
Analysis of Spatial and Temporal Distribution of Purinergic P2 Receptors in the Mouse Hippocampus



Abstract
:1. Introduction
2. Results
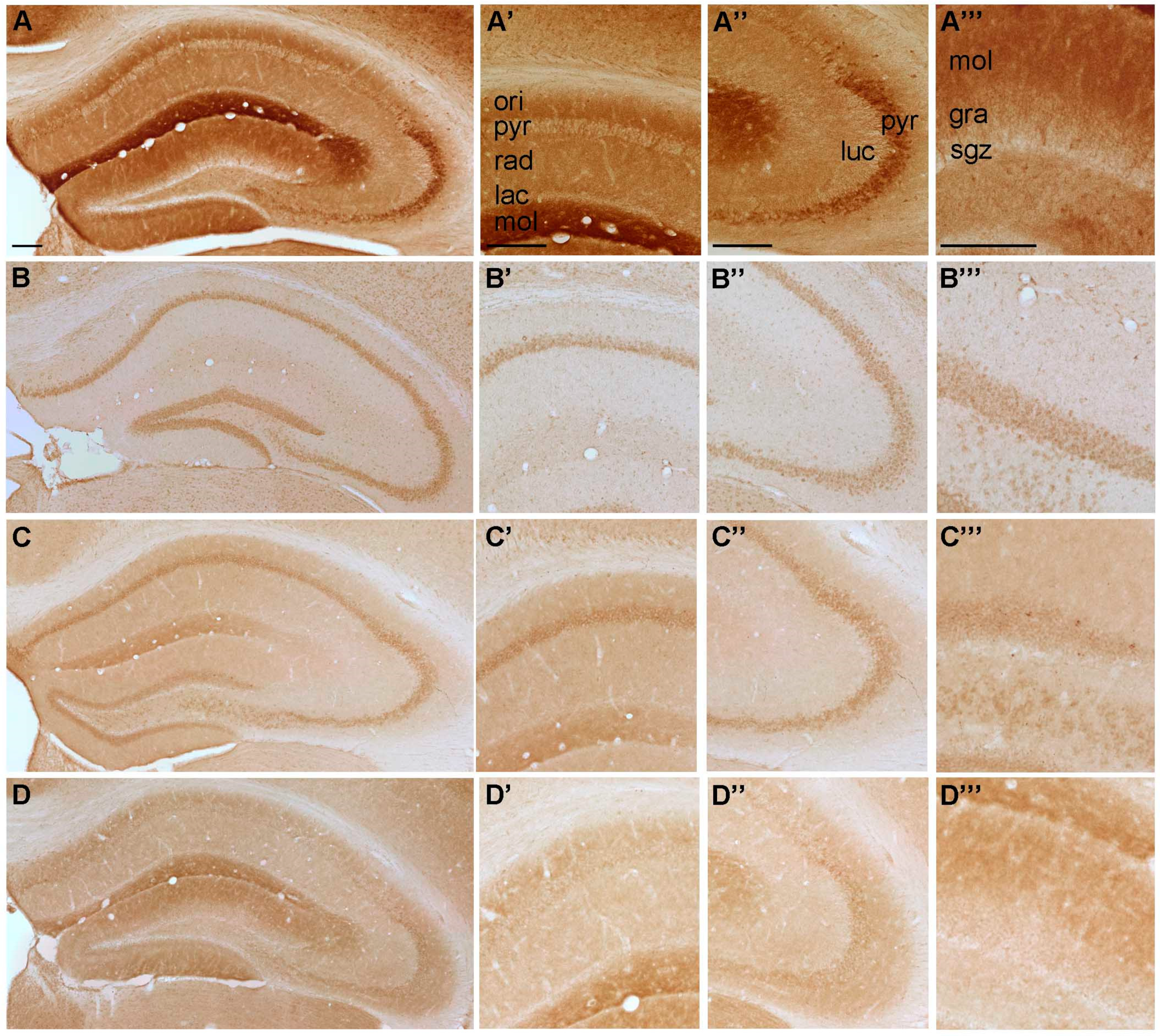
2.1. Spatial Distribution of P2-IR in the Hippocampus
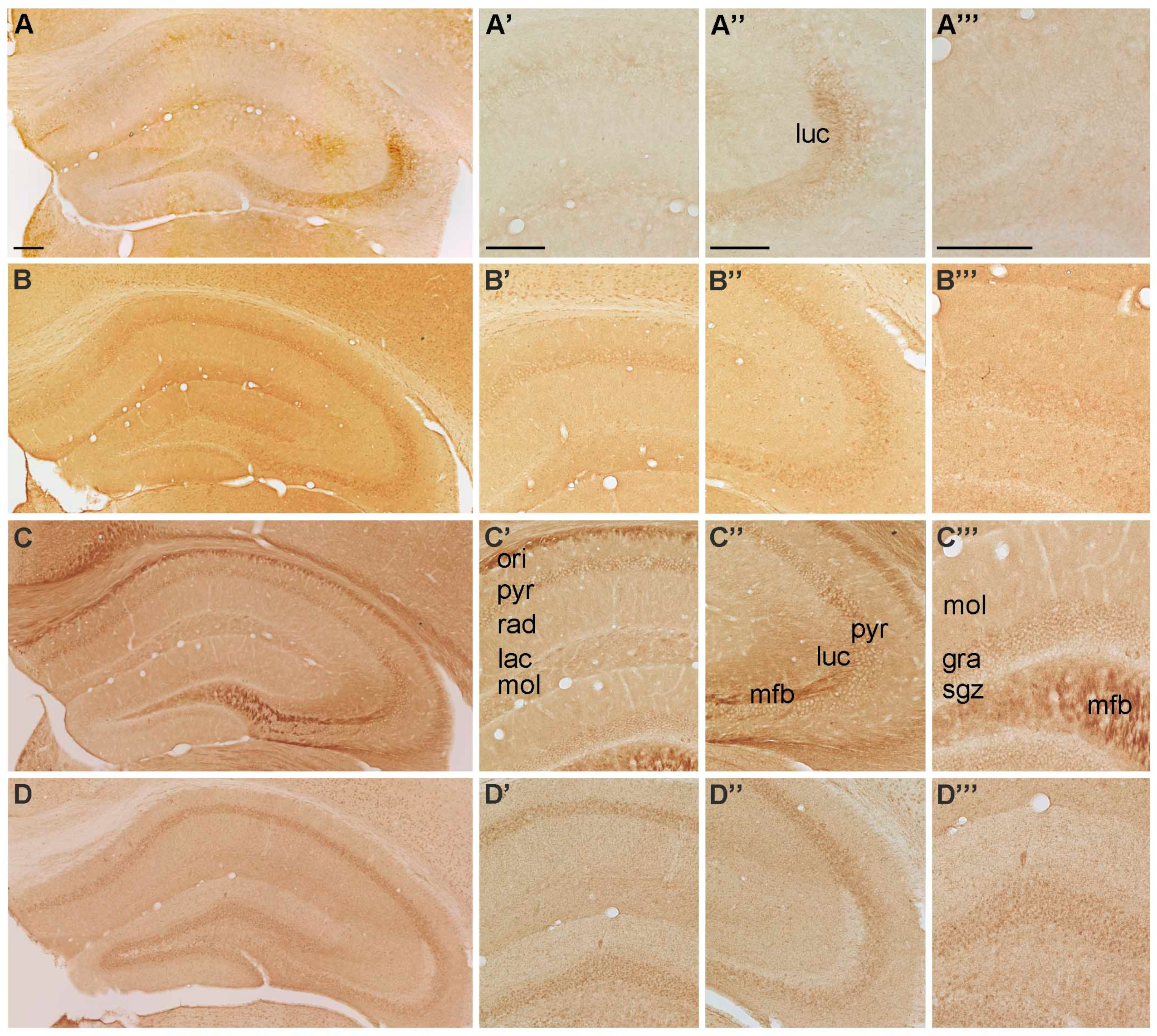
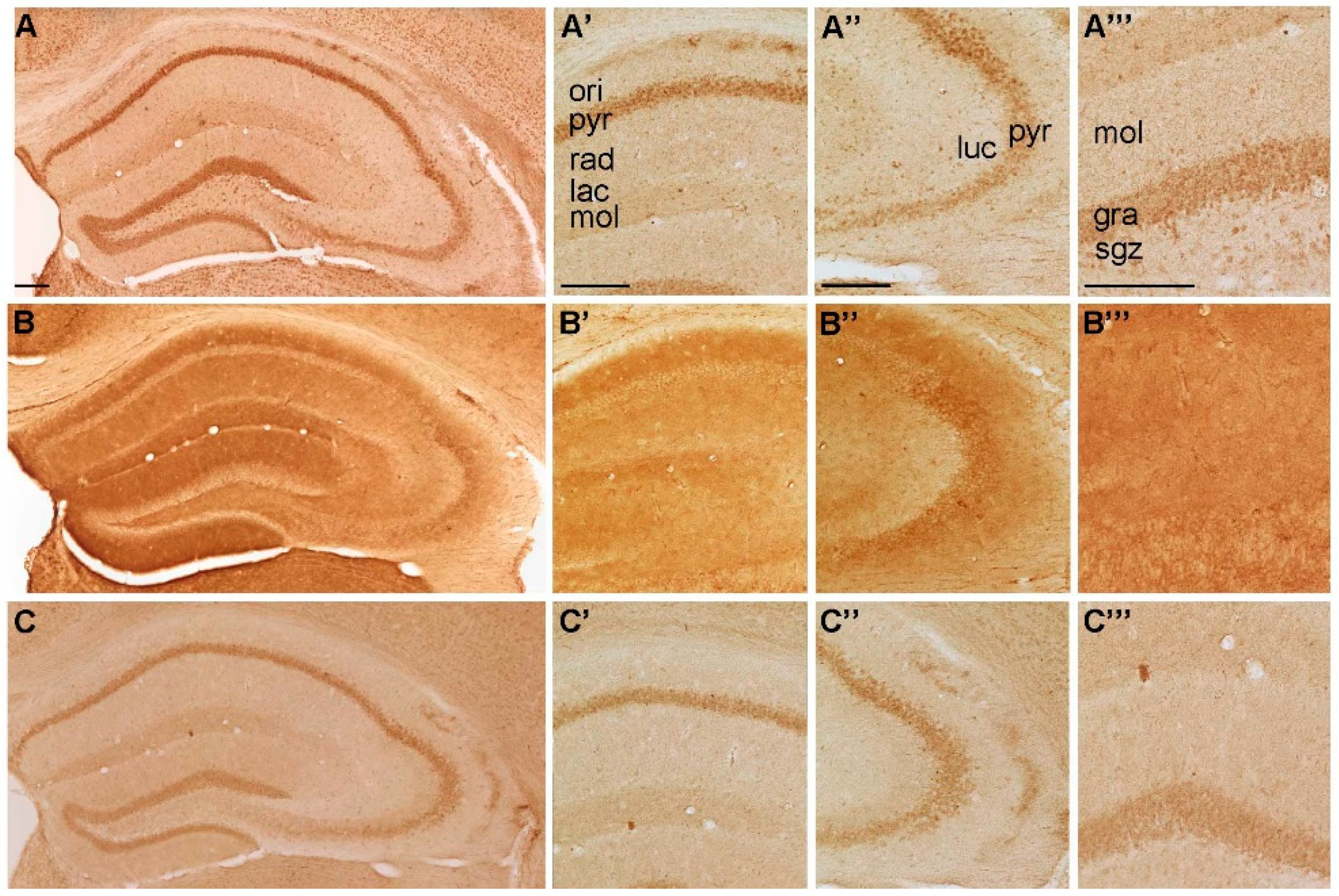
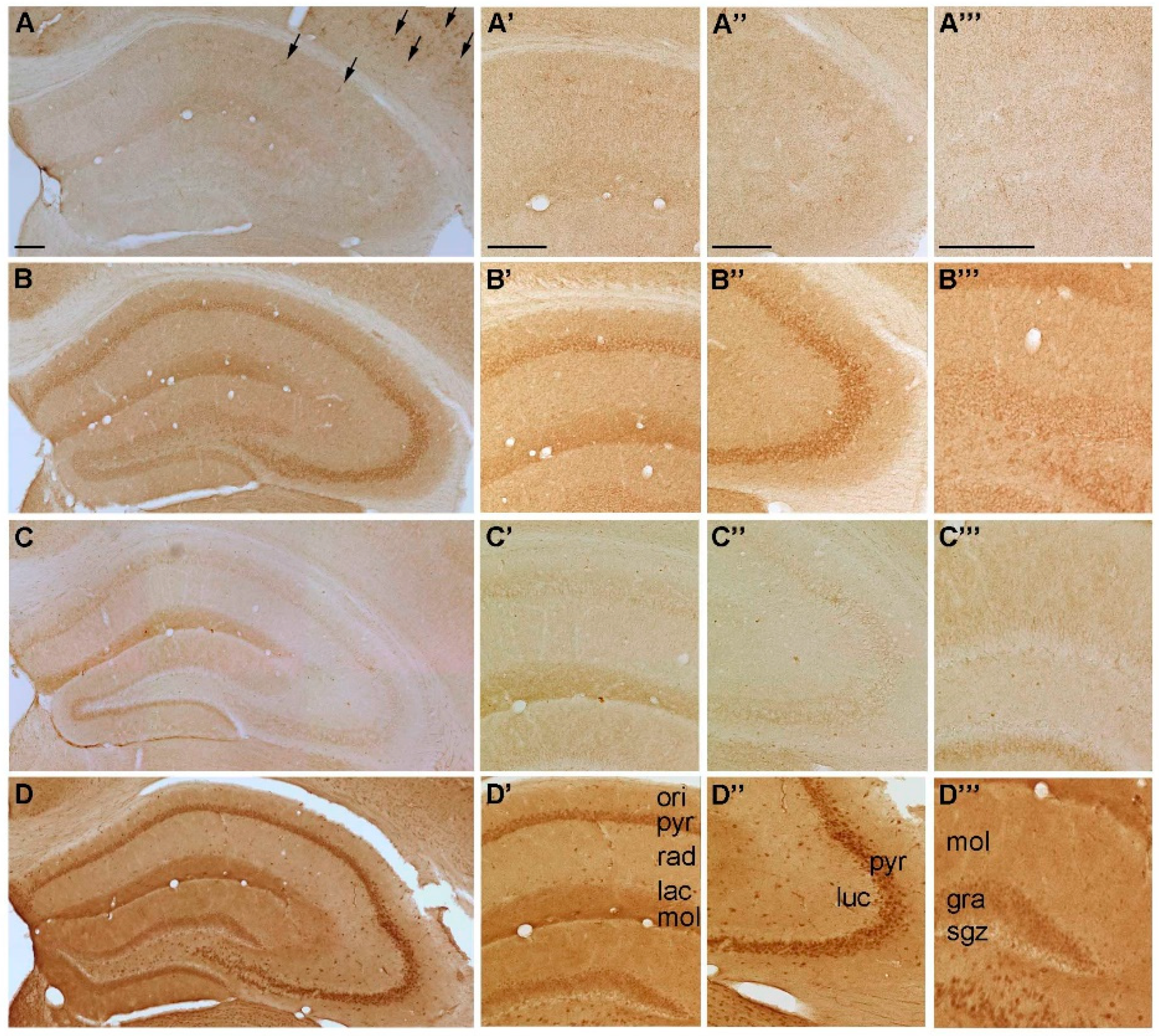
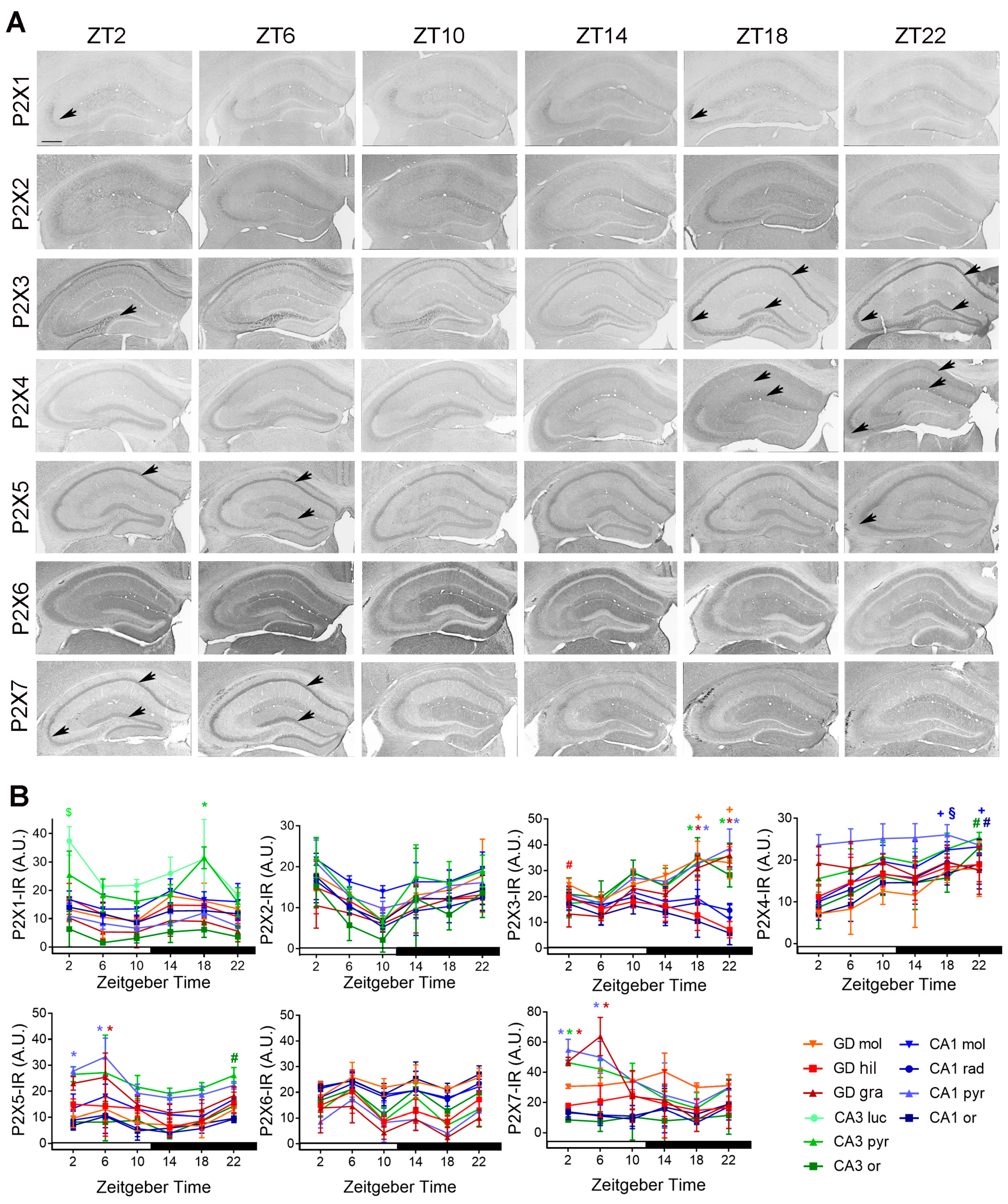
2.1.1. Spatial Distribution of P2X-IR
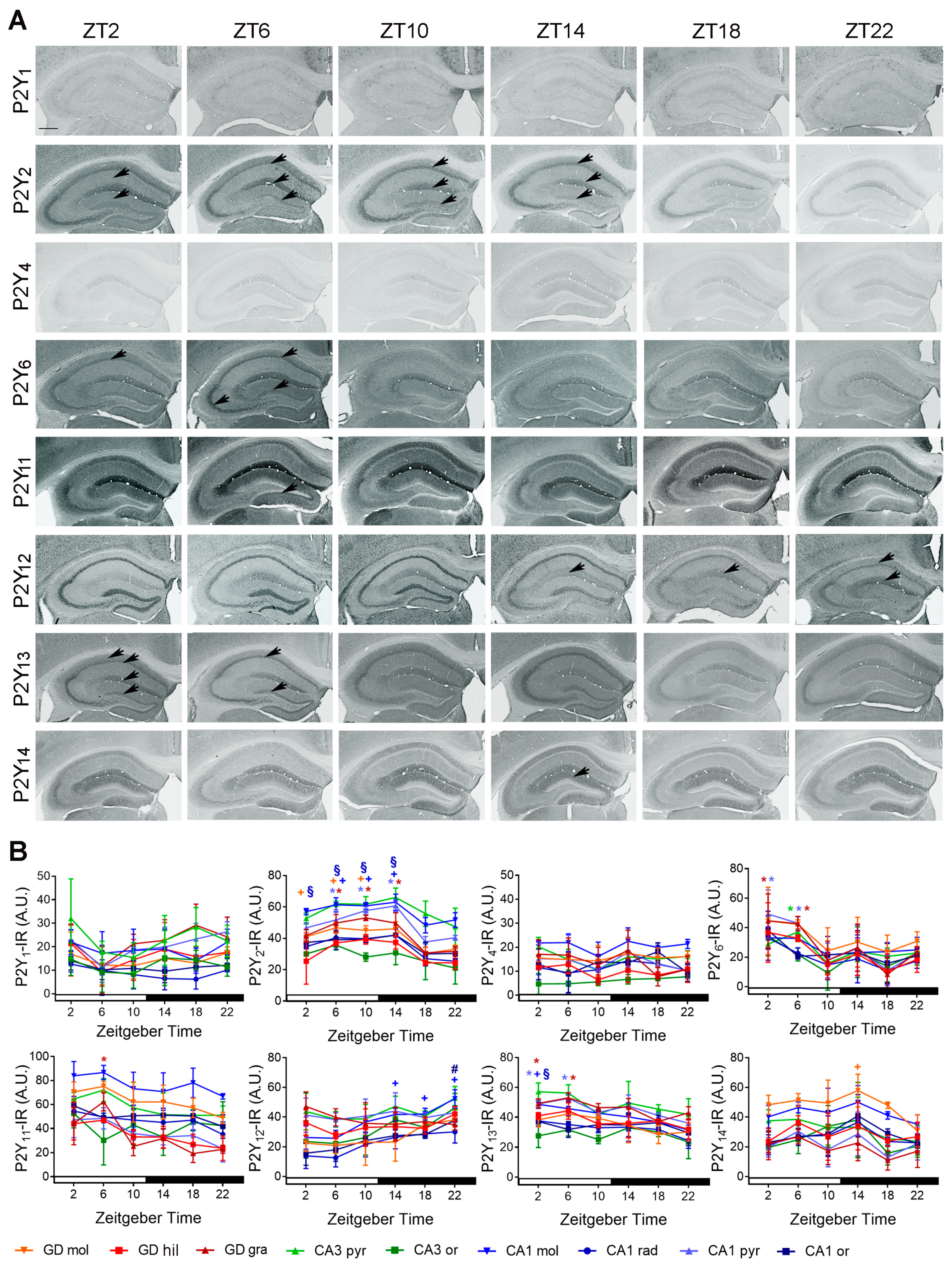
2.1.2. Spatial Distribution of P2Y-IR
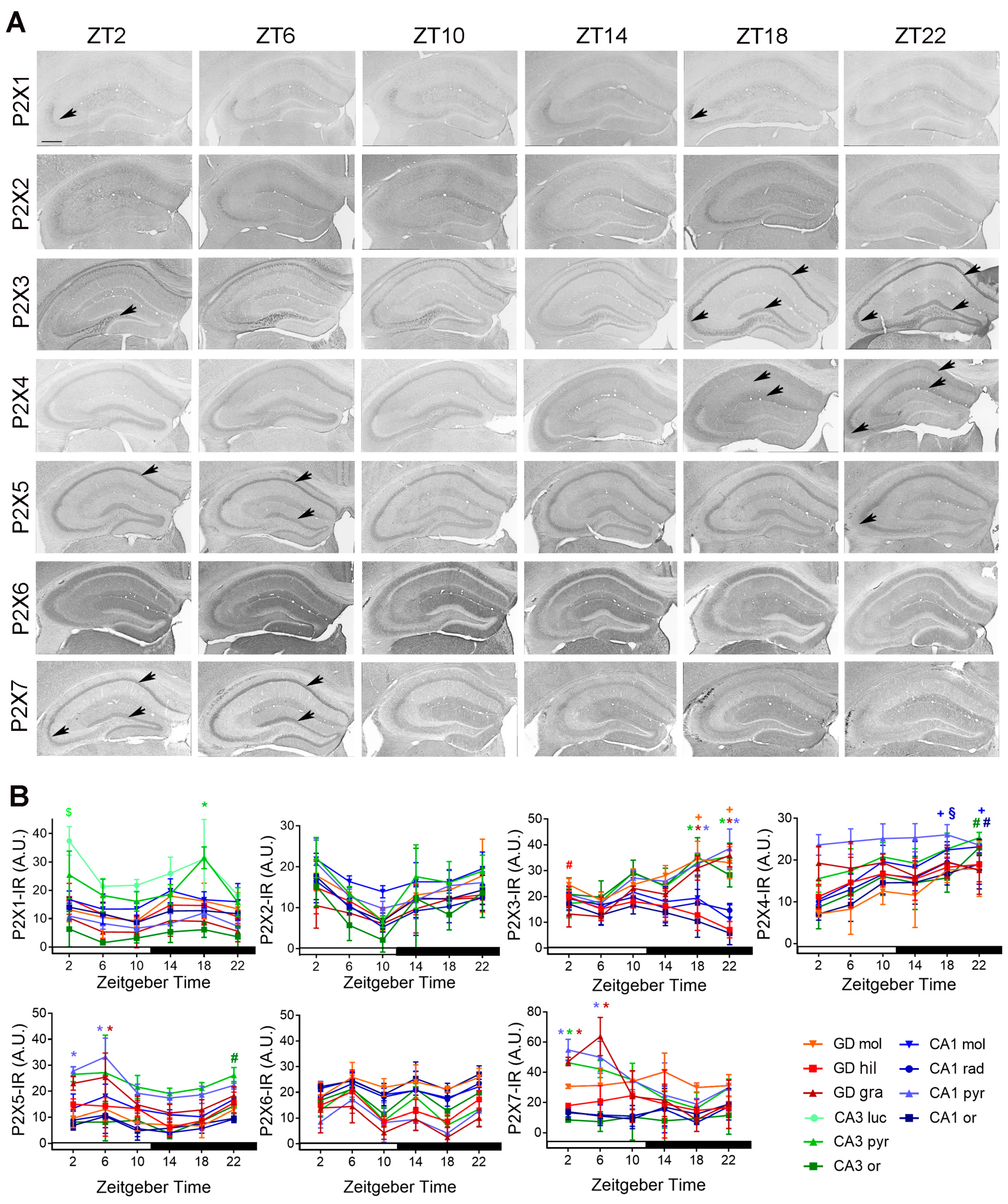
2.2. Temporal Distribution of P2-IR in the Hippocampus
2.2.1. Temporal Distribution of P2X-IR
2.2.2. Temporal Distribution of P2Y-IR
3. Discussion
4. Material and Methods
4.1. Animals and Tissue Processing
4.2. Brain Preparation, Immunohistochemistry and Timm Staining
4.3. Image Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, R.J.; Derkach, V.; Surprenant, A. ATP mediates fast synaptic transmission in mammalian neurons. Nature 1992, 357, 503–505. [Google Scholar] [CrossRef]
- Silinsky, E.M.; Gerzanich, V.; Vanner, S.M. ATP mediates excitatory synaptic transmission in mammalian neurones. Br. J. Pharmacol. 1992, 106, 762–763. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G.; Kennedy, C. Is there a basis for distinguishing two types of P2-purinoceptor? Gen. Pharmacol. 1985, 16, 433–440. [Google Scholar] [CrossRef]
- Koles, L.; Kato, E.; Hanuska, A.; Zadori, Z.S.; Al-Khrasani, M.; Zelles, T.; Rubini, P.; Illes, P. Modulation of excitatory neurotransmission by neuronal/glial signalling molecules: Interplay between purinergic and glutamatergic systems. Purinergic Signal. 2016, 12, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G. The past, present and future of purine nucleotides as signalling molecules. Neuropharmacology 1997, 36, 1127–1139. [Google Scholar] [CrossRef]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta 2008, 1783, 673–694. [Google Scholar] [CrossRef] [Green Version]
- Lazarowski, E.R.; Sesma, J.I.; Seminario-Vidal, L.; Kreda, S.M. Molecular mechanisms of purine and pyrimidine nucleotide release. Adv. Pharmacol. 2011, 61, 221–261. [Google Scholar] [CrossRef]
- Lazarowski, E.R. Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal. 2012, 8, 359–373. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G.; Fredholm, B.B.; Verkhratsky, A. Adenosine and ATP receptors in the brain. Curr. Top. Med. Chem. 2011, 11, 973–1011. [Google Scholar] [CrossRef]
- Fields, R.D.; Burnstock, G. Purinergic signalling in neuron-glia interactions. Nat. Rev. Neurosci. 2006, 7, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Abbracchio, M.P.; Burnstock, G. Purinergic signalling: Pathophysiological roles. Jpn. J. Pharmacol. 1998, 78, 113–145. [Google Scholar] [CrossRef] [Green Version]
- Neary, J.T.; Rathbone, M.P.; Cattabeni, F.; Abbracchio, M.P.; Burnstock, G. Trophic actions of extracellular nucleotides and nucleosides on glial and neuronal cells. Trends Neurosci. 1996, 19, 13–18. [Google Scholar] [CrossRef]
- Burnstock, G.; Knight, G.E. Cellular distribution and functions of P2 receptor subtypes in different systems. Int. Rev. Cytol. 2004, 240, 31–304. [Google Scholar]
- Mishra, S.K.; Braun, N.; Shukla, V.; Füllgabe, M.; Schomerus, C.; Korf, H.W.; Gachet, C.; Ikehara, Y.; Sevigny, J.; Robson, S.C.; et al. Extracellular nucleotide signaling in adult neural stem cells: Synergism with growth factor-mediated cellular proliferation. Development 2006, 133, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Illes, P.; Xu, G.Y.; Tang, Y. Purinergic Signaling in the Central Nervous System in Health and Disease. Neurosci. Bull. 2020, 36, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Illes, P.; Alexandre Ribeiro, J. Molecular physiology of P2 receptors in the central nervous system. Eur. J. Pharmacol. 2004, 483, 5–17. [Google Scholar] [CrossRef]
- Lommen, J.; Stahr, A.; Ingenwerth, M.; Ali, A.A.H.; von Gall, C. Time-of-day-dependent expression of purinergic receptors in mouse suprachiasmatic nucleus. Cell Tissue Res. 2017, 369, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.A.H.; Avakian, G.A.; Gall, C.V. The Role of Purinergic Receptors in the Circadian System. Int. J. Mol. Sci. 2020, 21, 3423. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.A.H.; Stahr, A.; Ingenwerth, M.; Theis, M.; Steinhäuser, C.; von Gall, C. Connexin30 and Connexin43 show a time-of-day dependent expression in the mouse suprachiasmatic nucleus and modulate rhythmic locomotor activity in the context of chronodisruption. Cell Commun. Signal. 2019, 17, 61. [Google Scholar] [CrossRef] [Green Version]
- Marpegan, L.; Swanstrom, A.E.; Chung, K.; Simon, T.; Haydon, P.G.; Khan, S.K.; Liu, A.C.; Herzog, E.D.; Beaule, C. Circadian regulation of ATP release in astrocytes. J. Neurosci. 2011, 31, 8342–8350. [Google Scholar] [CrossRef] [Green Version]
- Womac, A.D.; Burkeen, J.F.; Neuendorff, N.; Earnest, D.J.; Zoran, M.J. Circadian rhythms of extracellular ATP accumulation in suprachiasmatic nucleus cells and cultured astrocytes. Eur. J. Neurosci. 2009, 30, 869–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, A.; Vavra, V.; Svobodova, I.; Bendova, Z.; Vereb, G.; Zemkova, H. Potentiation of inhibitory synaptic transmission by extracellular ATP in rat suprachiasmatic nuclei. J. Neurosci. 2013, 33, 8035–8044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squire, L.R.; Wixted, J.T. The cognitive neuroscience of human memory since H.M. Annu. Rev. Neurosci. 2011, 34, 259–288. [Google Scholar] [CrossRef] [Green Version]
- Smarr, B.L.; Jennings, K.J.; Driscoll, J.R.; Kriegsfeld, L.J. A time to remember: The role of circadian clocks in learning and memory. Behav. Neurosci. 2014, 128, 283–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitti, F.L.; Siegelbaum, S.A. The hippocampal CA2 region is essential for social memory. Nature 2014, 508, 88–92. [Google Scholar] [CrossRef]
- van Strien, N.M.; Cappaert, N.L.; Witter, M.P. The anatomy of memory: An interactive overview of the parahippocampal-hippocampal network. Nat. Rev. Neurosci. 2009, 10, 272–282. [Google Scholar] [CrossRef]
- Rager, G.; Braak, W.Z.H.; Braak, E.; Nitsch, R.; Endhirn, E.A. Anatomie; Drenkhahn, D., Ed.; Elsevier: München, Germany, 2004. [Google Scholar]
- Florenzano, F.; Viscomi, M.T.; Mercaldo, V.; Longone, P.; Bernardi, G.; Bagni, C.; Molinari, M.; Carrive, P. P2X2R purinergic receptor subunit mRNA and protein are expressed by all hypothalamic hypocretin/orexin neurons. J. Comp. Neurol. 2006, 498, 58–67. [Google Scholar] [CrossRef]
- Xiang, Z.; Bo, X.; Oglesby, I.; Ford, A.; Burnstock, G. Localization of ATP-gated P2X2 receptor immunoreactivity in the rat hypothalamus. Brain Res. 1998, 813, 390–397. [Google Scholar] [CrossRef]
- Loesch, A.; Miah, S.; Burnstock, G. Ultrastructural localisation of ATP-gated P2X2 receptor immunoreactivity in the rat hypothalamo-neurohypophysial system. J. Neurocytol. 1999, 28, 495–504. [Google Scholar] [CrossRef]
- Chen, C.C.; Akopian, A.N.; Sivilotti, L.; Colquhoun, D.; Burnstock, G.; Wood, J.N. A P2X purinoceptor expressed by a subset of sensory neurons. Nature 1995, 377, 428–431. [Google Scholar] [CrossRef]
- Lewis, C.; Neidhart, S.; Holy, C.; North, R.A.; Buell, G.; Surprenant, A. Coexpression of P2X2 and P2X3 receptor subunits can account for ATP-gated currents in sensory neurons. Nature 1995, 377, 432–435. [Google Scholar] [CrossRef]
- Rubio, M.E.; Soto, F. Distinct Localization of P2X receptors at excitatory postsynaptic specializations. J. Neurosci. 2001, 21, 641–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran-Jimenez, M.J.; Matute, C. Immunohistochemical localization of the P2Y(1) purinergic receptor in neurons and glial cells of the central nervous system. Brain Res. Mol. Brain Res. 2000, 78, 50–58. [Google Scholar] [CrossRef]
- Communi, D.; Gonzalez, N.S.; Detheux, M.; Brezillon, S.; Lannoy, V.; Parmentier, M.; Boeynaems, J.M. Identification of a novel human ADP receptor coupled to G(i). J. Biol. Chem. 2001, 276, 41479–41485. [Google Scholar] [CrossRef] [Green Version]
- Laitinen, J.T.; Uri, A.; Raidaru, G.; Miettinen, R. [(35)S]GTPgammaS autoradiography reveals a wide distribution of G(i/o)-linked ADP receptors in the nervous system: Close similarities with the platelet P2Y(ADP) receptor. J. Neurochem. 2001, 77, 505–518. [Google Scholar] [CrossRef]
- Rodrigues, R.J.; Almeida, T.; Díaz-Hernández, M.; Marques, J.M.; Franco, R.; Solsona, C.; Miras-Portugal, M.T.; Ciruela, F.; Cunha, R.A. Presynaptic P2X1-3 and α3-containing nicotinic receptors assemble into functionally interacting ion channels in the rat hippocampus. Neuropharmacology 2016, 105, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Pankratov, Y.V.; Lalo, U.V.; Krishtal, O.A. Role for P2X receptors in long-term potentiation. J. Neurosci. 2002, 22, 8363–8369. [Google Scholar] [CrossRef]
- Burnstock, G. Physiopathological roles of P2X receptors in the central nervous system. Curr. Med. Chem. 2015, 22, 819–844. [Google Scholar] [CrossRef] [PubMed]
- Guzman, S.J.; Gerevich, Z. P2Y Receptors in Synaptic Transmission and Plasticity: Therapeutic Potential in Cognitive Dysfunction. Neural. Plast. 2016, 2016, 1207393. [Google Scholar] [CrossRef] [Green Version]
- Del Puerto, A.; Wandosell, F.; Garrido, J.J. Neuronal and glial purinergic receptors functions in neuron development and brain disease. Front. Cell Neurosci. 2013, 7, 197. [Google Scholar] [CrossRef] [Green Version]
- Tsao, A.; Sugar, J.; Lu, L.; Wang, C.; Knierim, J.J.; Moser, M.B.; Moser, E.I. Integrating time from experience in the lateral entorhinal cortex. Nature 2018, 561, 57–62. [Google Scholar] [CrossRef]
- Witter, M.P.; Naber, P.A.; van Haeften, T.; Machielsen, W.C.; Rombouts, S.A.; Barkhof, F.; Scheltens, P.; Lopes da Silva, F.H. Cortico-hippocampal communication by way of parallel parahippocampal-subicular pathways. Hippocampus 2000, 10, 398–410. [Google Scholar] [CrossRef]
- Weng, F.J.; Garcia, R.I.; Lutzu, S.; Alvina, K.; Zhang, Y.; Dushko, M.; Ku, T.; Zemoura, K.; Rich, D.; Garcia-Dominguez, D.; et al. Npas4 Is a Critical Regulator of Learning-Induced Plasticity at Mossy Fiber-CA3 Synapses during Contextual Memory Formation. Neuron 2018. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, R.J.; Almeida, T.; Richardson, P.J.; Oliveira, C.R.; Cunha, R.A. Dual presynaptic control by ATP of glutamate release via facilitatory P2X1, P2X2/3, and P2X3 and inhibitory P2Y1, P2Y2, and/or P2Y4 receptors in the rat hippocampus. J. Neurosci. 2005, 25, 6286–6295. [Google Scholar] [CrossRef]
- Seguela, P.; Haghighi, A.; Soghomonian, J.J.; Cooper, E. A novel neuronal P2x ATP receptor ion channel with widespread distribution in the brain. J. Neurosci. 1996, 16, 448–455. [Google Scholar] [CrossRef] [Green Version]
- George, J.; Cunha, R.A.; Mulle, C.; Amedee, T. Microglia-derived purines modulate mossy fibre synaptic transmission and plasticity through P2X4 and A1 receptors. Eur. J. Neurosci. 2016, 43, 1366–1378. [Google Scholar] [CrossRef]
- Remondes, M.; Schuman, E.M. Role for a cortical input to hippocampal area CA1 in the consolidation of a long-term memory. Nature 2004, 431, 699–703. [Google Scholar] [CrossRef]
- Kallarackal, A.J.; Kvarta, M.D.; Cammarata, E.; Jaberi, L.; Cai, X.; Bailey, A.M.; Thompson, S.M. Chronic stress induces a selective decrease in AMPA receptor-mediated synaptic excitation at hippocampal temporoammonic-CA1 synapses. J. Neurosci. 2013, 33, 15669–15674. [Google Scholar] [CrossRef] [Green Version]
- Scimemi, A.; Schorge, S.; Kullmann, D.M.; Walker, M.C. Epileptogenesis is associated with enhanced glutamatergic transmission in the perforant path. J. Neurophysiol. 2006, 95, 1213–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.A.H.; Abdel-Hafiz, L.; Tundo-Lavalle, F.; Hassan, S.A.; von Gall, C. P2Y2 deficiency impacts adult neurogenesis and related forebrain functions. FASEB J. 2021, 35, e21546. [Google Scholar] [CrossRef] [PubMed]
- Kukley, M.; Barden, J.A.; Steinhauser, C.; Jabs, R. Distribution of P2X receptors on astrocytes in juvenile rat hippocampus. Glia 2001, 36, 11–21. [Google Scholar] [CrossRef]
- Metzger, M.W.; Walser, S.M.; Aprile-Garcia, F.; Dedic, N.; Chen, A.; Holsboer, F.; Arzt, E.; Wurst, W.; Deussing, J.M. Genetically dissecting P2rx7 expression within the central nervous system using conditional humanized mice. Purinergic Signal. 2017, 13, 153–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boue-Grabot, E.; Pankratov, Y. Modulation of Central Synapses by Astrocyte-Released ATP and Postsynaptic P2X Receptors. Neural. Plast. 2017, 2017, 9454275. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, M.; Gachet, C.; Inoue, K.; Kato, F. Direct excitation of inhibitory interneurons by extracellular ATP mediated by P2Y1 receptors in the hippocampal slice. J. Neurosci. 2004, 24, 10835–10845. [Google Scholar] [CrossRef] [Green Version]
- Eckel-Mahan, K.L. Circadian Oscillations within the Hippocampus Support Memory Formation and Persistence. Front. Mol. Neurosci. 2012, 5, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhury, D.; Wang, L.M.; Colwell, C.S. Circadian regulation of hippocampal long-term potentiation. J. Biol. Rhythms. 2005, 20, 225–236. [Google Scholar] [CrossRef]
- Cao, X.; Li, L.P.; Wang, Q.; Wu, Q.; Hu, H.H.; Meng, Z.; Fang, Y.Y.; Zhang, J.; Li, S.J.; Xiong, W.C.; et al. Astrocyte-derived ATP modulates depressive-like behaviors. Nat. Med. 2013, 19, 773–777. [Google Scholar] [CrossRef]
- Dworak, M.; McCarley, R.W.; Kim, T.; Kalinchuk, A.V.; Basheer, R. Sleep and brain energy levels: ATP changes during sleep. J. Neurosci. 2010, 30, 9007–9016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sloviter, R.S. A simplified Timm stain procedure compatible with formaldehyde fixation and routine paraffin embedding of rat brain. Brain Res. Bull. 1982, 8, 771–774. [Google Scholar] [CrossRef]
- Longhurst, P.A.; Schwegel, T.; Folander, K.; Swanson, R. The human P2x1 receptor: Molecular cloning, tissue distribution, and localization to chromosome 17. Biochim. Biophys. Acta 1996, 1308, 185–188. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Chiozzi, P.; Falzoni, S.; Ferrari, D.; Sanz, J.M.; Venketaraman, V.; Baricordi, O.R. Cytolytic P2X purinoceptors. Cell Death Differ. 1998, 5, 191–199. [Google Scholar] [CrossRef]
- Alexander, K.; Niforatos, W.; Bianchi, B.; Burgard, E.C.; Lynch, K.J.; Kowaluk, E.A.; Jarvis, M.F.; van Biesen, T. Allosteric modulation and accelerated resensitization of human P2X(3) receptors by cibacron blue. J. Pharmacol. Exp. Ther. 1999, 291, 1135–1142. [Google Scholar]
- Burnstock, G. P2X receptors in sensory neurones. Br. J. Anaesth. 2000, 84, 476–488. [Google Scholar] [CrossRef]
- Oury, C.; Toth-Zsamboki, E.; Van Geet, C.; Thys, C.; Wei, L.; Nilius, B.; Vermylen, J.; Hoylaerts, M.F. A natural dominant negative P2X1 receptor due to deletion of a single amino acid residue. J. Biol. Chem. 2000, 275, 22611–22614. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Sachs, F. Inactivation of P2X2 purinoceptors by divalent cations. J. Physiol. 2000, 522 Pt 2, 199–214. [Google Scholar] [CrossRef]
- North, R.A.; Surprenant, A. Pharmacology of cloned P2X receptors. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 563–580. [Google Scholar] [CrossRef] [PubMed]
- Wirkner, K.; Stanchev, D.; Milius, D.; Hartmann, L.; Kato, E.; Zadori, Z.S.; Mager, P.P.; Rubini, P.; Nörenberg, W.; Illes, P. Regulation of the pH sensitivity of human P2X receptors by N-linked glycosylation. J. Neurochem. 2008, 107, 1216–1224. [Google Scholar] [CrossRef] [PubMed]
- Stanchev, D.; Blosa, M.; Milius, D.; Gerevich, Z.; Rubini, P.; Schmalzing, G.; Eschrich, K.; Schaefer, M.; Wirkner, K.; Illes, P. Cross-inhibition between native and recombinant TRPV1 and P2X(3) receptors. Pain 2009, 143, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Simonetti, M.; Birsa, N.; Ferrari, M.D.; van den Maagdenberg, A.M.J.M.; Giniatullin, R.; Nistri, A.; Fabbretti, E. Familial hemiplegic migraine Ca(v)2.1 channel mutation R192Q enhances ATP-gated P2X3 receptor activity of mouse sensory ganglion neurons mediating trigeminal pain. Mol. Pain 2010, 6, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, A.R.; Chavez-Valdez, R.; Ezell, T.; Donnelly, D.F.; Glover, J.C.; Gauda, E.B. Effect of development on [Ca2+]i transients to ATP in petrosal ganglion neurons: A pharmacological approach using optical recording. J. Appl. Physiol. 2012, 112, 1393–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnanasekaran, A.; Sundukova, M.; Hullugundi, S.; Birsa, N.; Bianchini, G.; Hsueh, Y.P.; Nistri, A.; Fabbretti, E. Calcium/calmodulin-dependent serine protein kinase (CASK) is a new intracellular modulator of P2X3 receptors. J. Neurochem. 2013, 126, 102–112. [Google Scholar] [CrossRef]
- Svennersten, K.; Hallén-Grufman, K.; de Verdier, P.J.; Wiklund, N.P.; Poljakovic, M. Localization of P2X receptor subtypes 2, 3 and 7 in human urinary bladder. BMC Urol. 2015, 15, 81. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.R.; Krabbe, S.; Novak, I. Purinergic receptors and calcium signalling in human pancreatic duct cell lines. Cell Physiol. Biochem. 2008, 22, 157–168. [Google Scholar] [CrossRef]
- Yeung, D.; Zablocki, K.; Lien, C.F.; Jiang, T.; Arkle, S.; Brutkowski, W.; Brown, J.; Lochmuller, H.; Simon, J.; Barnard, E.A.; et al. Increased susceptibility to ATP via alteration of P2X receptor function in dystrophic mdx mouse muscle cells. FASEB J. 2006, 20, 610–620. [Google Scholar] [CrossRef]
- Bulanova, E.; Budagian, V.; Orinska, Z.; Hein, M.; Petersen, F.; Thon, L.; Adam, D.; Bulfone-Paus, S. Extracellular ATP induces cytokine expression and apoptosis through P2X7 receptor in murine mast cells. J. Immunol. 2005, 174, 3880–3890. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.J.; Zheng, G.G.; Ma, X.T.; Yang, Y.H.; Li, G.; Rao, Q.; Nie, K.; Wu, K.F. Expression of P2X7 in human hematopoietic cell lines and leukemia patients. Leuk. Res. 2004, 28, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Budagian, V.; Bulanova, E.; Brovko, L.; Orinska, Z.; Fayad, R.; Paus, R.; Bulfone-Paus, S. Signaling through P2X7 receptor in human T cells involves p56lck, MAP kinases, and transcription factors AP-1 and NF-kappa B. J. Biol. Chem. 2003, 278, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.B.; Turner, J.J.O.; Fountain, S.J. Constitutive P2Y(2) receptor activity regulates basal lipolysis in human adipocytes. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.B.; Jung, S.R.; Hille, B.; Koh, D.S. Extracellular ATP protects pancreatic duct epithelial cells from alcohol-induced damage through P2Y1 receptor-cAMP signal pathway. Cell Biol. Toxicol. 2016, 32, 229–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, C.; Wegner, A.; Grosche, J.; Allgaier, C.; Illes, P.; Franke, H. P2 receptor expression in the dopaminergic system of the rat brain during development. Neuroscience 2007, 149, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Jelcic, M.; Wang, K.; Hui, K.L.; Cai, X.C.; Enyedi, B.; Luo, M.; Niethammer, P. A Photo-clickable ATP-Mimetic Reveals Nucleotide Interactors in the Membrane Proteome. Cell Chem. Biol. 2020, 27, 1073–1083.e12. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Miranda, E.; Molina-Aguilar, C.; González-Gallardo, A.; Vázquez-Martínez, O.; Díaz-Muñoz, M.; Vázquez-Cuevas, F.G. Increased Purinergic Responses Dependent on P2Y2 Receptors in Hepatocytes from CCl(4)-Treated Fibrotic Mice. Int. J. Mol. Sci. 2020, 21, 2305. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro Xavier, A.L.; Kress, B.T.; Goldman, S.A.; Lacerda de Menezes, J.R.; Nedergaard, M. A Distinct Population of Microglia Supports Adult Neurogenesis in the Subventricular Zone. J. Neurosci. 2015, 35, 11848–11861. [Google Scholar] [CrossRef]
- Fries, J.E.; Goczalik, I.M.; Wheeler-Schilling, T.H.; Kohler, K.; Guenther, E.; Wolf, S.; Wiedemann, P.; Bringmann, A.; Reichenbach, A.; Francke, M.; et al. Identification of P2Y receptor subtypes in human muller glial cells by physiology, single cell RT-PCR, and immunohistochemistry. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3000–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helenius, M.H.; Vattulainen, S.; Orcholski, M.; Aho, J.; Komulainen, A.; Taimen, P.; Wang, L.; de Jesus Perez, V.A.; Koskenvuo, J.W.; Alastalo, T.P. Suppression of endothelial CD39/ENTPD1 is associated with pulmonary vascular remodeling in pulmonary arterial hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 308, L1046–L1057. [Google Scholar] [CrossRef] [PubMed]
- Casieri, V.; Matteucci, M.; Pasanisi, E.M.; Papa, A.; Barile, L.; Fritsche-Danielson, R.; Lionetti, V. Ticagrelor Enhances Release of Anti-Hypoxic Cardiac Progenitor Cell-Derived Exosomes Through Increasing Cell Proliferation In Vitro. Sci. Rep. 2020, 10, 2494. [Google Scholar] [CrossRef]
- Yu, W.; Hill, W.G.; Robson, S.C.; Zeidel, M.L. Role of P2X4 Receptor in Mouse Voiding Function. Sci. Rep. 2018, 8, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, A.; Pike, C.J. Staining and Quantification of β-Amyloid Pathology in Transgenic Mouse Models of Alzheimer’s Disease. Methods Mol. Biol. 2020, 2144, 211–221. [Google Scholar] [CrossRef]
- Nógrádi, B.; Nyúl-Tóth, Á.; Kozma, M.; Molnar, K.; Patai, R.; Siklos, L.; Wilhelm, I.; Krizbai, I.A. Upregulation of Nucleotide-Binding Oligomerization Domain-, LRR- and Pyrin Domain-Containing Protein 3 in Motoneurons Following Peripheral Nerve Injury in Mice. Front. Pharmacol. 2020, 11, 584184. [Google Scholar] [CrossRef] [PubMed]
- Earnest, D.J.; Iadarola, M.; Yeh, H.H.; Olschowka, J.A. Photic regulation of c-fos expression in neural components governing the entrainment of circadian rhythms. Exp. Neurol. 1990, 109, 353–361. [Google Scholar] [CrossRef]
- Greenberg, M.E.; Ziff, E.B.; Greene, L.A. Stimulation of neuronal acetylcholine receptors induces rapid gene transcription. Science 1986, 234, 80–83. [Google Scholar] [CrossRef]
- Szekely, A.M.; Barbaccia, M.L.; Alho, H.; Costa, E. In primary cultures of cerebellar granule cells the activation of N-methyl-D-aspartate-sensitive glutamate receptors induces c-fos mRNA expression. Mol. Pharmacol. 1989, 35, 401–408. [Google Scholar] [PubMed]
- Brancaccio, M.; Patton, A.P.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Astrocytes Control Circadian Timekeeping in the Suprachiasmatic Nucleus via Glutamatergic Signaling. Neuron 2017, 93, 1420–1435.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, G.; Butt, A.M. P2Y and P2X purinoceptor mediated Ca2+ signalling in glial cell pathology in the central nervous system. Eur. J. Pharmacol. 2002, 447, 247–260. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Krishtal, O.A.; Burnstock, G. Purinoceptors on neuroglia. Mol. Neurobiol. 2009, 39, 190–208. [Google Scholar] [CrossRef]
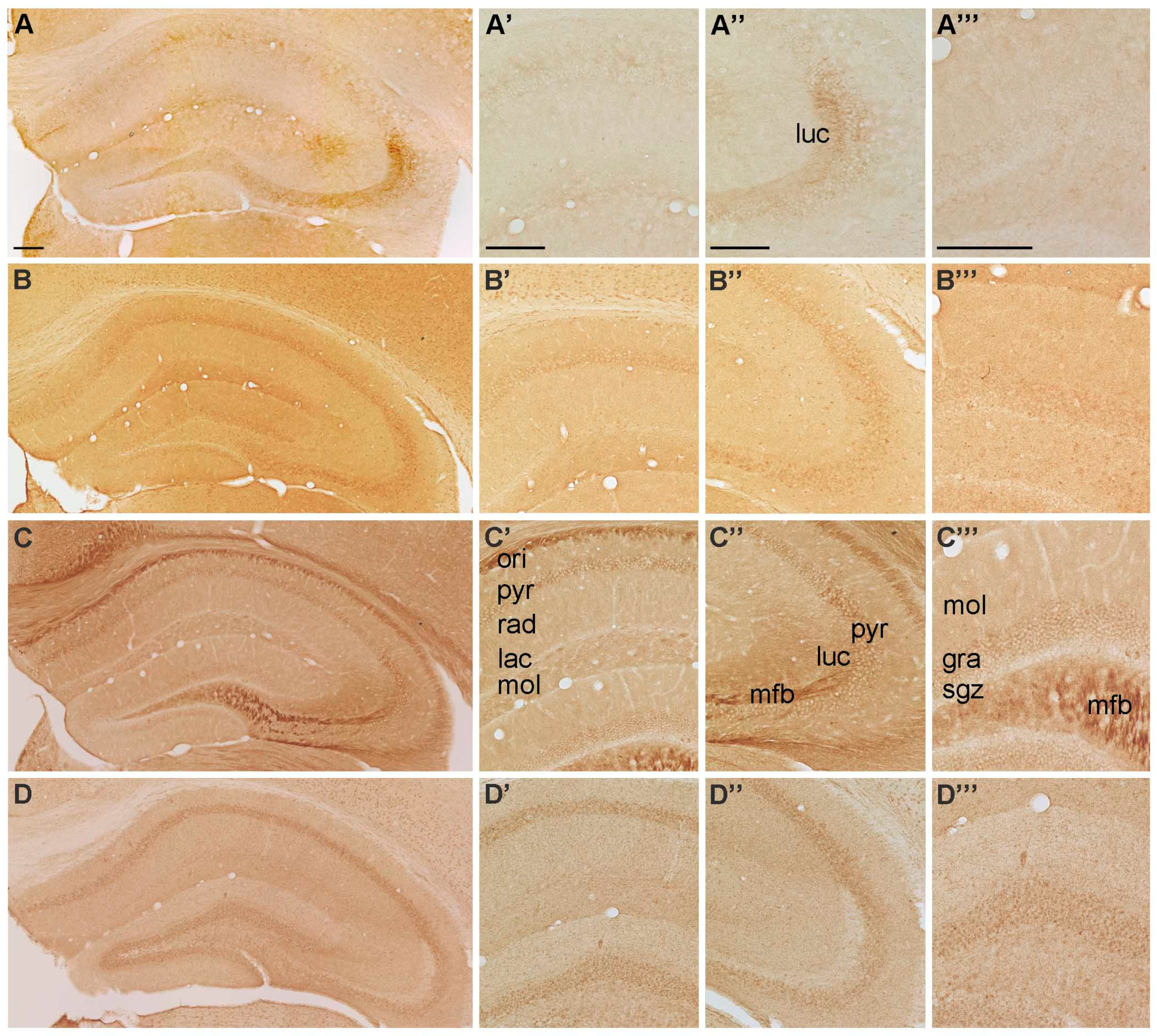
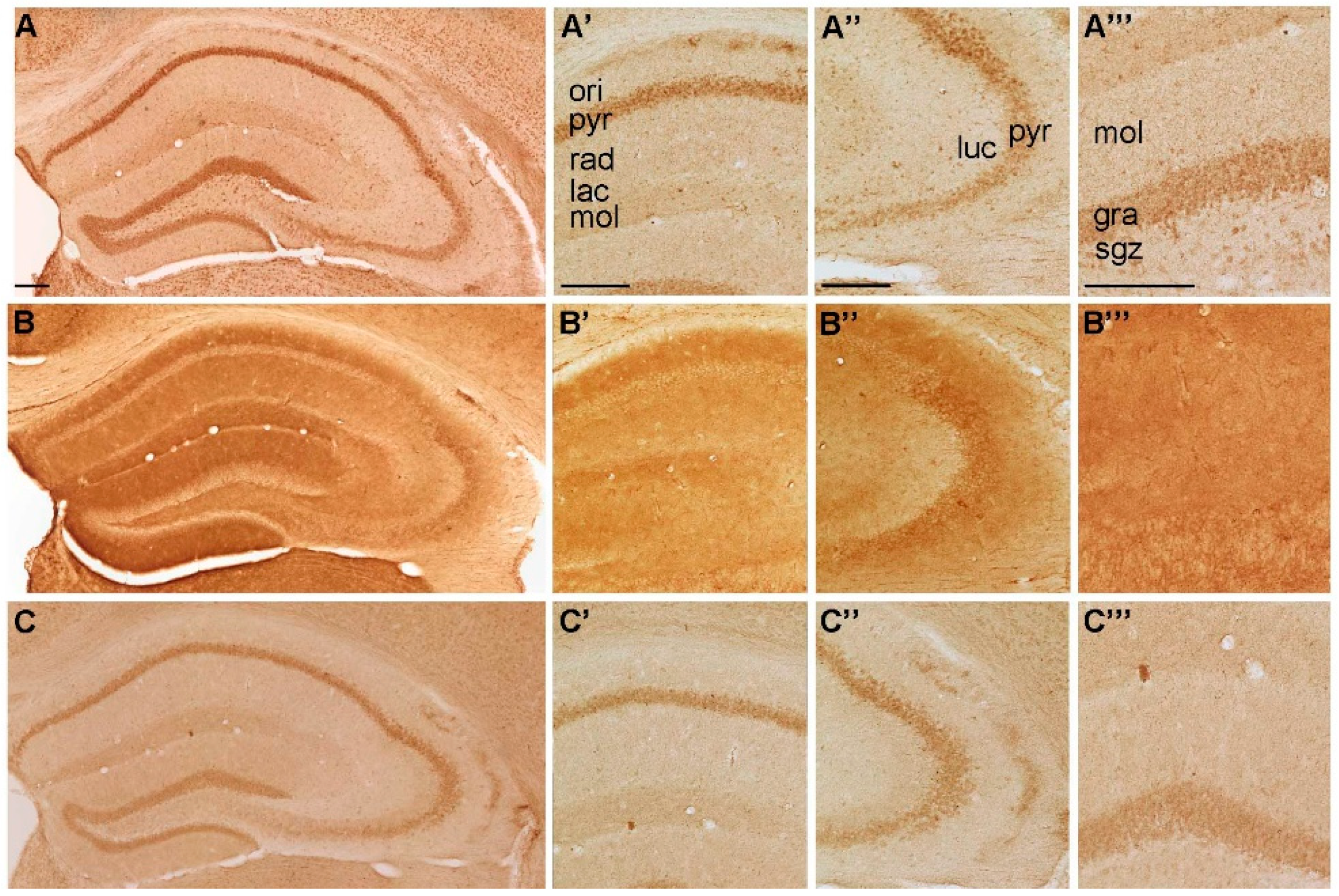
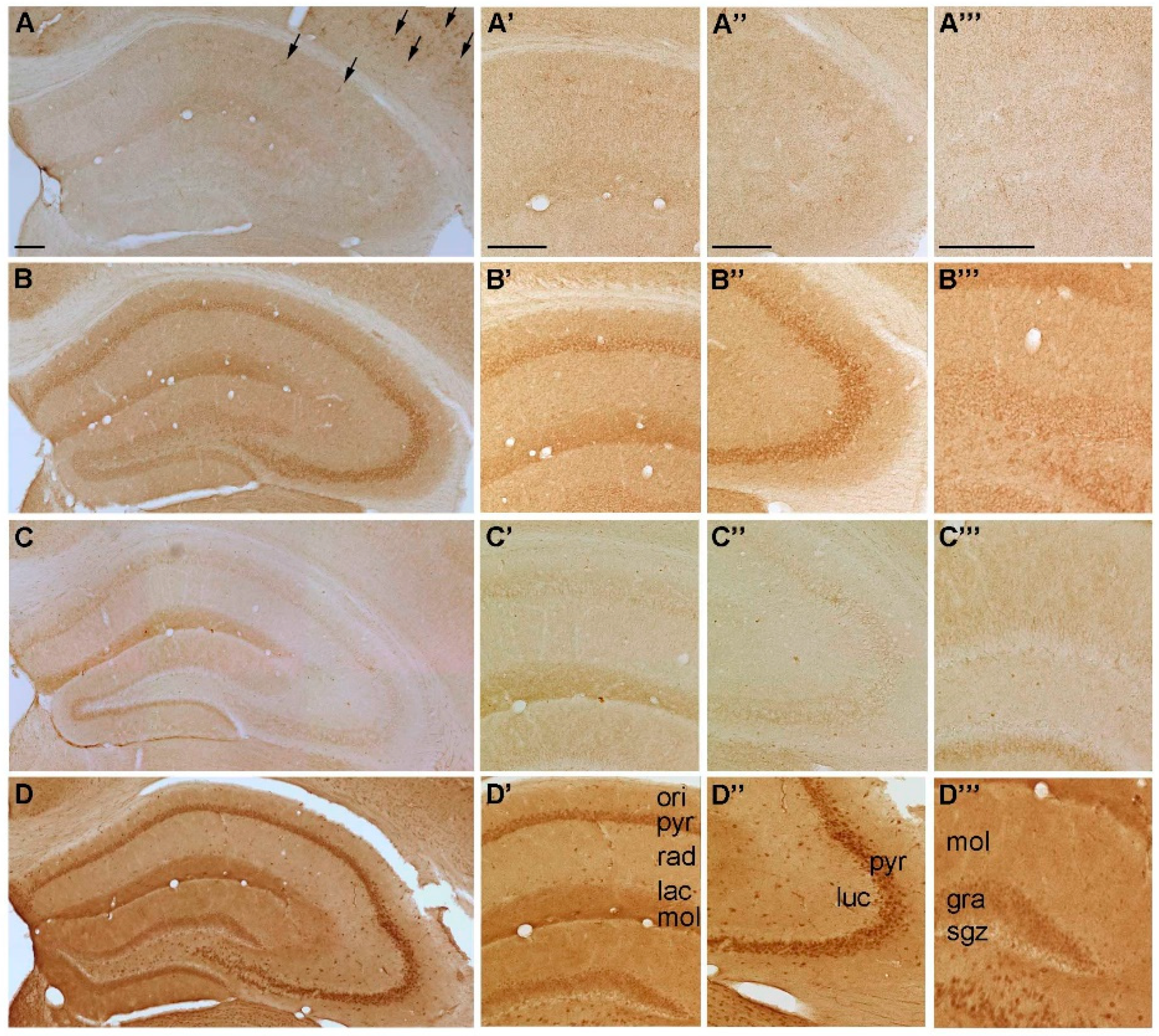
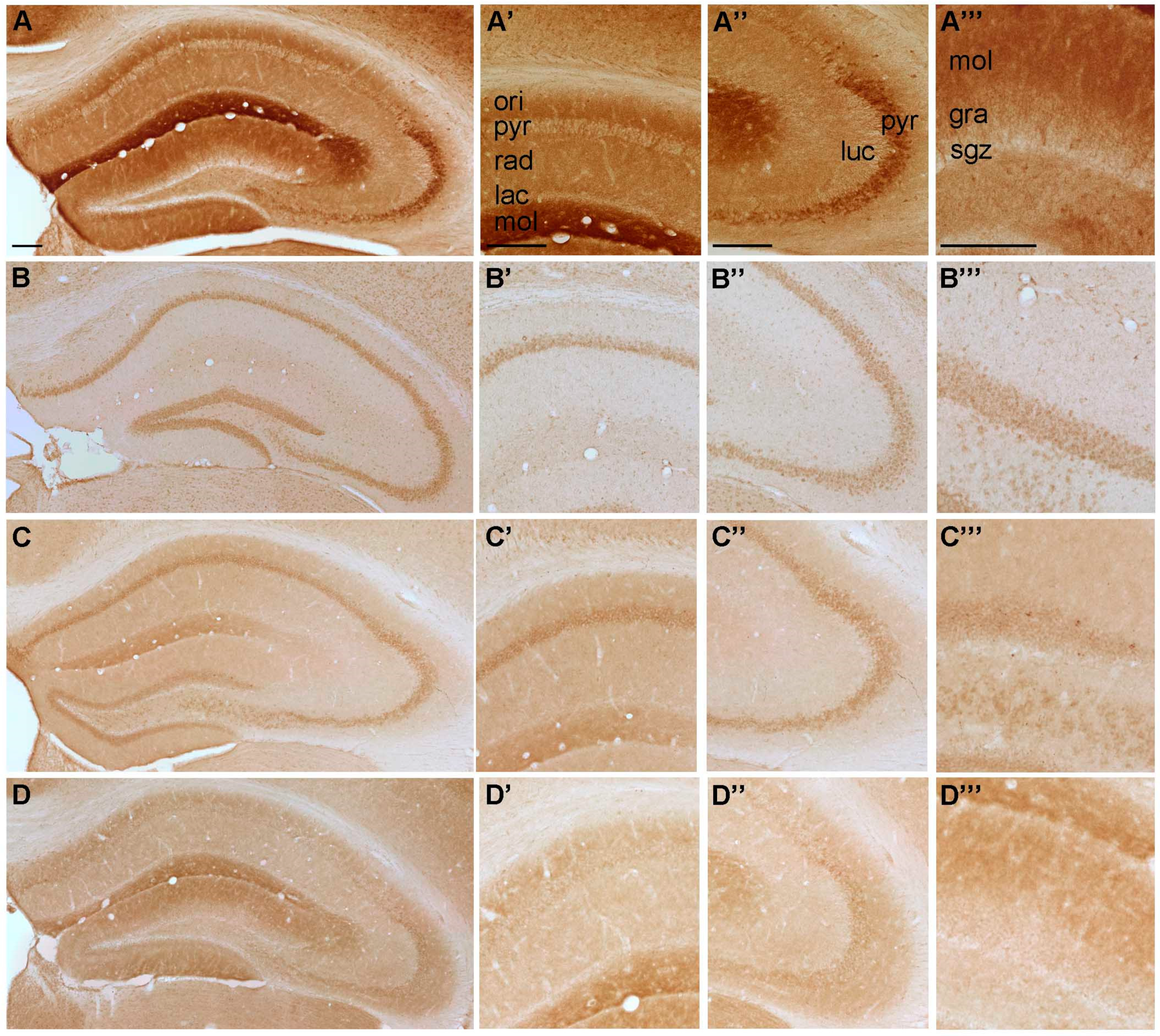


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P2X | P2Y | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 1 | 2 | 4 | 6 | 11 | 12 | 13 | 14 | |
| CA1 | |||||||||||||||
| ori | + | ++ | ++ | + | + | +++ | + | + | ++ | + | ++ | ++ | + | + | + |
| pyr | + | ++ | + | + | +++ | + | +++ | + | ++ | + | +++ | + | ++ | ++ | + |
| rad | + | ++ | + | 0 | + | ++ | + | + | + | + | ++ | ++ | + | + | + |
| lac | + | + | 0 | 0 | + | + | + | + | + | + | + | + | + | + | 0 |
| mol | ++ | ++ | + | + | ++ | +++ | ++ | + | ++ | ++ | +++ | ++++ | + | ++ | +++ |
| CA3 | |||||||||||||||
| ori | + | + | ++ | + | + | ++ | + | + | ++ | + | ++ | + | + | + | ++ |
| pyr | + | ++ | + | + | ++ | +++ | ++ | + | +++ | + | ++++ | +++ | ++ | ++ | + |
| luc | +++ | 0 | ++ | 0 | 0 | + | + | 0 | ++ | + | + | + | + | + | + |
| mol | +++ | + | ++ | 0 | 0 | ++ | 0 | 0 | ++ | 0 | ++ | ++ | + | + | ++ |
| DG | |||||||||||||||
| mol | + | ++ | + | + | + | +++ | + | + | + | + | ++ | +++ | + | + | ++ |
| gra | 0 | + | + | + | +++ | + | +++ | + | ++ | ++ | +++ | + | ++ | ++ | 0 |
| hil | ++ | ++ | +++ | + | ++ | ++ | + | + | ++ | + | ++ | ++ | + | + | + |
| P2X1 | 2 | 3 | 4 | 5 | 6 | 7 | P2Y1 | 2 | 4 | 6 | 11 | 12 | 13 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CA1 | |||||||||||||||
| ori | x | x | |||||||||||||
| pyr | x | x | x | x | x | x | |||||||||
| rad | x | x | x | ||||||||||||
| mol | x | x | x | x | |||||||||||
| CA3 | |||||||||||||||
| mol | |||||||||||||||
| luc | x | ||||||||||||||
| pyr | x | x | x | x | |||||||||||
| ori | x | x | |||||||||||||
| DG | |||||||||||||||
| mol | x | x | x | ||||||||||||
| gra | x | x | x | x | x | x | x | ||||||||
| hil | x |
| Zeitgeber Time | ||||||
|---|---|---|---|---|---|---|
| 2 | 6 | 10 | 14 | 18 | 22 | |
| P2X1 | + | + | ||||
| P2X2 | ||||||
| P2X3 | + | ++++ | ++++ | |||
| P2X4 | ++ | +++ | ||||
| P2X5 | + | ++ | + | |||
| P2X6 | ||||||
| P2X7 | +++ | ++ | ||||
| P2Y1 | ||||||
| P2Y2 | ++ | +++++ | +++++ | ++++ | ||
| P2Y4 | ||||||
| P2Y6 | ++ | +++ | ||||
| P2Y11 | + | |||||
| P2Y12 | + | + | ++ | |||
| P2Y13 | ++++ | ++ | ||||
| P2Y14 | + | |||||
| Antibody | Company and Number | Dilution | Reference |
|---|---|---|---|
| rabbit anti-P2X1 (H-100) | Santa Cruz, sc-25692 | 1:75 | [61,62,63,64,65,66,67] |
| rabbit anti-P2X2 | GeneTex, GTX10266 | 1:500 | [19] |
| rabbit anti-P2X3 (H-60) | Santa Cruz, sc-25694 | 1:50 | [68,69,70,71,72,73] |
| goat anti-P2X4 (N-15) | Santa Cruz, sc-15187 | 1:50 | [61,62,63,64,65] |
| goat anti-P2X5 (N-16) | Santa Cruz, sc-15191 | 1:25 | [19] |
| rabbit anti-P2X6 | LSBio, LS-C94426 | 1:100 | [19] |
| goat anti-P2X7 (L-20) | Santa Cruz, sc-15200 | 1:50 | [74,75,76,77,78] |
| rabbit anti-P2Y1 | Santa Cruz, sc-20123 | 1:50 | [79,80,81] |
| rabbit anti-P2Y2 | NovusBio, NB110-39032 | 1:100 | [19,82,83] |
| rabbit anti-P2Y4 | GeneTex, GTX87199 | 1:750 | [17] |
| rabbit anti-P2Y6 | Alomone labs, APR-011 | 1:100 | [84,85] |
| rabbit anti-P2Y11 | GeneTex, GTX108241 | 1:300 | [19,86] |
| rabbit anti-P2Y12 (P-14) | Santa Cruz, sc-27152 | 1:50 | [79,87,88] |
| rabbit anti-P2Y13 | LSBio, LS-C145104 | 1:250 | [19] |
| rabbit anti-P2Y14 | LSBio, LS-C120603 | 1:250 | [19] |
| goat anti-rabbit | Vector Laboratories, BA-1000 | 1:500 | [89] |
| rabbit anti-goat | Vector Laboratories, BA-5000 | 1:500 | [90] |
| Color | Intensity |
|---|---|
| yellow | absent |
| magenta | very weak |
| light blue | weak |
| dark blue | moderate |
| green | strong |
| red | very strong |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lommen, J.; Detken, J.; Harr, K.; von Gall, C.; Ali, A.A.H. Analysis of Spatial and Temporal Distribution of Purinergic P2 Receptors in the Mouse Hippocampus. Int. J. Mol. Sci. 2021, 22, 8078. https://doi.org/10.3390/ijms22158078
Lommen J, Detken J, Harr K, von Gall C, Ali AAH. Analysis of Spatial and Temporal Distribution of Purinergic P2 Receptors in the Mouse Hippocampus. International Journal of Molecular Sciences. 2021; 22(15):8078. https://doi.org/10.3390/ijms22158078
Chicago/Turabian StyleLommen, Julian, Julika Detken, Katharina Harr, Charlotte von Gall, and Amira A. H. Ali. 2021. "Analysis of Spatial and Temporal Distribution of Purinergic P2 Receptors in the Mouse Hippocampus" International Journal of Molecular Sciences 22, no. 15: 8078. https://doi.org/10.3390/ijms22158078
APA StyleLommen, J., Detken, J., Harr, K., von Gall, C., & Ali, A. A. H. (2021). Analysis of Spatial and Temporal Distribution of Purinergic P2 Receptors in the Mouse Hippocampus. International Journal of Molecular Sciences, 22(15), 8078. https://doi.org/10.3390/ijms22158078