
Acute, Low-Dose Neutron Exposures Adversely Impact Central Nervous System Function

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
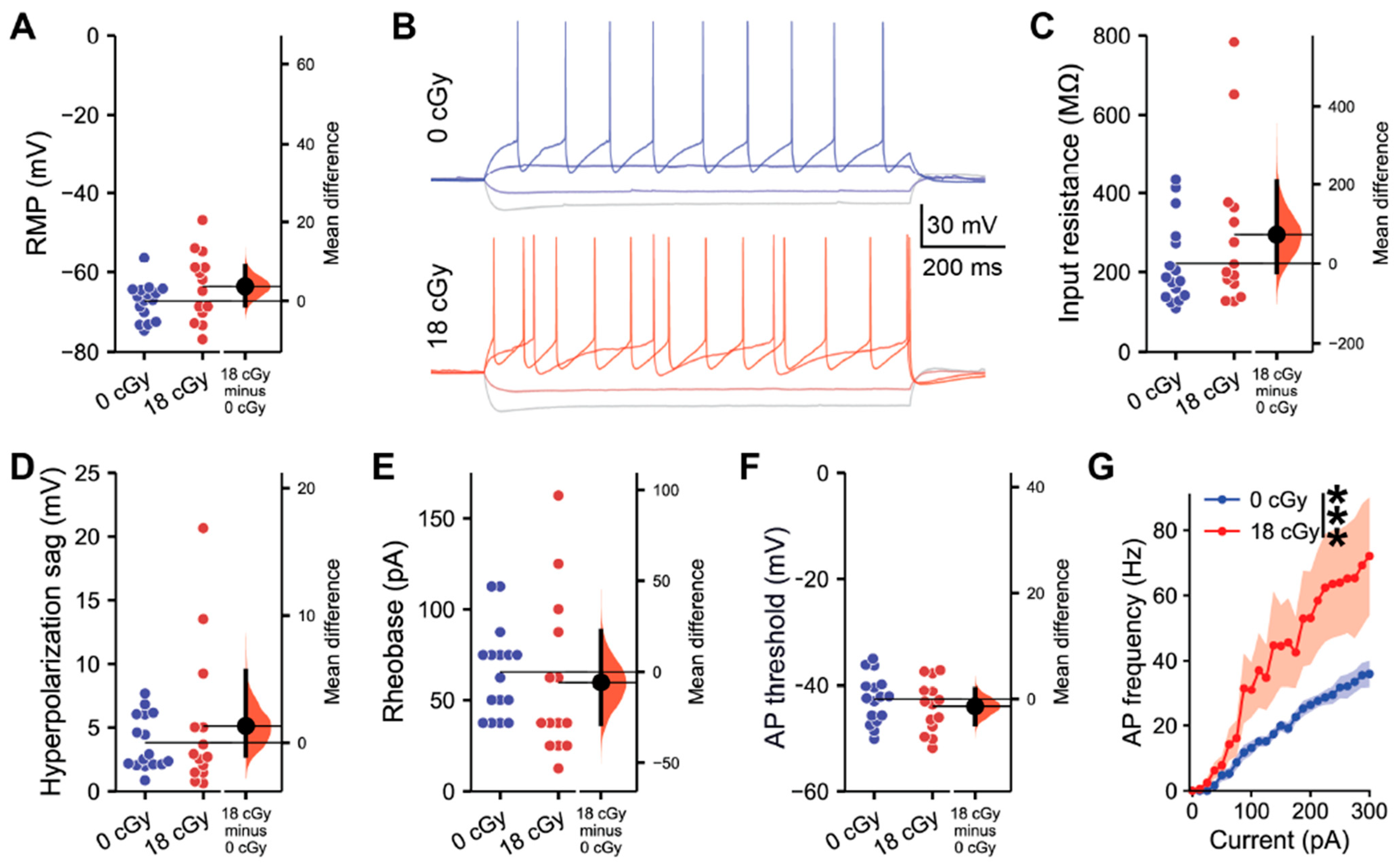
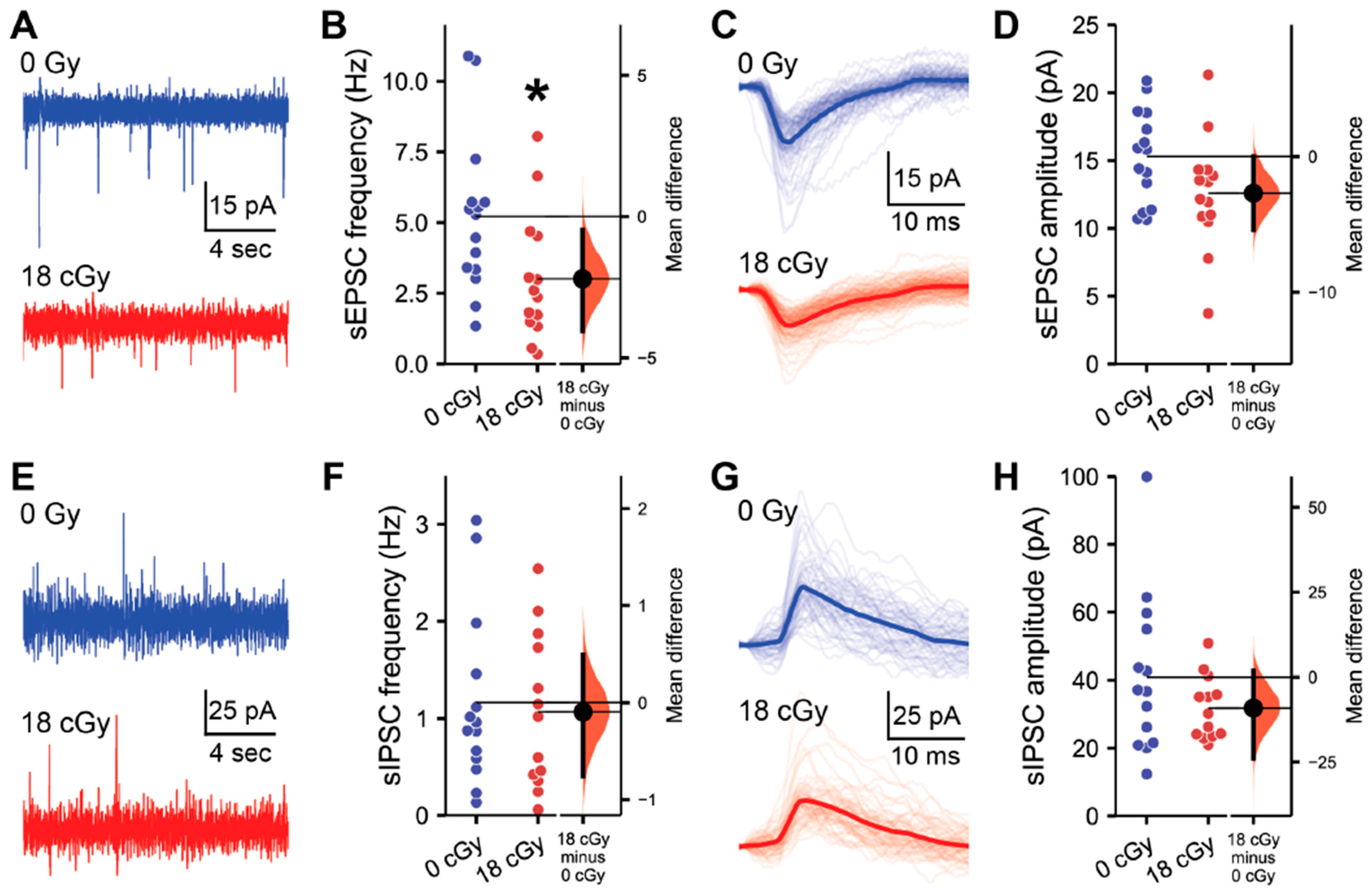
2.1. Acute Neutron Irradiation Alters Excitatory Hippocampal Neuronal Activity
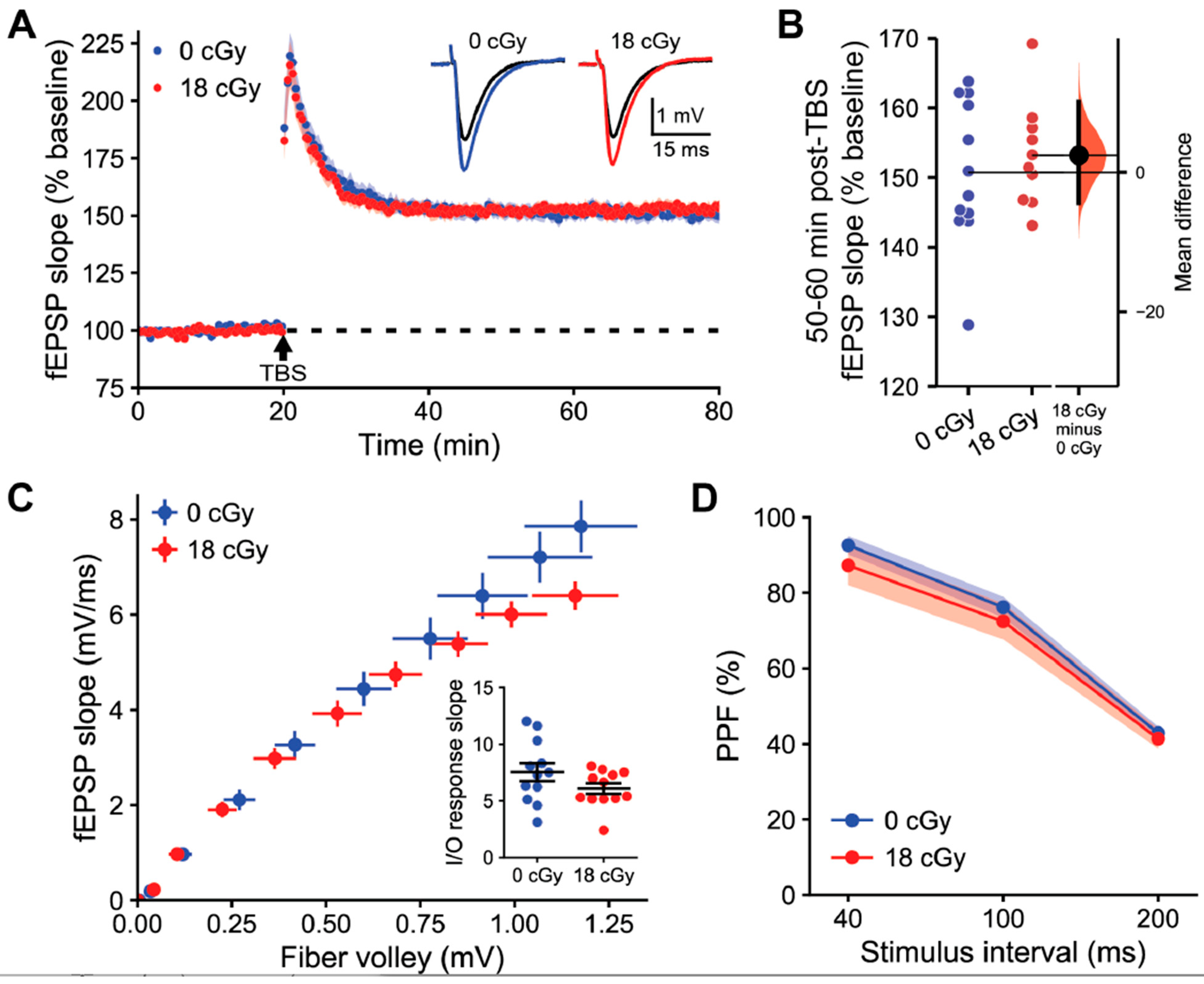
2.2. Hippocampal Long-Term Synaptic Plasticity Is Not Disrupted following Acute Neutron Irradiation
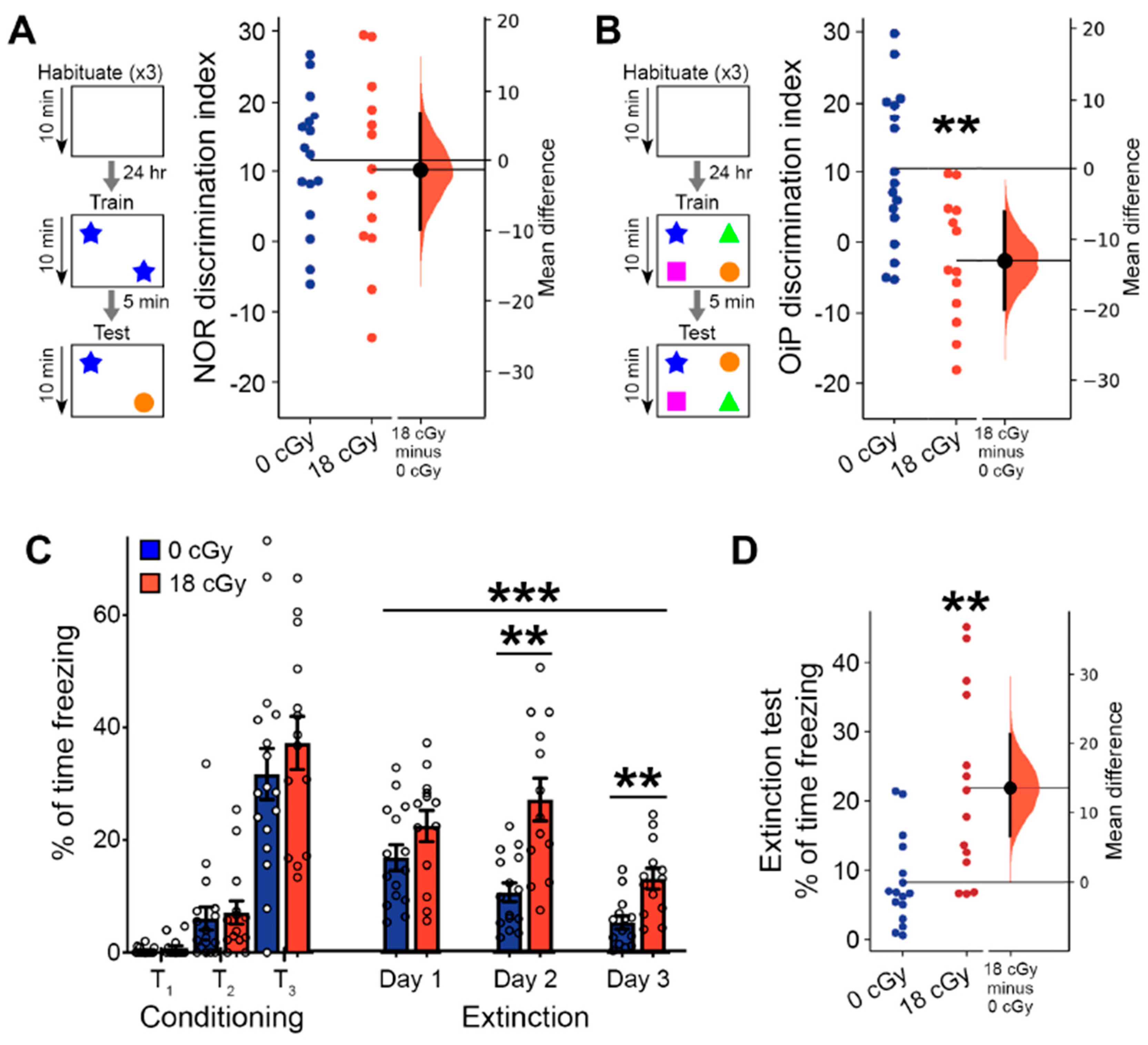
2.3. Acute Neutron Irradiated Induces Persistent Learning and Memory Deficits
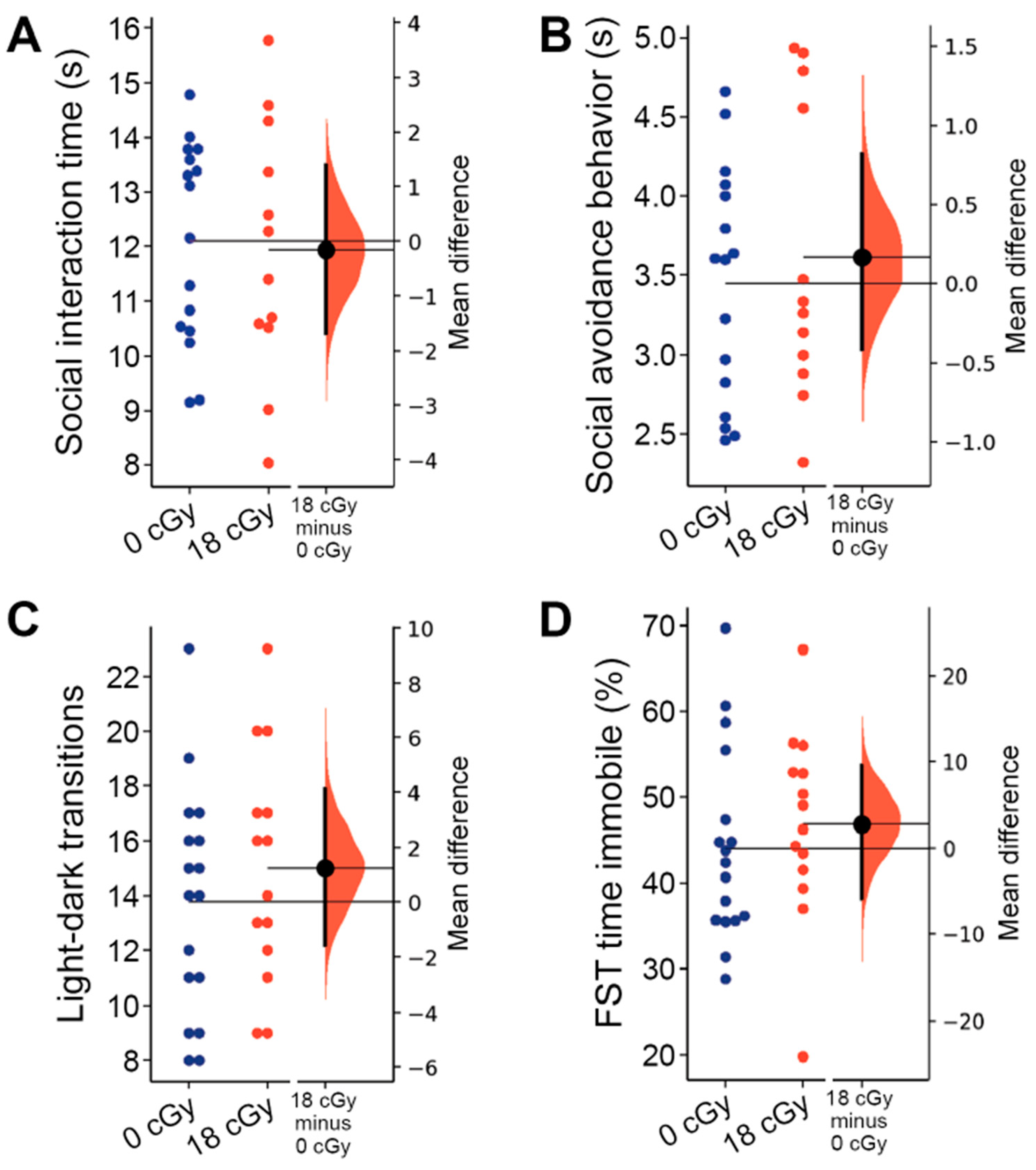
2.4. Social and Internalizing Behaviors Are Not Altered by Acute Neutron Irradiation
3. Discussion
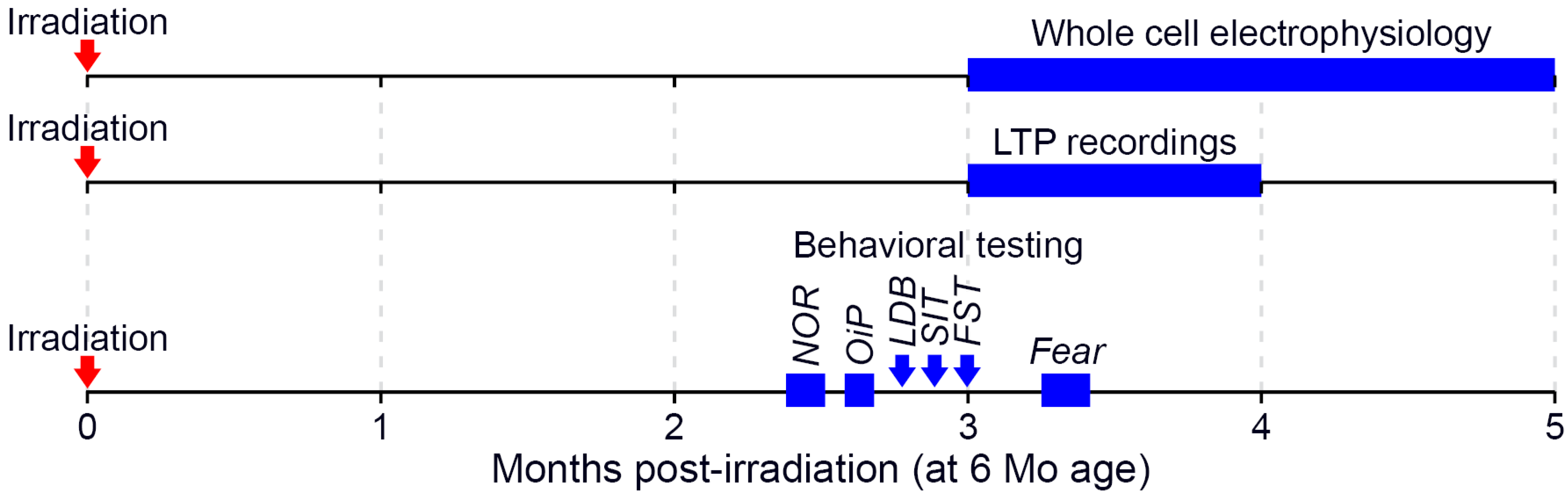
4. Materials and Methods
4.1. Animals
4.2. Neutron Irradiation and Dosimetry
4.3. Whole Cell Electrophysiology
4.4. Extracellular Field Recordings
4.5. Behavioral Testing
4.5.1. Episodic and Spatial Memory Testing
4.5.2. Social Interaction Testing
4.5.3. Anxiety- and Depression-Like Behavior Testing
4.5.4. Fear Extinction Testing
5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cucinotta, F.A.; Alp, M.; Sulzman, F.M.; Wang, M. Space radiation risks to the central nervous system. Life Sci. Space Res. 2014, 2, 54–69. [Google Scholar] [CrossRef] [Green Version]
- Nelson, G.A. Space Radiation and Human Exposures, A Primer. Radiat. Res. 2016, 185, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, L.C.; Slaba, T.C.; Guida, P.; Rusek, A. NASA’s first ground-based Galactic Cosmic Ray Simulator: Enabling a new era in space radiobiology research. PLoS Biol. 2020, 18, e3000669. [Google Scholar] [CrossRef]
- Slaba, T.C.; Blattnig, S.R.; Norbury, J.W.; Rusek, A.; La Tessa, C. Reference field specification and preliminary beam selection strategy for accelerator-based GCR simulation. Life Sci. Space Res. 2016, 8, 52–67. [Google Scholar] [CrossRef]
- Dobynde, M.I.; Shprits, Y.Y. Radiation environment created with GCRs inside a spacecraft. Life Sci. Space Res. 2020, 24, 116–121. [Google Scholar] [CrossRef]
- Parihar, V.K.; Allen, B.D.; Tran, K.K.; Chmielewski, N.N.; Craver, B.M.; Martirosian, V.; Morganti, J.M.; Rosi, S.; Vlkolinský, R.; Acharya, M.M.; et al. Targeted Overexpression of Mitochondrial Catalase Prevents Radiation-Induced Cognitive Dysfunction. Antioxid. Redox Signal. 2015, 22, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.M.; DeCicco-Skinner, K.L.; Hienz, R.D. Deficits in sustained attention and changes in dopaminergic protein levels following exposure to proton radiation are related to basal dopaminergic function. PLoS ONE 2015, 10, e0144556. [Google Scholar] [CrossRef]
- Raber, J.; Allen, A.R.; Sharma, S.; Allen, B.; Rosi, S.; Olsen, R.H.J.; Davis, M.J.; Eiwaz, M.; Fike, J.R.; Nelson, G.A. Effects of Proton and Combined Proton and 56Fe Radiation on the Hippocampus. Radiat. Res. 2016, 185, 20–30. [Google Scholar] [CrossRef]
- Parihar, V.K.; Maroso, M.; Syage, A.; Allen, B.D.; Angulo, M.C.; Soltesz, I.; Limoli, C.L. Persistent nature of alterations in cognition and neuronal circuit excitability after exposure to simulated cosmic radiation in mice. Exp. Neurol. 2018, 305, 44–55. [Google Scholar] [CrossRef]
- Rabin, B.M.; Carrihill-Knoll, K.L.; Shukitt-Hale, B. Comparison of the Effectiveness of Exposure to Low-LET Helium Particles (4He) and Gamma Rays (137Cs) on the Disruption of Cognitive Performance. Radiat. Res. 2015, 184, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Krukowski, K.; Feng, X.; Paladini, M.S.; Chou, A.; Sacramento, K.; Grue, K.; Riparip, L.K.; Jones, T.; Campbell-Beachler, M.; Nelson, G.; et al. Temporary microglia-depletion after cosmic radiation modifies phagocytic activity and prevents cognitive deficits. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Carr, H.; Alexander, T.C.; Groves, T.; Kiffer, F.; Wang, J.; Price, E.; Boerma, M.; Allen, A.R. Early effects of 16O radiation on neuronal morphology and cognition in a murine model. Life Sci. Space Res. 2018, 17, 63–73. [Google Scholar] [CrossRef]
- Parihar, V.K.; Allen, B.D.; Tran, K.K.; Macaraeg, T.G.; Chu, E.M.; Kwok, S.F.; Chmielewski, N.N.; Craver, B.M.; Baulch, J.E.; Acharya, M.M.; et al. What happens to your brain on the way to Mars. Sci. Adv. 2015, 1, e1400256. [Google Scholar] [CrossRef] [Green Version]
- Howe, A.; Kiffer, F.; Alexander, T.C.; Sridharan, V.; Wang, J.; Ntagwabira, F.; Rodriguez, A.; Boerma, M.; Allen, A.R. Long-term changes in cognition and physiology after low-dose 16 O irradiation. Int. J. Mol. Sci. 2019, 20, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whoolery, C.W.; Walker, A.K.; Richardson, D.R.; Lucero, M.J.; Reynolds, R.P.; Beddow, D.H.; Clark, K.L.; Shih, H.-Y.; LeBlanc, J.A.; Cole, M.G.; et al. Whole-Body Exposure to 28 Si-Radiation Dose-Dependently Disrupts Dentate Gyrus Neurogenesis and Proliferation in the Short Term and New Neuron Survival and Contextual Fear Conditioning in the Long Term. Radiat. Res. 2017, 188, 532–551. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Jewell, J.S.; Duncan, V.D.; Hadley, M.M.; Macadat, E.; Musto, A.E.; Tessa, C. La Impaired Attentional Set-Shifting Performance after Exposure to 5 cGy of 600 MeV/n 28Si Particles. Radiat. Res. 2018, 189, 273–282. [Google Scholar] [CrossRef]
- Britten, R.A.; Davis, L.K.; Johnson, A.M.; Keeney, S.; Siegel, A.; Sanford, L.D.; Singletary, S.J.; Lonart, G. Low (20 cGy) doses of 1 GeV/u (56)Fe--particle radiation lead to a persistent reduction in the spatial learning ability of rats. Radiat. Res. 2012, 177, 146–151. [Google Scholar] [CrossRef]
- Cherry, J.D.; Liu, B.; Frost, J.L.; Lemere, C.A.; Williams, J.P.; Olschowka, J.A.; O’Banion, M.K. Galactic Cosmic Radiation Leads to Cognitive Impairment and Increased Aβ Plaque Accumulation in a Mouse Model of Alzheimer’s Disease. PLoS ONE 2012, 7, e53275. [Google Scholar] [CrossRef] [PubMed]
- Wyrobek, A.J.; Britten, R.A. Individual variations in dose response for spatial memory learning among outbred wistar rats exposed from 5 to 20 cGy of (56) Fe particles. Environ. Mol. Mutagen. 2016, 57, 331–340. [Google Scholar] [CrossRef]
- Kiffer, F.; Alexander, T.; Anderson, J.; Groves, T.; McElroy, T.; Wang, J.; Sridharan, V.; Bauer, M.; Boerma, M.; Allen, A. Late Effects of 1H + 16O on Short-Term and Object Memory, Hippocampal Dendritic Morphology and Mutagenesis. Front. Behav. Neurosci. 2020, 14, 1–15. [Google Scholar] [CrossRef]
- Krukowski, K.; Grue, K.; Frias, E.S.; Pietrykowski, J.; Jones, T.; Nelson, G.A.; Rosi, S. Female mice are protected from space radiation-induced maladaptive responses. Brain. Behav. Immun. 2018, 74, 106–120. [Google Scholar] [CrossRef]
- Kiffer, F.; Carr, H.; Groves, T.; Anderson, J.E.; Alexander, T.; Wang, J.; Seawright, J.W.; Sridharan, V.; Carter, G.; Boerma, M.; et al. Effects of 1 H + 16 O Charged Particle Irradiation on Short-Term Memory and Hippocampal Physiology in a Murine Model. Radiat. Res. 2018, 189, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Yamazaki, J.; Torres, E.R.S.; Kirchoff, N.; Stagaman, K.; Sharpton, T.; Turker, M.S.; Kronenberg, A. Combined effects of three high-energy charged particle beams important for space flight on brain, behavioral and cognitive endpoints in B6D2F1 female and Male mice. Front. Physiol. 2019, 10, 179. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, I.V.; Schneider, C.J.; Bezaire, M.J.; Soltesz, I.; Vlkolinsky, R.; Nelson, G.A. Proton Radiation Alters Intrinsic and Synaptic Properties of CA1 Pyramidal Neurons of the Mouse Hippocampus. Radiat. Res. 2015, 183, 208–218. [Google Scholar] [CrossRef]
- Lee, S.-H.; Dudok, B.; Parihar, V.K.; Jung, K.M.; Zöldi, M.; Kang, Y.-J.; Maroso, M.; Alexander, A.L.; Nelson, G.A.; Piomelli, D.; et al. Neurophysiology of space travel: Energetic solar particles cause cell type-specific plasticity of neurotransmission. Brain Struct. Funct. 2017, 222, 2345–2357. [Google Scholar] [CrossRef] [Green Version]
- Klein, P.M.; Parihar, V.K.; Szabo, G.G.; Zöldi, M.; Angulo, M.C.; Allen, B.D.; Amin, A.N.; Nguyen, Q.-A.; Katona, I.; Baulch, J.E.; et al. Detrimental impacts of mixed-ion radiation on nervous system function. Neurobiol. Dis. 2021, 151, 105252. [Google Scholar] [CrossRef]
- Slaba, T.C.; Blattnig, S.R.; Clowdsley, M.S. Variation in lunar neutron dose estimates. Radiat. Res. 2011, 176, 827–841. [Google Scholar] [CrossRef]
- Norbury, J.W.; Slaba, T.C. Space radiation accelerator experiments—The role of neutrons and light ions. Life Sci. Space Res. 2014, 3, 90–94. [Google Scholar] [CrossRef]
- Khaksarighiri, S.; Guo, J.; Wimmer-Schweingruber, R.; Narici, L.; Lohf, H. Calculation of dose distribution in a realistic brain structure and the indication of space radiation influence on human brains. Life Sci. Space Res. 2020, 27, 33–48. [Google Scholar] [CrossRef]
- Stricklin, D.; VanHorne-Sealy, J.; Rios, C.I.; Scott Carnell, L.A.; Taliaferro, L.P. Neutron Radiobiology and Dosimetry. Radiat. Res. 2021, 195, 480–496. [Google Scholar] [CrossRef]
- Alard, J.P.; Bodez, V.; Tchirkov, A.; Nénot, M.L.; Arnold, J.; Crespin, S.; Rapp, M.; Verrelle, P.; Dionet, C. Simulation of neutron interactions at the single-cell level. Radiat. Res. 2002, 158, 650–656. [Google Scholar] [CrossRef]
- Norbury, J.W.; Slaba, T.C.; Aghara, S.; Badavi, F.F.; Blattnig, S.R.; Clowdsley, M.S.; Heilbronn, L.H.; Lee, K.; Maung, K.M.; Mertens, C.J.; et al. Advances in space radiation physics and transport at NASA. Life Sci. Space Res. 2019, 22, 98–124. [Google Scholar] [CrossRef]
- Zeitlin, C.; Hassler, D.M.; Cucinotta, F.A.; Ehresmann, B.; Wimmer-Schweingruber, R.F.; Brinza, D.E.; Kang, S.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Measurements of energetic particle radiation in transit to Mars on the Mars Science Laboratory. Science 2013, 340, 1080–1084. [Google Scholar] [CrossRef] [Green Version]
- Borak, T.B.; Heilbronn, L.H.; Krumland, N.; Weil, M.M. Design and dosimetry of a facility to study health effects following exposures to fission neutrons at low dose rates for long durations. Int. J. Radiat. Biol. 2019, 1063–1076. [Google Scholar] [CrossRef]
- Acharya, M.M.; Baulch, J.E.; Klein, P.M.; Baddour, A.A.D.; Apodaca, L.A.; Kramár, E.A.; Alikhani, L.; Garcia, C.; Angulo, M.C.; Batra, R.S.; et al. New Concerns for Neurocognitive Function during Deep Space Exposures to Chronic, Low Dose-Rate, Neutron Radiation. eNeuro 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Parihar, V.K.; Pasha, J.; Tran, K.K.; Craver, B.M.; Acharya, M.M.; Limoli, C.L. Persistent changes in neuronal structure and synaptic plasticity caused by proton irradiation. Brain Struct. Funct. 2015, 220, 1161–1171. [Google Scholar] [CrossRef] [Green Version]
- Aarts, E.; Verhage, M.; Veenvliet, J.V.; Dolan, C.V.; van der Sluis, S. A solution to dependency: Using multilevel analysis to accommodate nested data. Nat. Neurosci. 2014, 17, 491–496. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.; Tumkaya, T.; Aryal, S.; Choi, H.; Claridge-Chang, A. Moving beyond P values: Data analysis with estimation graphics. Nat. Methods 2019, 16, 565–566. [Google Scholar] [CrossRef]
- Calin-Jageman, R.J.; Cumming, G. Estimation for better inference in neuroscience. eNeuro 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Turrigiano, G.G. Too many cooks? Intrinsic and synaptic homeostatic mechanisms in cortical circuit refinement. Annu. Rev. Neurosci. 2011, 34, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Desai, N.S.; Rutherford, L.C.; Turrigiano, G.G. Plasticity in the intrinsic excitability of cortical pyramidal neurons. Nat. Neurosci. 1999, 2, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Echegoyen, J.; Neu, A.; Graber, K.D.; Soltesz, I. Homeostatic plasticity studies using in vivo hippocampal activity-blockade: Synaptic scaling, intrinsic plasticity and age-dependence. PLoS ONE 2007, 2, e700. [Google Scholar] [CrossRef] [Green Version]
- Turrigiano, G.G.; Leslie, K.R.; Desai, N.S.; Rutherford, L.C.; Nelson, S.B. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature 1998, 391, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.H.; Park, J.M.; Park, S.; Xiao, B.; Dehoff, M.H.; Kim, S.; Hayashi, T.; Schwarz, M.K.; Huganir, R.L.; Seeburg, P.H.; et al. Homeostatic Scaling Requires Group I mGluR Activation Mediated by Homer1a. Neuron 2010, 68, 1128–1142. [Google Scholar] [CrossRef] [Green Version]
- Gainey, M.A.; Hurvitz-Wolff, J.R.; Lambo, M.E.; Turrigiano, G.G. Synaptic scaling requires the GluR2 subunit of the AMPA receptor. J. Neurosci. 2009, 29, 6479–6489. [Google Scholar] [CrossRef] [Green Version]
- Mellor, J.; Nicoll, R.A.; Schmitz, D. Mediation of hippocampal mossy fiber long-term potentiation by presynaptic Ih channels. Science 2002, 295, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kang, N.; Jiang, L.; Nedergaard, M.; Kang, J. Activity-dependent long-term potentiation of intrinsic excitability in hippocampal CA1 pyramidal neurons. J. Neurosci. 2005, 25, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, N.L.; Wu, C.P.; Duan, S.; Poo, M.M. Bidirectional changes in spatial dendritic integration accompanying long-term synaptic modifications. Neuron 2003, 37, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Styr, B.; Slutsky, I. Imbalance between firing homeostasis and synaptic plasticity drives early-phase Alzheimer’s disease. Nat. Neurosci. 2018, 21, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Marder, E.; Goaillard, J.M. Variability, compensation and homeostasis in neuron and network function. Nat. Rev. Neurosci. 2006, 7, 563–574. [Google Scholar] [CrossRef]
- Schwartzkroin, P.A.; Wester, K. Long-lasting facilitation of a synaptic potential following tetanization in the in vitro hippocampal slice. Brain Res. 1975, 89, 107–119. [Google Scholar] [CrossRef]
- Lynch, M.A. Long-Term Potentiation and Memory. Physiol. Rev. 2004, 84, 87–136. [Google Scholar] [CrossRef]
- Sarnyai, Z.; Sibille, E.L.; Pavlides, C.; Fenster, R.J.; McEwen, B.S.; Tóth, M. Impaired hippocampal-dependent learning and functional abnormalities in the hippocampus in mice lacking serotonin1A receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 14731–14736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katafuchi, T.; Li, A.-J.; Hirota, S.; Kitamura, Y.; Hori, T. Impairment of spatial learning and hippocampal synaptic potentiation in c-kit mutant rats. Learn. Mem. 2000, 7, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finlay, J.M.; Dunham, G.A.; Isherwood, A.M.; Newton, C.J.; Nguyen, T.V.; Reppar, P.C.; Snitkovski, I.; Paschall, S.A.; Greene, R.W. Effects of prefrontal cortex and hippocampal NMDA NR1-subunit deletion on complex cognitive and social behaviors. Brain Res. 2015, 1600, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Ko, J. Neuroanatomical Substrates of Rodent Social Behavior: The Medial Prefrontal Cortex and Its Projection Patterns. Front. Neural Circuits 2017, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Winters, B.D.; Forwood, S.E.; Cowell, R.A.; Saksida, L.M.; Bussey, T.J. Double dissociation between the effects of peri-postrhinal cortex and hippocampal lesions on tests of object recognition and spatial memory: Heterogeneity of function within the temporal lobe. J. Neurosci. 2004, 24, 5901–5908. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.E.; Zola, S.M.; Squire, L.R. Impaired recognition memory rats after damage to the hippocampus. J. Neurosci. 2000, 20, 8853–8860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburton, E.C.; Brown, M.W. Neural circuitry for rat recognition memory. Behav. Brain Res. 2015, 285, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Barker, G.R.I.; Warburton, E.C. When is the hippocampus involved in recognition memory? J. Neurosci. 2011, 31, 10721–10731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, L.; Brown, M.W.; Aggleton, J.P.; Warburton, E.C. The medial dorsal thalamic nucleus and the medial prefrontal cortex of the rat function together to support associative recognition and recency but not item recognition. Learn. Mem. 2013, 20, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, O.Y.; de Souza Silva, M.A.; Yang, Y.-M.; Huston, J.P. The medial prefrontal cortex—hippocampus circuit that integrates information of object, place and time to construct episodic memory in rodents: Behavioral, anatomical and neurochemical properties. Neurosci. Biobehav. Rev. 2020, 113, 373–407. [Google Scholar] [CrossRef]
- Jo, Y.S.; Lee, I. Disconnection of the hippocampal-perirhinal cortical circuits severely disrupts object-place paired associative memory. J. Neurosci. 2010, 30, 9850–9858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirk, G.J.; Mueller, D. Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology 2008, 33, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Knapska, E.; Orsini, C.A.; Rabinak, C.A.; Zimmerman, J.M.; Maren, S. Fear extinction in rodents. Curr. Protoc. Neurosci. 2009, 47, 8–23. [Google Scholar] [CrossRef]
- VanElzakker, M.B.; Kathryn Dahlgren, M.; Caroline Davis, F.; Dubois, S.; Shin, L.M. From Pavlov to PTSD: The extinction of conditioned fear in rodents, humans, and anxiety disorders. Neurobiol. Learn. Mem. 2014, 113, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Gunaydin, L.A.; Grosenick, L.; Finkelstein, J.C.; Kauvar, I.V.; Fenno, L.E.; Adhikari, A.; Lammel, S.; Mirzabekov, J.J.; Airan, R.D.; Zalocusky, K.A.; et al. Natural Neural Projection Dynamics Underlying Social Behavior. Cell 2014, 157, 1535–1551. [Google Scholar] [CrossRef] [Green Version]
- Parihar, V.K.; Allen, B.D.; Caressi, C.; Kwok, S.; Chu, E.; Tran, K.K.; Chmielewski, N.N.; Giedzinski, E.; Acharya, M.M.; Britten, R.A.; et al. Cosmic radiation exposure and persistent cognitive dysfunction. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bourin, M.; Hascoët, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Otake, M.; Schull, W.J. Review: Radiation-related brain damage and growth retardation among the prenatally exposed atomic bomb survivors. Int. J. Radiat. Biol. 1998, 74, 159–171. [Google Scholar] [CrossRef]
- Takahashi, I.; Ohishi, W.; Mettler, F.A.; Ozasa, K.; Jacob, P.; Ban, N.; Lipshultz, S.E.; Stewart, F.A.; Nabika, T.; Niwa, Y.; et al. A report from the 2013 international workshop: Radiation and cardiovascular disease, Hiroshima, Japan. J. Radiol. Prot. 2013, 33, 869–880. [Google Scholar] [CrossRef]
- Ozasa, K. Epidemiological research on radiation-induced cancer in atomic bomb survivors. J. Radiat. Res. 2016, 57, i112–i117. [Google Scholar] [CrossRef] [PubMed]
- Mineyeva, O.A.; Barykina, N.V.; Bezriadnov, D.V.; Latushkin, S.T.; Ryazanov, A.I.; Unezhev, V.N.; Shuvaev, S.A.; Usova, S.V.; Lazutkin, A.A. Suppressed neurogenesis without cognitive deficits: Effects of fast neutron irradiation in mice. Neuroreport 2019, 30, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Ohmachi, Y.; Nakata, Y.; Hiraoka, T.; Hamano, T.; Fushiki, S.; Ogiu, T. Dose-response and large relative biological effectiveness of fast neutrons with regard to mouse fetal cerebral neuron apoptosis. J. Radiat. Res. 2006, 47, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-E.; Shin, M.-S.; Sung, Y.-H.; Kim, H.; Kang, J.-O.; Lim, B.-V.; Jee, Y.-S.; Kim, C.-J. Effects of Neutron Radiation on Apoptotic Cell Death and Cell Proliferation in Dentate Gyrus of Rats. Exp. Neurobiol. 2007, 16, 19–26. [Google Scholar]
- Yang, M.; Kim, J.-S.; Song, M.-S.; Kim, J.-C.; Shin, T.; Lee, S.-S.; Kim, S.-H.; Moon, C. Dose-response and relative biological effectiveness of fast neutrons: Induction of apoptosis and inhibition of neurogenesis in the hippocampus of adult mice. Int. J. Radiat. Biol. 2010, 86, 476–485. [Google Scholar] [CrossRef]
- Dyhrfjeld-Johnsen, J.; Morgan, R.J.; Földy, C.; Soltesz, I. Upregulated H-current in hyperexcitable CA1 dendrites after febrile seizures. Front. Cell. Neurosci. 2008, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Booker, S.A.; Simões de Oliveira, L.; Anstey, N.J.; Kozic, Z.; Dando, O.R.; Jackson, A.D.; Baxter, P.S.; Isom, L.L.; Sherman, D.L.; Hardingham, G.E.; et al. Input-Output Relationship of CA1 Pyramidal Neurons Reveals Intact Homeostatic Mechanisms in a Mouse Model of Fragile X Syndrome. Cell Rep. 2020, 32, 107988. [Google Scholar] [CrossRef]
- Dutta, S.M.; Hadley, M.M.; Peterman, S.; Jewell, J.S.; Duncan, V.D.; Britten, R.A. Quantitative Proteomic Analysis of the Hippocampus of Rats with GCR-Induced Spatial Memory Impairment. Radiat. Res. 2018, 189, 136–145. [Google Scholar] [CrossRef]
- Bayarchimeg, L.; Batmunkh, M.; Bugay, A.N.; Lkhagva, O. Evaluation of Radiation-Induced Damage in Membrane Ion Channels and Synaptic Receptors. Phys. Part. Nucl. Lett. 2019, 16, 54–62. [Google Scholar] [CrossRef]
- Saw, G.; Tang, F.R. Epigenetic regulation of the hippocampus, with special reference to radiation exposure. Int. J. Mol. Sci. 2020, 21, 9514. [Google Scholar] [CrossRef]
- Krishnan, B.; Natarajan, C.; Bourne, K.Z.; Alikhani, L.; Wang, J.; Sowa, A.; Groen, K.; Perry, B.; Dickstein, D.L.; Baulch, J.E.; et al. Chronic low dose neutron exposure results in altered neurotransmission properties of the hippocampus-prefrontal cortex axis in both mice and rats. Int. J. Mol. Sci. 2021, 22, 3668. [Google Scholar] [CrossRef]
- Allen, B.D.; Syage, A.R.; Maroso, M.; Baddour, A.A.D.; Luong, V.; Minasyan, H.; Giedzinski, E.; West, B.L.; Soltesz, I.; Limoli, C.L.; et al. Mitigation of helium irradiation-induced brain injury by microglia depletion. J. Neuroinflamm. 2020, 17, 159. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Merchán-Pérez, A.; Rial Verde, E.M.; Rodríguez, J.-R.; Defelipe, J.; Yuste, R. Ultrastructural, Molecular and Functional Mapping of GABAergic Synapses on Dendritic Spines and Shafts of Neocortical Pyramidal Neurons. Cereb. Cortex 2019, 29, 2771–2781. [Google Scholar] [CrossRef] [PubMed]
- Hersch, S.M.; White, E.L. Quantification of synapses formed with apical dendrites of golgi-impregnated pyramidal cells: Variability in thalamocortical inputs, but consistency in the ratios of asymmetrical to symmetrical synapses. Neuroscience 1981, 6, 1043–1051. [Google Scholar] [CrossRef]
- Buzsáki, G. Hippocampal sharp wave-ripple: A cognitive biomarker for episodic memory and planning. Hippocampus 2015, 25, 1073–1188. [Google Scholar] [CrossRef] [PubMed]
- Keiser, A.A.; Kramár, E.A.; Dong, T.; Shanur, S.; Pirodan, M.; Ru, N.; Acharya, M.M.; Baulch, J.E.; Limoli, C.L.; Wood, M.A. Systemic HDAC3 inhibition ameliorates impairments in synaptic plasticity caused by simulated galactic cosmic radiation exposure in male mice. Neurobiol. Learn. Mem. 2021, 178, 107367. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Kehl, S.J.; McLennan, H. Excitatory amino acids in synaptic transmission in the Schaffer collateral-commissural pathway of the rat hippocampus. J. Physiol. 1983, 334, 33–46. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Singer, W. Excitatory amino acid receptors and synaptic plasticity. Trends Pharmacol. Sci. 1990, 11, 290–296. [Google Scholar] [CrossRef]
- Kessels, H.W.; Malinow, R. Synaptic AMPA receptor plasticity and behavior. Neuron 2009, 61, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Alvarez, G.; Shetty, M.S.; Lu, B.; Yap, K.A.F.; Oh-Hora, M.; Sajikumar, S.; Bichler, Z.; Fivaz, M. Impaired spatial memory and enhanced long-term potentiation in mice with forebrain-specific ablation of the Stim genes. Front. Behav. Neurosci. 2015, 9, 1–12. [Google Scholar] [CrossRef]
- Kiffer, F.; Boerma, M.; Allen, A. Behavioral effects of space radiation: A comprehensive review of animal studies. Life Sci. Space Res. 2019, 21, 1–21. [Google Scholar] [CrossRef]
- Raber, J.; Marzulla, T.; Stewart, B.; Kronenberg, A.; Turker, M.S. 28Silicon Irradiation Impairs Contextual Fear Memory in B6D2F1 Mice. Radiat. Res. 2015, 183, 708–712. [Google Scholar] [CrossRef]
- Xu, Y.; Randers-Pehrson, G.; Turner, H.C.; Marino, S.A.; Geard, C.R.; Brenner, D.J.; Garty, G. Accelerator-Based Biological Irradiation Facility Simulating Neutron Exposure from an Improvised Nuclear Device. Radiat. Res. 2015, 184, 404–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laiakis, E.C.; Canadell, M.P.; Grilj, V.; Harken, A.D.; Garty, G.Y.; Astarita, G.; Brenner, D.J.; Smilenov, L.; Fornace, A.J. Serum lipidomic analysis from mixed neutron/X-ray radiation fields reveals a hyperlipidemic and pro-inflammatory phenotype. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Randers-Pehrson, G.; Marino, S.A.; Bigelow, A.W.; Akselrod, M.S.; Sykora, J.G.; Brenner, D.J. An accelerator-based neutron microbeam system for studies of radiation effects. Radiat. Prot. Dosim. 2011, 145, 373–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Randers-Pehrson, G.; Marino, S.A.; Garty, G.; Harken, A.; Brenner, D.J. Broad energy range neutron spectroscopy using a liquid scintillator and a proportional counter: Application to a neutron spectrum similar to that from an improvised nuclear device. Nucl. Instrum. Methods Phys. Res. A 2015, 794, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Rossi, H.H.; Bateman, J.L.; Bond, V.P.; Goodman, L.J.; Stickley, E.E. The dependence of RBE on the energy of fast neutrons. 1. Physical design and measurement of absorbed dose. Radiat. Res. 1960, 13, 503–520. [Google Scholar] [CrossRef]
- Ting, J.T.; Daigle, T.L.; Chen, Q.; Feng, G. Acute brain slice methods for adult and aging animals: Application of targeted patch clamp analysis and optogenetics. In Patch-Clamp Methods and Protocols; Martina, M., Taverna, S., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2014; Volume 1183, pp. 221–242. ISBN 978-1-4939-1095-3. [Google Scholar]
- Vogel-Ciernia, A.; Matheos, D.P.; Barrett, R.M.; Kramár, E.A.; Azzawi, S.; Chen, Y.; Magnan, C.N.; Zeller, M.; Sylvain, A.; Haettig, J.; et al. The neuron-specific chromatin regulatory subunit BAF53b is necessary for synaptic plasticity and memory. Nat. Neurosci. 2013, 16, 552–561. [Google Scholar] [CrossRef]
- Barker, G.R.I.; Bird, F.; Alexander, V.; Warburton, E.C. Recognition memory for objects, place, and temporal order: A disconnection analysis of the role of the medial prefrontal cortex and perirhinal cortex. J. Neurosci. 2007, 27, 2948–2957. [Google Scholar] [CrossRef] [Green Version]
- Winslow, J.T. Mouse Social Recognition and Preference. Curr. Protoc. Neurosci. 2004, 22, 8.16.1–8.16.16. [Google Scholar] [CrossRef]
- Sato, A.; Mizuguchi, M.; Ikeda, K. Social interaction test: A sensitive method for examining autism-related behavioral deficits. Protoc. Exch. 2013. [Google Scholar] [CrossRef]
- Petit-Demouliere, B.; Chenu, F.; Bourin, M. Forced swimming test in mice: A review of antidepressant activity. Psychopharmacology 2005, 177, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Cain, C.K.; Blouin, A.M.; Barad, M. Temporally Massed CS Presentations Generate More Fear Extinction than Spaced Presentations. J. Exp. Psychol. Anim. Behav. Process. 2003, 29, 323–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klein, P.M.; Alaghband, Y.; Doan, N.-L.; Ru, N.; Drayson, O.G.G.; Baulch, J.E.; Kramár, E.A.; Wood, M.A.; Soltesz, I.; Limoli, C.L. Acute, Low-Dose Neutron Exposures Adversely Impact Central Nervous System Function. Int. J. Mol. Sci. 2021, 22, 9020. https://doi.org/10.3390/ijms22169020
Klein PM, Alaghband Y, Doan N-L, Ru N, Drayson OGG, Baulch JE, Kramár EA, Wood MA, Soltesz I, Limoli CL. Acute, Low-Dose Neutron Exposures Adversely Impact Central Nervous System Function. International Journal of Molecular Sciences. 2021; 22(16):9020. https://doi.org/10.3390/ijms22169020
Chicago/Turabian StyleKlein, Peter M., Yasaman Alaghband, Ngoc-Lien Doan, Ning Ru, Olivia G. G. Drayson, Janet E. Baulch, Enikö A. Kramár, Marcelo A. Wood, Ivan Soltesz, and Charles L. Limoli. 2021. "Acute, Low-Dose Neutron Exposures Adversely Impact Central Nervous System Function" International Journal of Molecular Sciences 22, no. 16: 9020. https://doi.org/10.3390/ijms22169020
APA StyleKlein, P. M., Alaghband, Y., Doan, N.-L., Ru, N., Drayson, O. G. G., Baulch, J. E., Kramár, E. A., Wood, M. A., Soltesz, I., & Limoli, C. L. (2021). Acute, Low-Dose Neutron Exposures Adversely Impact Central Nervous System Function. International Journal of Molecular Sciences, 22(16), 9020. https://doi.org/10.3390/ijms22169020