
Molecular Mechanisms of Chemoresistance Induced by Cisplatin in NSCLC Cancer Therapy

 , , and
, , and
Abstract
:1. Lung Cancer from a Global Perspective
1.1. Histopathological Type
1.2. Treatment of NSCLC
1.3. The Effect of Cisplatin
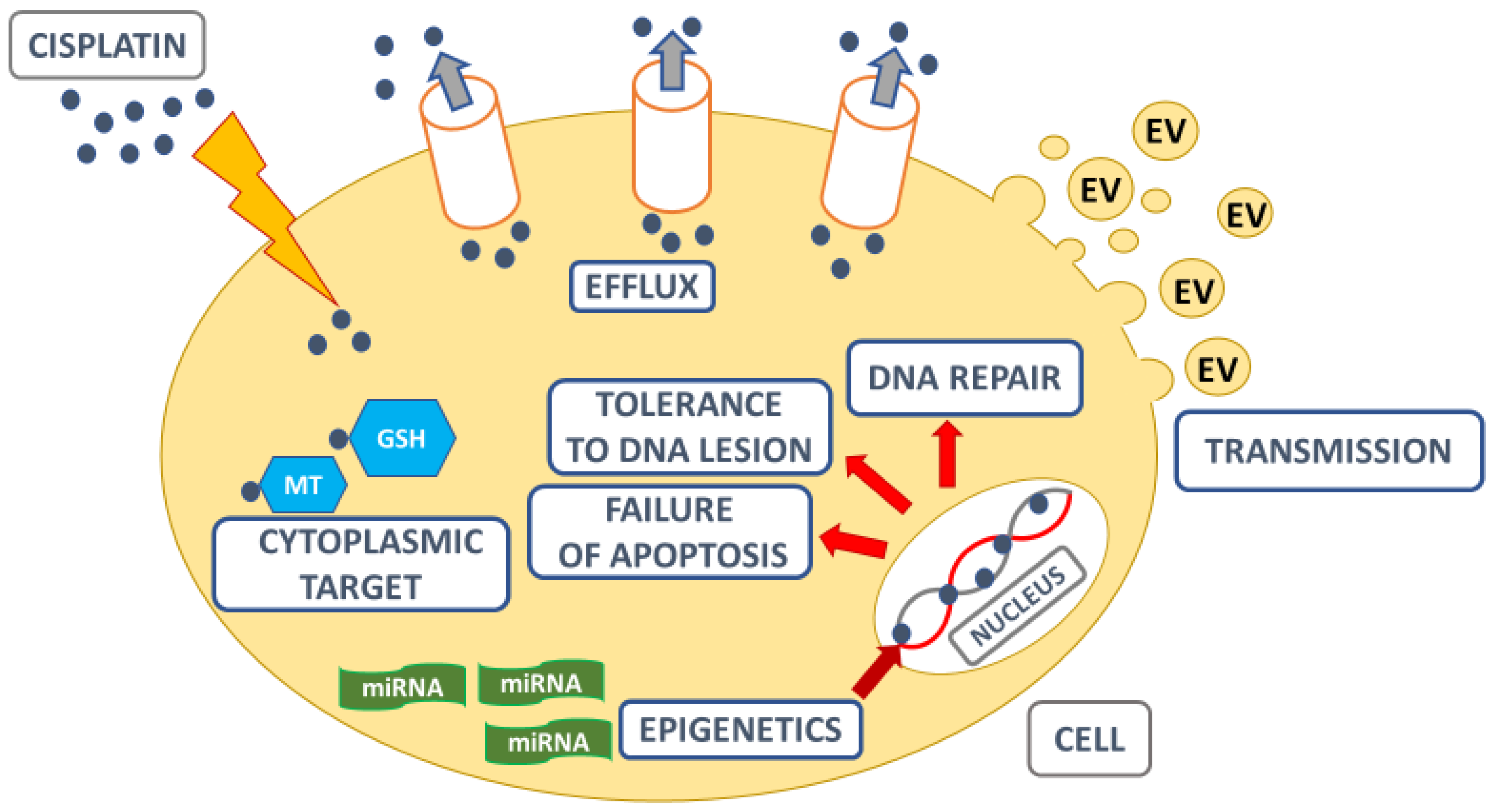
2. Mechanisms Underlying Cisplatin Resistance
2.1. Repair of DNA Damage
2.1.1. Nucleotide Excision Repair
2.1.2. Post-Replication Repair
2.1.3. Fanconi Anaemia and ICL Repair
2.1.4. Homologous Recombination Repair
2.2. Apoptosis
2.3. ABC Transporters
2.4. Epigenetic Regulation by miRNAs
MiRNAs Reducing the Cisplatin Resistance or Restoring the Sensitivity to Chemotherapeutics
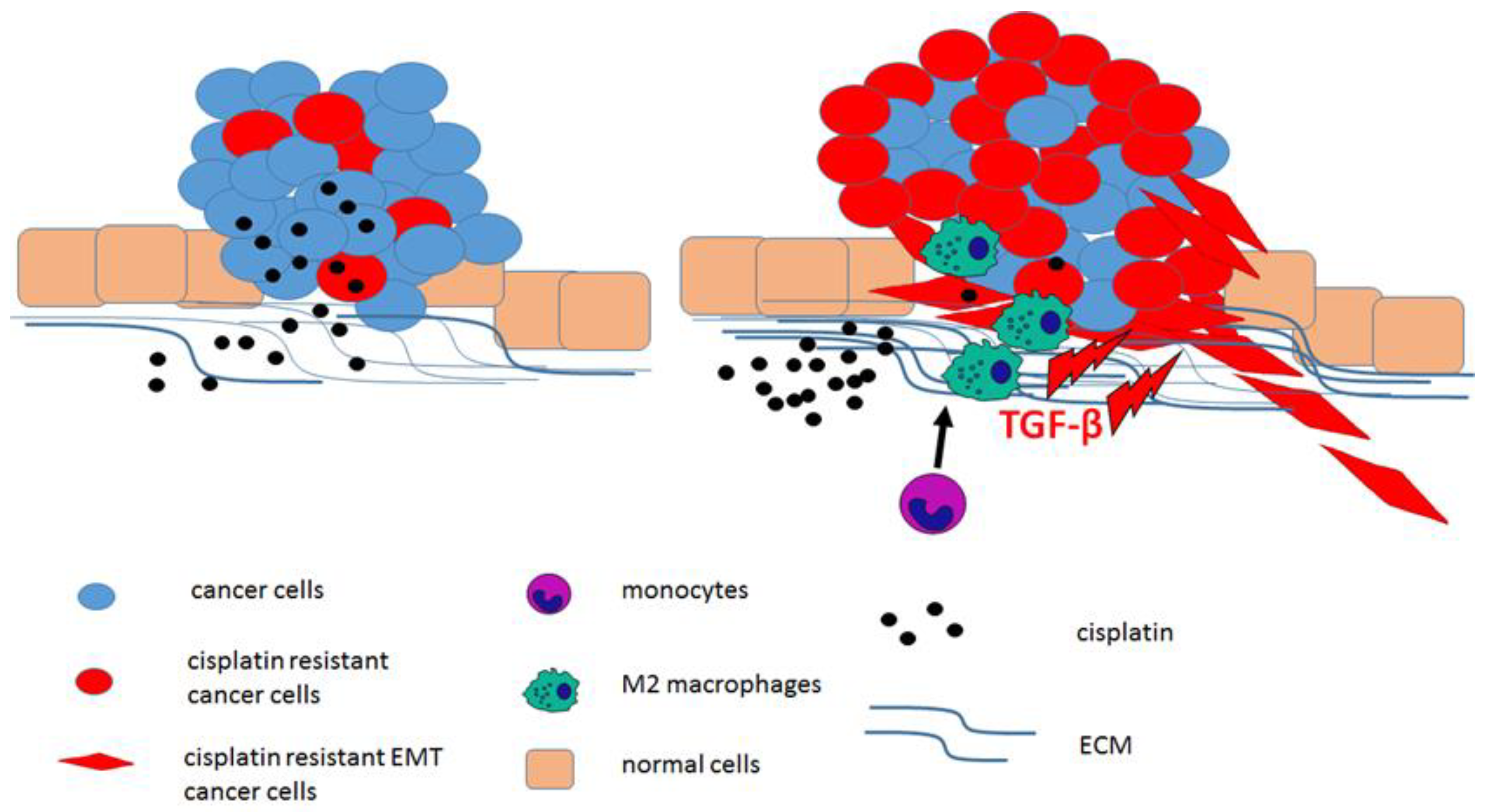
2.5. Cisplatin Resistance and the Tumour Microenvironment (TME)
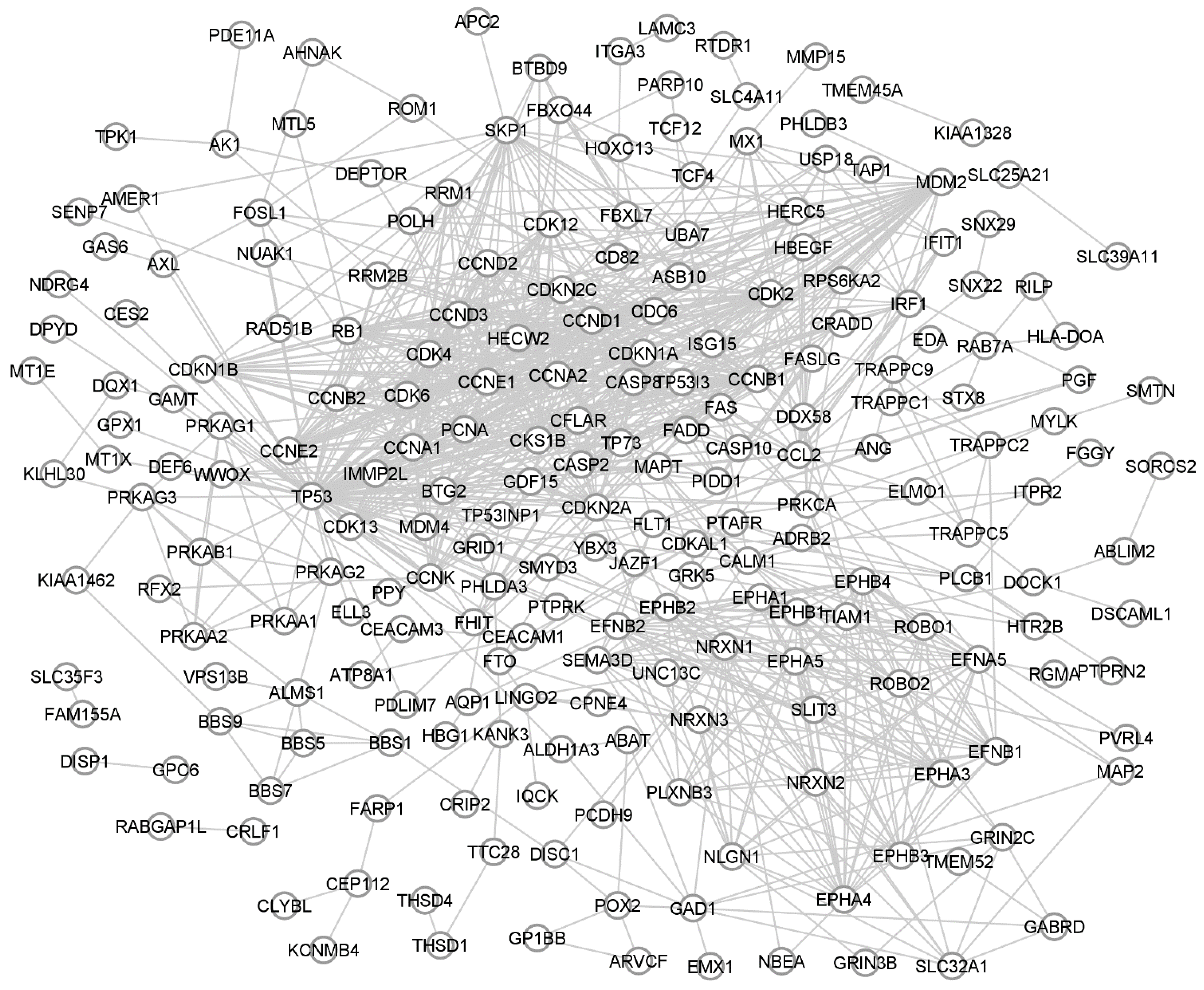
3. Protein-Protein Interaction Changes in NSCLC Caused by the Acquisition of Cisplatin Resistance
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roys, A.; Chang, X.; Liu, Y.; Xu, X.; Wu, Y.; Zuo, D. Resistance mechanisms and potent-targeted therapies of ROS1-positive lung cancer. Cancer Chemother. Pharmacol. 2019, 84, 679–688. [Google Scholar] [CrossRef]
- Kryczka, J.; Migdalska-Sęk, M.; Kordiak, J.; Kiszałkiewicz, J.M.; Pastuszak-Lewandoska, D.; Antczak, A.; Brzeziańska-Lasota, E. Serum Extracellular Vesicle-Derived miRNAs in Patients with Non-Small Cell Lung Cancer—Search for Non-Invasive Diagnostic Biomarkers. Diagnostics 2021, 11, 425. [Google Scholar] [CrossRef]
- Cancer Today. Available online: http://gco.iarc.fr/today/home (accessed on 15 January 2020).
- Blandin Knight, S.; Crosbie, P.A.; Balata, H.; Chudziak, J.; Hussell, T.; Dive, C. Progress and prospects of early detection in lung cancer. Open Biol. 2017, 7, 170070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romaszko, A.; Doboszyńska, A. Multiple primary lung cancer: A literature review. Adv. Clin. Exp. Med. 2018, 27, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Travis, W.D.; Brambilla, E.; Nicholson, A.G.; Yatabe, Y.; Austin, J.H.; Beasley, M.B.; Chirieac, L.R.; Dacic, S.; Duhig, E.; Flieder, D.B.; et al. The 2015 world health organization classification of lung tumors: Impact of genetic, clinical and radiologic advances Since the 2004 classification. J. Thorac. Oncol. 2015, 10, 1243–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.; Ridge, C.A.; Nicholson, A.G.; Mirsadraee, S. The 8th lung cancer TNM classification and clinical staging system: Review of the changes and clinical implications. Quant. Imaging Med. Surg. 2018, 8, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Osmani, L.; Askin, F.; Gabrielson, E.; Li, Q.K. Current WHO guidelines and the critical role of immunohistochemical markers in the subclassification of non-small cell lung carcinoma (NSCLC). Moving from targeted therapy to immunotherapy. Semin. Cancer Biol. 2018, 52, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.A.; Glynn, T.J. Epidemiology of lung cancer. Radiol. Clin. N. Am. 2000, 38, 453–470. [Google Scholar] [CrossRef]
- Greenhalgh, J.; Boland, A.; Bates, V.; Vecchio, F.; Dundar, Y.; Chaplin, M.; Green, J.A. First-line treatment of advanced epidermal growth factor receptor (EGFR) mutation positive non-squamous non-small cell lung cancer. Cochrane Database Syst. Rev. 2016, 25, CD010383. [Google Scholar] [CrossRef]
- Non-Small Cell Lung Cancer Treatment (PDQ®)–Health Professional Version. National Cancer Institute. Published 13 December 2019. Available online: https://www.cancer.gov/types/lung/hp/non-small-cell-lung-treatment-pdq (accessed on 12 August 2021).
- Basu, A.; Krishnamurthy, S. Cellular responses to cisplatin-induced DNA damage. J. Nucleic Acids 2010, 2010, e201367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cetintas, V.B.; Kucukaslan, A.S.; Kosova, B.; Tetik, A.; Selvi, N.; Cok, G.; Gunduz, C.; Eroglu, Z. Cisplatin resistance induced by decreased apoptotic activity in non-small-cell lung cancer cell lines. Cell Biol. Int. 2012, 36, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Goss, G.D.; Tsvetkova, E. Drug resistance and its significance for treatment decisions in non-small-cell lung cancer. Curr. Oncol. 2012, 19, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarin, N.; Engel, F.; Kalayda, G.V.; Mannewitz, M.; Cinatl, J., Jr.; Rothweiler, F.; Michaelis, M.; Saafan, H.; Ritter, C.A.; Jaehde, U.; et al. Cisplatin resistance in non-small cell lung cancer cells is associated with an abrogation of cisplatin-induced G2/M cell cycle arrest. PLoS ONE 2017, 12, e0181081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Vitale, I.; Michels, J.; Brenner, C.; Szabadkai, G.; Harel-Bellan, A.; Kroemer, G.J.C.D. Systems biology of cisplatin resistance: Past, present and future. Cell Death Dis. 2014, 5, e1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timerbaev, A.R.; Hartinger, C.G.; Aleksenko, S.S.; Keppler, B.K. Interactions of antitumor metallodrugs with serum proteins: Advances in characterization using modern analytical methodology. Chem. Rev. 2006, 106, 2224–2248. [Google Scholar] [CrossRef] [PubMed]
- Slater, A.F.; Nobel, C.S.; Maellaro, E.; Bustamante, J.; Kimland, M.; Orrenius, S. Nitrone spin traps and a nitroxide antioxidant inhibit a common pathway of thymocyte apoptosis. Biochem. J. 1995, 306, 771–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikuta, K.; Takemura, K.; Sasaki, K.; Kihara, M.; Nishimura, M.; Ueda, N.; Naito, S.; Lee, E.; Shimizu, E.; Yamauchi, A. Expression of multidrug resistance proteins and accumulation of cisplatin in human non-small cell lung cancer cells. Biol. Pharm. Bull. 2005, 28, 707–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosell, R.; Lord, R.V.N.; Taron, M.; Reguart, N. DNA repair and cisplatin resistance in non-small-cell lung cancer. Lung Cancer 2002, 38, 217–227. [Google Scholar] [CrossRef]
- Siddik, Z.H. Mechanisms of action of cancer chemotherapeutic agents: DNA-interactive alkylating agents and antitumour platinum-based drugs. In The Cancer Handbook; Alison, M.R., Ed.; John Wiley & Sons Ltd.: Houston, NJ, USA, 2005. [Google Scholar]
- Kryczka, J.; Boncela, J. Cell migration related to MDR—Another impediment to effective chemotherapy? Molecules 2018, 23, 331. [Google Scholar] [CrossRef] [Green Version]
- Sosa Iglesias, V.; Giuranno, L.; Dubois, L.J.; Theys, J.; Vooijs, M. Drug resistance in non-small cell lung cancer: A potential for NOTCH targeting? Front. Oncol. 2018, 8, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monzo, M.; Rosell, R.; Taron, M. Drug resistance in non-small cell lung cancer. Lung Cancer 2001, 34, S91–S94. [Google Scholar] [CrossRef]
- Fadejeva, I.; Olschewski, H.; Hrzenjak, A. MicroRNAs as regulators of cisplatin-resistance in non-small cell lung carcinomas. Oncotarget 2017, 8, 115754–115773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macerelli, M.; Ganzinelli, M.; Gouedard, C.; Broggini, M.; Garassino, M.C.; Linardou, H.; Damia, G.; Wiesmüller, L. Can the response to a platinum-based therapy be predicted by the DNA repair status in non-small cell lung cancer? Cancer Treat. Rev. 2016, 48, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, E.R.; Lippard, S.J. Structure, recognition, and processing of cisplatin-DNA adducts. Chem. Rev. 1999, 99, 2467–2498. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.; Saadat, N.; Myung, B.; Shekhar, M.P.V. Crosstalk between translesion synthesis, Fanconi anemia network, and homologous recombination repair pathways in interstrand DNA crosslink repair and development of chemoresistance. Mutat. Res. Rev. Mutat. Res. 2015, 763, 258–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatzemeier, U.; Von Pawel, J.; Gottfried, M.; Velde, G.T.; Mattson, K.; DeMarinis, F.; Harper, P.; Salvati, F.; Robinet, G.; Lucenti, A. Phase III comparative study of high-dose cisplatin versus a combination of paclitaxel and cisplatin in patients with advanced non-small-cell lung cancer. J. Clin. Oncol. 2000, 18, 3390–3399. [Google Scholar] [CrossRef]
- Ho, T.V.; Schärer, O.D. Translesion DNA synthesis polymerases in DNA interstrand crosslink repair. Environ. Mol. Mutagenesis 2010, 51, 552–566. [Google Scholar] [CrossRef]
- Chen, P.; Li, J.; Chen, Y.C.; Qian, H.; Chen, Y.J.; Su, J.Y.; Wu, M.; Lan, T. The functional status of DNA repair pathways determines the sensitization effect to cisplatin in non-small cell lung cancer cells. Cell Oncol. 2016, 39, 511–522. [Google Scholar] [CrossRef]
- Salehan, M.R.; Morse, H.R. DNA damage repair and tolerance: A role in chemotherapeutic drug resistance. Br. J. Biomed. Sci. 2013, 70, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Unk, I.; Hajdú, I.; Fátyol, K.; Szakál, B.; Blastyák, A.; Bermudez, V.; Hurwitz, J.; Prakash, L.; Prakash, S.; Haracska, L. Human SHPRH is a ubiquitin ligase for Mms2–Ubc13-dependent polyubiquitylation of proliferating cell nuclear antigen. Proc. Natl. Acad. Sci. USA 2006, 103, 18107–18112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, Y.; Suzuki, M.; Kawai, H.; Hishiki, A.; Hashimoto, H.; Masutani, C.; Hishida, T.; Suzuki, F.; Kamiya, K. En bloc transfer of polyubiquitin chains to PCNA in vitro is mediated by two different human E2-E3 pairs. Nucleic Acids Res. 2012, 40, 10394–10407. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.E.; Prakash, L.; Prakash, S. Pol31 and Pol32 subunits of yeast DNA polymerase δ are also essential subunits of DNA polymerase ζ. Proc. Natl. Acad. Sci. USA 2012, 109, 12455–12460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, T.-Q.; Li, R.; Shao, M.-H.; Ye, J.-Y.; Han, B.-H. RAD18 polymorphisms are associated with platinum-based chemotherapy toxicity in Chinese patients with non-small cell lung cancer. Acta Pharmacol. Sin. 2016, 37, 1490–1498. [Google Scholar] [CrossRef] [Green Version]
- Motegi, A.; Liaw, H.J.; Lee, K.Y.; Roest, H.P.; Maas, A.; Wu, X.; Myung, K. Polyubiquitination of proliferating cell nuclear antigen by HLTF and SHPRH prevents genomic instability from stalled replication forks. Proc. Natl. Acad. Sci. USA 2008, 105, 12411–12416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doles, J.; Oliver, T.G.; Cameron, E.R.; Hsu, G.; Jacks, T.; Walker, G.C.; Hemann, M.T. Suppression of Rev3, the catalytic subunit of Pol{zeta}, sensitizes drug-resistant lung tumors to chemotherapy. Proc. Natl. Acad. Sci. USA 2010, 107, 20786–20791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceppi, P.; Novello, S.; Cambieri, A.; Longo, M.; Monica, V.; Iacono, M.L.; Giaj-Levra, M.; Saviozzi, S.; Volante, M.; Papotti, M.; et al. Polymerase eta mRNA expression predicts survival of non-small cell lung cancer patients treated with platinum-based chemotherapy. Clin. Cancer Res. 2009, 15, 1039–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.; Huang, J. The Fanconi anemia pathway and DNA interstrand cross-link repair. Protein Cell 2011, 2, 704–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreassen, P.R.; D’Andrea, A.D.; Taniguchi, T. ATR couples FANCD2 monoubiquitination to the DNA-damage response. Genes Dev. 2004, 18, 1958–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, W.; Gao, L.; Aguila, B.; Kalvala, A.; Otterson, G.A.; Villalona-Calero, M.A. Fanconi anemia repair pathway dysfunction, a potential therapeutic target in lung cancer. Front. Oncol. 2014, 4, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Dexheimer, T.S.; Ai, Y.; Liang, Q.; Villamil, M.A.; Inglese, J.; Zhuang, Z. Selective and cell-active inhibitors of the USP1/UAF1 deubiquitinase complex reverse cisplatin resistance in non-small cell lung cancer cells. Chem. Biol. 2011, 18, 1390–1400. [Google Scholar] [CrossRef] [Green Version]
- Burkitt, K.; Ljungman, M. Phenylbutyrate interferes with the Fanconi anemia and BRCA pathway and sensitizes head and neck cancer cells to cisplatin. Mol. Cancer. 2008, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Chirnomas, D.; Taniguchi, T.; de la Vega, M.; Vaidya, A.P.; Vasserman, M.; Hartman, A.R.; Kennedy, R.; Foster, R.; Mahoney, J.; Seiden, M.V.; et al. Chemosensitization to cisplatin by inhibitors of the Fanconi anemia/BRCA pathway. Mol. Cancer Ther. 2006, 5, 952–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazón, G.; Mimitou, E.P.; Symington, L.S. SnapShot: Homologous Recombination in DNA Double-Strand Break Repair. Cell 2010, 142, e1–e648. [Google Scholar] [CrossRef] [Green Version]
- Daley, J.M.; Gaines, W.A.; Kwon, Y.; Sung, P. Regulation of DNA pairing in homologous recombination. Cold Spring Harb Perspect. Biol. 2014, 6, a017954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertl, H.A.; Russo, D.P.; Srivastava, N.; Brooks, J.T.; Dao, T.N.; LaRocque, J.R. The role of Blm helicase in homologous recombination, gene conversion tract length, and recombination between diverged sequences in Drosophila melanogaster. Genetics 2017, 207, 923–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Reed, E.; Li, Q.Q. Molecular basis of cellular response to cisplatin chemotherapy in non-small cell lung cancer (Review). Oncol. Rep. 2004, 12, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Han, C.Y.; Duan, F.G.; Fan, X.X.; Yao, X.J.; Parks, R.J.; Tang, Y.J.; Wang, M.F.; Liu, L.; Tsang, B.K.; et al. p53 sensitizes chemoresistant non-small cell lung cancer via elevation of reactive oxygen species and suppression of EGFR/PI3K/AKT signaling. Cancer Cell Int. 2019, 19, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Q.; Chen, Y.; Zhou, X. The janus face of p53-targeting ubiquitin ligases. Cells 2020, 9, 1656. [Google Scholar] [CrossRef] [PubMed]
- Zamagni, A.; Pasini, A.; Pirini, F.; Ravaioli, S.; Giordano, E.; Tesei, A.; Calistri, D.; Ulivi, P.; Fabbri, F.; Foca, F.; et al. CDKN1A upregulation and cisplatin-pemetrexed resistance in non-small cell lung cancer cells. Int. J. Oncol. 2020, 56, 1574–1584. [Google Scholar] [CrossRef] [Green Version]
- Dean, E.J.; Ward, T.; Pinilla, C.; Houghten, R.; Welsh, K.; Makin, G.; Ranson, M.; Dive, C. A small molecule inhibitor of XIAP induces apoptosis and synergises with vinorelbine and cisplatin in NSCLC. Br. J. Cancer 2010, 102, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Krepela, E.; Dankova, P.; Moravcikova, E.; Krepelova, A.; Prochazka, J.; Cermak, J.; Schützner, J.; Zatloukal, P.; Benkova, K. Increased expression of inhibitor of apoptosis proteins, survivin and XIAP in non-small cell lung carcinoma. Int. J. Oncol. 2009, 35, 1449–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivapurkar, N.; Reddy, J.; Chaudhary, P.M.; Gazdar, A.F. Apoptosis and lung cancer: A review. J. Cell Biochem. 2003, 88, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wang, X.J.; Huang, L.J.; Zhou, Y.A.; Tian, F.; Zhao, J.B.; Zhang, Z.P. High ABCG4 expression is associated with poor prognosis in non-small-cell lung cancer patients treated with cisplatin-based chemotherapy. PLoS ONE 2015, 10, e0135576. [Google Scholar]
- Prieto-Vila, M.; Takahashi, R.-U.; Usuba, W.; Kohama, I.; Ochiya, T. Drug resistance driven by cancer stem cells and their niche. Int. J. Mol. Sci. 2017, 18, 2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X. ABC Family Transporters. Adv. Exp. Med. Biol. 2019, 1141, 13–100. [Google Scholar]
- Shanker, M.; Willcutts, D.; Roth, J.A.; Ramesh, R. Drug resistance in lung cancer. Lung Cancer 2010, 1, 23–36. [Google Scholar]
- Wangari-Talbot, J.; Hopper-Borge, E. Drug resistance mechanisms in non-small cell lung carcinoma. J. Cancer Res. Updates 2013, 2, 265–282. [Google Scholar]
- Wang, Q.; Geng, F.; Zhou, H.; Chen, Y.; Du, J.; Zhang, X.; Song, D.; Zhao, H. MDIG promotes cisplatin resistance of lung adenocarcinoma by regulating ABC transporter expression via activation of the WNT/β-catenin signaling pathway. Oncol. Lett. 2019, 18, 4294–4307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ween, M.P.; Armstrong, M.A.; Oehler, M.K.; Ricciardelli, C. The role of ABC transporters in ovarian cancer progression and chemoresistance. Crit. Rev. Oncol. Hematol. 2015, 96, 220–256. [Google Scholar] [CrossRef] [PubMed]
- Vesel, M.; Rapp, J.; Feller, D.; Kiss, E.; Jaromi, L.; Meggyes, M.; Miskei, G.; Duga, B.; Smuk, G.; Laszlo, T.; et al. ABCB1 and ABCG2 drug transporters are differentially expressed in non-small cell lung cancers (NSCLC) and expression is modified by cisplatin treatment via altered Wnt signaling. Respir. Res. 2017, 18, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Yadav, T.; Rani, V. Exploring miRNA based approaches in cancer diagnostics and therapeutics. Crit. Rev. Oncol. Hematol. 2016, 98, 12–23. [Google Scholar] [CrossRef]
- Just, C.; Knief, J.; Lazar-Karsten, P.; Petrova, E.; Hummel, R.; Röcken, C.; Wellner, U.; Thorns, C. MicroRNAs as Potential Biomarkers for Chemoresistance in Adenocarcinomas of the Esophagogastric Junction. J. Oncol. 2019, 2019, 4903152. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.-W.; Wang, Y.-C.; Wang, L.; Chen, C.Y.; Lee, H. A low microRNA-630 expression confers resistance to tyrosine kinase inhibitors in EGFR-mutated lung adenocarcinomas via miR-630/YAP1/ERK feedback loop. Theranostics 2018, 8, 1256–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Gao, Y.; Zhang, K.; Li, C.; Pan, Y.; Chen, J.; Wang, R.; Chen, L. MicroRNAs as regulators of cisplatin resistance in lung cancer. CPB 2015, 37, 1869–1880. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-J.; Wu, D.-W.; Wang, G.-C.; Wang, Y.-C.; Chen, C.-Y.; Lee, H. MicroRNA-630 may confer favorable cisplatin-based chemotherapy and clinical outcomes in non-small cell lung cancer by targeting Bcl-2. Oncotarget 2018, 9, 13758–13767. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Morselli, E.; Vitale, I.; Kepp, O.; Senovilla, L.; Criollo, A.; Servant, N.; Paccard, C.; Hupé, P.; Robert, T.; et al. miR-181a and miR-630 regulate cisplatin-induced cancer cell death. Cancer Res. 2010, 70, 1793–1803. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Le Teuff, G.; Lacas, B.; Tsao, M.S.; Graziano, S.; Pignon, J.P.; Douillard, J.Y.; Le Chevalier, T.; Seymour, L.; Filipits, M.; et al. Prognostic and predictive effect of TP53 mutations in patients with non-small cell lung cancer from adjuvant cisplatin-based therapy randomized trials: A LACE-bio pooled analysis. J. Thorac Oncol. 2016, 11, 850–861. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.-H.; Wang, X.; Feng, Q. EGCG enhances the efficacy of cisplatin by downregulating hsa-miR-98-5p in NSCLC A549 cells. Nutr. Cancer 2014, 66, 636–644. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, L.-J.; Yang, Y.-C.; Wang, Z.-X.; Wang, R. MiR-224 promotes the chemoresistance of human lung adenocarcinoma cells to cisplatin via regulating G1/S transition and apoptosis by targeting p21(WAF1/CIP1). Br. J. Cancer 2014, 111, 339–354. [Google Scholar] [CrossRef]
- Li, Q.; Yang, Z.; Chen, M.; Liu, Y. Downregulation of microRNA-196a enhances the sensitivity of non-small cell lung cancer cells to cisplatin treatment. Int. J. Mol. Med. 2016, 37, 1067–1074. [Google Scholar] [CrossRef]
- Rahim, A.; Afzal, M.; Naveed, A.K. Genetic polymorphism of miRNA-196a and its target gene annexin-A1 expression based on ethnicity in Pakistani female breast cancer patients. Pak. J. Med. Sci. 2019, 35, 1598–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizzarro, V.; Belvedere, R.; Milone, M.R.; Pucci, B.; Lombardi, R.; Bruzzese, F.; Popolo, A.; Parente, L.; Budillon, A.; Petrella, A. Annexin A1 is involved in the acquisition and maintenance of a stem cell-like/aggressive phenotype in prostate cancer cells with acquired resistance to zoledronic acid. Oncotarget 2015, 6, 25076–25092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Min, J.; Wu, H.; Zhang, H.; Wang, C.; Tan, G.; Zhang, F. Annexin A1 is a potential biomarker of bone metastasis in small cell lung cancer. Oncol. Lett. 2021, 21, 141. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Lu, K.H.; Wang, K.M.; Sun, M.; Zhang, E.B.; Yang, J.S.; Yin, D.D.; Liu, Z.L.; Zhou, J.; Liu, Z.J. MicroRNA-196a promotes non-small cell lung cancer cell proliferation and invasion through targeting HOXA5. BMC Cancer 2012, 12, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Zhang, L.; Yao, Q.; Tao, Z. miR-15b regulates cisplatin resistance and metastasis by targeting PEBP4 in human lung adenocarcinoma cells. Cancer Gene Ther. 2015, 22, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Gkountakos, A.; Sartori, G.; Falcone, I.; Piro, G.; Ciuffreda, L.; Carbone, C.; Tortora, G.; Scarpa, A.; Bria, E.; Milella, M.; et al. PTEN in lung cancer: Dealing with the problem, building on new knowledge and turning the game around. Cancers 2019, 11, 1141. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Huang, L.; He, J.; Cai, S.; Weng, Y.; Huang, S.; Ma, S. PTEN inhibits non-small cell lung cancer cell growth by promoting G0/G1 arrest and cell apoptosis. Oncol. Lett. 2019, 17, 1333–1340. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Hu, C.P.; He, B.X.; Chen, X.; Lu, X.X.; Xie, M.X.; Chen, Q. PTEN expression is a prognostic marker for patients with non-small cell lung cancer: A systematic review and meta-analysis of the literature. Oncotarget 2016, 7, 57832–57840. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.H.; Fang, H.Y.; Yang, J.M.; Liu, S.L.; Yao, Q.H.; Fan, Y.J.; Gao, F.H. MicroRNA-92b acts as an oncogene by targeting PTEN/AKT in NSCLC. Cell Biochem. Funct. 2020, 38, 1100–1110. [Google Scholar] [CrossRef]
- Liu, Z.-L.; Wang, H.; Liu, J.; Wang, Z.-X. MicroRNA-21 (miR-21) expression promotes growth, metastasis, and chemo- or radioresistance in non-small cell lung cancer cells by targeting PTEN. Mol. Cell Biochem. 2013, 372, 35–45. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S.; Ma, F.; Zhang, W. miRNA-328 overexpression confers cisplatin resistance in non-small cell lung cancer via targeting of PTEN. Mol. Med. Rep. 2018, 18, 4563–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magee, P.; Shi, L.; Garofalo, M. Role of microRNAs in chemoresistance. Ann. Transl. Med. 2015, 3, 332. [Google Scholar] [PubMed]
- Acunzo, M.; Visone, R.; Romano, G.; Veronese, A.; Lovat, F.; Palmieri, D.; Bottoni, A.; Garofalo, M.; Gasparini, P.; Condorelli, G.; et al. miR-130a targets MET and induces TRAIL-sensitivity in NSCLC by downregulating miR-221 and 222. Oncogene 2012, 31, 634–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, M.; Qu, Q.; Wang, G.; Zhou, H. Let-7c sensitizes acquired cisplatin-resistant A549 cells by targeting ABCC2 and Bcl-XL. Pharmazie 2013, 68, 955–961. [Google Scholar]
- Jiang, Z.; Yin, J.; Fu, W.; Mo, Y.; Pan, Y.; Dai, L.; Huang, H.; Li, S.; Zhao, J. MiRNA 17 family regulates cisplatin-resistant and metastasis by targeting TGFbetaR2 in NSCLC. PLoS ONE 2014, 9, e94639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.Y.; Jiao, D.M.; Wang, J.; Hu, H.; Tang, X.; Chen, J.; Mou, H.; Lu, W. miR-206 regulates cisplatin resistance and EMT in human lung adenocarcinoma cells partly by targeting MET. Oncotarget 2016, 7, 24510–24526. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Chattopadhyay, D.; Chakrabarti, G. miR-17-5p downregulation contributes to paclitaxel resistance of lung cancer cells through altering beclin1 expression. PLoS ONE 2014, 9, e95716. [Google Scholar] [CrossRef] [Green Version]
- Czarnecka, K.H.; Szmyd, B.; Barańska, M.; Kaszkowiak, M.; Kordiak, J.; Antczak, A.; Pastuszak-Lewandoska, D.; Brzeziańska-Lasota, E. A strong decrease in TIMP3 expression mediated by the presence of miR-17 and 20a enables extracellular matrix remodeling in the NSCLC lesion surroundings. Front. Oncol. 2019, 9, 1372. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.-F.; Wang, J.-F.; Song, T.-T.; Zhang, J.; Wang, L. MiR-200b Inhibits Tumor Growth and Chemoresistance via Targeting p70S6K1 in Lung Cancer. Front. Oncol. 2020, 10, 643. [Google Scholar] [CrossRef]
- Chan, Y.T.; Lin, Y.C.; Lin, R.J.; Kuo, H.H.; Thang, W.C.; Chiu, K.P.; Yu, A.L. Concordant and discordant regulation of target genes by miR-31 and its isoforms. PLoS ONE 2013, 8, e58169. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-S.; Rossi, J.J. Molecular mechanisms of Dicer: Endonuclease and enzymatic activity. Biochem. J. 2017, 474, 1603–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.C.; Lima, N.; Da, S.; Sarian, L.O.; Matheu, A.; Ribeiro, M.L.; Derchain, S.F.M. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci. Rep. 2018, 8, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Wei, Q.; Koay, E.J.; Liu, Y.; Ning, B.; Bernard, P.W.; Zhang, N.; Han, H.; Katz, M.H.; Zhao, Z.; et al. Chemoresistance transmission via exosome-mediated EphA2 transfer in pancreatic cancer. Theranostics 2018, 8, 5986–5994. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Middleton, J.; Kim, T.; Laganà, A.; Piovan, C.; Secchiero, P.; Nuovo, G.J.; Cui, R.; Joshi, P.; Romano, G.; et al. A set of NF-κB-regulated microRNAs induces acquired TRAIL resistance in lung cancer. Proc. Natl. Acad. Sci. USA 2015, 112, E3355–E3364. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, M.; Di Leva, G.; Romano, G.; Nuovo, G.; Suh, S.S.; Ngankeu, A.; Taccioli, C.; Pichiorri, F.; Alder, H.; Secchiero, P.; et al. miR-221&222 regulate TRAIL resistance and enhance tumorigenicity through PTEN and TIMP3 downregulation. Cancer Cell. 2009, 16, 498–509. [Google Scholar]
- Larionova, I.; Cherdyntseva, N.; Liu, T.; Patysheva, M.; Rakina, M.; Kzhyshkowska, J. Interaction of tumor-associated macrophages and cancer chemotherapy. Oncoimmunology 2019, 8, 1596004. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-H.; Chang, J.-Y. New insights into mechanisms of cisplatin resistance: From tumor cell to microenvironment. Int. J. Mol. Sci. 2019, 20, 4136. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Kuo, K.T.; Wang, C.H.; Yeh, C.T.; Wang, Y. Cisplatin resistant lung cancer cells promoted M2 polarization of tumor-associated macrophages via the Src/CD155/MIF functional pathway. J. Exp. Clin. Cancer Res. 2019, 38, 180. [Google Scholar] [CrossRef] [Green Version]
- Kryczka, J.; Boncela, J. Leukocytes: The double-edged sword in fibrosis. Mediat. Inflamm. 2015, 2015, e652035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Kalantari, M.; Mohammadinejad, R.; Javaheri, T.; Sethi, G. Association of the Epithelial-Mesenchymal transition (EMT) with cisplatin resistance. Int. J. Mol. Sci. 2020, 21, 4002. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, W.; Wang, X.; Xu, C.; Zhang, N.; Di, W. Cisplatin-stimulated macrophages promote ovarian cancer migration via the CCL20-CCR6 axis. Cancer Lett. 2020, 472, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kirshberg, S.; Izhar, U.; Amir, G.; Demma, J.; Vernea, F.; Beider, K.; Shlomai, Z.; Wald, H.; Zamir, G.; Shapira, O.M.; et al. Involvement of CCR6/CCL20/IL-17 axis in NSCLC disease progression. PLoS ONE 2011, 6, e24856. [Google Scholar] [CrossRef] [PubMed]
- Wangpaichitr, M.; Kandemir, H.; Li, Y.Y.; Wu, C.; Nguyen, D.J.M.; Feun, L.G.; Kuo, M.T.; Savaraj, N. Relationship of metabolic alterations and PD-L1 expression in cisplatin resistant lung cancer. Cell Dev. Biol. 2017, 6, 183. [Google Scholar] [PubMed]
- Zhang, P.; Ma, Y.; Lv, C.; Huang, M.; Li, M.; Dong, B.; Liu, X.; An, G.; Zhang, W.; Zhang, J.; et al. Upregulation of programmed cell death ligand 1 promotes resistance response in non-small-cell lung cancer patients treated with neo-adjuvant chemotherapy. Cancer Sci. 2016, 107, 1563–1571. [Google Scholar] [CrossRef]
- Fournel, L.; Wu, Z.; Stadler, N.; Damotte, D.; Lococo, F.; Boulle, G.; Ségal-Bendirdjian, E.; Bobbio, A.; Icard, P.; Trédaniel, J.; et al. Cisplatin increases PD-L1 expression and optimizes immune check-point blockade in non-small cell lung cancer. Cancer Lett. 2019, 464, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Fan, X.; Zhu, W.; Huang, C.; Zhuang, W.; Xu, H.; Lin, X.; Hu, D.; Huang, Y.; Jiang, K.; et al. Prognostic significance of PD-L1 expression and tumor infiltrating lymphocyte in surgically resectable non-small cell lung cancer. Oncotarget 2017, 8, 83986–83994. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Yagishita, S.; Hagiwara, K.; Yoshioka, Y.; Kosaka, N.; Takeshita, F.; Fujiwara, T.; Tsuta, K.; Nokihara, H.; Tamura, T.; et al. The clinical relevance of the miR-197/CKS1B/STAT3-mediated PD-L1 network in chemoresistant non-small-cell lung cancer. Mol. Ther. 2015, 23, 717–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Collins, D.; Dolly, S.; McDonald, F.; O’Brien, M.E.R.; Yap, T.A. Targeting the PD-1/PD-L1 axis in non-small cell lung cancer. Curr. Probl. Cancer. 2017, 41, 111–124. [Google Scholar] [CrossRef]
- Abdel-Rahman, O. Correlation between PD-L1 expression and outcome of NSCLC patients treated with anti-PD-1/PD-L1 agents: A meta-analysis. Crit Rev. Oncol. Hematol. 2016, 101, 75–85. [Google Scholar] [CrossRef]
- Gettinger, S.N.; Horn, L.; Gandhi, L.; Spigel, D.R.; Antonia, S.J.; Rizvi, N.A.; Powderly, J.D.; Heist, R.S.; Carvajal, R.D.; Jackman, D.M.; et al. Overall survival and long-term safety of nivolumab (anti-programmed death 1 antibody, BMS-936558, ONO-4538) in patients with previously treated advanced non-small-cell lung cancer. J. Clin. Oncol. 2015, 33, 2004–2012. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, K.; Graulus, G.J.; Mesotten, L.; Thomeer, M.; Derveaux, E.; Noben, J.P.; Guedens, W.; Adriaensens, P. The metabolic landscape of lung cancer: New insights in a disturbed glucose metabolism. Front. Oncol. 2019, 9, 1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The warburg effect: How does it benefit cancer cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, B.; Mohd Omar, M.F.; Soong, R. The Warburg effect and drug resistance. Br. J. Pharmacol. 2016, 173, 970–979. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; Coller, H.A. Partners in the Warburg effect. eLife 2016, 5, e15938. [Google Scholar] [CrossRef]
- Home—GEO—NCBI. Available online: https://www.ncbi.nlm.nih.gov/geo/ (accessed on 14 April 2021).
- Makondi, P.T.; Chu, C.-M.; Wei, P.-L.; Chang, Y.-J. Prediction of novel target genes and pathways involved in irinotecan-resistant colorectal cancer. PLoS ONE 2017, 12, e0180616. [Google Scholar] [CrossRef] [Green Version]
- STRING: Functional Protein Association Networks. Available online: https://string-db.org/ (accessed on 14 April 2021).
- Otasek, D.; Morris, J.H.; Bouças, J.; Pico, A.R.; Demchak, B. Cytoscape automation: Empowering workflow-based network analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [Green Version]
- DAVID Functional Annotation Bioinformatics Microarray Analysis. Available online: https://david.ncifcrf.gov/ (accessed on 14 April 2021).
- KEGG PATHWAY Database. Available online: https://www.kegg.jp/kegg/pathway.html (accessed on 14 April 2021).

{kind=link}
{kind=link}
{kind=link}
| miRNA | Target Gene(s) | Chemoresistance | References |
|---|---|---|---|
| ↑miRNA-15b | ↓PEBP4 | TKIs resistance | [80] |
| ↑miRNA-20 | ↓MMP2 | paclitaxel resistance | [93] |
| ↑miRNA-21, | ↓PTEN, BCL2, IGF1R | TKIs resistance | [67] |
| ↑miRNA-21,miRNA-30c, ↑miRNA-100 | ↓caspase-3, caspase-8, TRAF7, FoxO3a ↑NF-κB signalling | TRAIL resistance | [99] |
| ↑miRNA-21, miRNA-92b, miRNA-328 | ↓PTEN | cisplatin resistance | [84,85,86] |
| ↑miRNA-31, | ↓DICER1, ABCB9 | cisplatin resistance | [96] |
| miRNA-98-5p | ↓MAPK signalling | cisplatin resistance EGCG-cisplatin resistance | [73] |
| ↑miRNA-30b/c, miRNA-221/222 | ↓APAF-1, BIM ↓SRC, PKC-ε | TKIs resistance | [100] |
| ↑miRNA-196a | ↓Annexin-A1, HOXA5 | cisplatin resistance | [75,79] |
| ↑miRNA-221/222 | ↓PTEN, TIMP3 | TRAIL resistance | [87,88] |
| ↑miRNA-224 | ↓p21WAF1/CIP | cisplatin resistance | [69,74] |
| ↑miRNA-630 | ↓CDC7 | cisplatin resistance | [27,69] |
| ↑Let-7b | ↓ABCC2, BCL-XL | ↑sensitivity to cisplatin | [89] |
| ↑miRNA-17-5p | ↓BECN1, TGFβR2 | ↑sensitivity to cisplatin ↑sensitivity to paclitaxel | [92] |
| ↑miRNA-17-5p | ↑TIMP | reversing of EMT phenotype, ECM remodelling | [93] |
| ↑miRNA-130 | ↓MET, miRNA-221, miRNA-222. | ↑sensitivity to TRAIL | [88] |
| ↑miRNA-200, miRNA-206 | ↓p70S6K1, ↓HIF-1α | ↑sensitivity to cisplatin | [94] |
| Gene | Betweenness | Degree | Adj. p Value | p Value | Fold Change |
|---|---|---|---|---|---|
| TP53 | 0.499325647 | 86 | 0.52935846 | 2.04 × 10−1 | 1.3603568 |
| MDM2 | 0.04074928 | 41 | 0.00026943 | 1.32 × 10−6 | 10.4840794 |
| CDKN1A | 0.02606798 | 40 | 0.00032013 | 1.83 × 10−6 | 10.1108217 |
| PCNA | 0.005309797 | 24 | 0.02632428 | 1.48 × 10−3 | 4.3818849 |
| POLH | 0.0002545446 | 5 | 0.00054359 | 4.25 × 10−6 | 9.1942649 |
| KEGG | Pathway Name | Count in Network | FDR |
|---|---|---|---|
| hsa01524 | Platinum drug resistance | 9 of 70 | 2.58 × 10−5 |
| hsa04210 | Apoptosis | 27 of 135 | 7.38 × 10−23 |
| hsa04310 | Wnt signalling pathway | 11 of 143 | 0.00016 |
| hsa04115 | p53 signalling pathway | 22 of 68 | 9.40 × 10−19 |
| hsa04151 | PI3K-Akt signalling pathway | 21 of 348 | 2.28 × 10−6 |
| hsa04510 | Focal adhesion | 10 of 197 | 0.0049 |
| hsa04530 | Tight junction | 11 of 167 | 0.00048 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kryczka, J.; Kryczka, J.; Czarnecka-Chrebelska, K.H.; Brzeziańska-Lasota, E. Molecular Mechanisms of Chemoresistance Induced by Cisplatin in NSCLC Cancer Therapy. Int. J. Mol. Sci. 2021, 22, 8885. https://doi.org/10.3390/ijms22168885
Kryczka J, Kryczka J, Czarnecka-Chrebelska KH, Brzeziańska-Lasota E. Molecular Mechanisms of Chemoresistance Induced by Cisplatin in NSCLC Cancer Therapy. International Journal of Molecular Sciences. 2021; 22(16):8885. https://doi.org/10.3390/ijms22168885
Chicago/Turabian StyleKryczka, Jolanta, Jakub Kryczka, Karolina H. Czarnecka-Chrebelska, and Ewa Brzeziańska-Lasota. 2021. "Molecular Mechanisms of Chemoresistance Induced by Cisplatin in NSCLC Cancer Therapy" International Journal of Molecular Sciences 22, no. 16: 8885. https://doi.org/10.3390/ijms22168885
APA StyleKryczka, J., Kryczka, J., Czarnecka-Chrebelska, K. H., & Brzeziańska-Lasota, E. (2021). Molecular Mechanisms of Chemoresistance Induced by Cisplatin in NSCLC Cancer Therapy. International Journal of Molecular Sciences, 22(16), 8885. https://doi.org/10.3390/ijms22168885