
Lymphoid Organ Proteomes Identify Therapeutic Efficacy Biomarkers Following the Intracavitary Administration of Curcumin in a Highly Invasive Rat Model of Peritoneal Mesothelioma

 ,
, 
Abstract
1. Introduction
2. Results
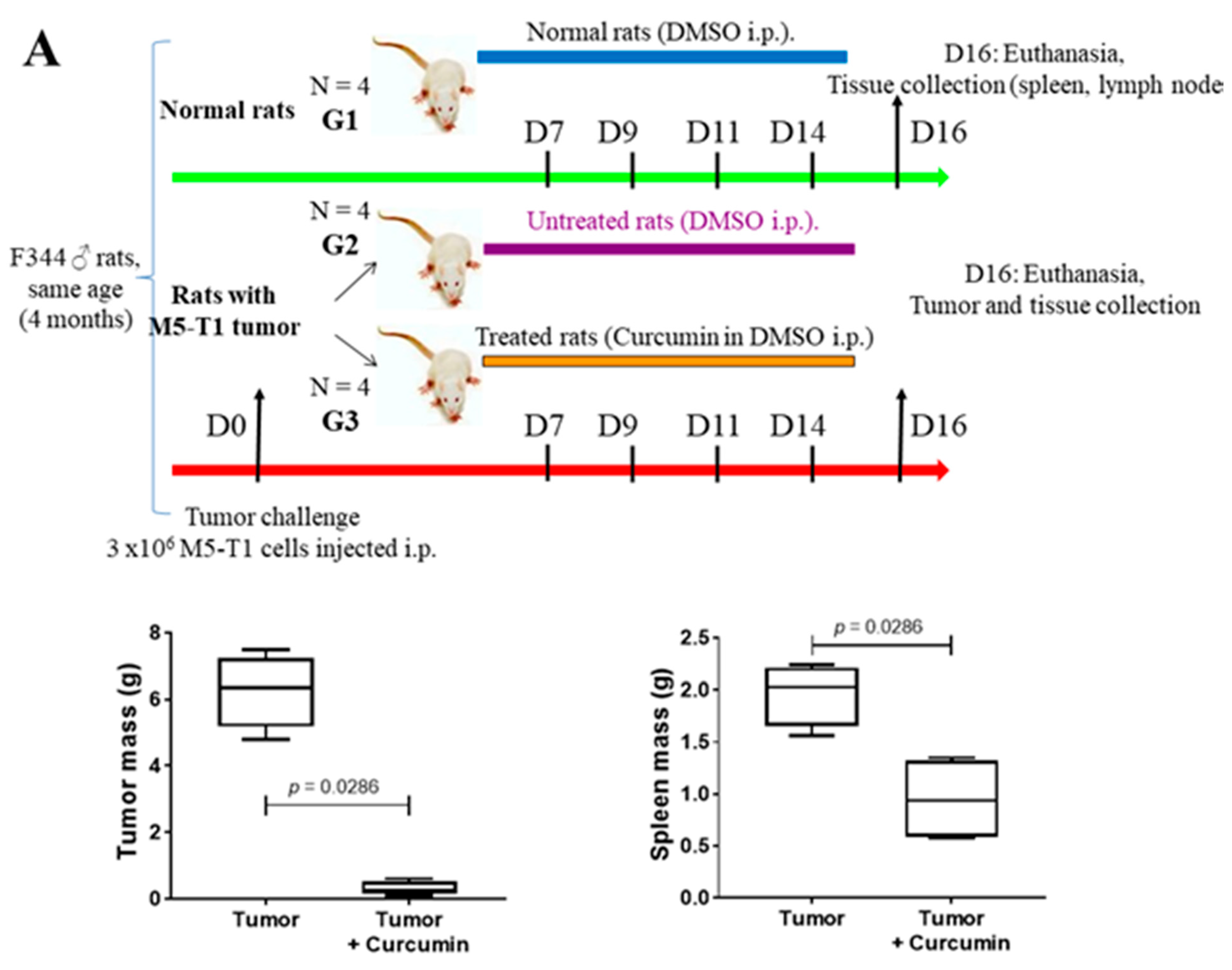
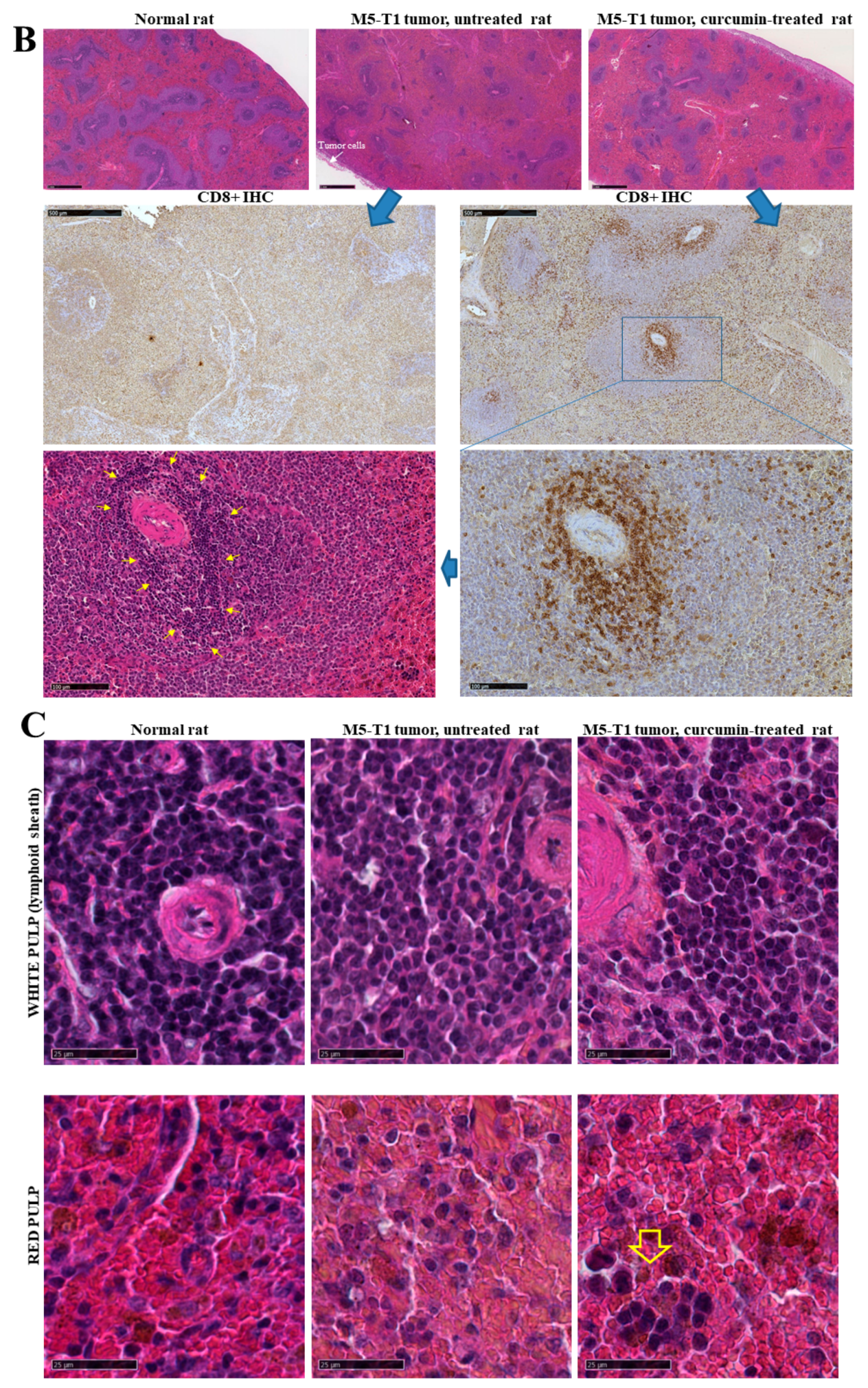
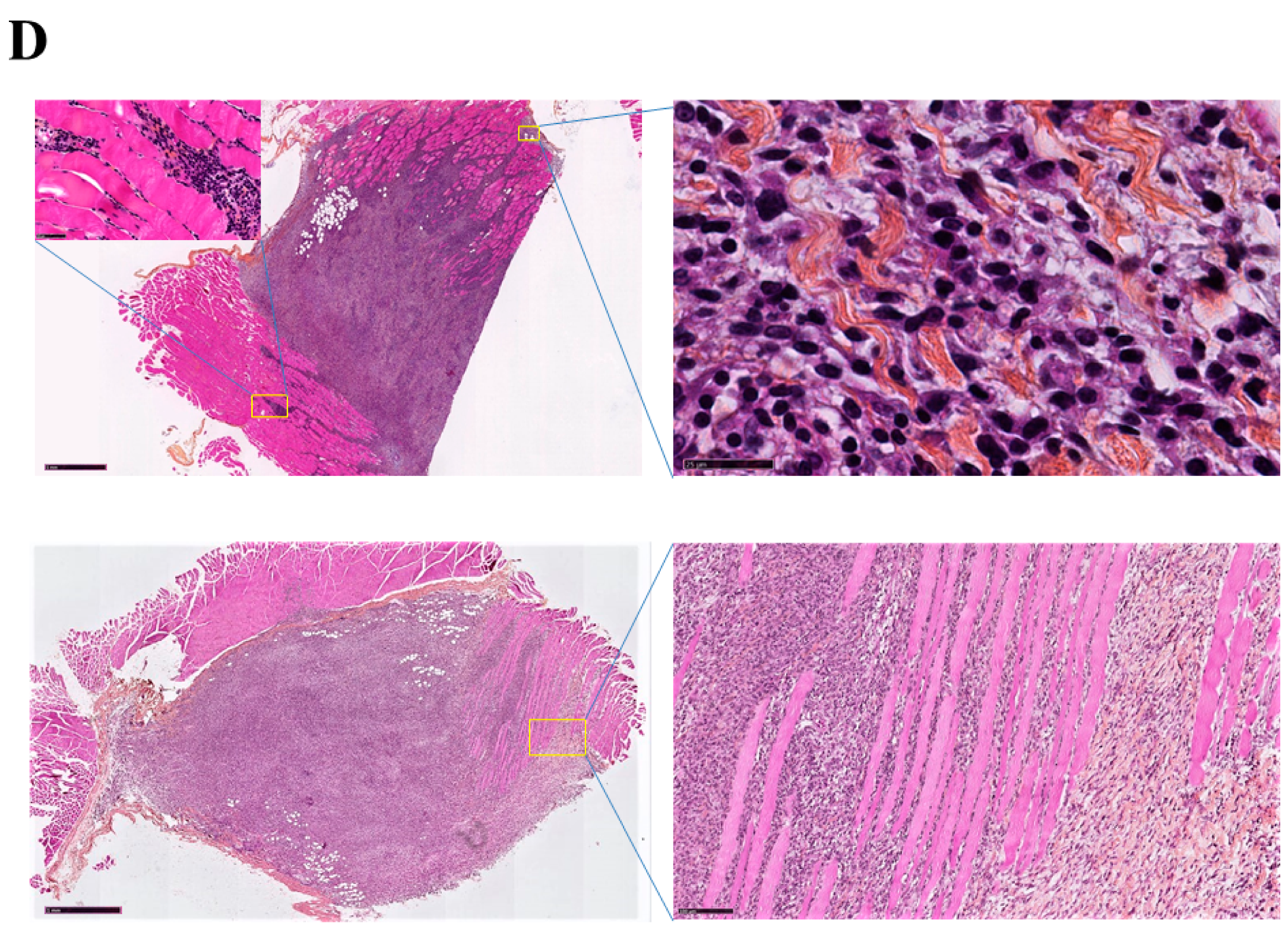
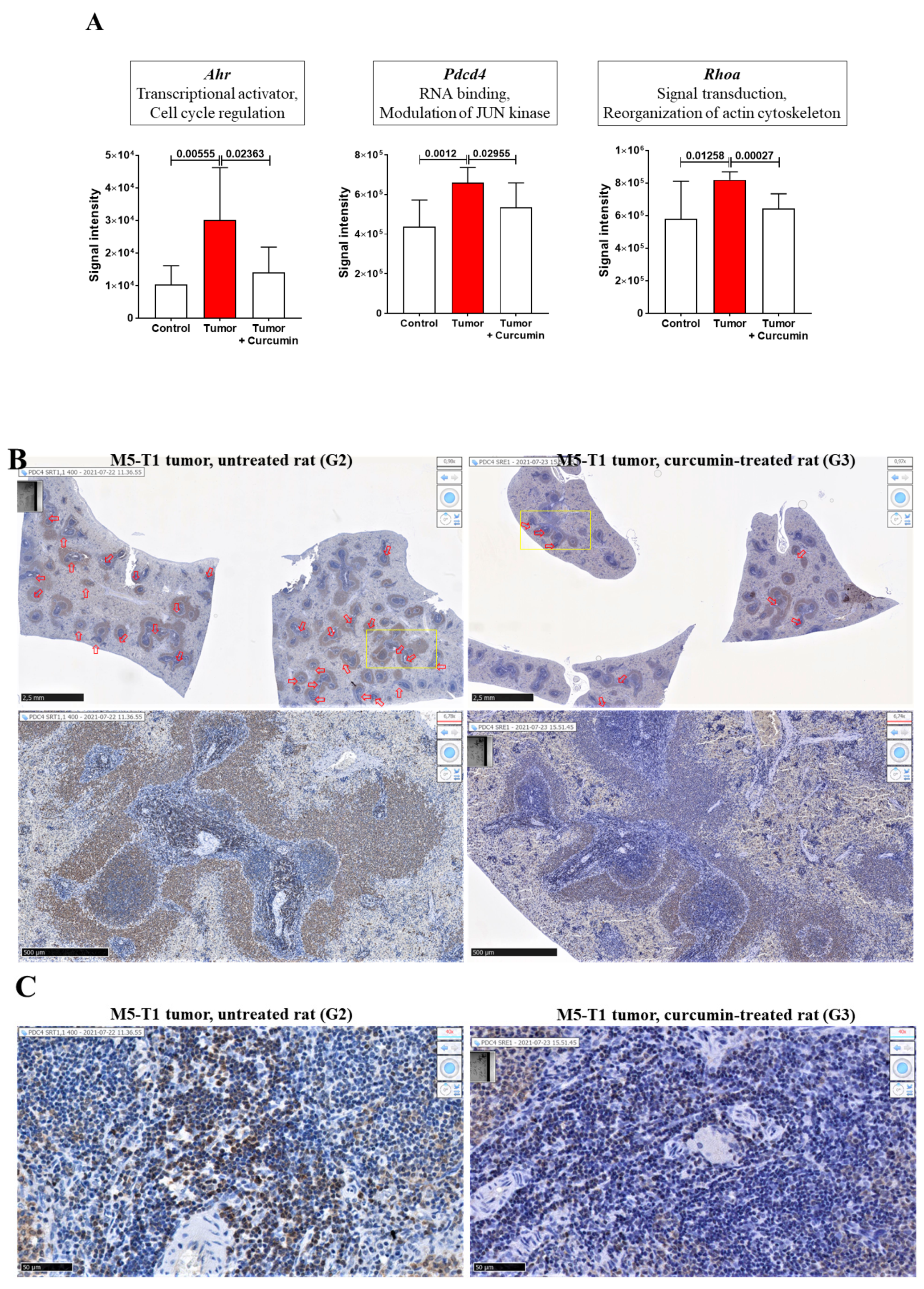
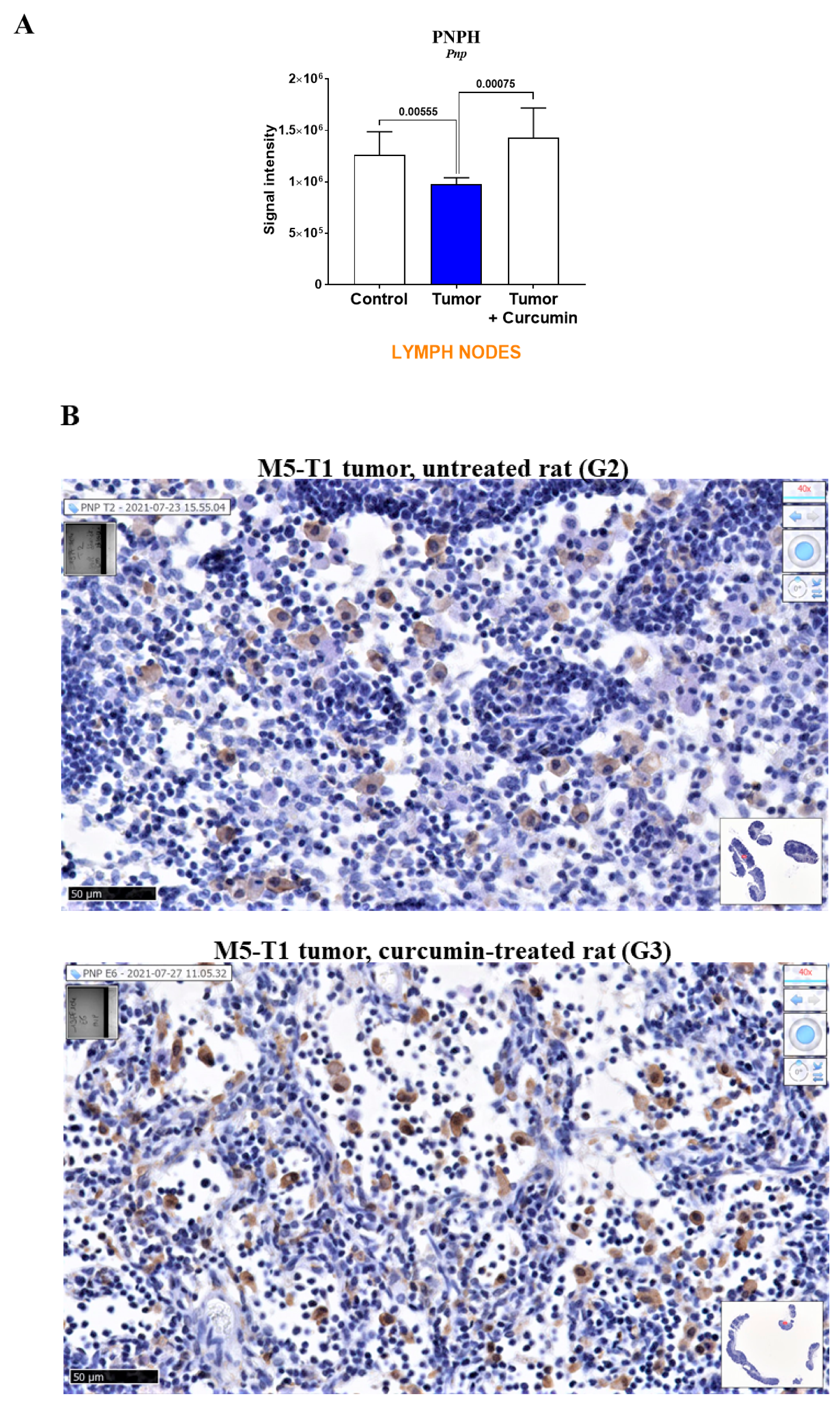
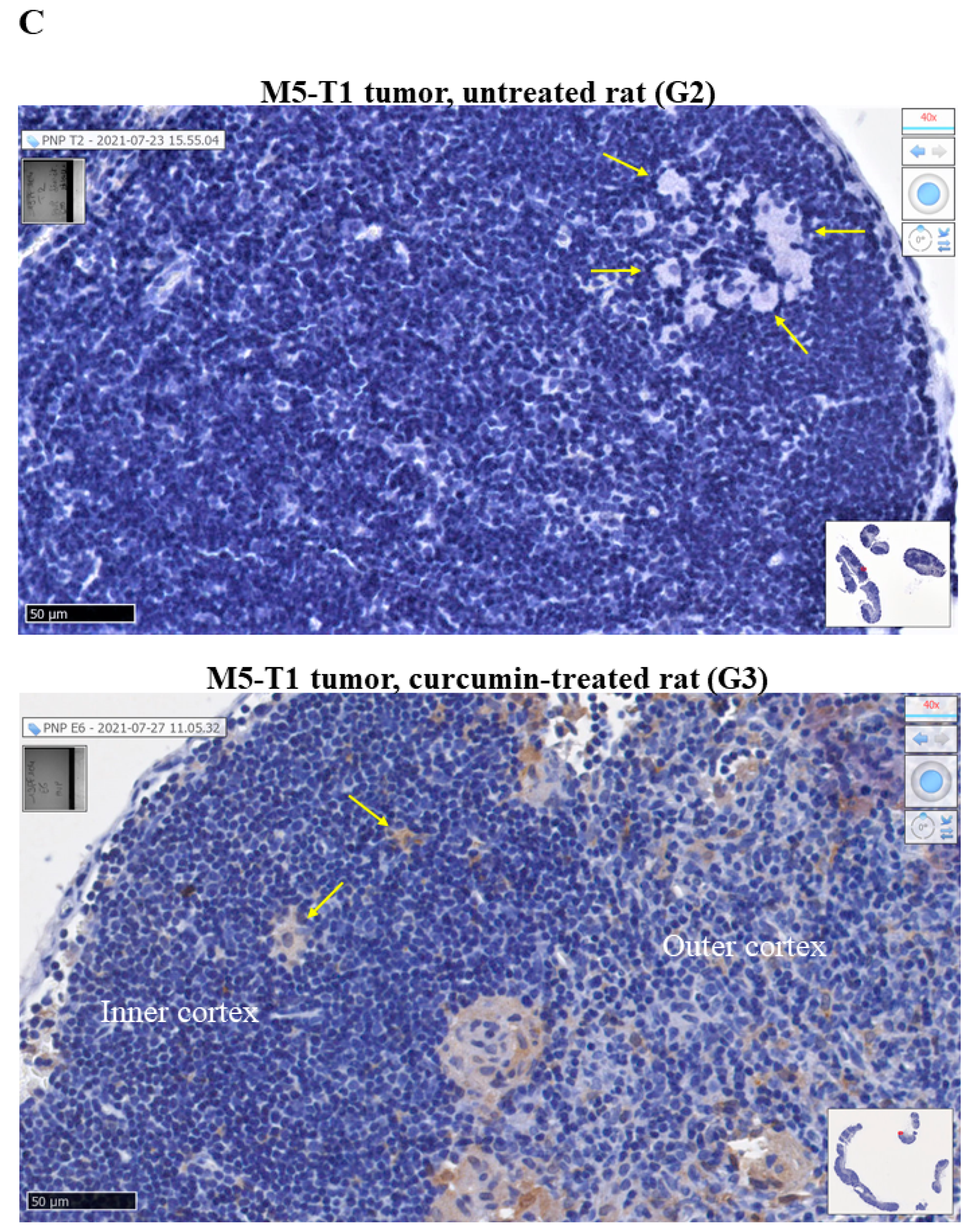
2.1. Histological Features of the Immune Response in the Spleen and in Residual Tumors of Curcumin-Treated Rats
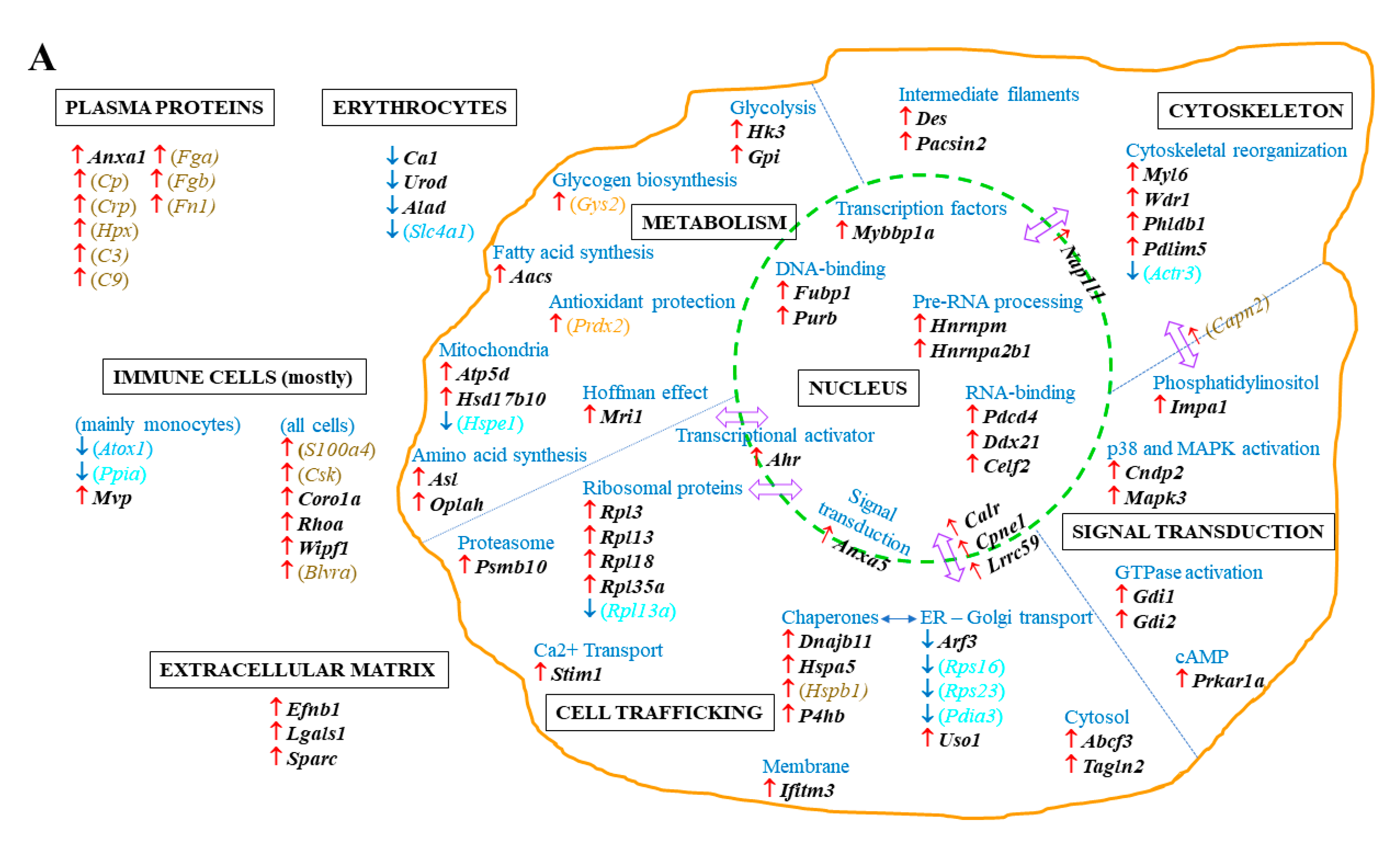
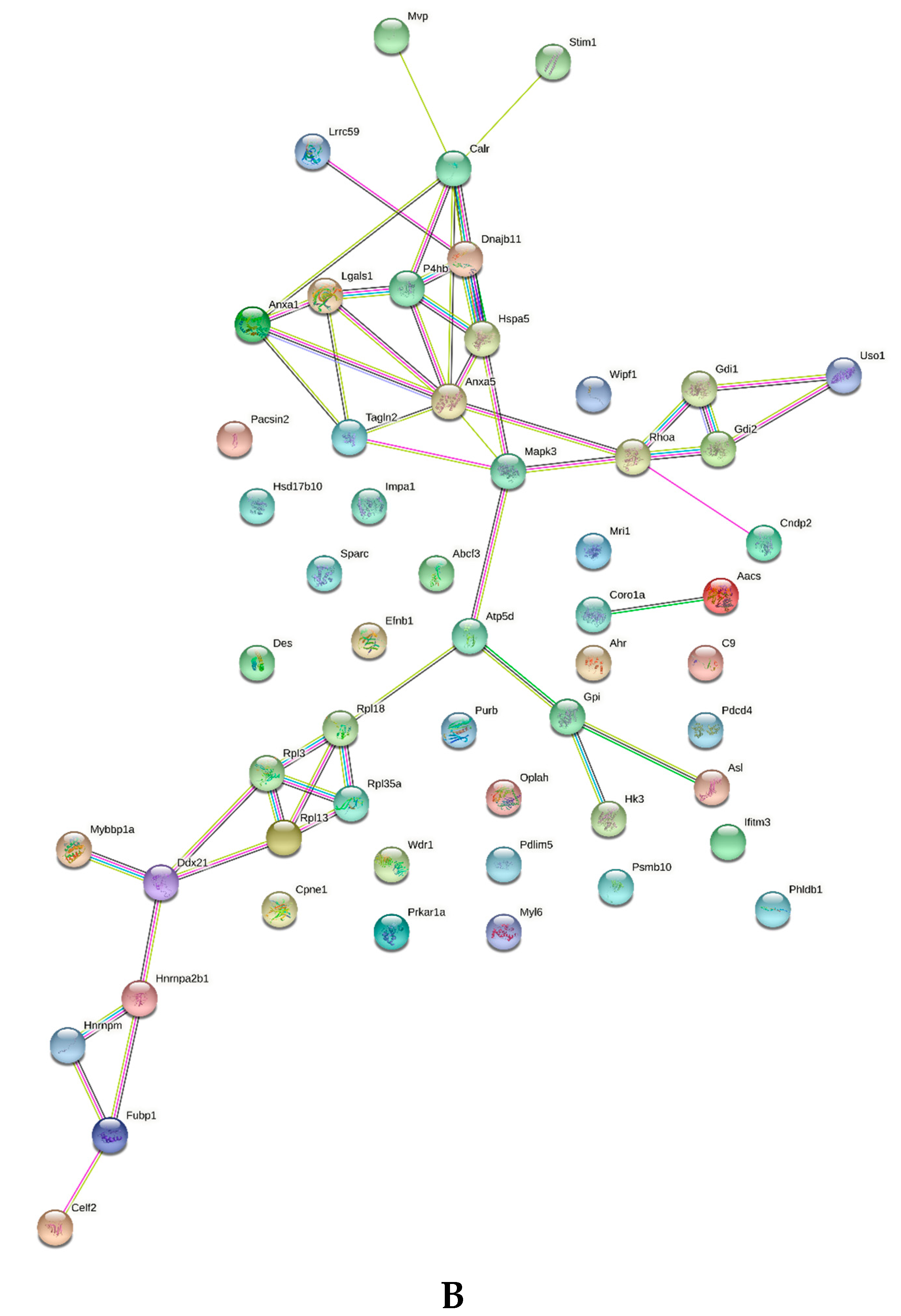
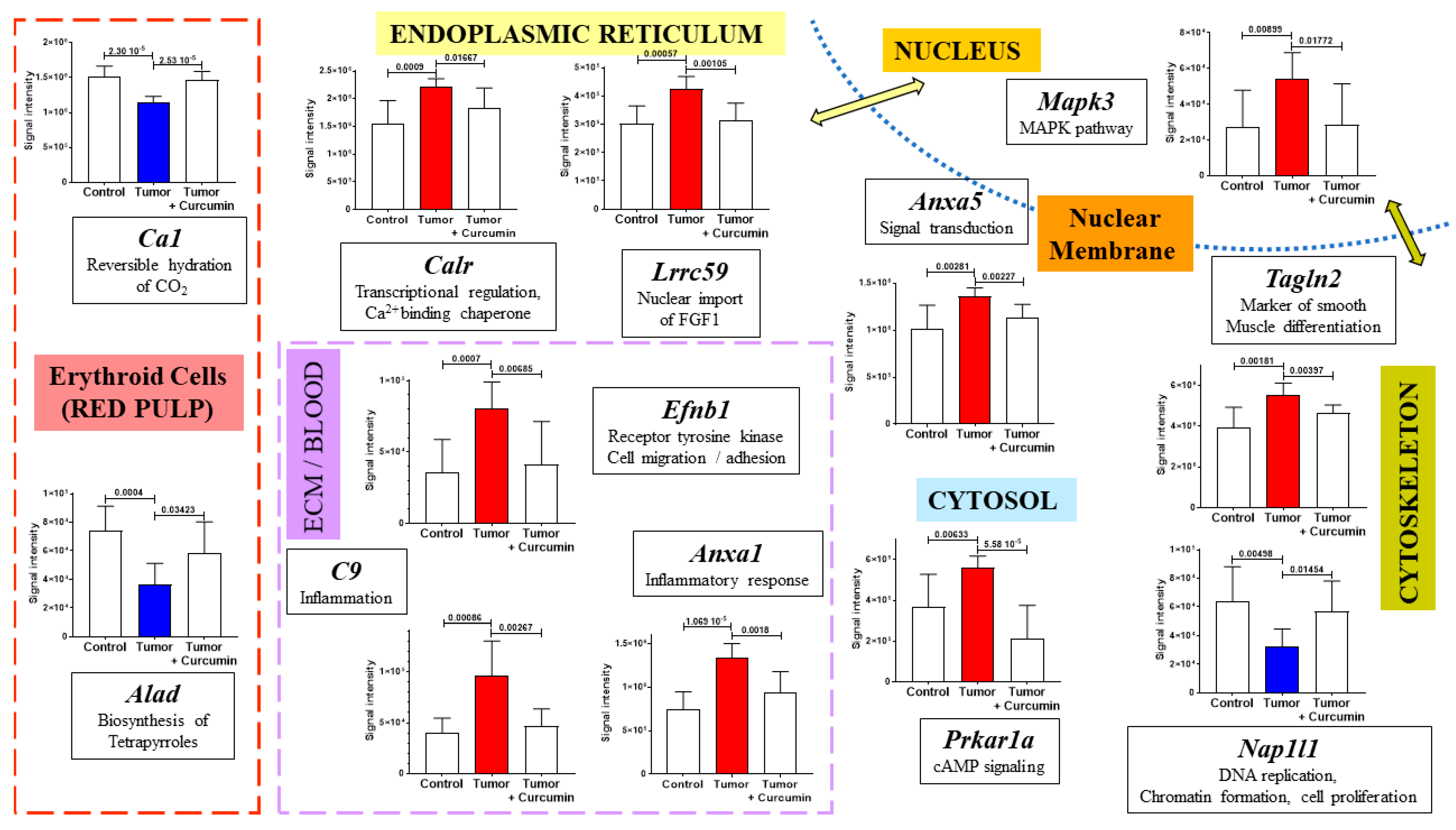
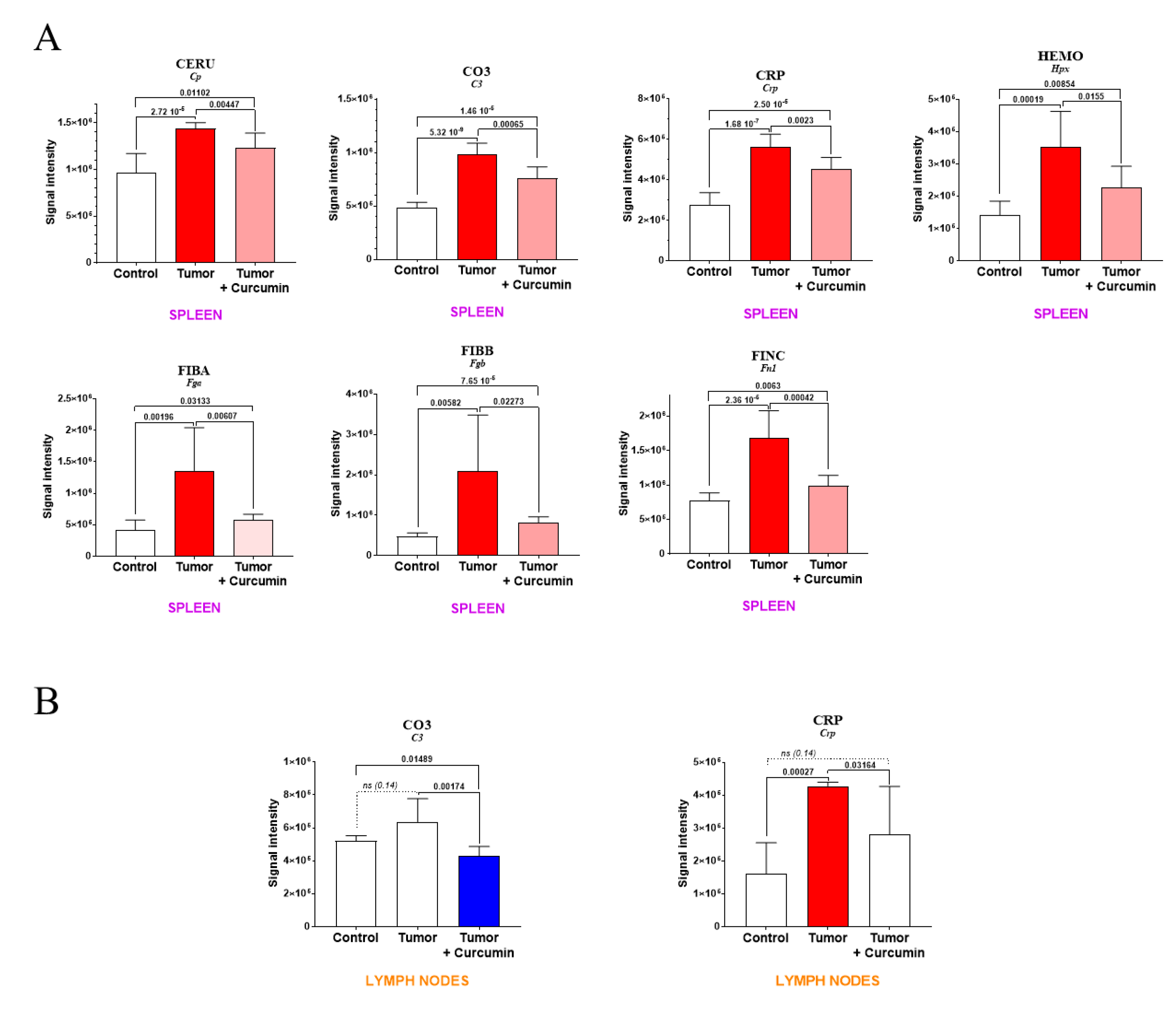
2.2. Spleen Biomarkers of Tumor Progression and Curcumin-Induced Immune Response
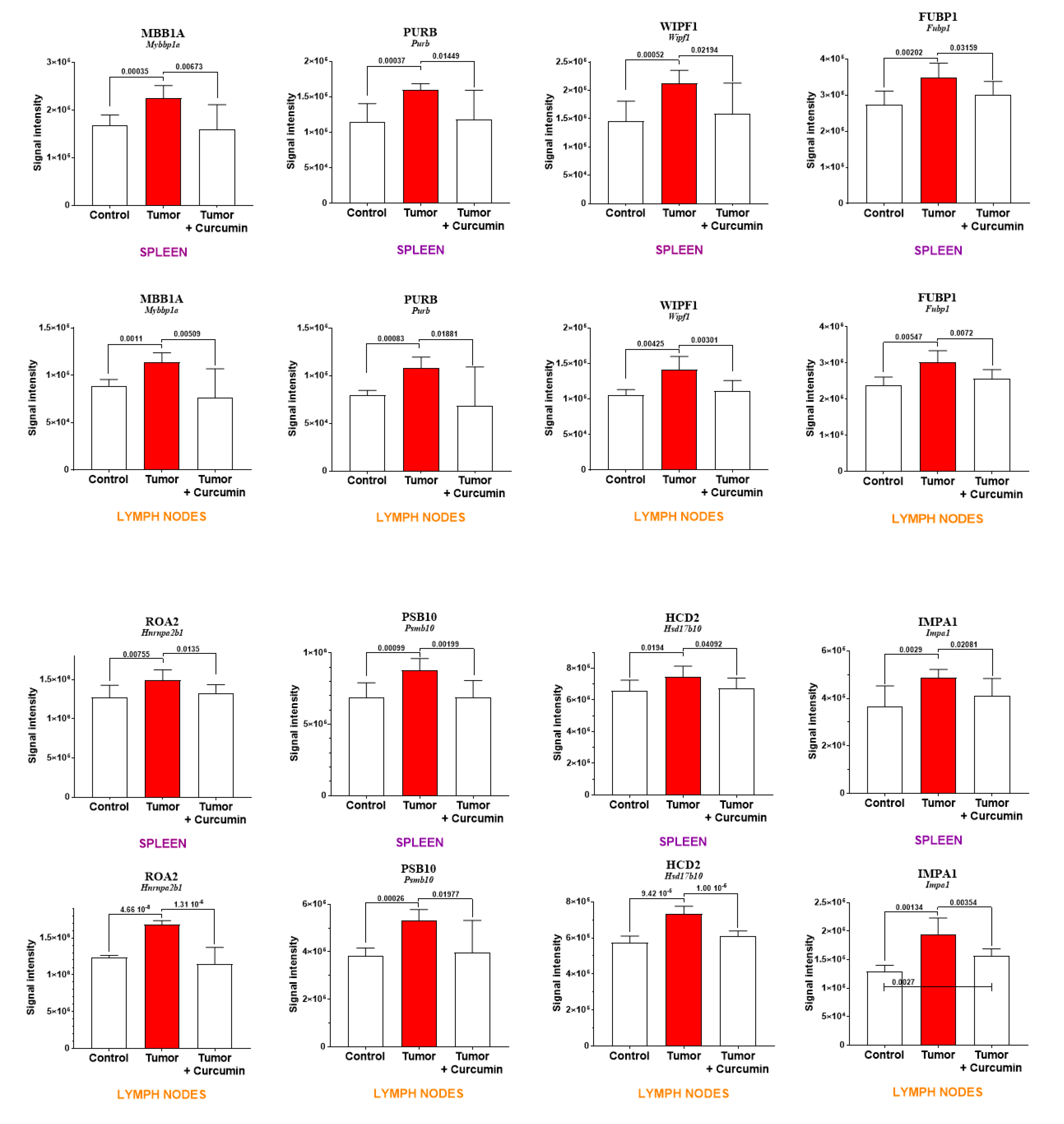
2.3. Common and Specific Biomarkers of Curcumin-Induced Effects in Spleen and Lymph Nodes
2.4. Tumor Progression and Curcumin-Induced Effects on Plasma Biomarkers
3. Discussion
4. Materials and Methods
4.1. Experimental Procedures in Rats
4.2. Histological Analyses
4.3. Proteomic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muniraj, N.; Siddharth, S.; Sharma, D. Bioactive compounds: Multi-targeting silver bullets for preventing and treating breast cancer. Cancers 2019, 11, 1563. [Google Scholar] [CrossRef]
- El-Magboub, A.; Rojsitthisak, P.; Muangnoi, C.; Wichitnithad, W.; Romero, R.; Haworth, I.S. Biological targets and pharmacology of curcumin. In Curcumin Synthesis, Emerging Role in Pain Management and Health Implications; Pouliquen, D.L., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2014; pp. 103–133. [Google Scholar]
- Anighoro, A.; Bajorah, J.; Rastelli, G. Polypharmacology: Challenges and opportunities in drug discovery. J. Med. Chem. 2014, 57, 7874–7887. [Google Scholar] [CrossRef]
- Karuppasamy, R.; Veerappapillai, S.; Maiti, S.; Shin, W.-H.; Kihara, D. Current progress and future perspectives of polypharmacology: From the view of non-small cell lung cancer. Semin. Cancer Biol. 2021, 68, 84–91. [Google Scholar] [CrossRef]
- Weng, W.; Goel, A. Curcumin and colorectal cancer: An update and current perspective on this natural medicine. In Seminars in Cancer Biology; Academic Press: Amsterdam, The Netherland, 2020; pp. 1–14. [Google Scholar] [CrossRef]
- Ryskalin, L.; Biagioni, F.; Busceti, C.L.; Frati, A.; Fornai, F. The multi-faceted effect of curcumin in glioblastoma from rescuing cell clearance to autophagy-independent effects. Molecules 2020, 25, 4839. [Google Scholar] [CrossRef] [PubMed]
- Pouliquen, D.L.; Nawrocki-Raby, B.; Nader, J.; Blandin, S.; Robard, M.; Birembaut, P.; Grégoire, M. Evaluation of intracavitary administration of curcumin for the treatment of sarcomatoid mesothelioma. Oncotarget 2017, 8, 57552–57573. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pouliquen, D.L.; Boissard, A.; Henry, C.; Blandin, S.; Richomme, P.; Coqueret, O.; Guette, C. Curcumin treatment identifies therapeutic targets within biomarkers of liver colonization by highly invasive mesothelioma cells—Potential links with sarcomas. Cancers 2020, 12, 3384. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Gordon, O.N.; Edwards, R.L.; Luis, P.B. Degradation of curcumin: From mechanism to biological implications. J. Agric. Food Chem. 2015, 63, 7606–7614. [Google Scholar] [CrossRef]
- D’Andrea, M.A.; Reddy, G.K. Systemic antitumor effects and abscopal responses in melanoma patients receiving radiation therapy. Oncology 2020, 98, 202–215. [Google Scholar] [CrossRef]
- Bronte, V.; Pittet, M.J. The spleen in local and systemic regulation of immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef]
- Ciolino, H.P.; Daschner, P.J.; Wang, T.T.Y.; Yeh, G.C. Effect of curcumin on the aryl hydrocarbon receptor and cytochrome P450 1A1 in MCF-7 human breast carcinoma cells. Biochem. Pharmacol. 1998, 56, 197–206. [Google Scholar] [CrossRef]
- Rinaldi, A.L.; Morse, M.A.; Fields, H.W.; Rothas, D.A.; Pei, P.; Rodrigo, K.A.; Renner, R.J.; Mallery, S.R. Curcumin activates the aryl hydrocarbon receptor yet significantly inhibits (-)-Benzo(a)pyrene-7R-trans-7, 8-dihydrobiol bioactivation in oral squamous cell carcinoma cells and oral mucosa. Cancer Res. 2002, 62, 5451–5456. [Google Scholar]
- Nishiumi, S.; Yoshida, K.-I.; Ashida, H. Curcumin suppresses the transformation of an aryl hydrocarbon receptor through its phosphorylation. Arch. Biochem. Biophys. 2007, 466, 267–273. [Google Scholar] [CrossRef]
- Choi, H.; Chun, Y.-S.; Shin, Y.J.; Ye, S.K.; Kim, M.-S.; Park, J.-W. Curcumin attenuates cytochrome P450 induction in response to 2,3,7,8-tetrachlorodibenzo-p-dioxin by ROS-dependently degrading AhR and ARNT. Cancer Sci. 2008, 99, 2518–2524. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Gupta, S.; Maru, G.B. Dietary curcumin modulates transcriptional regulators of phase I and phase II enzymes in benzo[a]pyrene-treated mice: Mechanism of its anti-initiating action. Carcinogenesis 2008, 29, 1022–1032. [Google Scholar] [CrossRef]
- Ciftci, O.; Tanyildizi, S.; Godekmerdan, A. Protective effect of curcumin on immune system and body weight gain on rats intoxicated with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Immunopharmacol. Immunotoxicol. 2010, 32, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Mishra, M.; Sharma, A.; Shukla, A.K.; Mudiam, M.K.R.; Patel, D.K.; Dam, K.R.; Chowdhuri, D.K. Genotoxicity and apoptosis in Drosophila melanogaster exposed to benzene, toluene and xylene: Attenuation by quercetin and curcumin. Toxicol. Appl. Pharmacol. 2011, 253, 14–30. [Google Scholar] [CrossRef]
- Mohammadi-Bardbori, A.; Akbarizadeh, A.R.; Delju, F.; Rannug, A. Chromatin remodeling by curcumin alters endogenous aryl hydrocarbon receptor signaling. Chem. Biol. Interact. 2016, 252, 19–27. [Google Scholar] [CrossRef]
- Nakai, R.; Fukuda, S.; Kawase, M.; Yamashita, Y.; Ashida, H. Curcumin and its derivatives inhibit 2,3,7,8-tetrachloro-dibenzo-p-dioxin-induced expression of drug metabolizing enzymes through aryl hydrocarbon receptor-mediated pathway. Biosci. Biotechnol. Biochem. 2018, 82, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Kam, A.; Li, K.M.; Razmovski-Naumovski, V.; Nammi, S.; Chan, K.; Grau, G.E.; Li, G.Q. Curcumin reduces tumour necrosis factor-enhanced annexin V-positive microparticle release in human vascular endothelial cells. J. Pharm. Pharm. Sci. 2015, 18, 424. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.; Chaves, L.; Eadon, M.T.; Chang, A.; Quigg, R.J.; Alexander, J.J. Curcumin alleviates immune-complex-mediated glomerulonephritis in factor-H-deficient mice. Immunology 2013, 139, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Ravindranathan, P.; Pasham, D.; Balaji, U.; Cardenas, J.; Gu, J.; Toden, S.; Goel, A. A combination of curcumin and oligomeric proanthocyanidins offer superior anti-tumorigenic properties in colorectal cancer. Sci. Rep. 2018, 8, 13869. [Google Scholar] [CrossRef]
- Rabinovitch, G.A.; Alonso, C.R.; Sotomayor, C.E.; Durand, S.; Bocco, J.L.; Riera, C.M. Molecular mechanisms implicated in galectin-1-induced apoptosis: Activation of the AP-1 transcription factor and downregulation of Bcl-2. Cell Death Differ. 2000, 7, 747–753. [Google Scholar] [CrossRef]
- Brandt, B.; Abou-Eladab, E.F.; Tiedge, M.; Walzel, H. Role of the JNK/c-Jun/AP-1 signaling pathway in galectin-1-induced T-cell death. Cell Death Dis. 2010, 1, e23. [Google Scholar] [CrossRef]
- Thiyagarajan, S.; Thirumalai, K.; Nirmala, S.; Biswas, J.; Krishnakumar, S. Effect of curcumin on lung resistance-related protein (LRP) in retinoblastoma cells. Curr. Eye Res. 2009, 34, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Mudduluru, G.; George-William, J.N.; Muppala, S.; Asangani, I.A.; Kumarswamy, R.; Nelson, L.D.; Allgayer, H. Curcumin regulates miR-21 expression and inhibits invasion and metastasis in colorectal cancer. Biosci. Rep. 2011, 31, 185–197. [Google Scholar] [CrossRef]
- Yang, C.H.; Yue, J.; Sims, M.; Pfeffer, L.M. The curcumin analog EF24 targets NF-kB and miRNA-21, and has potent anticancer activity in vitro and in vivo. PLoS ONE 2013, 8, e71130. [Google Scholar] [CrossRef]
- Chen, J.; Xu, T.; Chen, C. The critical roles of miR-21 in anti-cancer effects of curcumin. Ann. Transl. Med. 2015, 3, 330. [Google Scholar] [CrossRef]
- Lelli, D.; Pedone, C.; Sahebkar, A. Curcumin and treatment of melanoma: The potential role of microRNAs. Biomed. Pharmacother. 2017, 88, 832–834. [Google Scholar] [CrossRef]
- Tan, X.; Kim, G.; Lee, D.; Oh, J.; Kim, M.; Piao, C.; Lee, J.; Lee, M.S.; Jeong, J.H.; Lee, M. A curcumin-loaded polymeric micelle as a carrier of a microRNA-21 antisense-oligonucleotide for enhanced anti-tumor effects in a glioblastoma animal model. Biomater. Sci. 2018, 6, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Gahnbari, M.; Tasbandi, A.; Sahebkar, A. Regulation of microRNA-21 expression by natural products in cancer. Phytother. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, M.; Luo, Z.; Zhang, W.; Zhu, D.; Lu, Y.; Wu, J.; Yao, X. Protective effect of curcumin against irinotecan-induced intestinal mucosal injury via attenuation of NF-kB activation, oxidative stress and endoplasmic reticulum stress. Int. J. Oncol. 2019, 54, 1376–1386. [Google Scholar] [CrossRef]
- Wang, R.; Li, J.; Zhao, Y.; Li, Y.; Yin, L. Investigating the therapeutic potential and mechanism of curcumin in breast cancer based on RNA sequencing and bioinformatics analysis. Breast Cancer 2018, 25, 206–212. [Google Scholar] [CrossRef]
- Zhang, P.; Bai, H.; Liu, G.; Wang, H.; Chen, F.; Zhang, B.; Zeng, P.; Wu, C.; Peng, C.; Huang, C.; et al. MicroRNA-33b, upregulated by EF24, a curcumin analog, suppresses the epithelial-to-mesenchymal transition (EMT) and migratory potential of melanoma cells by targeting HMGA2. Toxicol. Lett. 2015, 234, 151–161. [Google Scholar] [CrossRef]
- Qin, L.; Qin, J.; Zhen, X.; Yang, Q.; Huang, L. Curcumin protects against hepatic stellate cells activation and migration by inhibiting the CXCL12/CXCR4 biological axis in liver fibrosis: A study in vitro and in vivo. Biomed. Pharmacother. 2018, 101, 599–607. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, X.; Xu, Y.; Shen, Z.; Cheng, H.; Cheng, F.; Liu, X.; Wang, R. RhoA/Rho-kinase triggers epithelial-mesenchymal transition in mesothelial cells and contributes to the pathogenesis of dialysis-related peritoneal fibrosis. Oncotarget 2018, 9, 14397–14412. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.; Kemmerling, U.; Aguayo, F.; Bleak, T.C.; Muñoz, J.P.; Calaf, G.M. Curcumin rescues breast cells from epithelial-mesenchymal transition and invasion induced by anti-miR-34a. Int. J. Oncol. 2020, 56, 480–493. [Google Scholar] [CrossRef]
- Killian, P.H.; Kronski, E.; Michalik, K.M.; Barbieri, O.; Astigiano, S.; Sommerhoff, C.; Pfeffer, U.; Nerlich, A.G.; Bachmeier, B. Curcumin inhibits prostate cancer metastasis in vivo by targeting the inflammatory cytokines CXCL1 and -2. Carcinogenesis 2012, 33, 2507–2519. [Google Scholar] [CrossRef]
- Shin, D.H.; Nam, J.H.; Lee, E.S.; Zhang, Y.; Kim, S.J. Inhibition of Ca2+ release-activated Ca2+ channel (CRAC) by curcumin and caffeic acid phenethyl ester (CAPE) via electrophilic addition to a cysteine residue of Orai1. Biochem. Biophys. Res. Comm. 2012, 428, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhuang, Z.; Su, Q.; He, J.; Li, H. Curcumin has anti-proliferative and pro-apoptotic effects on tongue cancer in vitro: A study with bioinformatics analysis and in vitro experiments. Drug Des. Devel. Ther. 2020, 14, 509–518. [Google Scholar] [CrossRef]
- Taverna, S.; Fontana, S.; Monteleone, F.; Pucci, M.; Saieva, L.; De Caro, V.; Cardinale, V.G.; Giallombardo, M.; Vicario, E.; Rolfo, C.; et al. Curcumin modulates chronic myelogenous leukemia exosomes composition and affects angiogenic phenotype via exosomal miR-21. Oncotarget 2016, 7, 30420–30439. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.P.; Jin, H.; Shi, J.D.; Zhu, L.M.; Suo, Y.; Lu, G.; Liu, A.; Wang, T.C.; Yang, C.S. Curcumin induces the differentiation of myeloid-derived suppressor cells and inhibits their interaction with cancer cells and related tumor growth. Cancer Prev. Res. 2012, 5, 205–215. [Google Scholar] [CrossRef]
- Dudekula, K.; Le Bihan, T. Data from quantitative label free proteomics analysis of rat spleen. Data Brief. 2015, 8, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Frankel, L.B.; Christoffersen, N.R.; Jacobsen, A.; Lindow, M.; Krogh, A.; Lund, A.H. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J. Biol. Chem. 2008, 283, 1026–1033. [Google Scholar] [CrossRef]
- O’Connor, K.L.; Chen, M. Dynamic functions of RhoA in tumor cell migration and invasion. Small GTPases 2013, 4, 141–147. [Google Scholar] [CrossRef]
- Greenstein, J.D.; Peake, P.W.; Charlesworth, J.A. The kinetics and distribution of C9 and SC5b-9 in vivo: Effects of complement activation. Clin. Exp. Immunol. 1995, 100, 40–46. [Google Scholar] [CrossRef]
- O’Brien, R.M.; Cannon, A.; Reynolds, J.V.; Lysaght, J.; Lynam-Lennon, N. Complement in tumourigenesis and the response to cancer therapy. Cancers 2021, 13, 1209. [Google Scholar] [CrossRef]
- Tanaka, T.; Hozumi, Y.; Iino, M.; Goto, K. NAP1L1 regulates NF-κB signaling pathway acting on anti-apoptotic Mcl-1 gene expression. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1759–1768. [Google Scholar] [CrossRef]
- Çevik, R.E.; Cesarec, M.; Da Silva Filipe, A.; Licastro, D.; McLauchlan, J.; Marcello, A. Hepatitis C virus NS5A targets nucleosome assembly protein NAP1L1 to control the innate cellular response. J. Virol. 2017, 91, e00880-17. [Google Scholar] [CrossRef]
- Nishihara, H.; Hwang, M.; Kizaba-Kondoh, S.; Eckmann, L.; Insel, P.A. Cyclic AMP promotes cAMP-responsive element-binding protein-dependent induction of cellular inhibitor of apoptosis protein-2 and suppresses apoptosis of colon cancer cells through ERK1/2 and p38 MAPK. J. Biol. Chem. 2004, 279, 26176–26183. [Google Scholar] [CrossRef]
- Feng, R.; Ye, J.; Zhou, C.; Qi, L.; Fu, Z.; Yan, B.; Liang, Z.; Li, R.; Zhai, W. Calreticulin down-regulation inhibits the cell growth, invasion and cell cycle progression of human hepatocellular carcinoma cells. Diagn. Pathol. 2015, 10, 149. [Google Scholar] [CrossRef] [PubMed]
- Fucikova, J.; Kasikova, L.; Truxova, I.; Laco, J.; Skapa, P.; Ryska, A.; Spisek, R. Relevance of the chaperone-like protein calreticulin for the biological behavior and clinical outcome of cancer. Immunol. Lett. 2018, 193, 25–34. [Google Scholar] [CrossRef]
- Schcolnik-Cabrera, A.; Oldak, B.; Juárez, M.; Cruz-Rivera, M.; Flisser, A.; Mendlovic, F. Calreticulin in phagocytosis and cancer: Opposite roles in immune response outcomes. Apoptosis 2019, 24, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Pouliquen, D.L.; Boissard, A.; Coqueret, O.; Guette, C. Biomarkers of tumor invasiveness in proteomics (Review). Int. J. Oncol. 2020, 57, 409–432. [Google Scholar] [CrossRef] [PubMed]
- Nader, J.S.; Guillon, J.; Petit, C.; Boissard, A.; Franconi, F.; Blandin, S.; Lambot, S.; Grégoire, M.; Verrièle, V.; Nawrocki-Raby, B.; et al. S100A4 is a biomarker of tumorigenesis, EMT, invasion, and colonization of host organs in experimental malignant mesothelioma. Cancers 2020, 12, 939. [Google Scholar] [CrossRef]
- Shiuan, E.; Chen, J. Eph receptor tyrosine kinases in tumor immunity. Cancer Res. 2016, 76, 6452–6457. [Google Scholar] [CrossRef]
- Iwasaki, K.; Ninomiya, R.; Shin, T.; Nomura, T.; Kajiwara, T.; Hijiya, N.; Moriyama, M.; Mimata, H.; Hamada, F. Chronic hypoxia-induced slug promotes invasive behavior of prostate cancer cells by activating expression of ephrin-B1. Cancer Sci. 2018, 109, 3159–3170. [Google Scholar] [CrossRef]
- Perrera, C.; Colombo, R.; Valsasina, B.; Carpinelli, P.; Troiani, S.; Modugno, M.; Gianellini, L.; Cappella, P.; Isacchi, A.; Moll, J.; et al. Identification of Myb-binding protein 1A (MYBBP1A) as a novel substrate for Aurora B kinase. J. Biol. Chem. 2010, 285, 11775–11785. [Google Scholar] [CrossRef]
- George, B.; Horn, D.; Bayo, P.; Zaoui, K.; Flechtenmacher, C.; Grabe, N.; Plinkert, P.; Krizhanovsky, V.; Hess, J. Regulation and function of Myb-binding protein 1A (MYBBP1A) in cellular senescence and pathogenesis of head and neck cancer. Cancer Lett. 2015, 358, 191–199. [Google Scholar] [CrossRef]
- Weng, X.; Wu, J.; Lv, Z.; Peng, C.; Chen, J.; Zhang, C.; He, B.; Tong, R.; Hu, W.; Ding, C.; et al. Targeting Mybbp1a suppresses HCC progression via inhibiting IGF1/AKT pathway by CpG islands hypo-methylation dependent promotion of IGFBP5. EBioMedicine 2019, 44, 225–236. [Google Scholar] [CrossRef]
- Nahálkova, J. The molecular mechanisms associated with PIN7, a protein-protein interaction network of seven pleiotropic proteins. J. Theor. Biol. 2020, 487, 110124. [Google Scholar] [CrossRef]
- Kelm, R.J., Jr.; Lamba, G.S.; Levis, J.E.; Holmes, C.E. Characterization of purine-rich element binding protein B as a novel biomarker in acute myelogenous leukemia prognostication. J. Cell Biochem. 2018, 119, 2073–2083. [Google Scholar] [CrossRef]
- Jang, M.; Park, B.C.; Kang, S.; Chi, S.-W.; Cho, S.; Chung, S.J.; Lee, S.C.; Bae, K.-H.; Park, S.G. Far upstream element-binding protein-1, a novel caspase substrate, acts as a cross-talker between apoptosis and the c-myc oncogene. Oncogene 2009, 28, 1529–1536. [Google Scholar] [CrossRef]
- Malz, M.; Weber, A.; Singer, S.; Riehmer, V.; Bissinger, M.; Riener, M.-O.; Longerich, T.; Soll, C.; Vogel, A.; Angel, P.; et al. Overexpression of far upstream element binding proteins: A mechanism regulating proliferation and migration in liver cancer cells. Hepatology 2009, 50, 1130–1139. [Google Scholar] [CrossRef]
- Debaize, L.; Troadec, M.-B. The master regulator FUBP1: Its emerging role in normal cell function and malignant development. Cell. Mol. Life Sci. 2019, 76, 259–281. [Google Scholar] [CrossRef]
- Ma, C.; Huang, Z.; Wu, Z.; Di, C.; Lin, X.; Huang, M.; Hong, H.; Yin, H. Overexpression of FUBP1 is associated with human cervical carcinoma development and prognosis. Life Sci. 2021, 269, 119098. [Google Scholar] [CrossRef]
- García, E.; Jones, G.E.; Machesky, L.M.; Antón, I.M. WIP: WASP-interacting proteins at invadopodia and podosomes. Eur. J. Cell Biol. 2012, 91, 869–877. [Google Scholar] [CrossRef]
- Gryaznova, T.; Kropyvko, S.; Burdyniuk, M.; Gubar, O.; Kryklyva, V.; Tsyba, L.; Rynditch, A. Intersectin adaptor proteins are associated with actin-regulating protein WIP in invadopodia. Cell. Signal. 2015, 27, 1499–1508. [Google Scholar] [CrossRef]
- Pan, Y.; Lu, F.; Xiong, P.; Pan, M.; Zhang, Z.; Lin, X.; Pan, M.; Huang, H. WIPF1 antagonizes the tumor suppressive effect of miR-141/200c and is associated with poor survival in patients with PDAC. J. Exp. Clin. Cancer Res. 2018, 37, 167. [Google Scholar] [CrossRef]
- Ramesh, N.; Antón, I.M.; Martínez-Quiles, N.; Geha, R.S. Waltzing with WASP. Trends Cell Biol. 1999, 9, 15–19. [Google Scholar] [CrossRef]
- Groot, M.; Lee, H. Sorting mechanisms for microRNAs into extracellular vesicles and their associated diseases. Cells 2020, 9, 1044. [Google Scholar] [CrossRef]
- Fabbiano, F.; Corsi, J.; Gurrieri, E.; Trevisan, C.; Notarangelo, M.; D’Agostino, V.G. RNA packaging into extracellular vesicles: An orchestra of RNA-binding proteins? J. Extracell. Vesicles 2020, 10, e12043. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-W.; Wang, P.; Liu, J.-Q.; Zhang, H.; Xi, W.-D.; Jia, X.-H.; Wang, K.-K. Coordinated regulation of the immunoproteasome subunits by PML/RAR and PU.1 in acute promyelocytic leukemia. Oncogene 2014, 33, 2700–2708. [Google Scholar] [CrossRef] [PubMed]
- Morosov, A.V.; Karpov, V.L. Proteasomes and several aspects of their heterogeneity relevant to cancer. Front. Oncol. 2019, 9, 761. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Gene | Full Name | G2/G1 | G3/G2 | G3/G1 |
|---|---|---|---|---|---|
| AACS | Aacs | Acetoacetyl-CoA synthetase | ↑ p = 0.01991 | ↓ p = 0.02754 | ns (p = 0.69) |
| ABCF3 | Abcf3 | ATP-binding cassette sub-family F member 3 | ↑ p = 0.0423 | ↓ p = 0.0127 | ns (p = 0.21) |
| AHR | Ahr | Aryl hydrocarbon receptor | ↑ p = 0.00555 | ↓ p = 0.02363 | ns (p = 0.29) |
| ANXA1 | Anxa1 | Annexin A1 | ↑ p = 1.69 10−5 | ↓ p = 0.0018 | ns (p = 0.11) |
| ANXA5 | Anxa5 | Annexin A5 | ↑ p = 0.00281 | ↓ p = 0.00227 | ns (p = 0.27) |
| ARLY | Asl | Argininosuccinate lyase | ↑ p = 0.01787 | ↓ p = 0.00459 | ns (p = 0.65) |
| ATPD | Atp5d | ATP synthase subunit delta, mitochondrial | ↑ p = 0.03331 | ↓ p = 0.00153 | ns (p = 0.53) |
| CALR | Calr | Calreticulin | ↑ p = 0.0009 | ↓ p = 0.01667 | ns (p = 0.17) |
| CELF2 | Celf2 | CUGBP Elav-like family member 2 | ↑ p = 0.04365 | ↓ p = 0.03967 | ns (p = 0.25) |
| CNDP2 | Cndp2 | Cytosolic non-specific dipeptidase | ↑ p = 0.01447 | ↓ p = 0.02765 | ns (p = 0.16) |
| CO9 | C9 | Complement component C9 | ↑ p = 0.00086 | ↓ p = 0.00267 | ns (p = 0.45) |
| COR1A | Coro1a | Coronin-1A | ↑ p = 0.00729 | ↓ p = 0.00078 | ns (p = 0.79) |
| CPNE1 | Cpne1 | Copine-1 | ↑ p = 0.02077 | ↓ p = 0.00269 | ns (p = 0.35) |
| DDX21 | Ddx21 | Nucleolar RNA helicase 2 | ↑ p = 0.00579 | ↓ p = 0.02476 | ns (p = 0.22) |
| DESM | Des | Desmin | ↑ p = 0.01363 | ↓ p = 0.01693 | ns (p = 0.51) |
| DJB11 | Dnajb11 | DnaJ homolog subfamily B member 11 | ↑ p = 0.02069 | ↓ p = 0.00836 | ns (p = 0.85) |
| EFNB1 | Efnb1 | Ephrin-B1 | ↑ p = 0.0007 | ↓ p = 0.00685 | ns (p = 0.67) |
| FUBP1 | Fubp1 | Far upstream element-binding protein 1 | ↑ p = 0.00202 | ↓ p = 0.03159 | ns (p = 0.14) |
| G6PI | Gpi | Glucose-6-phosphate isomerase | ↑ p = 0.03101 | ↓ p = 0.00557 | ns (p = 0.07) |
| GDIA | Gdi1 | Rab GDP dissociation inhibitor alpha | ↑ p = 0.00306 | ↓ p = 0.01494 | ns (p = 0.23) |
| GDIB | Gdi2 | Rab GDP dissociation inhibitor beta | ↑ p = 0.01183 | ↓ p = 0.0371 | ns (p = 0.34) |
| GRP78 | Hspa5 | 78kDa glucose-regulated protein | ↑ p = 0.00475 | ↓ p = 0.00064 | ns (p = 0.21) |
| HCD2 | Hsd17b10 | 3-hydroxyacyl-CoA dehydrogenase type-2 | ↑ p = 0.0194 | ↓ p = 0.04092 | ns (p = 0.70) |
| HNRPM | Hnrnpm | Heterogeneous nuclear ribonucleoprotein M | ↑ p = 0.00659 | ↓ p = 0.0007 | ns (p = 0.54) |
| HXK3 | Hk3 | Hexokinase-3 | ↑ p = 0.03094 | ↓ p = 0.00115 | ns (p = 0.96) |
| IFM3 | Ifitm3 | Interferon-induced transmembrane protein 3 | ↑ p = 0.00314 | ↓ p = 0.0218 | ns (p = 0.95) |
| IMPA1 | Impa1 | Inositol monophosphatase 1 | ↑ p = 0.0029 | ↓ p = 0.02081 | ns (p = 0.29) |
| KAP0 | Prkar1a | cAMP-dependent protein kinase type I-alpha regulatory subunit | ↑ p = 0.00633 | ↓ p = 5.58 10−5 | ns (p = 0.08) |
| LEG1 | Lgals1 | Galectin-1 | ↑ p = 0.01128 | ↓ p = 0.00304 | ns (p = 0.30) |
| LRC59 | Lrrc59 | Leucine-rich repeat-containing protein 59 | ↑ p = 0.00057 | ↓ p = 0.00105 | ns (p = 0.76) |
| MBB1A | Mybbp1a | Myb-binding protein 1A | ↑ p = 0.00035 | ↓ p = 0.00673 | ns (p = 0.71) |
| MK03 | Mapk3 | Mitogen-activated protein kinase 3 | ↑ p = 0.00899 | ↓ p = 0.01772 | ns (p = 0.91) |
| MTNA | Mri1 | Methylthioribose-1-phosphate isomerase | ↑ p = 0.00444 | ↓ p = 0.0179 | ns (p = 0.86) |
| MVP | Mvp | Major vault protein | ↑ p = 0.00743 | ↓ p = 0.02179 | ns (p = 0.54) |
| MYL6 | Myl6 | Myosin light polypeptide 6 | ↑ p = 0.0052 | ↓ p = 0.01144 | ns (p = 0.75) |
| OPLA | Oplah | 5-oxoprolinase | ↑ p = 0.04606 | ↓ p = 0.03249 | ns (p = 0.52) |
| PACN2 | Pacsin2 | Protein kinase C and casein substrate in neurons 2 protein | ↑ p = 0.03552 | ↓ p = 0.02471 | ns (p = 0.99) |
| PDCD4 | Pdcd4 | Programmed cell death protein 4 | ↑ p = 0.0012 | ↓ p = 0.02955 | ns (p = 0.16) |
| PDIA1 | P4hb | Protein disulfide isomerase | ↑ p = 0.03143 | ↓ p = 0.03366 | ns (p = 0.47) |
| PDLI5 | Pdlim5 | PDZ and LIM domain protein 5 | ↑ p = 0.03056 | ↓ p = 0.03767 | ns (p = 0.64) |
| PHLB1 | Phldb1 | Pleckstrin homolog-like domain family B member 1 | ↑ p = 0.03397 | ↓ p = 0.01971 | ns (p = 0.35) |
| PSB10 | Psmb10 | Proteasome subunit beta type-10 | ↑ p = 0.00099 | ↓ p = 0.00199 | ns (p = 0.97) |
| PURB | Purb | Transcriptional activator protein Pur-beta | ↑ p = 0.00037 | ↓ p = 0.01449 | ns (p = 0.86) |
| RHOA | Rhoa | Transforming protein RhoA | ↑ p = 0.01258 | ↓ p = 0.00027 | ns (p = 0.48) |
| RL13 | Rpl13 | 60S ribosomal protein L13 | ↑ p = 0.00388 | ↓ p = 0.00536 | ns (p = 0.59) |
| RL18 | Rpl18 | 60S ribosomal protein L18 | ↑ p = 0.00386 | ↓ p = 0.0196 | ns (p = 0.45) |
| RL3 | Rpl3 | 60S ribosomal protein L3 | ↑ p = 0.00187 | ↓ p = 0.00629 | ns (p = 0.38) |
| RL35A | Rpl35a | 60S ribosomal protein L35a | ↑ p = 0.03126 | ↓ p = 0.01536 | ns (p = 0.31) |
| ROA2 | Hnrnpa2b1 | Heterogeneous nuclear ribonucleoprotein s A2/B1 | ↑ p = 0.00755 | ↓ p = 0.0135 | ns (p = 0.39) |
| SPRC | Sparc | SPARC | ↑ p = 0.044 | ↓ p = 0.02796 | ns (p = 0.36) |
| STIM1 | Stim1 | Stromal interaction molecule 1 | ↑ p = 0.02664 | ↓ p = 0.03078 | ns (p = 0.61) |
| TAGL2 | Tagln2 | Transgelin-2 | ↑ p = 0.00181 | ↓ p = 0.00397 | ns (p = 0.08) |
| USO1 | Uso1 | General vesicular transport factor p115 | ↑ p = 0.01675 | ↓ p = 0.01326 | ns (p = 0.88) |
| WDR1 | Wdr1 | WD repeat-containing protein 1 | ↑ p = 0.01835 | ↓ p = 0.01242 | ns (p = 0.93) |
| WIPF1 | Wipf1 | WAS/WASL-interacting protein family member 1 | ↑ p = 0.00052 | ↓ p = 0.02194 | ns (p = 0.56) |
| ARF3 | Arf3 | ADP-ribosylation factor 3 | ↓ p = 0.03262 | ↑ p = 0.04184 | ns (p = 0.23) |
| CAH1 | Ca1 | Carbonic anhydrase 1 | ↓ p = 2.30 × 10−5 | ↑ p = 2.53 × 10−5 | ns (p = 0.47) |
| DCUP | Urod | Uroporphyrinogen decarboxylase | ↓ p = 0.03293 | ↑ p = 0.04001 | ns (p = 0.92) |
| HEM2 | Alad | Delta-aminolevulinic acid dehydratase | ↓ p = 0.0004 | ↑ p = 0.03423 | ns (p = 0.14) |
| NP1L1 | Nap1l1 | Nucleosome assembly protein 1-like 1 | ↓ p = 0.00498 | ↑ p = 0.01454 | ns (p = 0.53) |
| BIEA | Blvra | Biliverdin reductase A | ↑ p = 3.00 10−5 | ↓ p = 0.00726 | ↑ p = 0.00148 |
| CAN2 | Capn2 | Calpain-2 catalytic subunit | ↑ p = 1.09 × 10−5 | ↓ p = 0.02941 | p = 0.01691 |
| CERU | Cp | Ceruloplasmin | ↑ p = 2.72 × 10−5 | ↓ p = 0.00447 | ↑ p = 0.01102 |
| CO3 | C3 | Complement C3 | ↑ p = 5.32 × 10−9 | ↓ p = 0.00065 | ↑ p = 1.46 × 10−5 |
| CRP | Crp | C-reactive protein | ↑ p = 1.68 × 10−7 | ↓ p = 0.0023 | ↑ p = 2.50 × 10−5 |
| CSK | Csk | Tyrosine-protein kinase CSK | ↑ p = 6.65 × 10−5 | ↓ p = 0.00093 | ↑ p = 0.0141 |
| FIBA | Fga | Fibrinogen alpha chain | ↑ p = 0.00196 | ↓ p = 0.00607 | ↑ p = 0.03133 |
| FIBB | Fgb | Fibrinogen beta chain | ↑ p = 0.00582 | ↓ p = 0.02273 | ↑ p = 7.65 × 10−5 |
| FINC | Fn1 | Fibronectin | ↑ p = 2.36 × 10−5 | ↓ p = 0.00042 | ↑ p = 0.0063 |
| HEMO | Hpx | Hemopexin | ↑ p = 0.00019 | ↓ p = 0.0155 | ↑ p = 0.00854 |
| HSPB1 | Hspb1 | Heat shock protein beta-1 | ↑ p = 9.81 × 10−5 | ↓ p = 0.02131 | p = 0.0003 |
| S10A4 | S100a4 | Protein S100-A4 | ↑ p = 0.00015 | ↓ p = 0.00305 | ↑ p = 0.00534 |
| ATOX1 | Atox1 | Copper transport protein ATOX1 | ↓ p = 0.00085 | ↑ p = 0.03533 | ↓ p = 0.02508 |
| B3AT | Slc4a1 | Band 3 anion transport protein | ↓ p = 0.04023 | ↑ p = 0.00039 | ↑ p = 0.00089 |
| GYS2 | Gys2 | Glycogen [starch] synthase, liver | ↑ p = 0.00021 | ↑ p = 0.02082 | ↑ p = 0.0006 |
| PRDX2 | Prdx2 | Peroxiredoxin-2 | ↑ p = 0.0008 | ↑ p = 0.02675 | ↑ p = 0.00086 |
| ARP3 | Actr3 | Actin-related protein 3 | ↓ p = 0.0345 | ↓ p = 0.00646 | ↓ p = 0.00054 |
| CH10 | Hspe1 | 10 kDa heat shock protein, mitochondrial | ↓ p = 0.0041 | ↓ p = 0.04784 | ↓ p = 0.00055 |
| PDIA3 | Pdia3 | Protein disulfide isomerase A3 | ↓ p = 0.00265 | ↓ p = 0.00268 | ↓ p = 4.86 × 10−5 |
| PPIA | Ppia | Peptidyl-prolyl cis-trans isomerase A | ↓ p = 0.02202 | ↓ p = 0.01276 | ↓ p = 0.00125 |
| RL13A | Rpl13a | 60S ribosomal protein L13a | ↓ p = 0.02799 | ↓ p = 0.01762 | ↓ p = 0.00137 |
| RS16 | Rps16 | 40S ribosomal protein S16 | ↓ p = 0.00333 | ↓ p = 0.04225 | ↓ p = 0.0003 |
| RS23 | Rps23 | 40S ribosomal protein S23 | ↓ p = 0.00851 | ↓ p = 0.01798 | ↓ p = 6.27 × 10−5 |
| Authors (Year) | Biological Model/Topic | |
|---|---|---|
| AHR (Ahr) | Ciolino et al. [12] | MCF-7 Human breast carcinoma cells |
| Rinaldi et al. [13] | Human oral squamous cell carcinoma cells | |
| Nishiumi et al. [14] | Mouse hepatoma Hepa-1c1c7 | |
| Choi et al. [15] | Hep3B, MCF-7, HEK 293 human cells… | |
| Garg et al. [16] | Mice (in vivo) | |
| Cifti et al. [17] | Rats (in vivo) | |
| Singh et al. [18] | Drosophila larvae | |
| Mohammadi-Bardbori et al. [19] | Human hepatoma | |
| Nakai et al. [20] | Mouse hepatoma | |
| ANXA5 (Anxa5) | Kam et al. [21] | EAhy926 human endothelial cells |
| CO9 (C9) | Jacob et al. [22] | Mice (in vivo) |
| GRP78 (Hspa5) | Ravindranathan et al. [23] | Colorectal cancer cells |
| LEG1 (Lgals1) | Rabinovitch et al. [24] | Normal rat T cells (from spleen) |
| Brandt et al. [25] | Human leukemic T cells | |
| MVP (Mvp) | Thiyagarajan et al. [26] | Y79 human retinoblastoma cells |
| PDCD4 (Pdcd4) | Mudduluru et al. [27] | Rko and HCT116 human colorectal cancer cells |
| Yang et al. [28] | DU145 human prostate cancer cells, B16 murine melanoma cells | |
| Chen et al. [29] | Review: microRNAs regulation | |
| Lelli et al. [30] | Review: melanoma | |
| Tan et al. [31] | C6 rat experimental glioblastoma | |
| Shakeri et al. [32] | Review: microRNA-21 | |
| PDIA1 (P4hb) | Ouyang et al. [33] | Mouse intestinal mucosa (in vivo) |
| PSB10 (Psmb10) | Wang et al. [34] | MCF-7 Human breast carcinoma cells |
| RHOA (Rhoa) | Zhang et al. [35] | Lu1205 and A375 human melanoma cells |
| Qin et al. [36] | HSC-T6 rat cells, rat liver fibrosis (in vivo) | |
| Wang et al. [37] | HMrSV5 human peritoneal mesothelial cells | |
| Gallardo et al. [38] | MCF-10F and MDA-MB-231 human breast cancer cells | |
| SPRC (Sparc) | Kilian et al. [39] | PC-3 human prostate cancer cells + xenografts in CD-1 mice (in vivo) |
| Wang et al. [34] | MCF-7 Human breast carcinoma cells | |
| STIM1 (Stim1) | Shin et al. [40] | HEK293 human embryonic kidney cells |
| TAGL2 (Tagln2) | Ma et al. [41] | CAL 27 human tongue cancer cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouliquen, D.L.; Boissard, A.; Henry, C.; Blandin, S.; Coqueret, O.; Guette, C. Lymphoid Organ Proteomes Identify Therapeutic Efficacy Biomarkers Following the Intracavitary Administration of Curcumin in a Highly Invasive Rat Model of Peritoneal Mesothelioma. Int. J. Mol. Sci. 2021, 22, 8566. https://doi.org/10.3390/ijms22168566
Pouliquen DL, Boissard A, Henry C, Blandin S, Coqueret O, Guette C. Lymphoid Organ Proteomes Identify Therapeutic Efficacy Biomarkers Following the Intracavitary Administration of Curcumin in a Highly Invasive Rat Model of Peritoneal Mesothelioma. International Journal of Molecular Sciences. 2021; 22(16):8566. https://doi.org/10.3390/ijms22168566
Chicago/Turabian StylePouliquen, Daniel L., Alice Boissard, Cécile Henry, Stéphanie Blandin, Olivier Coqueret, and Catherine Guette. 2021. "Lymphoid Organ Proteomes Identify Therapeutic Efficacy Biomarkers Following the Intracavitary Administration of Curcumin in a Highly Invasive Rat Model of Peritoneal Mesothelioma" International Journal of Molecular Sciences 22, no. 16: 8566. https://doi.org/10.3390/ijms22168566
APA StylePouliquen, D. L., Boissard, A., Henry, C., Blandin, S., Coqueret, O., & Guette, C. (2021). Lymphoid Organ Proteomes Identify Therapeutic Efficacy Biomarkers Following the Intracavitary Administration of Curcumin in a Highly Invasive Rat Model of Peritoneal Mesothelioma. International Journal of Molecular Sciences, 22(16), 8566. https://doi.org/10.3390/ijms22168566