

Umbelliferone Ameliorates Benign Prostatic Hyperplasia by Inhibiting Cell Proliferation and G1/S Phase Cell Cycle Progression through Regulation of STAT3/E2F1 Axis


Abstract
:1. Introduction
2. Results
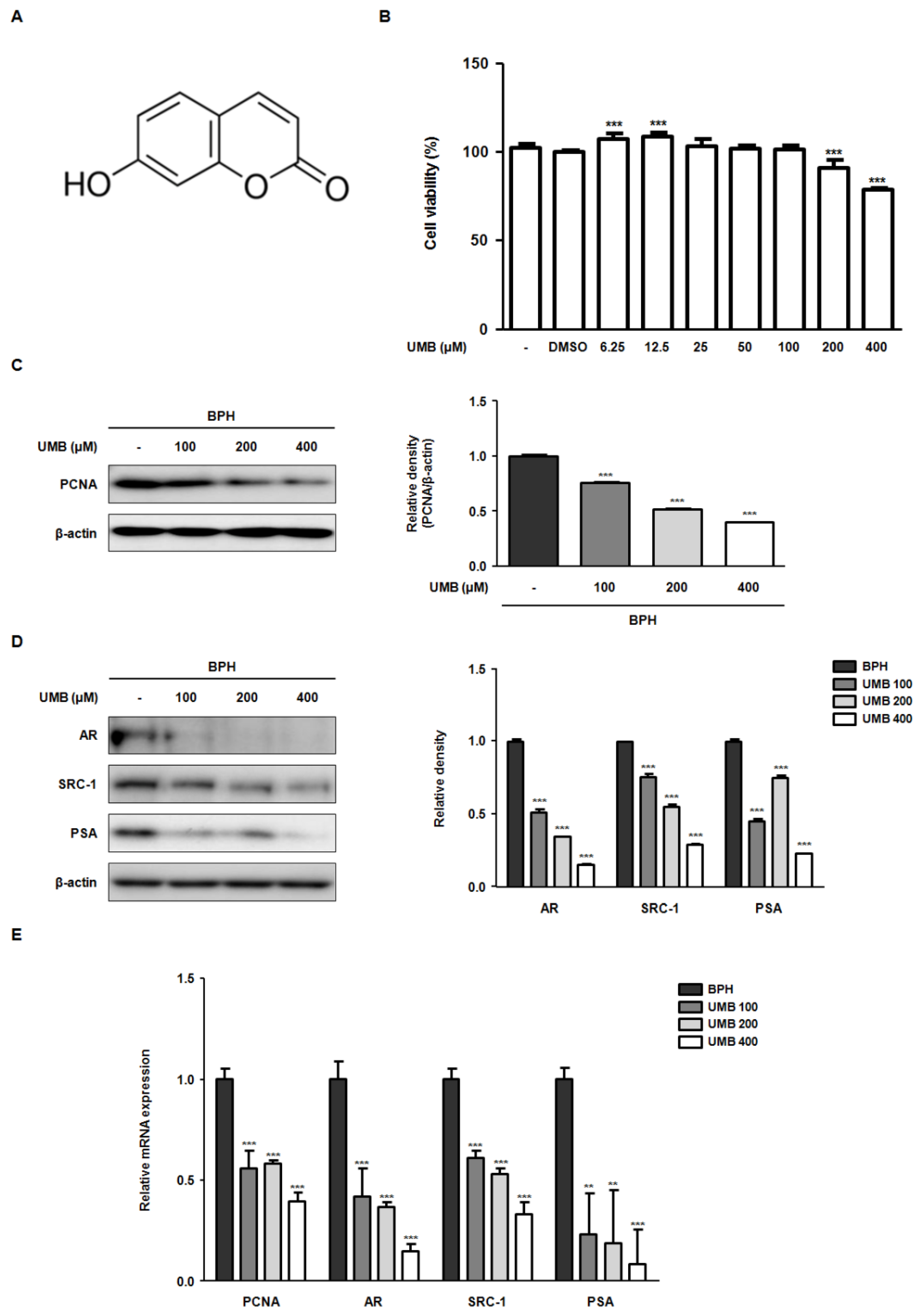
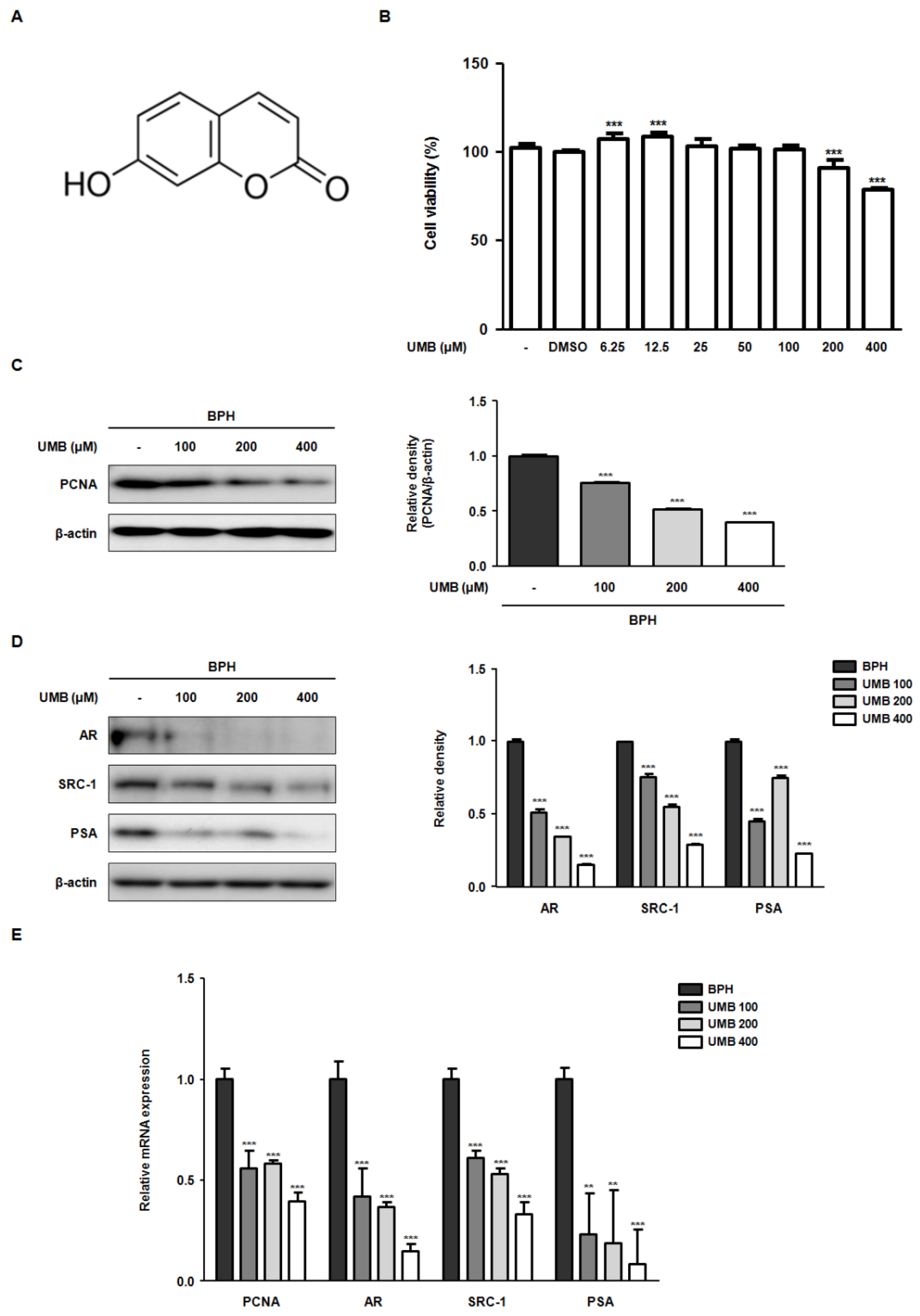
2.1. Effect of UMB on the Blockade of Androgen/AR Signaling in BPH-1 Cells
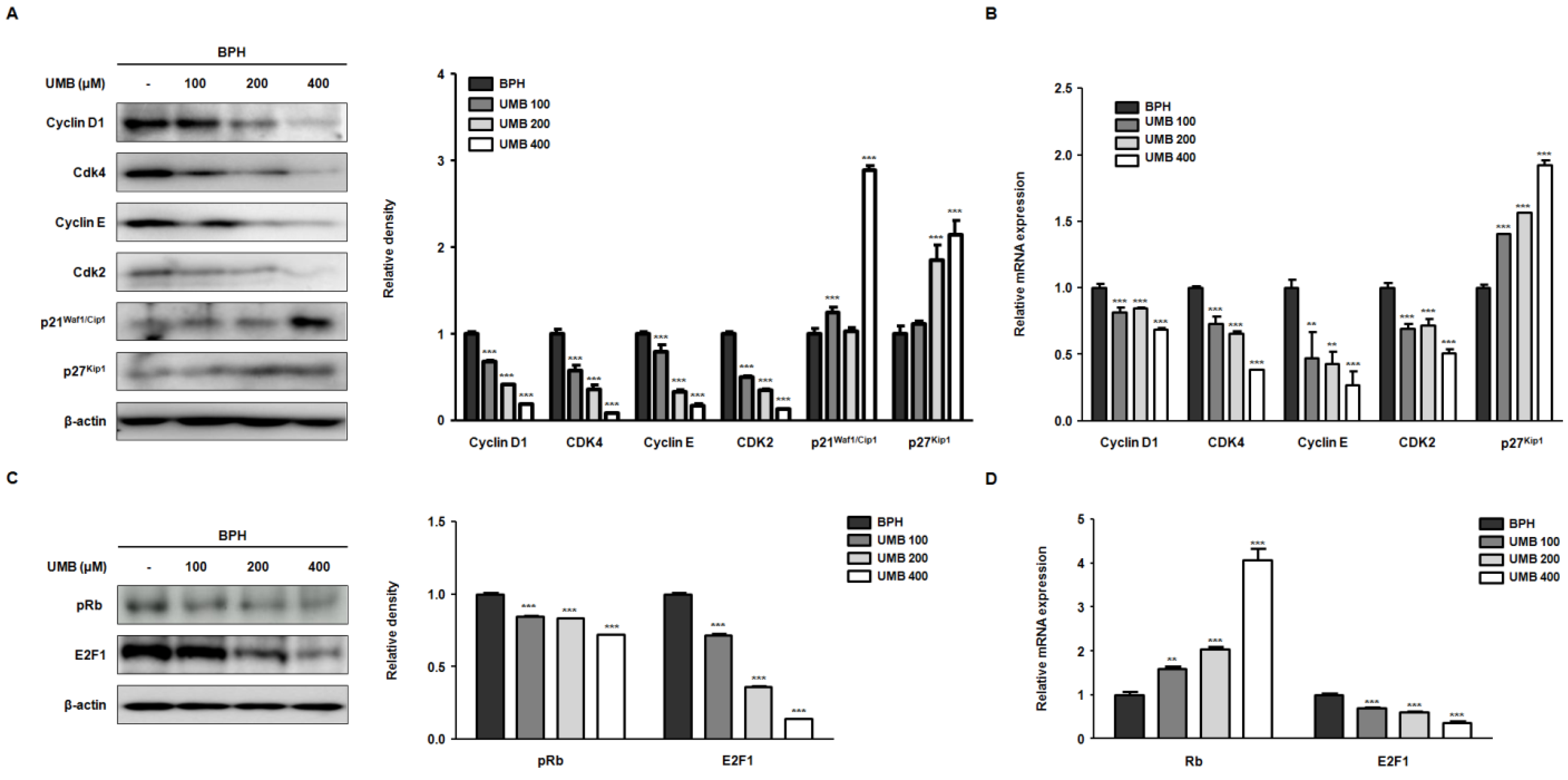
2.2. Effect of UMB on G1/S Phase Cell Cycle Arrest in BPH-1 Cells
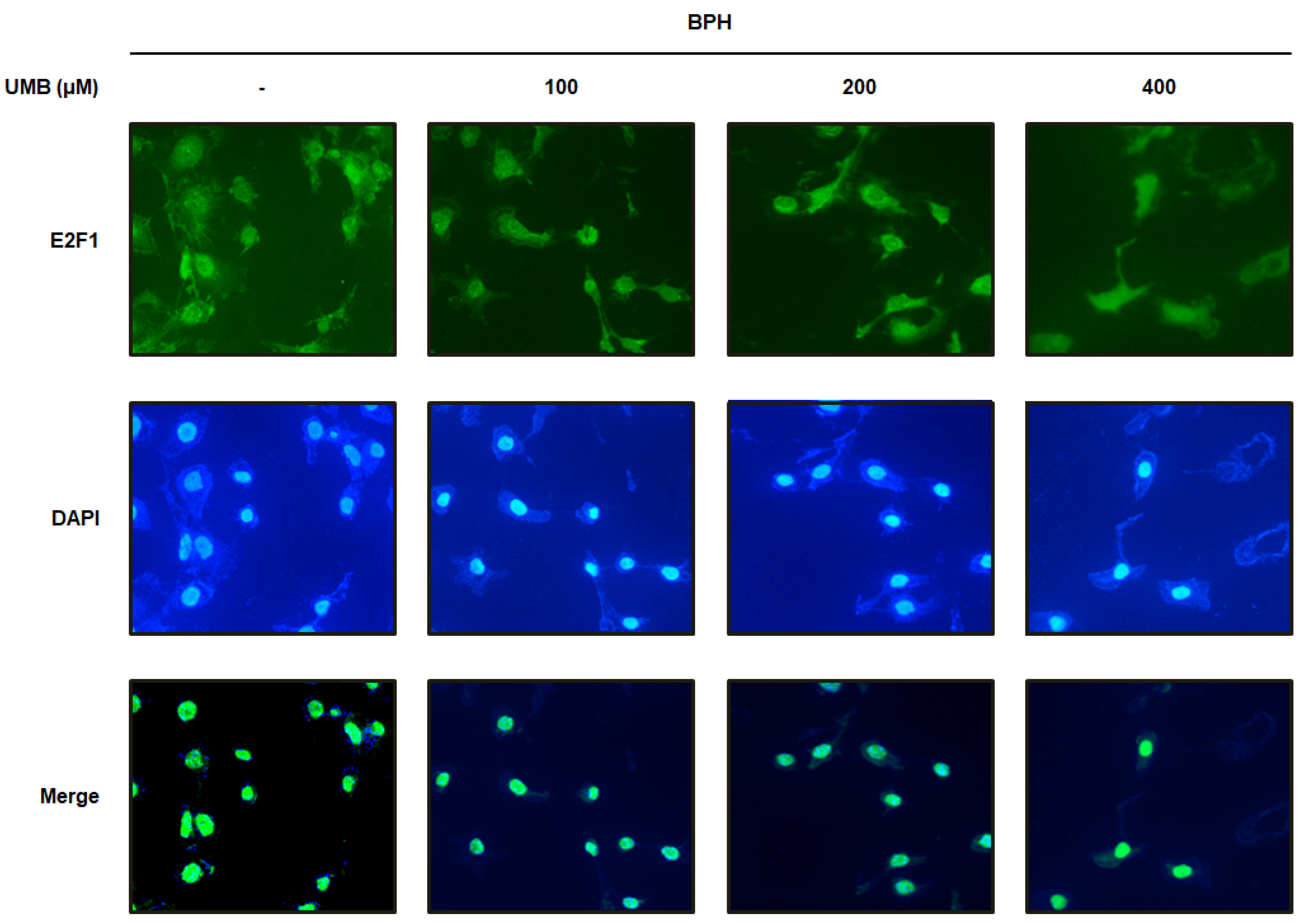
2.3. Effect of UMB Treatment on E2F1 Expression in BPH-1 Cells
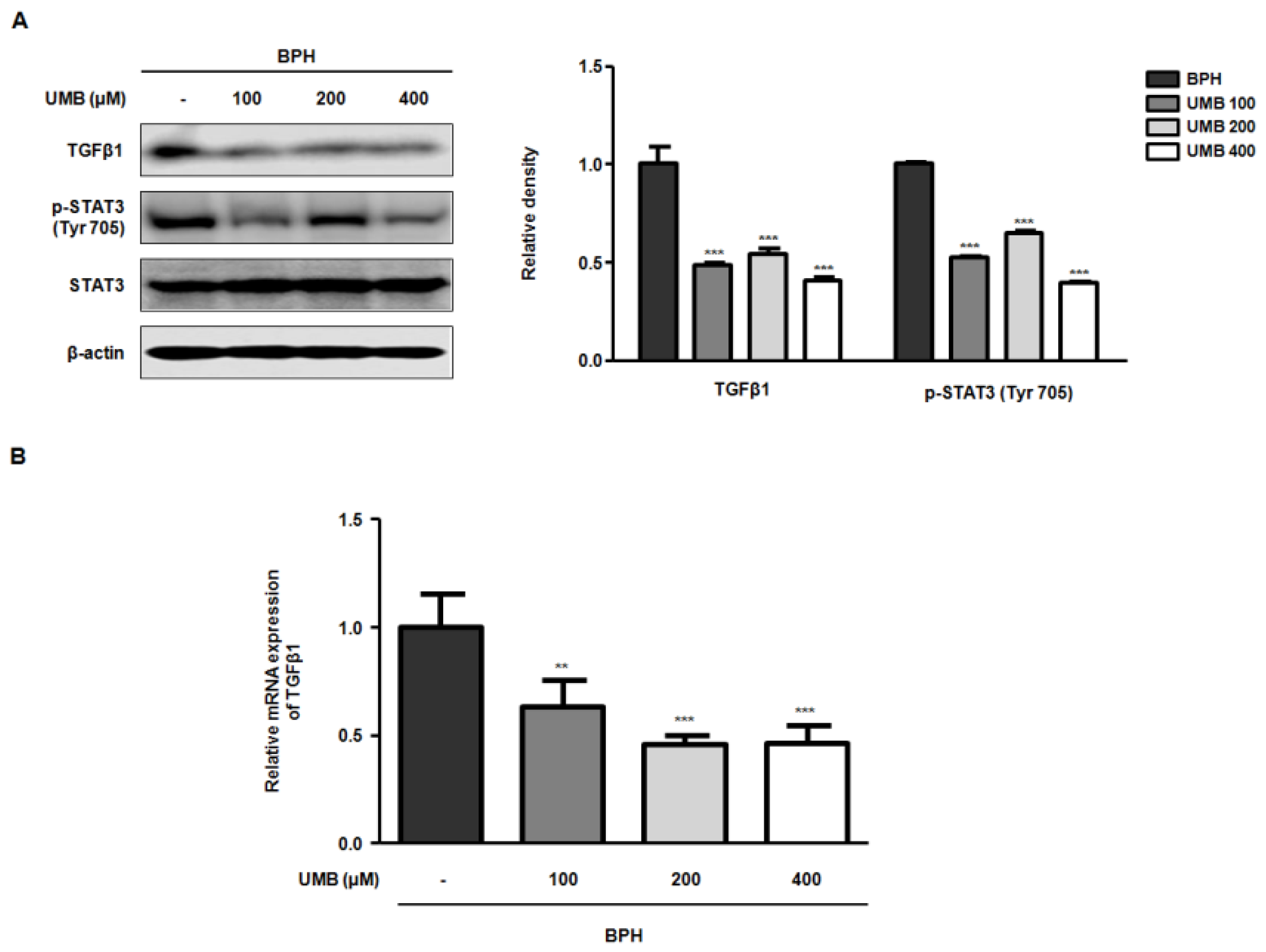
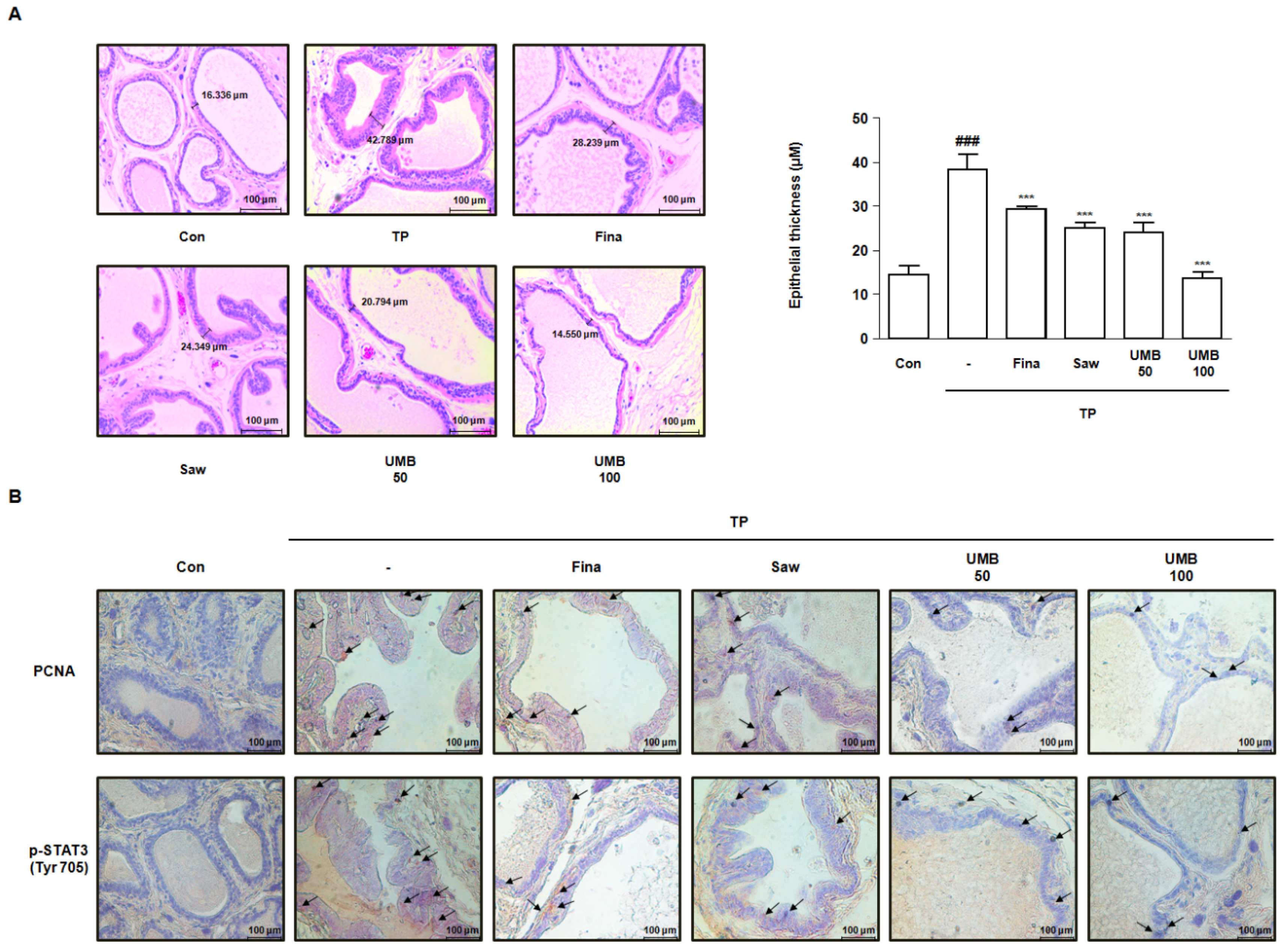
2.4. Effect of UMB Treatment on TGFβ1 and p-STAT3 (Tyr 705) Expression in BPH-1 Cells
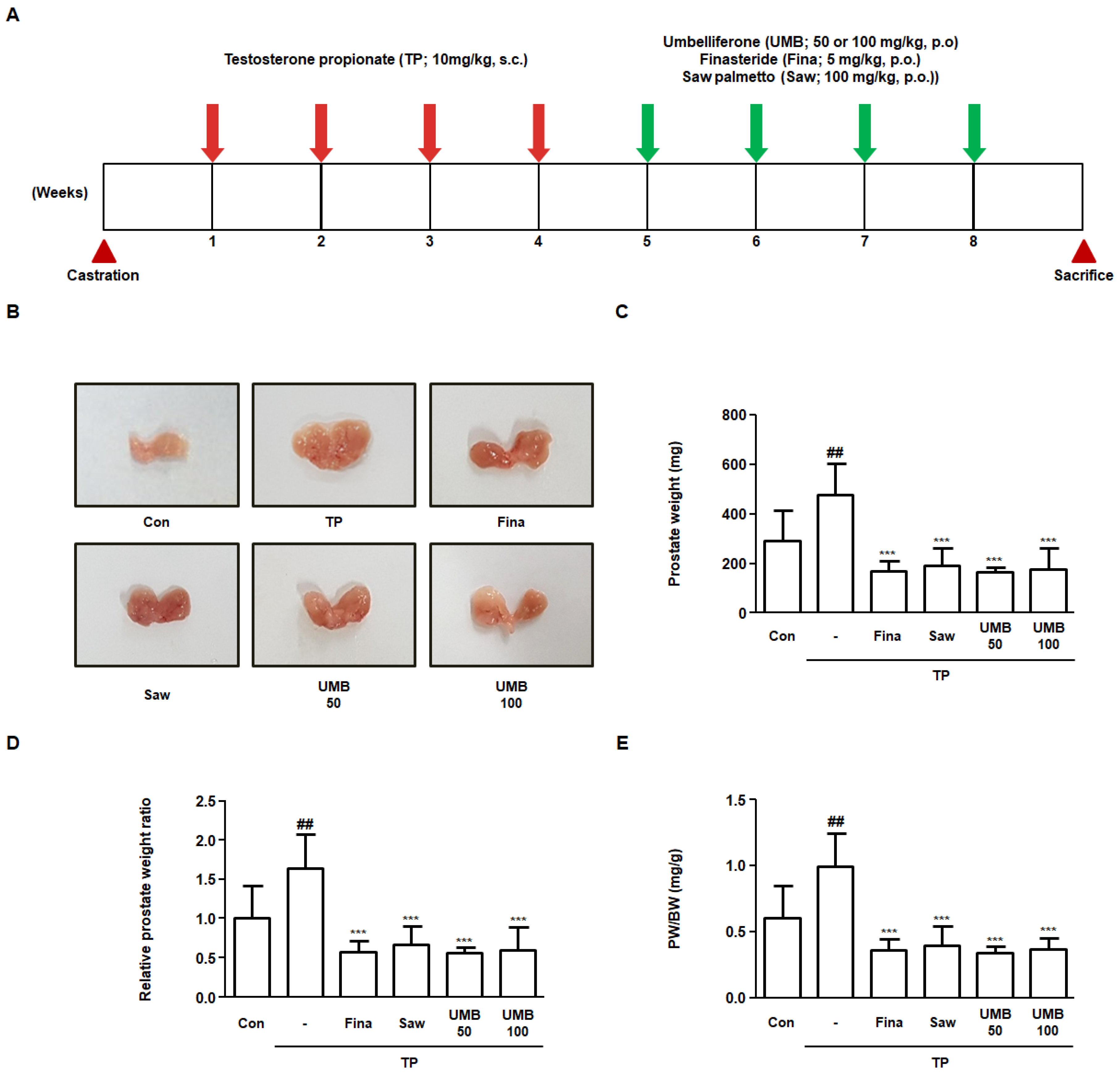
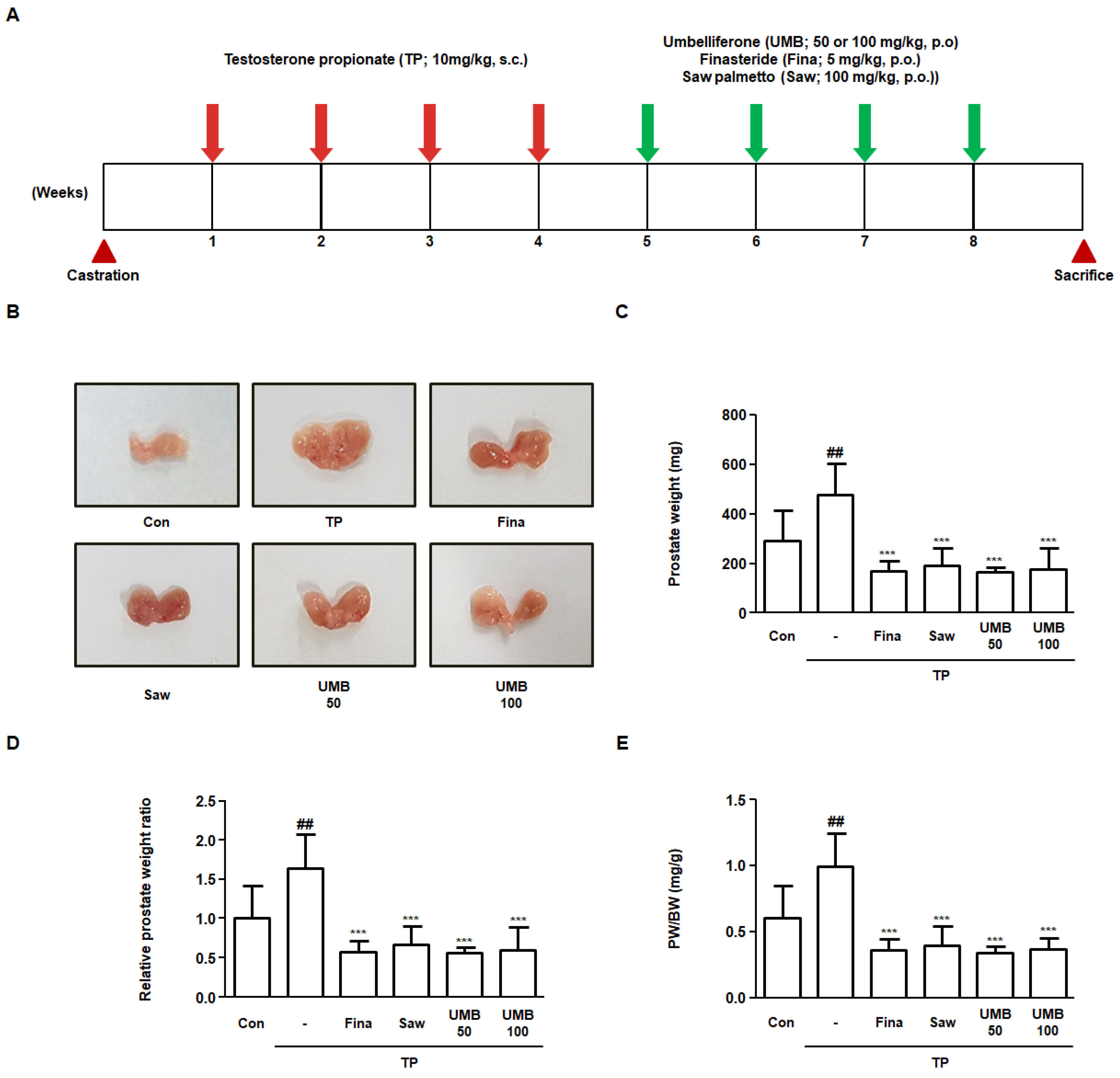
2.5. Effect of UMB on Prostate Enlargement in TP-Induced Rats
2.6. Effect of UMB on Histological Alterations in TP-Induced Rats
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of UMB and Cell Culture
4.3. Cell Viability Assays
4.4. Western Blot Analysis
4.5. Isolation of Total RNA and Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
4.6. Immunofluorescence Staining
4.7. Animals
4.8. Hematoxylin and Eosin (H&E) Staining and Immunohistochemistry (IHC)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AR | Androgen receptor |
| BPH | Benign prostatic hyperplasia |
| DHT | Dihydrotestosterone |
| E2F1 | E2F transcription factor 1 |
| Fina | Finasteride |
| PCNA | Proliferating cell nuclear antigen |
| PSA | Prostate-specific antigen |
| Saw | Saw palmetto |
| STAT3 | Signal transducer and activator of transcription 3 |
| TP | Testosterone propionate |
| TGFβ1 | Transforming growth factor beta 1 |
| UMB | Umbelliferone |
References
- Youn, D.H.; Park, J.; Kim, H.L.; Jung, Y.; Kang, J.; Jeong, M.Y.; Sethi, G.; Seok Ahn, K.; Um, J.Y. Chrysophanic acid reduces testosterone-induced benign prostatic hyperplasia in rats by suppressing 5alpha-reductase and extracellular signal-regulated kinase. Oncotarget 2017, 8, 9500–9512. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Fan, X.R.; Xie, H.; He, Q.H.; Nie, Y.S.; Zhang, M.; Yan, M. Anti-Inflammatory and Antioxidant Effects of Kelong-Capsule on Testosterone-Induced Benign Prostatic Hyperplasia in Rats. Evid. Based Complement. Altern. Med. eCAM 2018, 2018, 5290514. [Google Scholar] [CrossRef]
- Sheng, J.; Yang, Y.; Cui, Y.; He, S.; Wang, L.; Liu, L.; He, Q.; Lv, T.; Han, W.; Yu, W.; et al. M2 macrophage-mediated interleukin-4 signalling induces myofibroblast phenotype during the progression of benign prostatic hyperplasia. Cell Death Dis. 2018, 9, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Shin, J.; Choi, H.; Jo, A.; Pan, S.; Bae, D.; Lee, Y.; Choi, C. Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5alpha-Reductase and Androgen Receptor Activities in a Rat Model. Nutrients 2017, 9, 1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, Y.S.; Karna, K.K.; Choi, B.R.; Park, J.K. Finasteride and Erectile Dysfunction in Patients with Benign Prostatic Hyperplasia or Male Androgenetic Alopecia. World J. Men’s Health 2019, 37, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, S.A.; Chung, D.E.; Lee, R.K.; Scofield, S.; Te, A.E. A 5-year retrospective analysis of 5alpha-reductase inhibitors in men with benign prostatic hyperplasia: Finasteride has comparable urinary symptom efficacy and prostate volume reduction, but less sexual side effects and breast complications than dutasteride. Int. J. Clin. Pract. 2012, 66, 1052–1055. [Google Scholar] [PubMed]
- D’Amico, R.; Genovese, T.; Cordaro, M.; Siracusa, R.; Gugliandolo, E.; Peritore, A.F.; Interdonato, L.; Crupi, R.; Cuzzocrea, S.; Di Paola, R.; et al. Palmitoylethanolamide/Baicalein Regulates the Androgen Receptor Signaling and NF-kappaB/Nrf2 Pathways in Benign Prostatic Hyperplasia. Antioxidants 2021, 10, 1014. [Google Scholar] [CrossRef]
- Kwon, Y. Use of saw palmetto (Serenoa repens) extract for benign prostatic hyperplasia. Food Sci. Biotechnol. 2019, 28, 1599–1606. [Google Scholar] [CrossRef]
- Pais, P. Potency of a novel saw palmetto ethanol extract, SPET-085, for inhibition of 5alpha-reductase II. Adv. Ther. 2010, 27, 555–563. [Google Scholar] [CrossRef]
- Powell, S.M.; Christiaens, V.; Voulgaraki, D.; Waxman, J.; Claessens, F.; Bevan, C.L. Mechanisms of androgen receptor signalling via steroid receptor coactivator-1 in prostate. Endocr. Relat. Cancer 2004, 11, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Han, S.J.; Tsai, S.Y.; DeMayo, F.J.; Xu, J.; Tsai, M.J.; O’Malley, B.W. Roles of steroid receptor coactivator (SRC)-1 and transcriptional intermediary factor (TIF) 2 in androgen receptor activity in mice. Proc. Natl. Acad. Sci. USA 2005, 102, 9487–9492. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Xu, W.; Lin, J.; Peng, J.; Hong, Z. Qianliening capsule treats benign prostatic hyperplasia via induction of prostatic cell apoptosis. Mol. Med. Rep. 2013, 7, 848–854. [Google Scholar] [CrossRef] [Green Version]
- Robles, L.D.; Frost, A.R.; Davila, M.; Hutson, A.D.; Grizzle, W.E.; Chakrabarti, R. Down-regulation of Cdc6, a cell cycle regulatory gene, in prostate cancer. J. Biol. Chem. 2002, 277, 25431–25438. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Banerjee, S.; Acosta, E.P.; Lillard, J.W.; Singh, R. Resveratrol induces cell cycle arrest and apoptosis with docetaxel in prostate cancer cells via a p53/ p21WAF1/CIP1 and p27KIP1 pathway. Oncotarget 2017, 8, 17216–17228. [Google Scholar] [CrossRef] [Green Version]
- Guadagnin, E.; Narola, J.; Bonnemann, C.G.; Chen, Y.W. Tyrosine 705 Phosphorylation of STAT3 Is Associated with Phenotype Severity in TGFbeta1 Transgenic Mice. Biomed Res. Int. 2015, 2015, 843743. [Google Scholar] [CrossRef] [Green Version]
- Sherry, M.M.; Reeves, A.; Wu, J.K.; Cochran, B.H. STAT3 is required for proliferation and maintenance of multipotency in glioblastoma stem cells. Stem Cells 2009, 27, 2383–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaee, R.; Behravan, E.; Behravan, J.; Soltani, F.; Naderi, Y.; Emami, B.; Iranshahi, M. Antigenotoxic activities of the natural dietary coumarins umbelliferone, herniarin and 7-isopentenyloxy coumarin on human lymphocytes exposed to oxidative stress. Drug Chem. Toxicol. 2014, 37, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.F.; Wang, J.N.; Li, Z.; Wei, B.; Jin, J.; Gao, L.; Li, H.D.; Li, J.; Chen, H.Y.; Meng, X.M. 7-Hydroxycoumarin protects against cisplatin-induced acute kidney injury by inhibiting necroptosis and promoting Sox9-mediated tubular epithelial cell proliferation. Phytomedicine Int. J. Phytother. Phytopharm. 2020, 69, 153202. [Google Scholar] [CrossRef] [PubMed]
- Vijayalakshmi, A.; Sindhu, G. Umbelliferone arrest cell cycle at G0/G1 phase and induces apoptosis in human oral carcinoma (KB) cells possibly via oxidative DNA damage. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 92, 661–671. [Google Scholar] [CrossRef]
- Germoush, M.O.; Othman, S.I.; Al-Qaraawi, M.A.; Al-Harbi, H.M.; Hussein, O.E.; Al-Basher, G.; Alotaibi, M.F.; Elgebaly, H.A.; Sandhu, M.A.; Allam, A.A.; et al. Umbelliferone prevents oxidative stress, inflammation and hematological alterations, and modulates glutamate-nitric oxide-cGMP signaling in hyperammonemic rats. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 102, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Gonzalez, J.S.; Prado-Garcia, H.; Aguilar-Cazares, D.; Molina-Guarneros, J.A.; Morales-Fuentes, J.; Mandoki, J.J. Apoptosis and cell cycle disturbances induced by coumarin and 7-hydroxycoumarin on human lung carcinoma cell lines. Lung Cancer 2004, 43, 275–283. [Google Scholar] [CrossRef]
- Shen, J.Q.; Zhang, Z.X.; Shen, C.F.; Liao, J.Z. Anticarcinogenic effect of Umbelliferone in human prostate carcinoma: An in vitro study. J. B.U. Off. J. Balk. Union Oncol. 2017, 22, 94–101. [Google Scholar]
- Liu, J.; Fang, T.; Li, M.; Song, Y.; Li, J.; Xue, Z.; Li, J.; Bu, D.; Liu, W.; Zeng, Q.; et al. Pao Pereira Extract Attenuates Testosterone-Induced Benign Prostatic Hyperplasia in Rats by inhibiting 5alpha-Reductase. Sci. Rep. 2019, 9, 19703. [Google Scholar] [CrossRef]
- Song, J.H.; Hwang, B.; Chung, H.J.; Moon, B.; Kim, J.W.; Ko, K.; Kim, B.W.; Kim, W.R.; Kim, W.J.; Myung, S.C.; et al. Peanut Sprout Extracts Cultivated with Fermented Sawdust Medium Inhibits Benign Prostatic Hyperplasia In Vitro and In Vivo. World J. Men’s Health 2020, 38, 385–396. [Google Scholar] [CrossRef]
- Zhao, X.; Harashima, H.; Dissmeyer, N.; Pusch, S.; Weimer, A.K.; Bramsiepe, J.; Bouyer, D.; Rademacher, S.; Nowack, M.K.; Novak, B.; et al. A general G1/S-phase cell-cycle control module in the flowering plant Arabidopsis thaliana. PLoS Genet. 2012, 8, e1002847. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, L.A. Inhibition of E2F1 activity and cell cycle progression by arsenic via retinoblastoma protein. Cell Cycle 2017, 16, 2058–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Su, H.; Liao, X.; Han, C.; Yu, T.; Zhu, G.; Wang, X.; Winkler, C.A.; O’Brien, S.J.; Peng, T. Marker of proliferation Ki-67 expression is associated with transforming growth factor beta 1 and can predict the prognosis of patients with hepatic B virus-related hepatocellular carcinoma. Cancer Manag. Res. 2018, 10, 679–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldera, V.; Mellai, M.; Annovazzi, L.; Valente, G.; Tessitore, L.; Schiffer, D. Stat3 expression and its correlation with proliferation and apoptosis/autophagy in gliomas. J. Oncol. 2008, 2008, 219241. [Google Scholar] [CrossRef] [Green Version]
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20 (Suppl. 3), S11–S18. [Google Scholar] [CrossRef] [Green Version]
- Tao, R.; Miao, L.; Yu, X.; Orgah, J.O.; Barnabas, O.; Chang, Y.; Liu, E.; Fan, G.; Gao, X. Cynomorium songaricum Rupr demonstrates phytoestrogenic or phytoandrogenic like activities that attenuates benign prostatic hyperplasia via regulating steroid 5-alpha-reductase. J. Ethnopharmacol. 2019, 235, 65–74. [Google Scholar] [CrossRef]
- Tombal, B. A holistic approach to androgen deprivation therapy: Treating the cancer without hurting the patient. Urol. Int. 2009, 83, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Kandil, S.; Westwell, A.D.; McGuigan, C. 7-Substituted umbelliferone derivatives as androgen receptor antagonists for the potential treatment of prostate and breast cancer. Bioorg. Med. Chem. Lett. 2016, 26, 2000–2004. [Google Scholar] [CrossRef] [Green Version]
- Park, H.S.; Seo, C.S.; Wijerathne, C.U.; Jeong, H.Y.; Moon, O.S.; Seo, Y.W.; Won, Y.S.; Son, H.Y.; Lim, J.H.; Kwun, H.J. Effect of Veratrum maackii on Testosterone Propionate-Induced Benign Prostatic Hyperplasia in Rats. Biol. Pharm. Bull. 2019, 42, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Altintas, D.M.; Shukla, M.S.; Goutte-Gattat, D.; Angelov, D.; Rouault, J.P.; Dimitrov, S.; Samarut, J. Direct cooperation between androgen receptor and E2F1 reveals a common regulation mechanism for androgen-responsive genes in prostate cells. Mol. Endocrinol. 2012, 26, 1531–1541. [Google Scholar] [CrossRef] [Green Version]
- Glorian, V.; Allegre, J.; Berthelet, J.; Dumetier, B.; Boutanquoi, P.M.; Droin, N.; Kayaci, C.; Cartier, J.; Gemble, S.; Marcion, G.; et al. DNA damage and S phase-dependent E2F1 stabilization requires the cIAP1 E3-ubiquitin ligase and is associated with K63-poly-ubiquitination on lysine 161/164 residues. Cell Death Dis. 2017, 8, e2816. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Liu, B.; Gu, Q.; Su, L.; Yu, Y.; Zhu, Z. Stat6 cooperates with Sp1 in controlling breast cancer cell proliferation by modulating the expression of p21(Cip1/WAF1) and p27 (Kip1). Cell. Oncol. 2013, 36, 79–93. [Google Scholar] [CrossRef]
- Jimenez-Orozco, F.A.; Lopez-Gonzalez, J.S.; Nieto-Rodriguez, A.; Velasco-Velazquez, M.A.; Molina-Guarneros, J.A.; Mendoza-Patino, N.; Garcia-Mondragon, M.J.; Elizalde-Galvan, P.; Leon-Cedeno, F.; Mandoki, J.J. Decrease of cyclin D1 in the human lung adenocarcinoma cell line A-427 by 7-hydroxycoumarin. Lung Cancer 2001, 34, 185–194. [Google Scholar] [CrossRef]
- Chakraborty, D.; Sumova, B.; Mallano, T.; Chen, C.W.; Distler, A.; Bergmann, C.; Ludolph, I.; Horch, R.E.; Gelse, K.; Ramming, A.; et al. Activation of STAT3 integrates common profibrotic pathways to promote fibroblast activation and tissue fibrosis. Nat. Commun. 2017, 8, 1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Jin, B.R.; An, H.J. Psoralea corylifolia L. extract ameliorates benign prostatic hyperplasia by regulating prostate cell proliferation and apoptosis. J. Ethnopharmacol. 2021, 273, 113844. [Google Scholar] [CrossRef]
- Jin, B.R.; Cheon, S.Y.; Kim, H.J.; Kim, M.S.; Lee, K.H.; An, H.J. Anti-Proliferative Effects of Standardized Cornus officinalis on Benign Prostatic Epithelial Cells via the PCNA/E2F1-Dependent Cell Cycle Pathway. Int. J. Mol. Sci. 2020, 21, 9567. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| PCNA | TTAAACGGTTGCAGGCGTAG | AGGAAAGTCTAGCTGGTTTCGG |
| AR | GAGCCAGGTGTAGTGTGTGC | TCGTCCACGTGTAAGTTGCG |
| SRC-1 | GCTGGTATCCTTCCTTAGTG | TGGCGTTGCTTGTTGTGGTG |
| PSA | ATAGGATTGCCCAGGCAGAA | CTAAGGGTAAAAGCAGGGAGAGAGT |
| Cyclin D1 | ACGGCCGAGAAGCTGTGCATC | CCTCCGCCTCTGGCATTTTGGAG |
| Cdk4 | ATGGCTACCTCTCGATATGAGC | CATTGGGGACTCTCACACTCT |
| Cyclin E | GACGGGGAGCTCAAAACTGA | TACAACGGAGCCCAGAACAC |
| Cdk2 | TTCTATGCCTGATTACAAGCC | CTGGCTTGGTCACATCCT |
| p27Kip1 | AACGTGCGAGTGTCTAACGG | CCCTCTAGGGGTTTGTGATTCT |
| Rb | ATGGTTCACCTCGAACACCC | TTTCGACACAACCCTGTCCC |
| E2F1 | AAGAACCGCTGTTGTCCCG | TCGAGGCCGAAGTGGTAGTC |
| TGFβ1 | CTATCGACATGGAGCTGGTGAAG | CGTGGAGCTGAAGCAATAGTTGG |
| β-actin | GGCCAGGTCATCACCATTGG | CTTTGCGGATGTCCACGTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Jin, B.-R.; An, H.-J. Umbelliferone Ameliorates Benign Prostatic Hyperplasia by Inhibiting Cell Proliferation and G1/S Phase Cell Cycle Progression through Regulation of STAT3/E2F1 Axis. Int. J. Mol. Sci. 2021, 22, 9019. https://doi.org/10.3390/ijms22169019
Kim H-J, Jin B-R, An H-J. Umbelliferone Ameliorates Benign Prostatic Hyperplasia by Inhibiting Cell Proliferation and G1/S Phase Cell Cycle Progression through Regulation of STAT3/E2F1 Axis. International Journal of Molecular Sciences. 2021; 22(16):9019. https://doi.org/10.3390/ijms22169019
Chicago/Turabian StyleKim, Hyo-Jung, Bo-Ram Jin, and Hyo-Jin An. 2021. "Umbelliferone Ameliorates Benign Prostatic Hyperplasia by Inhibiting Cell Proliferation and G1/S Phase Cell Cycle Progression through Regulation of STAT3/E2F1 Axis" International Journal of Molecular Sciences 22, no. 16: 9019. https://doi.org/10.3390/ijms22169019
APA StyleKim, H.-J., Jin, B.-R., & An, H.-J. (2021). Umbelliferone Ameliorates Benign Prostatic Hyperplasia by Inhibiting Cell Proliferation and G1/S Phase Cell Cycle Progression through Regulation of STAT3/E2F1 Axis. International Journal of Molecular Sciences, 22(16), 9019. https://doi.org/10.3390/ijms22169019