
The Effect of Chronic Iloperidone Treatment on Cytochrome P450 Expression and Activity in the Rat Liver: Involvement of Neuroendocrine Mechanisms



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. The Effect of Chronic Treatment with Iloperidone on the CYP Activity in Rat Liver Microsomes
2.2. The Influence of Two-Week Treatment with Iloperidone on the CYP Protein Level in Rat Liver Microsomes
2.3. The Effect of Iloperidone Treatment on the mRNA Level of CYP Enzymes in the Liver
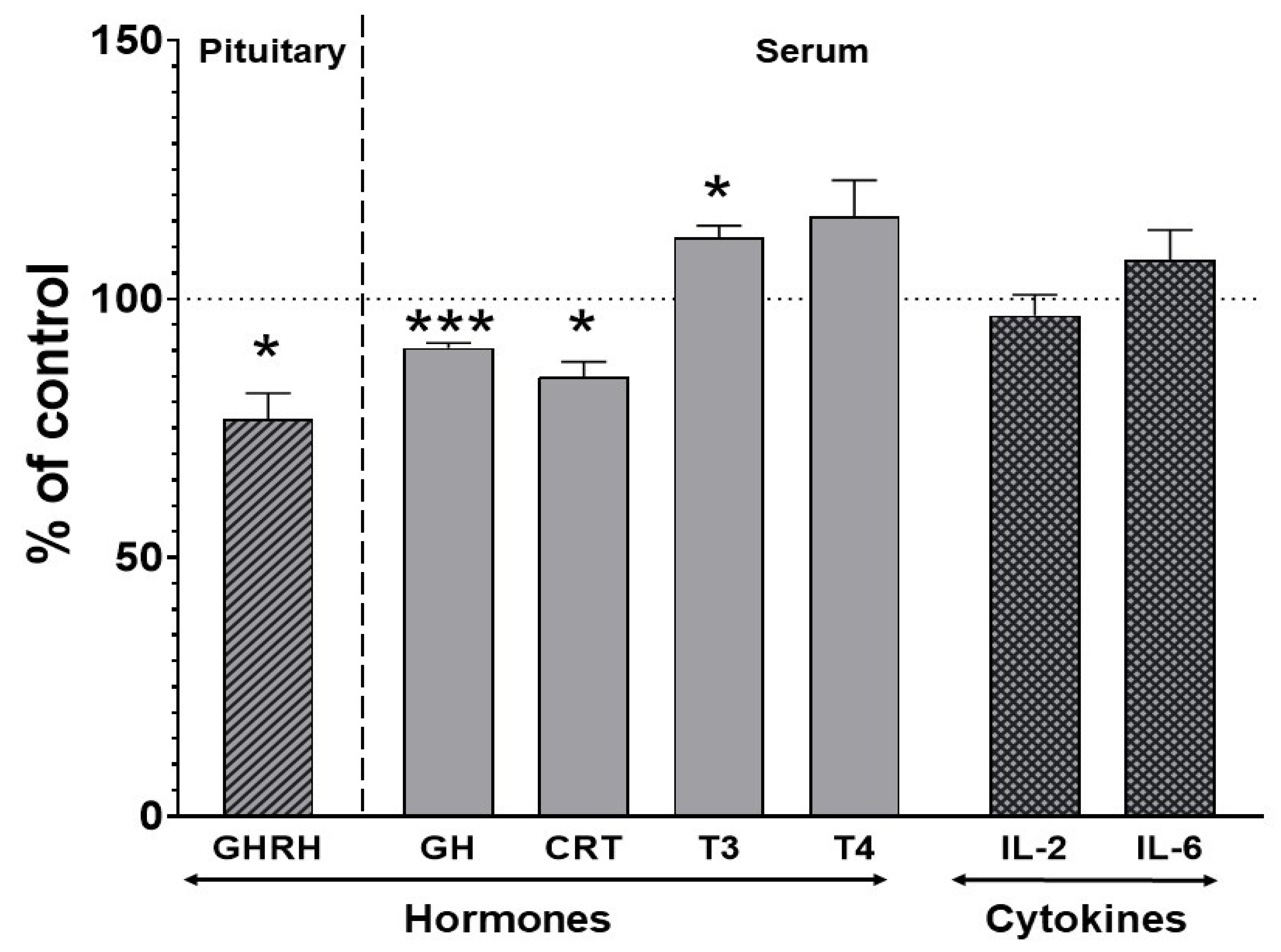
2.4. The Effect of Two-Week Treatment with Iloperidone on the Pituitary GHRH and Serum Concentrations of Hormones and Cytokines
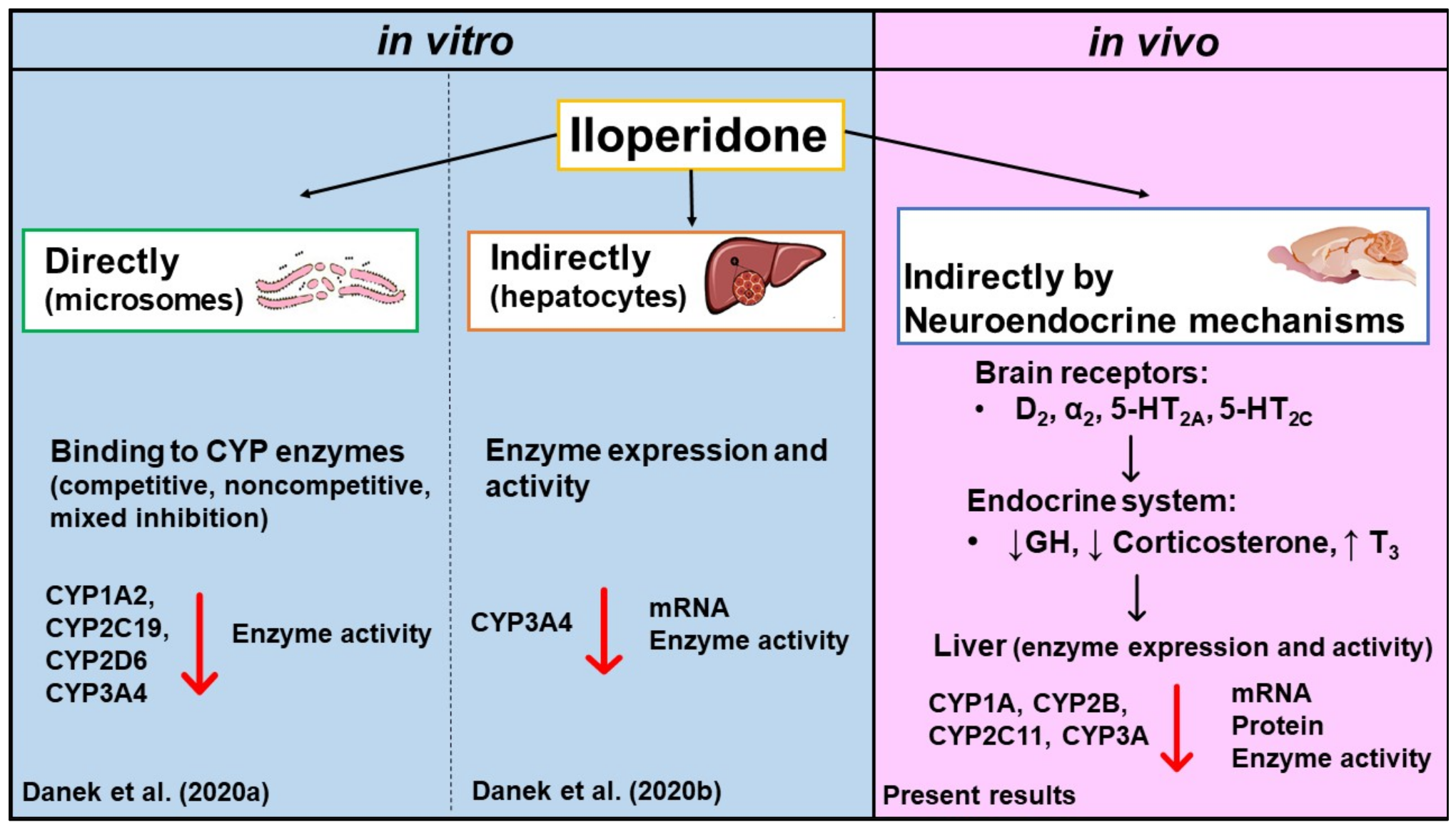
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Drug Treatment and Liver Sample Preparation
4.4. CYP Enzyme Activities in the Liver
4.5. Determination of CYP Proteins in Liver Microsomes
4.6. Analysis of mRNA Level in the Liver
4.7. Analysis of Hormones and Cytokines in the Pituitary and Blood Serum
4.8. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Munro, A.W.; McLean, K.J.; Grant, J.L.; Makris, T.M. Structure and function of the cytochrome P450 peroxygenase enzymes. Biochem. Soc. Trans. 2018, 46, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.J. Iloperidone. CNS Drugs 2009, 23, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Snyder, G.L.; Vanover, K.E. Dopamine Targeting Drugs for the Treatment of Schizophrenia: Past, Present and Future. Curr. Top. Med. Chem. 2016, 16, 3385–3403. [Google Scholar] [CrossRef] [Green Version]
- Citrome, L. Iloperidone for schizophrenia: A review of the efficacy and safety profile for this newly commercialised second-generation antipsychotic. Int. J. Clin. Pract. 2009, 63, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Citrome, L. A Review of the Pharmacology, Efficacy and Tolerability of Recently Approved and Upcoming Oral Antipsychotics: An Evidence-Based Medicine Approach. CNS Drugs 2013, 27, 879–911. [Google Scholar] [CrossRef] [PubMed]
- Tarazi, F.; Stahl, S.M. Iloperidone, asenapine and lurasidone: A primer on their current status. Expert Opin. Pharmacother. 2012, 13, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- Cutler, A.J.; Kalali, A.H.; Mattingly, G.W.; Kunovac, J.; Meng, X. Long-term safety and tolerability of iloperidone: Results from a 25-week, open-label extension trial. CNS Spectr. 2013, 18, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Tonin, F.; Wiens, A.; Fernandez-Llimos, F.; Pontarolo, R. Iloperidone in the treatment of schizophrenia: An evidence-based review of its place in therapy. Core Evid. 2016, 11, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Daniel, W.A. The influence of long-term treatment with psychotropic drugs on cytochrome P450: The involvement of different mechanisms. Expert Opin. Drug Metab. Toxicol. 2005, 1, 203–217. [Google Scholar] [CrossRef]
- Deodhar, M.; Al Rihani, S.; Arwood, M.; Darakjian, L.; Dow, P.; Turgeon, J.; Michaud, V. Mechanisms of CYP450 Inhibition: Understanding Drug-Drug Interactions Due to Mechanism-Based Inhibition in Clinical Practice. Pharmaceuticals 2020, 12, 846. [Google Scholar] [CrossRef]
- Wójcikowski, J.; Daniel, W.A. The brain dopaminergic system as an important center regulating liver cytochrome P450 in the rat. Expert Opin. Drug Metab. Toxicol. 2009, 5, 631–645. [Google Scholar] [CrossRef]
- Sadakierska-Chudy, A.; Haduch, A.; Rysz, M.; Gołembiowska, K.; Daniel, W.A. The role of brain noradrenergic system in the regulation of liver cytochrome P450 expression. Biochem. Pharmacol. 2013, 86, 800–807. [Google Scholar] [CrossRef]
- Bromek, E.; Wójcikowski, J.; Daniel, W.A. Involvement of the paraventricular (PVN) and arcuate (ARC) nuclei of the hypothalamus in the central noradrenergic regulation of liver cytochrome P450. Biochem. Pharmacol. 2013, 86, 1614–1620. [Google Scholar] [CrossRef]
- Bromek, E.; Daniel, W.A. The regulation of liver cytochrome P450 expression and activity by the brain serotonergic system in different experimental models. Expert Opin. Drug Metab. Toxicol. 2021, 17, 413–424. [Google Scholar] [CrossRef]
- Wojcikowski, J.; Daniel, W.A. The Role of the Nervous System in the Regulation of Liver Cytochrome P450. Curr. Drug Metab. 2011, 12, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Danek, P.; Bromek, E.; Daniel, W.A. The Influence of Long-Term Treatment with Asenapine on Liver Cytochrome P450 Expression and Activity in the Rat. The Involvement of Different Mechanisms. Pharmaceuticals 2021, 14, 629. [Google Scholar] [CrossRef]
- Danek, P.J.; Wójcikowski, J.; Daniel, W.A. The atypical neuroleptics iloperidone and lurasidone inhibit human cytochrome P450 enzymes in vitro. Evaluation of potential metabolic interactions. Pharmacol. Rep. 2020, 72, 1685–1694. [Google Scholar] [CrossRef] [Green Version]
- Danek, P.J.; Wójcikowski, J.; Daniel, W.A. Asenapine and iloperidone decrease the expression of major cytochrome P450 enzymes CYP1A2 and CYP3A4 in human hepatocytes. A significance for drug-drug interactions during combined therapy. Toxicol. Appl. Pharmacol. 2020, 406, 115239. [Google Scholar] [CrossRef] [PubMed]
- Waxman, D.J.; O’Connor, C. Growth Hormone Regulation of Sex-Dependent Liver Gene Expression. Mol. Endocrinol. 2006, 20, 2613–2629. [Google Scholar] [CrossRef] [PubMed]
- Waxman, D.J.; Holloway, M.G. Sex Differences in the Expression of Hepatic Drug Metabolizing Enzymes. Mol. Pharmacol. 2009, 76, 215–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monostory, K.; Kőhalmy, K.; Prough, R.A.; Kóbori, L.; Vereczkey, L. The effect of synthetic glucocorticoid, dexamethasone on CYP1A1 inducibility in adult rat and human hepatocytes. FEBS Lett. 2004, 579, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monostory, K.; Pascussi, J.-M.; Kóbori, L.; Dvorak, Z. Hormonal regulation of CYP1A expression. Drug Metab. Rev. 2009, 41, 547–572. [Google Scholar] [CrossRef]
- Audet-Walsh, E.; Auclair-Vincent, S.; Anderson, A. Glucocorticoids and phenobarbital induce murineCYP2Bgenes by independent mechanisms. Expert Opin. Drug Metab. Toxicol. 2009, 5, 1501–1511. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, Z.; Pavek, P. Regulation of drug-metabolizing cytochrome P450 enzymes by glucocorticoids. Drug Metab. Rev. 2010, 42, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Monostory, K.; Dvorak, Z. Steroid Regulation of Drug-Metabolizing Cytochromes P450. Curr. Drug Metab. 2011, 12, 154–172. [Google Scholar] [CrossRef]
- Yamazoe, Y.; Murayama, N.; Shimada, M.; Kato, R. Thyroid hormone suppression of hepatic levels of phenobarbital-inducible P-450b AND P-450e and other neonatal P-450S in hypophysectomized rats. Biochem. Biophys. Res. Commun. 1989, 160, 609–614. [Google Scholar] [CrossRef]
- Murayama, N.; Shimada, M.; Yamazoe, Y.; Kato, R. Difference in the Susceptibility of Two Phenobarbital-Inducible Forms, P450IIB1 and P450IIB2, to Thyroid Hormone- and Growth Hormone-Induced Suppression in Rat Liver: Phenobarbi-tal-Inducible P450IIB2 Suppression by Thyroid Hormone Acting Directly, but Not through the Pituitary System. Mol. Pharmacol. 1991, 39, 811–817. [Google Scholar]
- Brtko, J.; Dvorak, Z. Role of Retinoids, Rexinoids and Thyroid Hormone in the Expression of Cytochrome P450 Enzymes. Curr. Drug Metab. 2011, 12, 71–88. [Google Scholar] [CrossRef]
- Pascussi, J.-M.; Gerbal-Chaloin, S.; Duret, C.; Daujat-Chavanieu, M.; Vilarem, M.-J.; Maurel, P. The Tangle of Nuclear Receptors that Controls Xenobiotic Metabolism and Transport: Crosstalk and Consequences. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Tolson, A.H.; Wang, H. Regulation of drug-metabolizing enzymes by xenobiotic receptors: PXR and CAR. Adv. Drug Deliv. Rev. 2010, 62, 1238–1249. [Google Scholar] [CrossRef] [Green Version]
- Daujat-Chavanieu, M.; Gerbal-Chaloin, S. Regulation of CAR and PXR Expression in Health and Disease. Cells 2020, 9, 2395. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, X.C.; Baldwin, W.S. CAR and PXR: Xenosensors of endocrine disrupters? Chem. Interact. 2005, 155, 111–128. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, S.; Wang, J.; Renukuntla, J.; Sirimulla, S.; Chen, J. A comprehensive review of cytochrome P450 2E1 for xenobiotic metabolism. Drug Metab. Rev. 2019, 51, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Ingelman-Sundberg, M.; Ronis, M.J.; Lindros, K.O.; Eliasson, E.; Zhukov, A. Ethanol-Inducible Cytochrome P4502E1: Regula-tion, Enzymology and Molecular Biology. Alcohol Alcohol. Suppl. 1994, 2, 131–139. [Google Scholar]
- Song, B.-J.; Abdelmegeed, M.A.; Cho, Y.-E.; Akbar, M.; Rhim, J.S.; Song, M.-K.; Hardwick, J.P. Contributing Roles of CYP2E1 and Other Cytochrome P450 Isoforms in Alcohol-Related Tissue Injury and Carcinogenesis. Adv. Exp. Med. Biol. 2019, 1164, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Koop, D.R.; Tierney, D.J. Multiple mechanisms in the regulation of ethanol-inducible cytochrome P450IIE1. BioEssays 1990, 12, 429–435. [Google Scholar] [CrossRef]
- Gonzalez, F.J.; Ueno, T.; Umeno, M.; Song, B.J.; Veech, R.L.; Gelboin, H.V. Microsomal Ethanol Oxidizing System: Transcrip-tional and Posttranscriptional Regulation of Cytochrome P450, CYP2E1. Alcohol Alcohol. Suppl. 1991, 1, 97–101. [Google Scholar]
- Eliasson, E.; Mkrtchian, S.; Ingelman-Sundberg, M. Hormone- and Substrate-Regulated Intracellular Degradation of Cyto-chrome P450 (2E1) Involving MgATP-Activated Rapid Proteolysis in the Endoplasmic Reticulum Membranes. J. Biol. Chem. 1992, 267, 15765–15769. [Google Scholar] [CrossRef]
- Martini, R.; Ingelman-Sundberg, M.; Murray, M. Pretranslational and Posttranslational Regulation of Rat Hepatic CYPs 3A2 and 2E1 by Disulfiram. Biochem. Pharmacol. 1997, 54, 1323–1329. [Google Scholar] [CrossRef]
- Novak, R.F.; Woodcroft, K.J. The alcohol-inducible form of cytochrome P450 (CYP2E1): Role in toxicology and regulation of expression. Arch. Pharmacal Res. 2000, 23, 267–282. [Google Scholar] [CrossRef]
- Chen, G.-F.; Ronis, M.J.J.; Ingelman-Sundberg, M.; Badger, T.M. Hormonal regulation of microsomal cytochrome P4502E1 and P450 reductase in rat liver and kidney. Xenobiotica 1999, 29, 437–451. [Google Scholar] [CrossRef]
- Williams, M.T.; Simonet, L.C. Effects of growth hormone on cytochrome P-450j. Biochem. Biophys. Res. Commun. 1988, 155, 392–397. [Google Scholar] [CrossRef]
- Tani, Y.; Yamamoto, H.; Kawaji, A.; Mizuno, H.; Fukushige, J.; Hosokawa, T.; Doi, K. Hepatic cytochrome P450 and flavin-containing monooxygenase in male Nts:Mini rat, a transgenic rat carrying antisense RNA transgene for rat growth hormone. Toxicol. Lett. 1999, 106, 159–169. [Google Scholar] [CrossRef]
- Takahashi, J.; Furuhata, Y.; Ikeda, A.; Takahashi, M.; Iwata, H.; Kazusaka, A.; Fujita, S. Characterization of hepatic cytochrome P450 isozyme composition in the transgenic rat expressing low level human growth hormone. Xenobiotica 1999, 29, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Son, M.H.; Kang, K.W.; Kim, E.J.; Ryu, J.H.; Cho, H.; Kim, S.H.; Kim, W.B.; Kim, S.G. Role of glucose utilization in the restoration of hypophysectomy-induced hepatic cytochrome P450 2E1 by growth hormone in rats. Chem. Interact. 2000, 127, 13–28. [Google Scholar] [CrossRef]
- Zhang, S.; Cawley, G.F.; Eyer, C.S.; Backes, W.L. Altered Ethylbenzene-Mediated Hepatic CYP2E1 Expression in Growth Hormone-Deficient Dwarf Rats. Toxicol. Appl. Pharmacol. 2002, 179, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Doody, E.E.; Groebner, J.L.; Walker, J.R.; Frizol, B.M.; Tuma, D.J.; Fernandez, D.J.; Tuma, P.L. Ethanol metabolism by alcohol dehydrogenase or cytochrome P450 2E1 differentially impairs hepatic protein trafficking and growth hormone signaling. Am. J. Physiol. Liver Physiol. 2017, 313, G558–G569. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.M.; Coon, M.J. Regulation of Rabbit Cytochrome P450 2E1 Expression in HepG2 Cells by Insulin and Thyroid Hor-mone. Mol. Pharmacol. 1998, 54, 740–747. [Google Scholar] [PubMed]
- Spina, E.; De Leon, J. Clinically relevant interactions between newer antidepressants and second-generation antipsychotics. Expert Opin. Drug Metab. Toxicol. 2014, 10, 721–746. [Google Scholar] [CrossRef]
- Strupczewski, J.T.; Bordeau, K.J.; Chiang, Y.; Glamkowski, E.J.; Conway, P.G.; Corbett, R.; Hartman, H.; Szewczak, M.R.; Wilmot, C.A.; Helsley, G.C. 3-[[(Aryloxy)alkyl]piperidinyl]-1,2-Benzisoxazoles as D2/5-HT2 Antagonists with Potential Atypical Antipsychotic Activity: Antipsychotic Profile of Iloperidone (HP 873). J. Med. Chem. 1995, 38, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Boyda, H.N.; Procyshyn, R.M.; Pang, C.C.Y.; Hawkes, E.; Wong, D.; Jin, C.H.; Honer, W.G.; Barr, A.M. Metabolic Side-Effects of the Novel Second-Generation Antipsychotic Drugs Asenapine and Iloperidone: A Comparison with Olanzapine. PLoS ONE 2013, 8, e53459. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.K.; Tarazi, F.I. Long-Term Effects of Iloperidone on Cerebral Serotonin and Adrenoceptor Subtypes. J. Mol. Neurosci. 2018, 66, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Daniel, W.A.; Haduch, A.; Wójcikowski, J. Inhibition and possible induction of rat CYP2D after short- and long-term treatment with antidepressants. J. Pharm. Pharmacol. 2002, 54, 1545–1552. [Google Scholar] [CrossRef]
- Kot, M.; Daniel, W.A. Relative contribution of rat cytochrome P450 isoforms to the metabolism of caffeine: The pathway and concentration dependence. Biochem. Pharmacol. 2008, 75, 1538–1549. [Google Scholar] [CrossRef]
- Daniel, W.A.; Haduch, A.; Syrek, M.; Boksa, J. Direct and indirect interactions between antidepressant drugs and CYP2C6 in the rat liver during long-term treatment. Eur. Neuropsychopharmacol. 2006, 16, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Haduch, A.; Wójcikowski, J.; Daniel, W.A. Effect of neuroleptics on cytochrome P450 2C11 (CYP2C11) in rat liver. Pharmacol. Rep. 2011, 63, 1491–1499. [Google Scholar] [CrossRef]
- Wójcikowski, J.; Haduch, A.; Daniel, W.A. Effect of classic and atypical neuroleptics on cytochrome P450 3A (CYP3A) in rat liver. Pharmacol. Rep. 2012, 64, 1411–1418. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danek, P.J.; Kuban, W.; Daniel, W.A. The Effect of Chronic Iloperidone Treatment on Cytochrome P450 Expression and Activity in the Rat Liver: Involvement of Neuroendocrine Mechanisms. Int. J. Mol. Sci. 2021, 22, 8447. https://doi.org/10.3390/ijms22168447
Danek PJ, Kuban W, Daniel WA. The Effect of Chronic Iloperidone Treatment on Cytochrome P450 Expression and Activity in the Rat Liver: Involvement of Neuroendocrine Mechanisms. International Journal of Molecular Sciences. 2021; 22(16):8447. https://doi.org/10.3390/ijms22168447
Chicago/Turabian StyleDanek, Przemysław J., Wojciech Kuban, and Władysława A. Daniel. 2021. "The Effect of Chronic Iloperidone Treatment on Cytochrome P450 Expression and Activity in the Rat Liver: Involvement of Neuroendocrine Mechanisms" International Journal of Molecular Sciences 22, no. 16: 8447. https://doi.org/10.3390/ijms22168447
APA StyleDanek, P. J., Kuban, W., & Daniel, W. A. (2021). The Effect of Chronic Iloperidone Treatment on Cytochrome P450 Expression and Activity in the Rat Liver: Involvement of Neuroendocrine Mechanisms. International Journal of Molecular Sciences, 22(16), 8447. https://doi.org/10.3390/ijms22168447