye, W.; Li, B. The mechanism of sublethal chlorantraniliprole exposure causing silkworm pupation metamorphosis defects. Pest. Manag. Sci. 2020, 76, 2838–2845. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, L.; Liu, X.; Peng, Y.; Liang, G.; Xiao, H. Dissecting the roles of FTZ-F1 in larval molting and pupation, and the sublethal effects of methoxyfenozide on Helicoverpa armigera. Pest. Manag. Sci. 2021, 77, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.F.; Fu, J.; Mu, L.L.; Guo, W.C.; Li, G.Q. Two Leptinotarsa uridine diphosphate N-acetylglucosamine pyrophosphorylases are specialized for chitin synthesis in larval epidermal cuticle and midgut peritrophic matrix. Insect Biochem. Mol. Biol. 2016, 68, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ren, X.; Liu, Y.; Smagghe, G.; Liang, P.; Gao, X. MiR-189942 regulates fufenozide susceptibility by modulating ecdysone receptor isoform B in Plutella xylostella (L.). Pestic. Biochem. Physiol. 2020, 163, 235–240. [Google Scholar] [CrossRef]
- Xu, Q.-Y.; Deng, P.; Zhang, Q.; Li, A.; Fu, K.-Y.; Guo, W.-C.; Li, G.Q. Ecdysone receptor isoforms play distinct roles in larval-pupal-adult transition in Leptinotarsa decemlineata. Insect Sci. 2019, 27, 487–499. [Google Scholar] [CrossRef]
- Wu, J.J.; Mu, L.L.; Kang, W.N.; Ze, L.J.; Shen, C.H.; Jin, L.; Anjum, A.A.; Li, G.Q. RNA interference targeting ecdysone receptor blocks the larval-pupal transition in Henosepilachna vigintioctopunctata. Insect Sci. 2021, 28, 419–429. [Google Scholar] [CrossRef]
- Lenaerts, C.; Van Wielendaele, P.; Peeters, P.; Vanden Broeck, J.; Marchal, E. Ecdysteroid signalling components in metamorphosis and development of the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2016, 75, 10–23. [Google Scholar] [CrossRef]
- Shen, C.; Xu, Q.-Y.; Mu, L.-L.; Fu, K.-Y.; Guo, W.-C.; Li, G.Q. Involvement of Leptinotarsa hormone receptor 38 in the larval-pupal transition. Gene 2020, 751, 144779. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.-Y.; Deng, P.; Li, A.; Zhang, Q.; Mu, L.-L.; Fu, K.-Y.; Guo, W.-C.; Li, G.Q. Functional characterization of ultraspiracle in Leptinotarsa decemlineata using RNA interference assay. Insect Mol. Biol. 2019, 28, 676–688. [Google Scholar] [CrossRef]
- Ren, X.-L.; Chen, R.-R.; Zhang, Y.; Ma, Y.; Cui, J.-J.; Han, Z.-J.; Mu, L.-L.; Li, G.Q. A Spodoptera exigua Cadherin Serves as a Putative Receptor for Bacillus thuringiensis Cry1Ca Toxin and Shows Differential Enhancement of Cry1Ca and Cry1Ac Toxicity. Appl. Environ. Microbiol. 2013, 79, 5576–5583. [Google Scholar] [CrossRef] [Green Version]
- Luan, J.B.; Ghanim, M.; Liu, S.S.; Czosnek, H. Silencing the ecdysone synthesis and signaling pathway genes disrupts nymphal development in the whitefly. Insect Biochem. Mol. Biol. 2013, 43, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Swevers, L.; Soin, T.; Mosallanejad, H.; Iatrou, K.; Smagghe, G. Ecdysteroid signaling in ecdysteroid-resistant cell lines from the polyphagous noctuid pest Spodoptera exigua. Insect Biochem. Mol. Biol. 2008, 38, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X.; Niu, C.D.; Zhang, Y.; Jia, Y.L.; Zhang, Y.J.; Zhang, Y.; Zhang, Y.Q.; Gao, C.F.; Wu, S.F. The NompC channel regulates Nilaparvata lugens proprioception and gentle-touch response. Insect Biochem. Mol. Biol. 2019, 106, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yuan, M.; Shakeel, M.; Zhang, Y.-J.; Wang, S.; Wang, X.; Zhan, S.; Kang, T.; Li, J. Selection and Evaluation of Reference Genes for Expression Analysis Using qRT-PCR in the Beet Armyworm Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae). PLoS ONE 2014, 9, e84730. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Error Rate (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|---|
| Control group 1 | 44,923,180 | 6,783,400,180 | 44,335,152 | 6,636,272,205 | 0.0236 | 98.66 | 95.48 | 46.75 |
| Control group 2 | 46,298,736 | 6,991,109,136 | 45,854,460 | 6,864,878,635 | 0.0230 | 98.88 | 96.17 | 49.38 |
| Control group 3 | 46,272,298 | 6,987,116,998 | 45,733,494 | 6,851,482,844 | 0.0231 | 98.85 | 96.06 | 49.08 |
| MET (LC30) 1 | 48,311,340 | 7,295,012,340 | 47,908,742 | 7,186,907,127 | 0.0227 | 99.01 | 96.49 | 49.14 |
| MET (LC30) 2 | 49,407,600 | 7,460,547,600 | 48,903,418 | 7,321,874,857 | 0.0229 | 98.93 | 96.28 | 49.00 |
| MET (LC30) 3 | 48,680,700 | 7,350,785,700 | 48,177,980 | 7,222,758,714 | 0.0229 | 98.91 | 96.21 | 48.38 |
| UniGenes | Gene Bank | Homologous Genes | Log2FC | Padjust | Organisms |
|---|---|---|---|---|---|
| TRINITY_DN68411_c0_g1 | ACD39740.1 | ultraspiracle protein (USP) | 1.43053001 | 0.000710809 | S. exigua |
| TRINITY_DN68123_c0_g1 | XP_022816977.1 | probable nuclear hormone receptor HR3 isoform X6 (HR3) | 1.319971145 | 0.407707359 | S. litura |
| TRINITY_DN67384_c0_g4 | GU296540.1 | ecdysone receptor A (EcRA) | 0.511687018 | 0.509814187 | S. exigua |
| TRINITY_DN66888_c3_g2 | XP_022822951.1 | nuclear hormone receptor FTZ-F1 beta (FTZ-F1) | −0.85620132 | 0.101532649 | S. litura |
| TRINITY_DN35412_c0_g1 | XM_022967707.1 | PREDICTED: ecdysone-induced protein 74EF (E74) | −0.86981036 | 1 | S. litura |
| TRINITY_DN62913_c0_g1 | XP_021187825.1 | hormone receptor 4 isoform X1 (HR4) | −3.19753521 | 8.62 × 10−9 | H. armigera |
| TRINITY_DN68583_c0_g4 | XP_022820857.1 | probable nuclear hormone receptor HR38 (HR38) | −6.28201224 | 3.90 × 10−40 | S. litura |
| TRINITY_DN65723_c0_g2 | ACV97159.1 | glucose-6-phosphate isomerase (G6PI) | 1.468813444 | 0.000855695 | S. exigua |
| TRINITY_DN66137_c0_g2 | ABY86218.1 | trehalase-1 (TREA) | 0.83888701 | 0.486117572 | S. exigua |
| TRINITY_DN67124_c0_g6 | ABM66814.2 | trehalose 6-phosphate synthase (TPS) | −0.15968318 | 0.840417477 | S. exigua |
| TRINITY_DN63532_c0_g3 | XP_022834981.1 | hexokinase type 2 isoform X6 (HK) | −0.54980536 | 0.337950088 | S. litura |
| TRINITY_DN68071_c2_g3 | XP_022827210.1 | Phosphoacetyl glucosamine mutase (PGM) | −0.81471571 | 0.132596543 | S. litura |
| TRINITY_DN65476_c0_g2 | AAZ03545.1 | chitin synthase A (CHSA) | −1.99147489 | 2.42017 × 10−8 | S. exigua |
| TRINITY_DN66504_c1_g4 | XP_022821069.1 | glutamine--fructose-6-phosphate aminotransferase (GFAT) | −4.07457569 | 3.01573 × 10−29 | S. litura |
| TRINITY_DN67977_c1_g2 | ACN29686.1 | UDP-N-acetylglucosamine pyrophosphorylase (UAP) | −5.02829161 | 1.86507 × 10−40 | S. exigua |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Ma, Y.; Ma, X.; Hu, H.; Wang, D.; Song, X.; Ren, X.; Ma, Y. Combined Transcriptomic Analysis and RNA Interference Reveal the Effects of Methoxyfenozide on Ecdysone Signaling Pathway of Spodoptera exigua. Int. J. Mol. Sci. 2021, 22, 9080. https://doi.org/10.3390/ijms22169080
Zhang Z, Ma Y, Ma X, Hu H, Wang D, Song X, Ren X, Ma Y. Combined Transcriptomic Analysis and RNA Interference Reveal the Effects of Methoxyfenozide on Ecdysone Signaling Pathway of Spodoptera exigua. International Journal of Molecular Sciences. 2021; 22(16):9080. https://doi.org/10.3390/ijms22169080
Chicago/Turabian StyleZhang, Zhixian, Yajie Ma, Xiaoyan Ma, Hongyan Hu, Dan Wang, Xianpeng Song, Xiangliang Ren, and Yan Ma. 2021. "Combined Transcriptomic Analysis and RNA Interference Reveal the Effects of Methoxyfenozide on Ecdysone Signaling Pathway of Spodoptera exigua" International Journal of Molecular Sciences 22, no. 16: 9080. https://doi.org/10.3390/ijms22169080
APA StyleZhang, Z., Ma, Y., Ma, X., Hu, H., Wang, D., Song, X., Ren, X., & Ma, Y. (2021). Combined Transcriptomic Analysis and RNA Interference Reveal the Effects of Methoxyfenozide on Ecdysone Signaling Pathway of Spodoptera exigua. International Journal of Molecular Sciences, 22(16), 9080. https://doi.org/10.3390/ijms22169080