
A Novel Oxygen Carrier (M101) Attenuates Ischemia-Reperfusion Injuries during Static Cold Storage in Steatotic Livers

 ,
,  , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
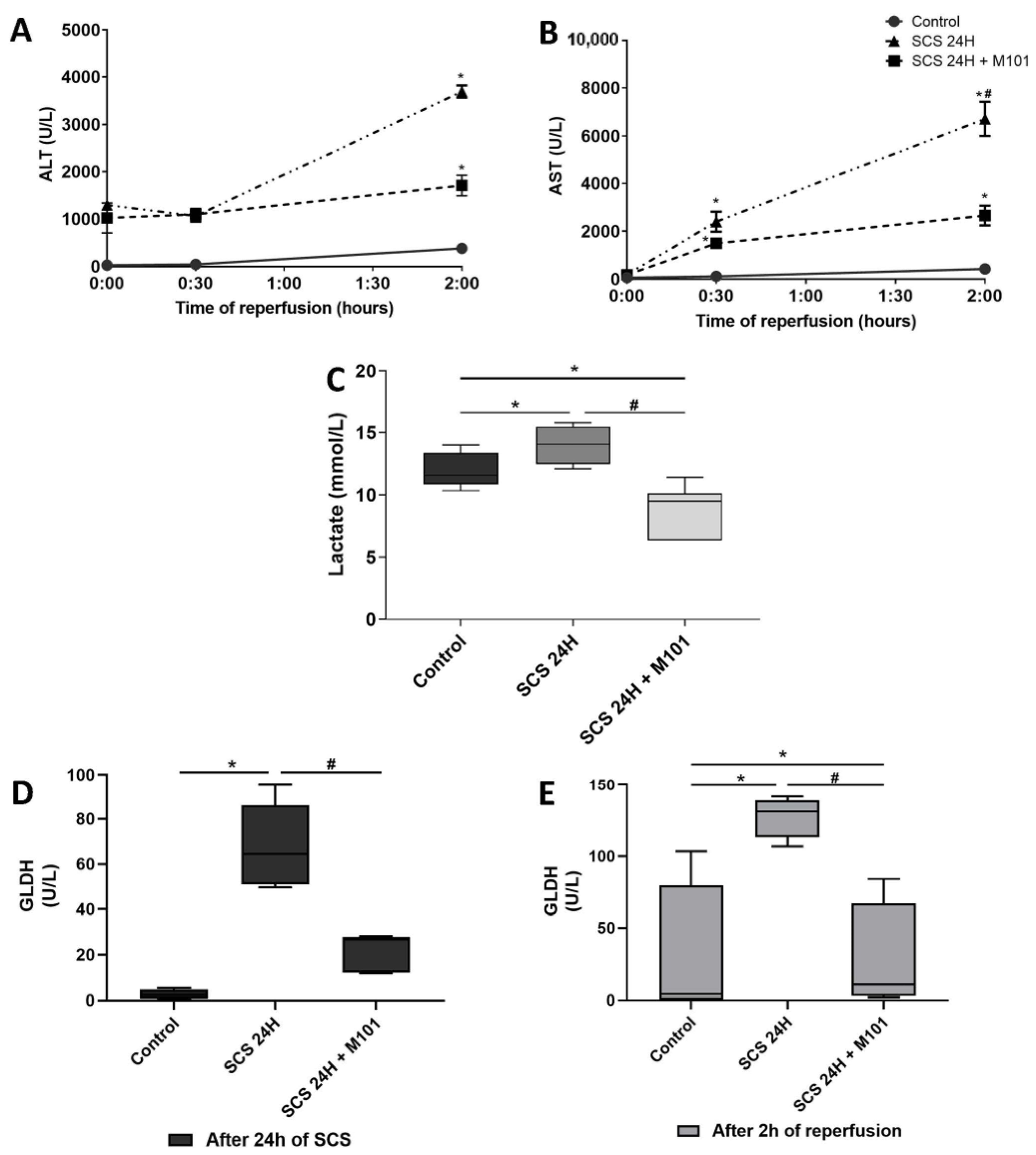
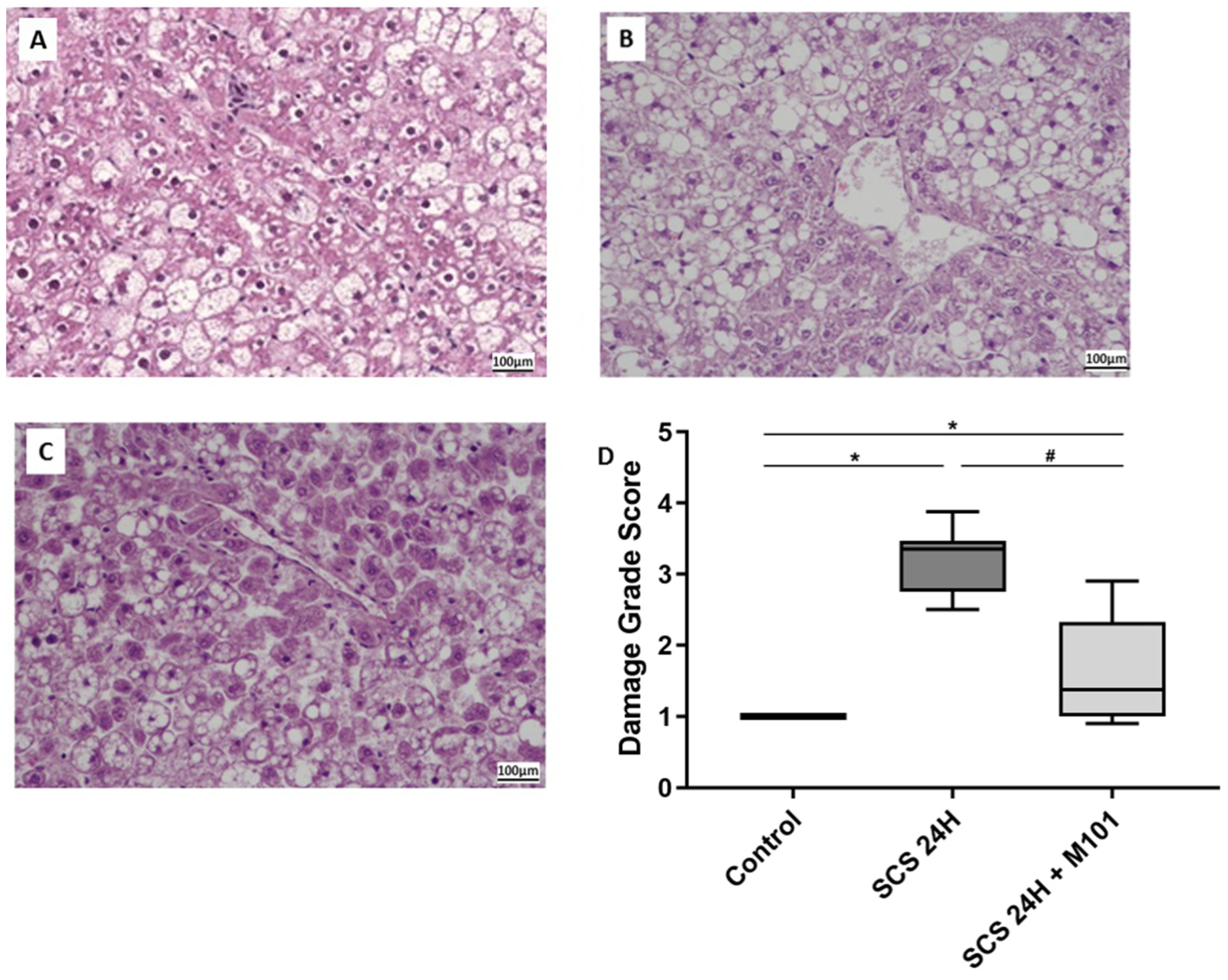
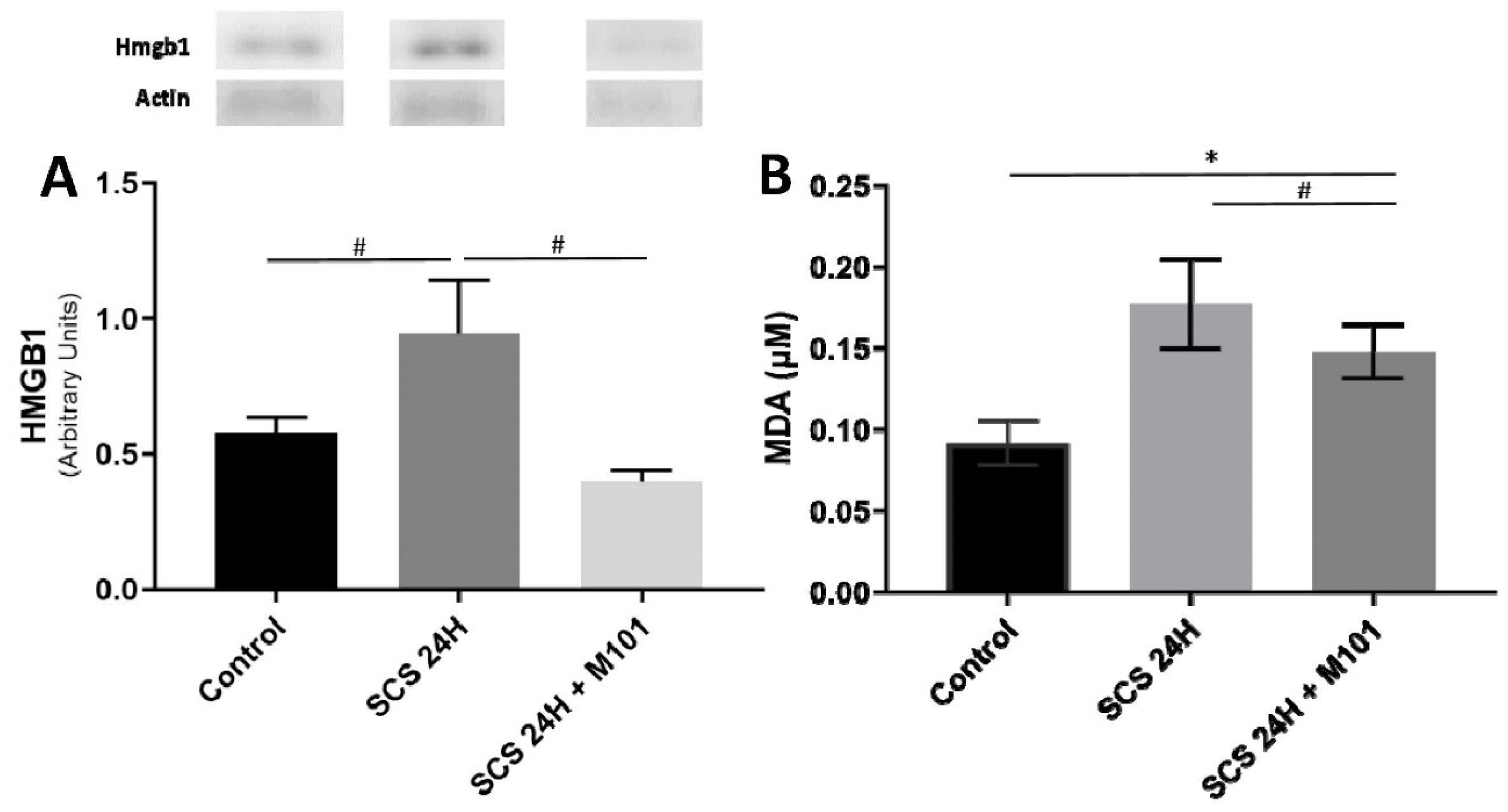
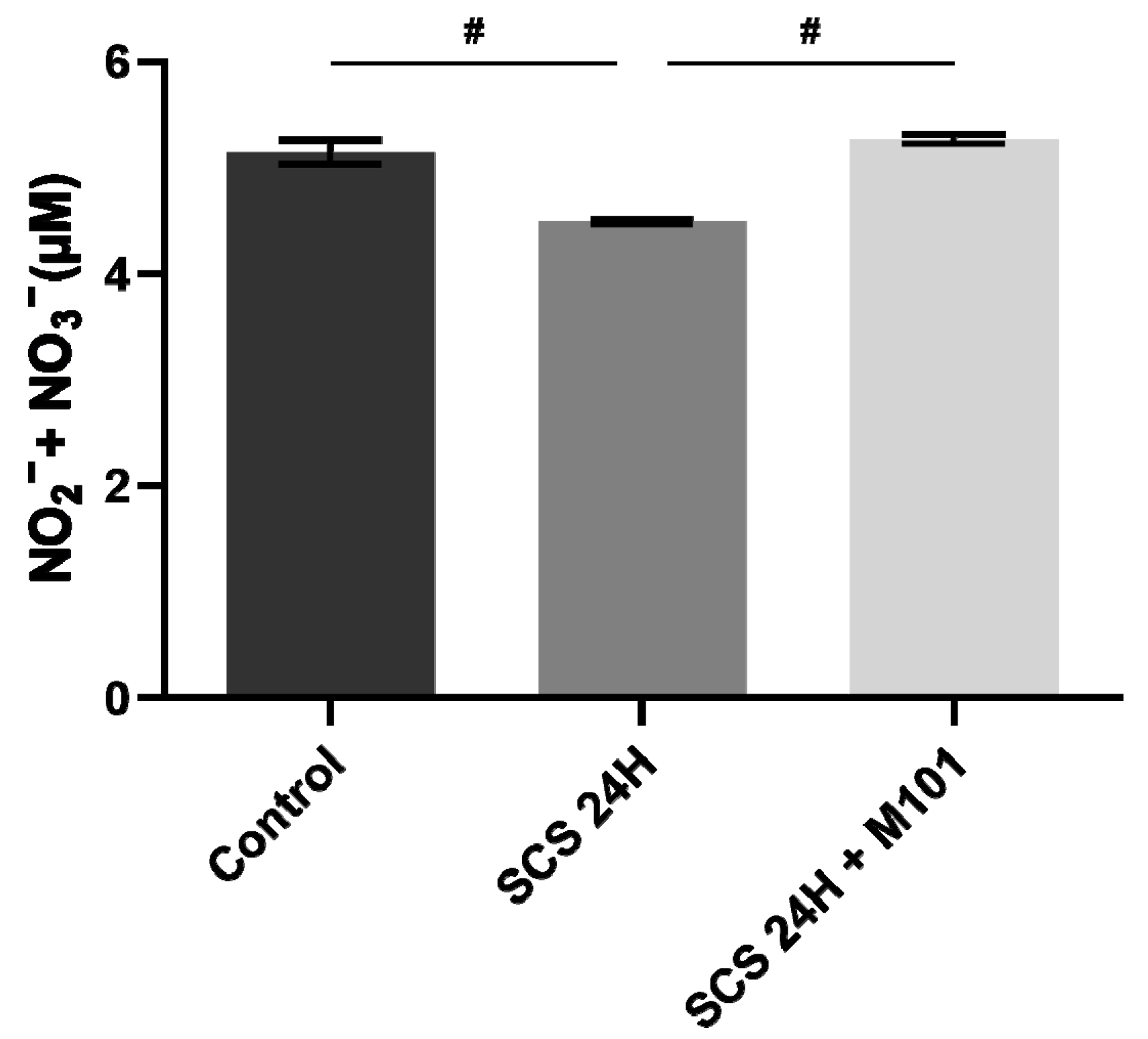
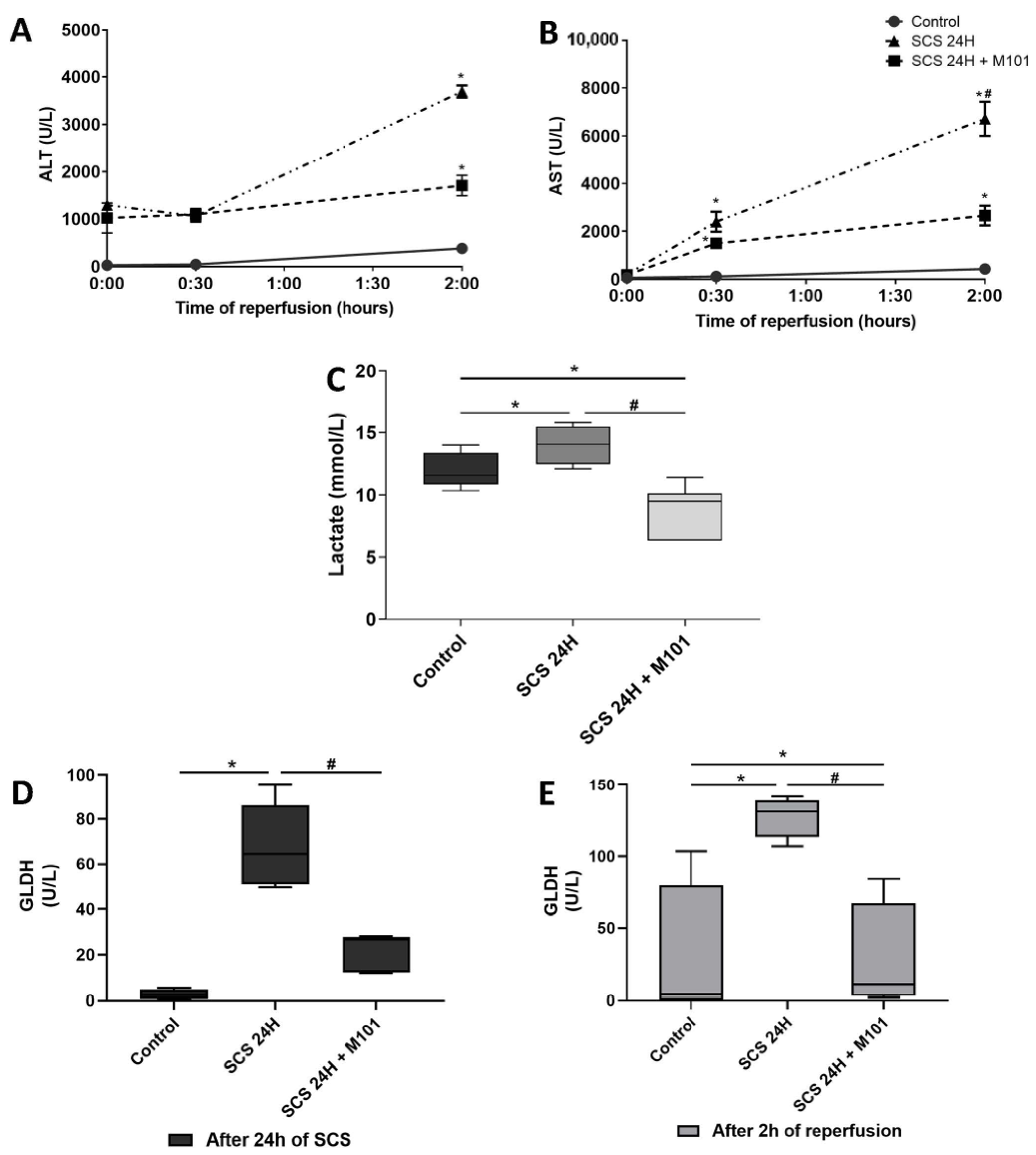
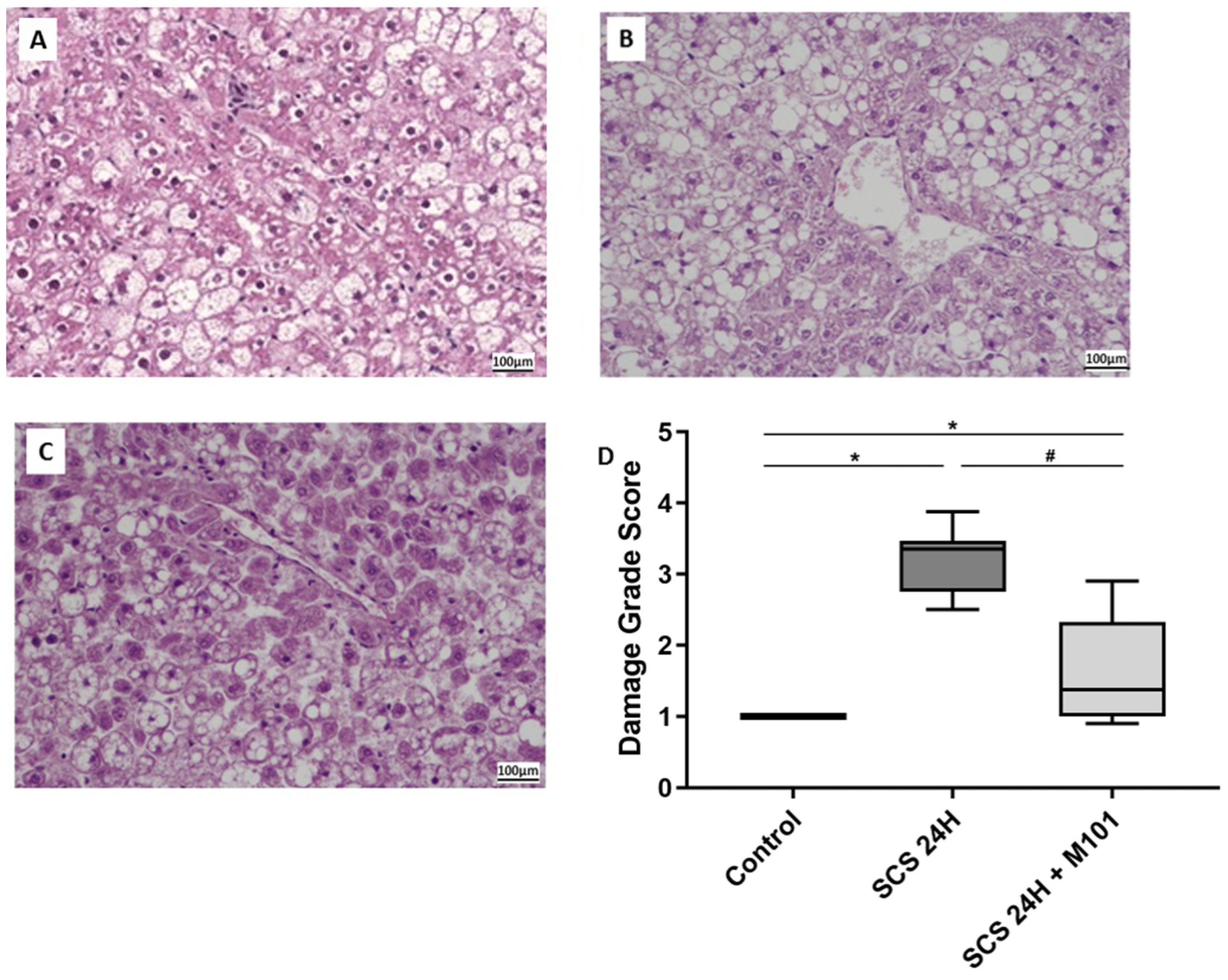
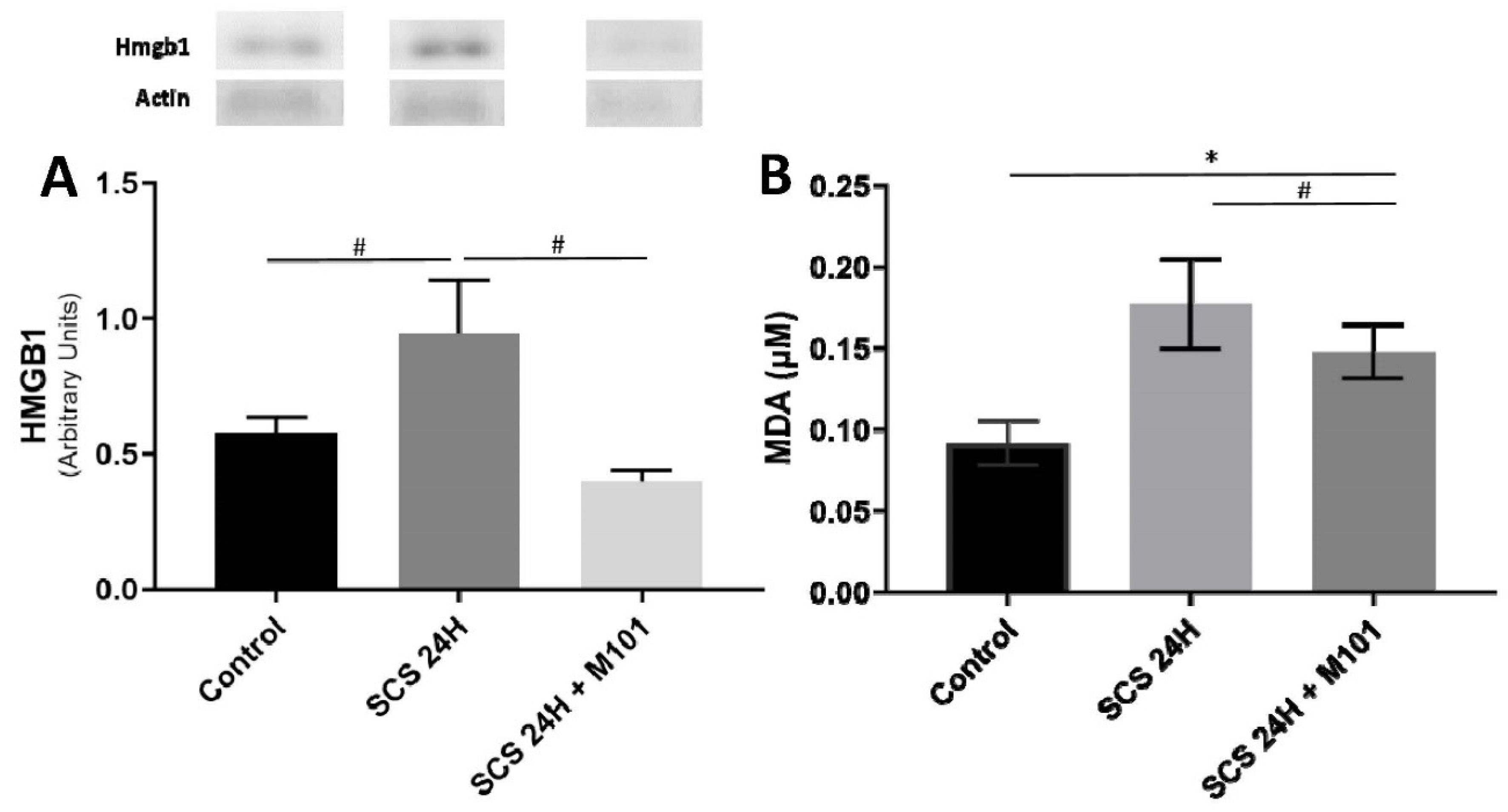
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. HEMO2life® and Preservation Solution
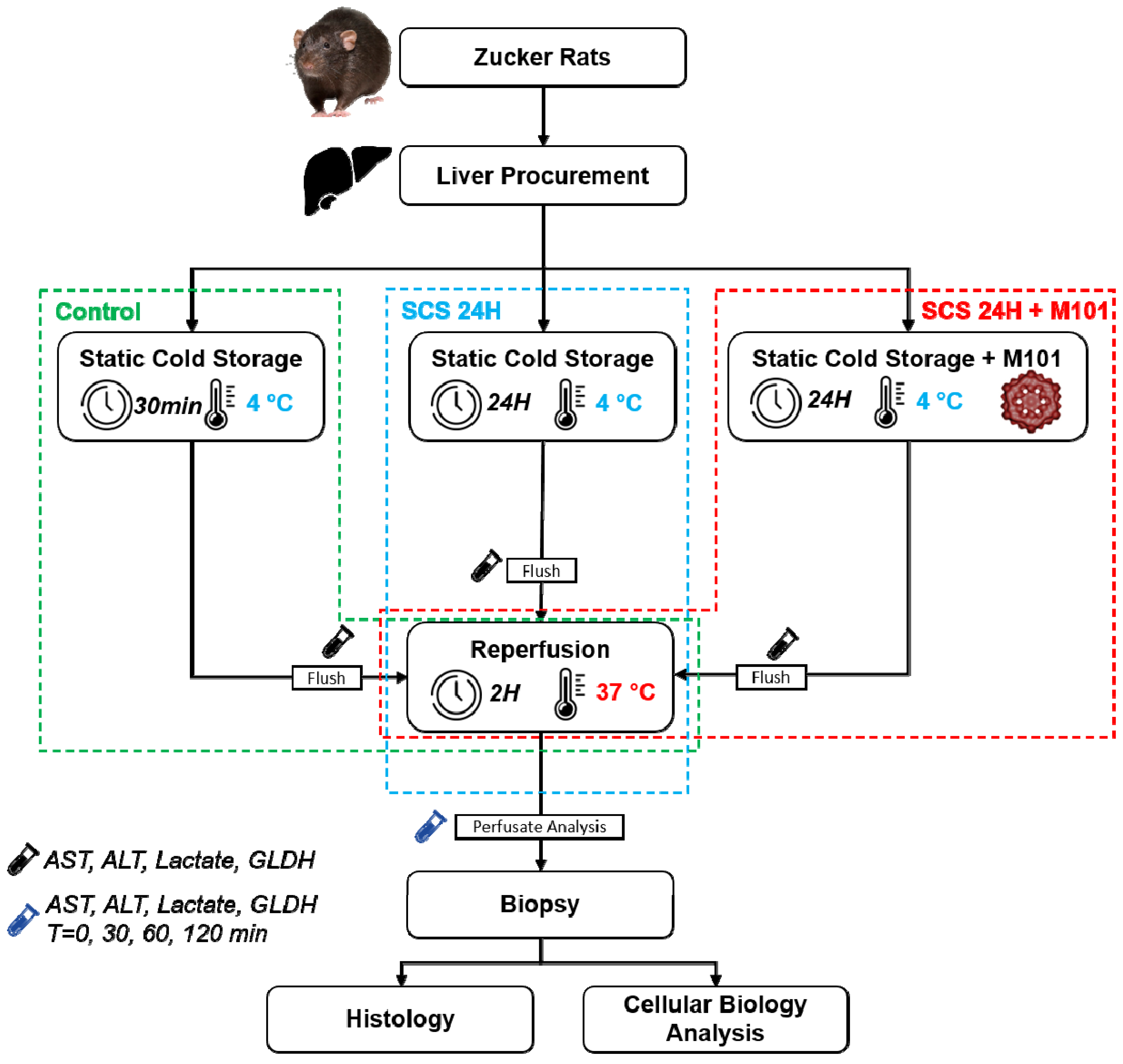
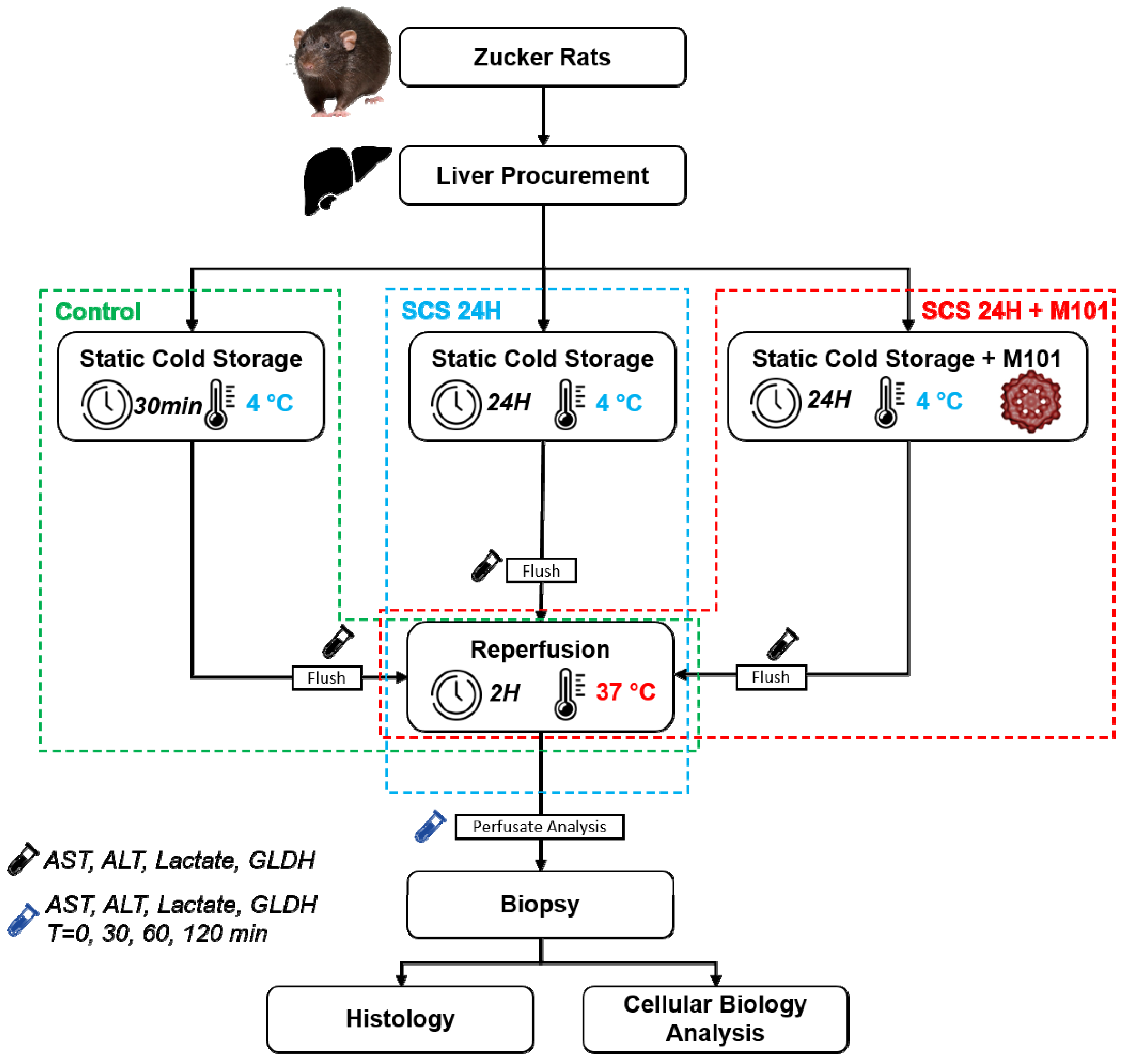
4.3. Liver Procurement and Experimental Groups
4.4. Histology
4.5. Biochemical Determination
4.6. Western Blot
4.7. ELISA
4.8. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet 2018, 391, 1301–1314. [Google Scholar] [CrossRef]
- Torbenson, M. Hepatic Adenomas. Surg. Pathol. Clin. 2018, 11, 351–366. [Google Scholar] [CrossRef]
- Halliday, N.; Westbrook, R.H. Liver transplantation: Need, indications, patient selection and pre-transplant care. Br. J. Hosp. Med. 2017, 78, 252–259. [Google Scholar] [CrossRef]
- Nair, A.; Hashimoto, K. Extended criteria donors in liver transplantation-from marginality to mainstream. Hepatobiliary Surg. Nutr. 2018, 7, 386–388. [Google Scholar] [CrossRef]
- Boteon, Y.L.; Boteon, A.P.C.D.S.; Attard, J.; Mergental, H.; Mirza, D.F.; Bhogal, R.H.; Afford, S.C. Ex situ machine perfusion as a tool to recondition steatotic donor livers: Troublesome features of fatty livers and the role of defatting therapies. A systematic review. Arab. Archaeol. Epigr. 2018, 18, 2384–2399. [Google Scholar] [CrossRef] [Green Version]
- Zamboni, F.; Franchello, A.; David, E.; Rocca, G.; Ricchiuti, A.; Lavezzo, B.; Rizzetto, M.; Salizzoni, M. Effect of macrovescicular steatosis and other donor and recipient characteristics on the outcome of liver transplantation. Clin. Transplant. 2001, 15, 53–57. [Google Scholar] [CrossRef] [PubMed]
- McCormack, L.; Dutkowski, P.; El-Badry, A.M.; Clavien, P.-A. Liver transplantation using fatty livers: Always feasible? J. Hepatol. 2011, 54, 1055–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llacuna, L.; Marí, M.; Lluis, J.M.; García-Ruiz, C.; Fernández-Checa, J.C.; Morales, A. Reactive Oxygen Species Mediate Liver Injury Through Parenchymal Nuclear Factor-κB Inactivation in Prolonged Ischemia/Reperfusion. Am. J. Pathol. 2009, 174, 1776–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nastos, C.; Kalimeris, K.; Papoutsidakis, N.; Tasoulis, M.-K.; Lykoudis, P.M.; Theodoraki, K.; Nastou, D.; Smyrniotis, V.; Arkadopoulos, N. Global consequences of liver ischemia/reperfusion injury. Oxid. Med. Cell Longev. 2014, 2014, 906965. [Google Scholar] [CrossRef] [Green Version]
- Dar, W.A.; Sullivan, E.; Bynon, J.S.; Eltzschig, H.; Ju, C. Ischaemia reperfusion injury in liver transplantation: Cellular and molecular mechanisms. Liver. Int. 2019, 39, 788–801. [Google Scholar] [CrossRef] [Green Version]
- Claes, G.; Aarell, M.; Brunius, U. Kidney preservation with continuing perfusion. Nord. Med. 1970, 84, 604–613. [Google Scholar]
- Leemkuil, M.; Lier, G.; Engelse, M.A.; Ploeg, R.J.; de Koning, E.J.P.; Hart, N.A.T.; Krikke, C.; Leuvenink, H.G.D. Hypothermic Oxygenated Machine Perfusion of the Human Donor Pancreas. Transplant. Direct 2018, 4, e388. [Google Scholar] [CrossRef] [PubMed]
- Meister, F.A.; Czigany, Z.; Bednarsch, J.; Böcker, J.; Amygdalos, I.; Santana, D.A.M.; Rietzler, K.; Moeller, M.; Tolba, R.; Boor, P.; et al. Hypothermic Oxygenated Machine Perfusion of Extended Criteria Kidney Allografts from Brain Dead Donors: Protocol for a Prospective Pilot Study. JMIR Res. Protoc. 2019, 8, e14622. [Google Scholar] [CrossRef] [PubMed]
- Karangwa, S.; Panayotova, G.; Dutkowski, P.; Porte, R.; Guarrera, J.; Schlegel, A. Hypothermic machine perfusion in liver transplantation. Int. J. Surg. 2020, 82, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Brüggenwirth, I.M.; van Leeuwen, O.B.; de Vries, Y.; Bodewes, S.B.; Adelmeijer, J.; Wiersema-Buist, J.; Lisman, T.; Martins, P.N.; de Meijer, V.E.; Porte, R.J. Extended hypothermic oxygenated machine perfusion enables ex situ preservation of porcine livers for up to 24 hours. JHEP Rep. 2020, 2, 100092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodewes, S.B.; Van Leeuwen, O.B.; Thorne, A.M.; Lascaris, B.; Ubbink, R.; Lisman, T.; Monbaliu, D.; De Meijer, V.E.; Nijsten, M.W.N.; Porte, R.J. Oxygen Transport during Ex Situ Machine Perfusion of Donor Livers Using Red Blood Cells or Artificial Oxygen Carriers. Int. J. Mol. Sci. 2020, 22, 235. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, X.; Guan, J. An oxygen release system to augment cardiac progenitor cell survival and differentiation under hypoxic condition. Biomaterials 2012, 33, 5914–5923. [Google Scholar] [CrossRef]
- Gupta, A.S. Hemoglobin-based Oxygen Carriers: Current State-of-the-art and Novel Molecules. Shock 2019, 52, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Li, C.; Guan, Y.; Dang, Y.; Li, X.; Fan, Z.; Shen, J.; Ma, L.; Guan, J. High oxygen preservation hydrogels to augment cell survival under hypoxic condition. Acta. Biomater. 2020, 105, 56–67. [Google Scholar] [CrossRef]
- Bhattacharjee, R.N.; Patel, S.V.; Sun, Q.; Jiang, L.; Richard-Mohamed, M.; Ruthirakanthan, A.; Aquil, S.; Al-Ogaili, R.; Juriasingani, S.; Sener, A.; et al. Renal Protection Against Ischemia Reperfusion Injury: Hemoglobin-based Oxygen Carrier-201 Versus Blood as an Oxygen Carrier in Ex Vivo Subnormothermic Machine Perfusion. Transplantation 2020, 104, 482–489. [Google Scholar] [CrossRef]
- Inci, I.; Arni, S.; Iskender, I.; Citak, N.; Rodriguez, J.M.; Weisskopf, M.; Opitz, I.; Weder, W.; Frauenfelder, T.; Krafft, M.P.; et al. Functional, Metabolic and Morphologic Results of Ex Vivo Donor Lung Perfusion with a Perfluorocarbon-Based Oxygen Carrier Nanoemulsion in a Large Animal Transplantation Model. Cells 2020, 9, 2501. [Google Scholar] [CrossRef]
- Martins, P.N.; Berendsen, T.A.; Yeh, H.; Bruinsma, B.G.; Izamis, M.-L.; Dries, S.O.D.; Gillooly, A.R.; Porte, R.; Yarmush, M.L.; Uygun, K.; et al. Oxygenated UW Solution Decreases ATP Decay and Improves Survival After Transplantation of DCD Liver Grafts. Transplantation 2019, 103, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Thuillier, R.; Dutheil, D.; Trieu, M.T.N.; Mallet, V.; Allain, G.; Rousselot, M.; Denizot, M.; Goujon, J.-M.; Zal, F.; Hauet, T. Supplementation with a New Therapeutic Oxygen Carrier Reduces Chronic Fibrosis and Organ Dysfunction in Kidney Static Preservation. Arab. Archaeol. Epigr. 2011, 11, 1845–1860. [Google Scholar] [CrossRef]
- Teh, E.S.; Zal, F.; Polard, V.; Menasché, P.; Chambers, D.J. HEMO2life as a protective additive to Celsior solution for static storage of donor hearts prior to transplantation. Artif. Cells Nanomed. Biotechnol. 2017, 45, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Glorion, M.; Polard, V.; Favereau, F.; Hauet, T.; Zal, F.; Fadel, E.; Sage, E. Prevention of ischemia-reperfusion lung injury during static cold preservation by supplementation of standard preservation solution with HEMO2life® in pig lung transplantation model. Artif. Cells Nanomed. Biotechnol. 2017, 46, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alix, P.; Val-Laillet, D.; Turlin, B.; Ben Mosbah, I.; Burel, A.; Bobillier, E.; Bendavid, C.; Delpy, E.; Zal, F.; Corlu, A.; et al. Adding the oxygen carrier M101 to a cold-storage solution could be an alternative to HOPE for liver graft preservation. JHEP Rep. 2020, 2, 100119. [Google Scholar] [CrossRef] [PubMed]
- Le Meur, Y.; Badet, L.; Essig, M.; Thierry, A.; Büchler, M.; Drouin, S.; Deruelle, C.; Morelon, E.; Pesteil, F.; Delpech, P.; et al. First-in-human use of a marine oxygen carrier (M101) for organ preservation: A safety and proof-of-principle study. Arab. Archaeol. Epigr. 2020, 20, 1729–1738. [Google Scholar] [CrossRef]
- Batool, F.; Stutz, C.; Petit, C.; Benkirane-Jessel, N.; Delpy, E.; Zal, F.; Leize-Zal, E.; Huck, O. A therapeutic oxygen carrier isolated from Arenicola marina decreased P. gingivalis induced inflammation and tissue destruction. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Mallet, V.; Dutheil, D.; Polard, V.; Rousselot, M.; Leize, E.; Hauet, T.; Goujon, J.M.; Zal, F. Dose-Ranging Study of the Performance of the Natural Oxygen Transporter HEMO2Life in Organ Preservation. Artif. Organs. 2014, 38, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, T.; Polard, V.; Rousselot, M.; Lotte, A.; Raouane, M.; Lehn, P.; Opolon, P.; Leize, E.; Deutsch, E.; Zal, F.; et al. In vivo biodistribution and oxygenation potential of a new generation of oxygen carrier. J. Biotechnol. 2014, 187, 1–9. [Google Scholar] [CrossRef]
- Lemaire, F.; Sigrist, S.; Delpy, E.; Cherfan, J.; Peronet, C.; Zal, F.; Bouzakri, K.; Pinget, M.; Maillard, E. Beneficial effects of the novel marine oxygen carrier M101 during cold preservation of rat and human pancreas. J. Cell. Mol. Med. 2019, 23, 8025–8034. [Google Scholar] [CrossRef]
- Ostrea, E.M.; Cepeda, E.E.; Fleury, C.A.; Balun, J.E. Red Cell Membrane Lipid Peroxidation and Hemolysis Secondary to Phototherapy. Acta. Paediatr. 1985, 74, 378–381. [Google Scholar] [CrossRef]
- Yang, R.; Zou, X.; Tenhunen, J.; Tønnessen, T.I. HMGB1 and Extracellular Histones Significantly Contribute to Systemic Inflammation and Multiple Organ Failure in Acute Liver Failure. Mediat. Inflamm. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Varadarajan, R.; Golden-Mason, L.; Young, L.; McLoughlin, P.; Nolan, N.; McEntee, G.; Traynor, O.; Geoghegan, J.; Hegarty, J.E.; O’Farrelly, C. Nitric Oxide in Early Ischaemia Reperfusion Injury during Human Orthotopic Liver Transplantation. Transplantation 2004, 78, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.T.; Martins, L.M.; Guimarães, J.E.; Sambade, C.; Vasconcelos, M.H. Jos Specific downregulation of bcl-2 and xIAP by RNAi enhances the effects of chemotherapeutic agents in MCF-7 human breast cancer cells. Cancer Gene Ther. 2004, 11, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akar, U.; Chaves-Reyez, A.; Barria, M.; Tari, A.; Sanguino, A.; Kondo, Y.; Kondo, S.; Arun, B.; Lopez-Berestein, G.; Ozpolat, B. Silencing of Bcl-2 expression by small interfering RNA induces autophagic cell death in MCF-7 breast cancer cells. Autophagy 2008, 4, 669–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathore, S.; Datta, G.; Kaur, I.; Malhotra, P.; Mohmmed, A. Disruption of cellular homeostasis induces organelle stress and triggers apoptosis like cell-death pathways in malaria parasite. Cell Death Dis. 2015, 6, e1803. [Google Scholar] [CrossRef] [Green Version]
- Allard, M.-A.; Castro-Benitez, C.; Imai, K.; Selten, J.; Lopez, A.; Sebagh, M.; Lemoine, A.; Cunha, A.S.; Cherqui, D.; Castaing, D.; et al. Suitability of livers for transplantation when treated by normothermic machine perfusion. Clin. Transplantayion 2018, 32, e13256. [Google Scholar] [CrossRef]
- Nagrath, D.; Xu, H.; Tanimura, Y.; Zuo, R.; Berthiaume, F.; Avila, M.; Yarmush, R.; Yarmush, M.L. Metabolic preconditioning of donor organs: Defatting fatty livers by normothermic perfusion ex vivo. Metab. Eng. 2009, 11, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, J.; Hannaert, P.; Kasil, A.; Thuillier, R.; Leize, E.; Delpy, E.; Steichen, C.; Goujon, J.M.; Zal, F.; Hauet, T. Efficacy of the natural oxygen transporter HEMO 2 life ® in cold preservation in a preclinical porcine model of donation after cardiac death. Transpl. Int. 2019, 32, 985–996. [Google Scholar] [CrossRef]
- Jipa, L.-N.; Tomescu, D.; Droc, G. The interrelation between arterial lactate levels and postoperative outcome following liver transplantation. Rom. J. Anaesth. Intensiv. Care 2014, 21, 106–112. [Google Scholar]
- Wu, J.-F.; Wu, R.-Y.; Chen, J.; Ou-Yang, B.; Chen, M.-Y.; Guan, X.-D. Early lactate clearance as a reliable predictor of initial poor graft function after orthotopic liver transplantation. Hepatobiliary Pancreat. Dis. Int. 2011, 10, 587–592. [Google Scholar] [CrossRef]
- Golse, N.; Guglielmo, N.; El Metni, A.; Frosio, F.; Cosse, C.; Naili, S.; Ichaï, P.; Ciacio, O.; Pittau, G.; Allard, M.-A.; et al. Arterial Lactate Concentration at the End of Liver Transplantation is an Early Predictor of Primary Graft Dysfunction. Ann. Surg. 2019, 270, 131–138. [Google Scholar] [CrossRef]
- Le Pape, F.; Cosnuau-Kemmat, L.; Richard, G.; Dubrana, F.; Férec, C.; Zal, F.; Leize, E.; Delépine, P. HEMOXCell, a New Oxygen Carrier Usable as an Additive for Mesenchymal Stem Cell Culture in Platelet Lysate-Supplemented Media. Artif. Organs. 2017, 41, 359–371. [Google Scholar] [CrossRef]
- Kim, J.-S.; He, L.; Lemasters, J.J. Mitochondrial permeability transition: A common pathway to necrosis and apoptosis. Biochem. Biophys. Res. Commun. 2003, 304, 463–470. [Google Scholar] [CrossRef]
- Milkovic, L.; Gasparovic, A.C.; Zarkovic, N. Overview on major lipid peroxidation bioactive factor 4-hydroxynonenal as pluripotent growth-regulating factor. Free. Radic. Res. 2015, 49, 850–860. [Google Scholar] [CrossRef]
- Schneider, C.; Porter, N.A.; Brash, A.R. Routes to 4-Hydroxynonenal: Fundamental Issues in the Mechanisms of Lipid Peroxidation. J. Biol. Chem. 2008, 283, 15539–15543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, A.; Panisello-Rosello, A.; Castro-Benitez, C.; Adam, R. Glycocalyx Preservation and NO Production in Fatty Livers—The Protective Role of High Molecular Polyethylene Glycol in Cold Ischemia Injury. Int. J. Mol. Sci. 2018, 19, 2375. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Meng, Z.; Liu, Y.; Patel, R.P.; Lang, J.D. The Hepatoprotective Effect of Sodium Nitrite on Cold Ischemia-Reperfusion Injury. J. Transplant. 2012, 2012, 635179. [Google Scholar] [CrossRef] [PubMed]
- Abu-Amara, M.; Yang, S.Y.; Seifalian, A.; Davidson, B.; Fuller, B. The nitric oxide pathway—evidence and mechanisms for protection against liver ischaemia reperfusion injury. Liver Int. 2012, 32, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Bhatraju, P.; Crawford, J.; Hall, M.; Lang, J.D. Inhaled nitric oxide: Current clinical concepts. Nitric. Oxide. 2015, 50, 114–128. [Google Scholar] [CrossRef]
- Di Pasqua, L.; Berardo, C.; Cagna, M.; Verta, R.; Collotta, D.; Nicoletti, F.; Ferrigno, A.; Collino, M.; Vairetti, M. Metabotropic Glutamate Receptor Blockade Reduces Preservation Damage in Livers from Donors after Cardiac Death. Int. J. Mol. Sci. 2021, 22, 2234. [Google Scholar] [CrossRef]
- Maejima, Y.; Adachi, S.; Morikawa, K.; Ito, H.; Isobe, M. Nitric oxide inhibits myocardial apoptosis by preventing caspase-3 activity via S-nitrosylation. J. Mol. Cell. Cardiol. 2005, 38, 163–174. [Google Scholar] [CrossRef]
- Cursio, R.; Colosetti, P.; Gugenheim, J. Autophagy and Liver Ischemia-Reperfusion Injury. Bio. Med. Res. Int. 2015, 2015, 417590. [Google Scholar] [CrossRef]
- Yan, W.-J.; Dong, H.-L.; Xiong, L. The protective roles of autophagy in ischemic preconditioning. Acta. Pharmacol. Sin. 2013, 34, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Marquez, R.T.; Xu, L. Bcl-2:Beclin 1 complex: Multiple, mechanisms regulating autophagy/apoptosis toggle switch. Am. J. Cancer Res. 2012, 2, 214–221. [Google Scholar] [PubMed]
- Kang, R.; Livesey, K.M.; Zeh, I.H.J.; Lotze, M.T.; Tang, D. HMGB1 as an autophagy sensor in oxidative stress. Autophagy 2011, 7, 904–906. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Kang, R.; Livesey, K.M.; Cheh, C.-W.; Farkas, A.M.; Loughran, P.; Hoppe, G.; Bianchi, M.E.; Tracey, K.J.; Zeh, H.J.; et al. Endogenous HMGB1 regulates autophagy. J. Cell Biol. 2010, 190, 881–892. [Google Scholar] [CrossRef]
- Levy, J.H. Hemoglobin-Based Oxygen Carriers. In Blood Substitutes; The American Council on Science and Health: New York, NY, USA, 2009. [Google Scholar]
- Smani, Y.; Labrude, P.; Vigneron, C.; Faivre, B. Les transporteurs d’oxygène à base d’hémoglobine et les tentatives de substituer les globules rouges. Transfus. Clin. Biol. 2007, 14, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Thuillier, R.; Delpy, E.; Matillon, X.; Kaminski, J.; Kasil, A.; Soussi, D.; Danion, J.; Sauvageon, Y.; Rod, X.; Donatini, G.; et al. Preventing acute kidney injury during transplantation: The application of novel oxygen carriers. Expert Opin. Investig. Drugs 2019, 28, 643–657. [Google Scholar] [CrossRef]
- Adam, R.; Delvart, V.; Karam, V.; Ducerf, C.; Navarro, F.; Letoublon, C.; Belghiti, J.; Pezet, D.; Castaing, D.; Le Treut, Y.P.; et al. Compared Efficacy of Preservation Solutions in Liver Transplantation: A Long-Term Graft Outcome Study From the European Liver Transplant Registry. Arab. Archaeol. Epigr. 2015, 15, 395–406. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asong-Fontem, N.; Panisello-Rosello, A.; Lopez, A.; Imai, K.; Zal, F.; Delpy, E.; Rosello-Catafau, J.; Adam, R. A Novel Oxygen Carrier (M101) Attenuates Ischemia-Reperfusion Injuries during Static Cold Storage in Steatotic Livers. Int. J. Mol. Sci. 2021, 22, 8542. https://doi.org/10.3390/ijms22168542
Asong-Fontem N, Panisello-Rosello A, Lopez A, Imai K, Zal F, Delpy E, Rosello-Catafau J, Adam R. A Novel Oxygen Carrier (M101) Attenuates Ischemia-Reperfusion Injuries during Static Cold Storage in Steatotic Livers. International Journal of Molecular Sciences. 2021; 22(16):8542. https://doi.org/10.3390/ijms22168542
Chicago/Turabian StyleAsong-Fontem, Njikem, Arnau Panisello-Rosello, Alexandre Lopez, Katsunori Imai, Franck Zal, Eric Delpy, Joan Rosello-Catafau, and René Adam. 2021. "A Novel Oxygen Carrier (M101) Attenuates Ischemia-Reperfusion Injuries during Static Cold Storage in Steatotic Livers" International Journal of Molecular Sciences 22, no. 16: 8542. https://doi.org/10.3390/ijms22168542
APA StyleAsong-Fontem, N., Panisello-Rosello, A., Lopez, A., Imai, K., Zal, F., Delpy, E., Rosello-Catafau, J., & Adam, R. (2021). A Novel Oxygen Carrier (M101) Attenuates Ischemia-Reperfusion Injuries during Static Cold Storage in Steatotic Livers. International Journal of Molecular Sciences, 22(16), 8542. https://doi.org/10.3390/ijms22168542