
Role of Klotho in Hyperglycemia: Its Levels and Effects on Fibroblast Growth Factor Receptors, Glycolysis, and Glomerular Filtration

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
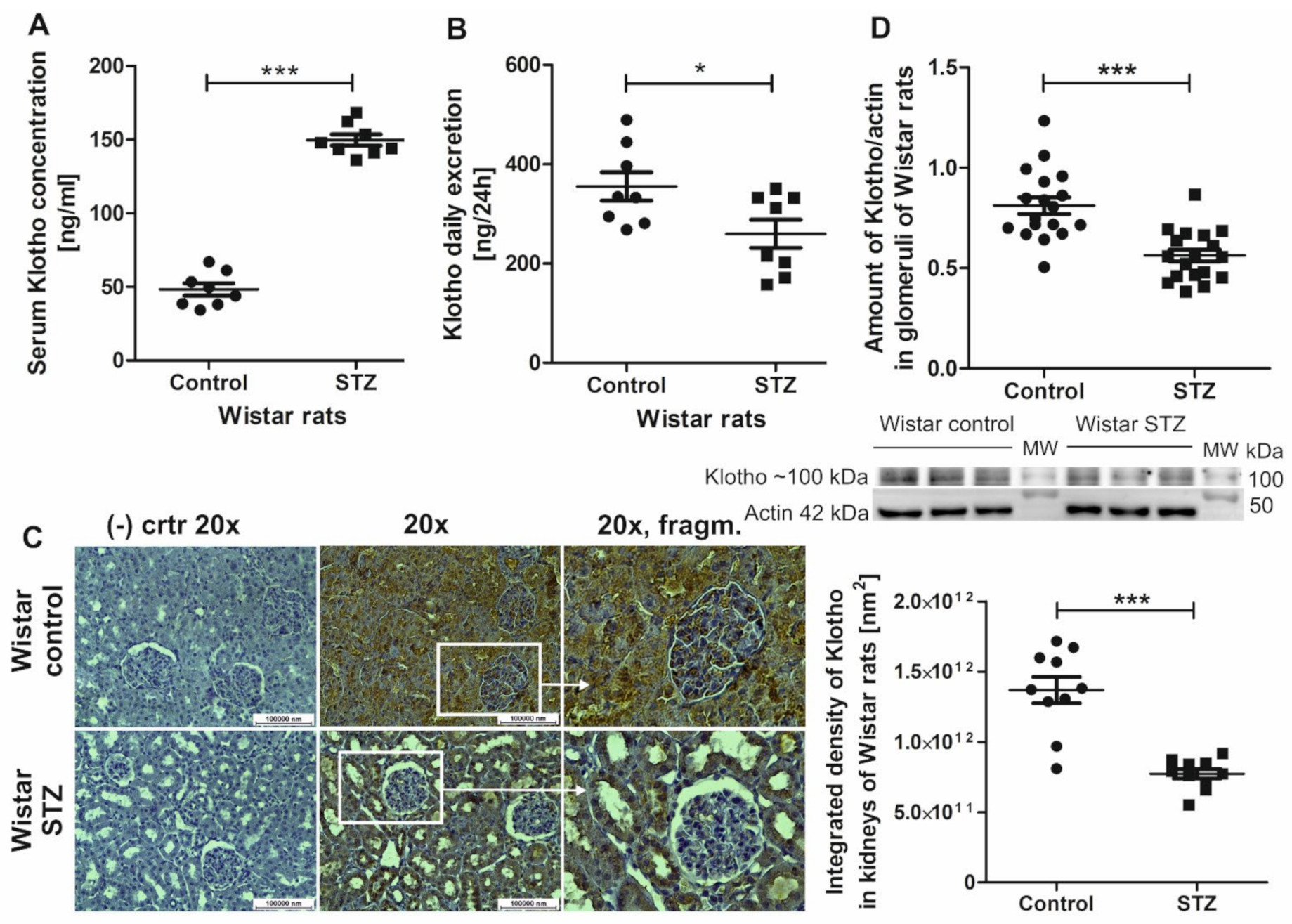
2.1. Klotho Levels Increase in Serum and Decrease in Renal Tissue and Urine in Diabetic Rats
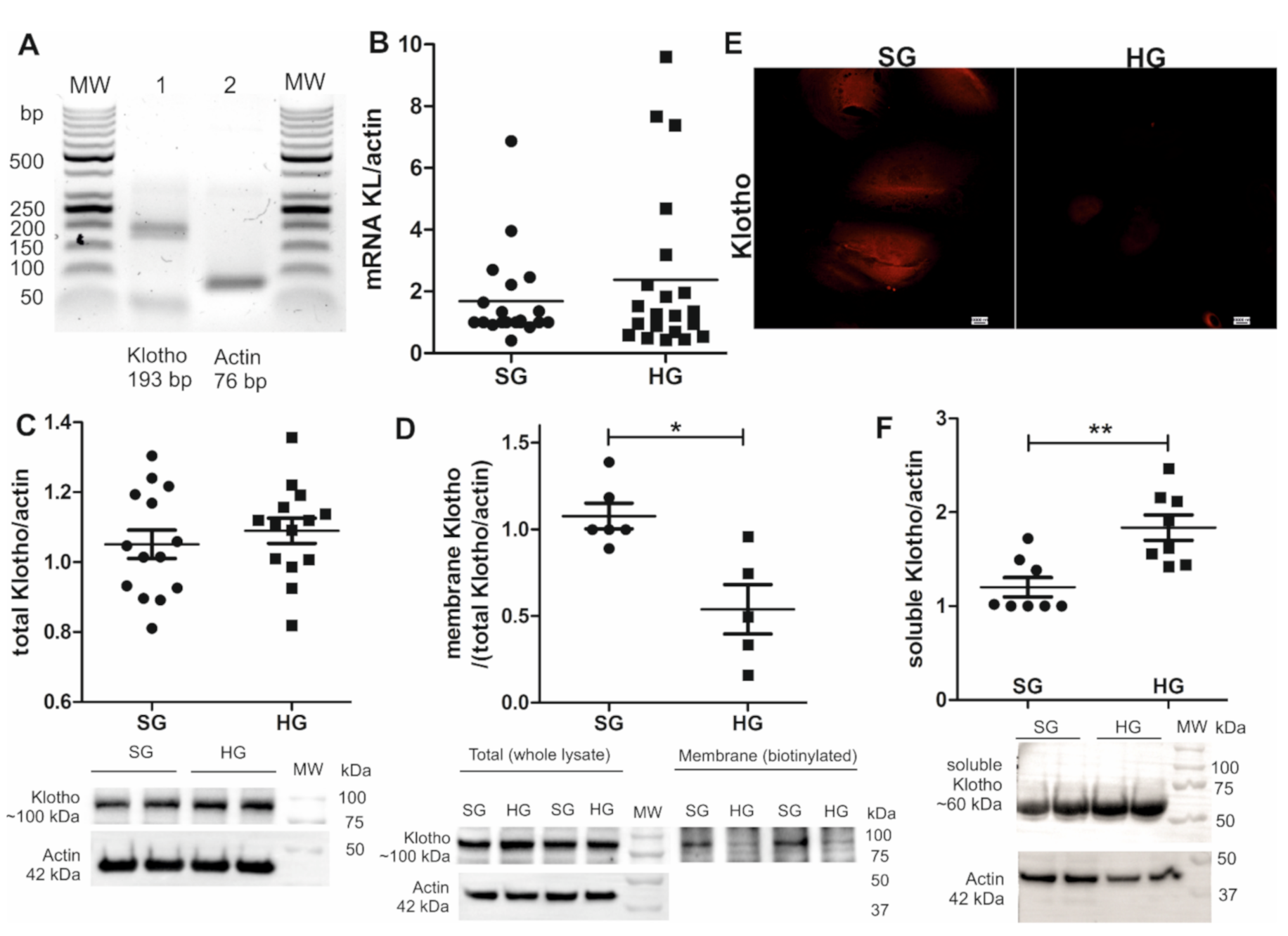
2.2. Shedding of Soluble Klotho Increases in Hyperglycemia and Causes a Decrease in the Amount of Podocyte Membrane-Bound Klotho
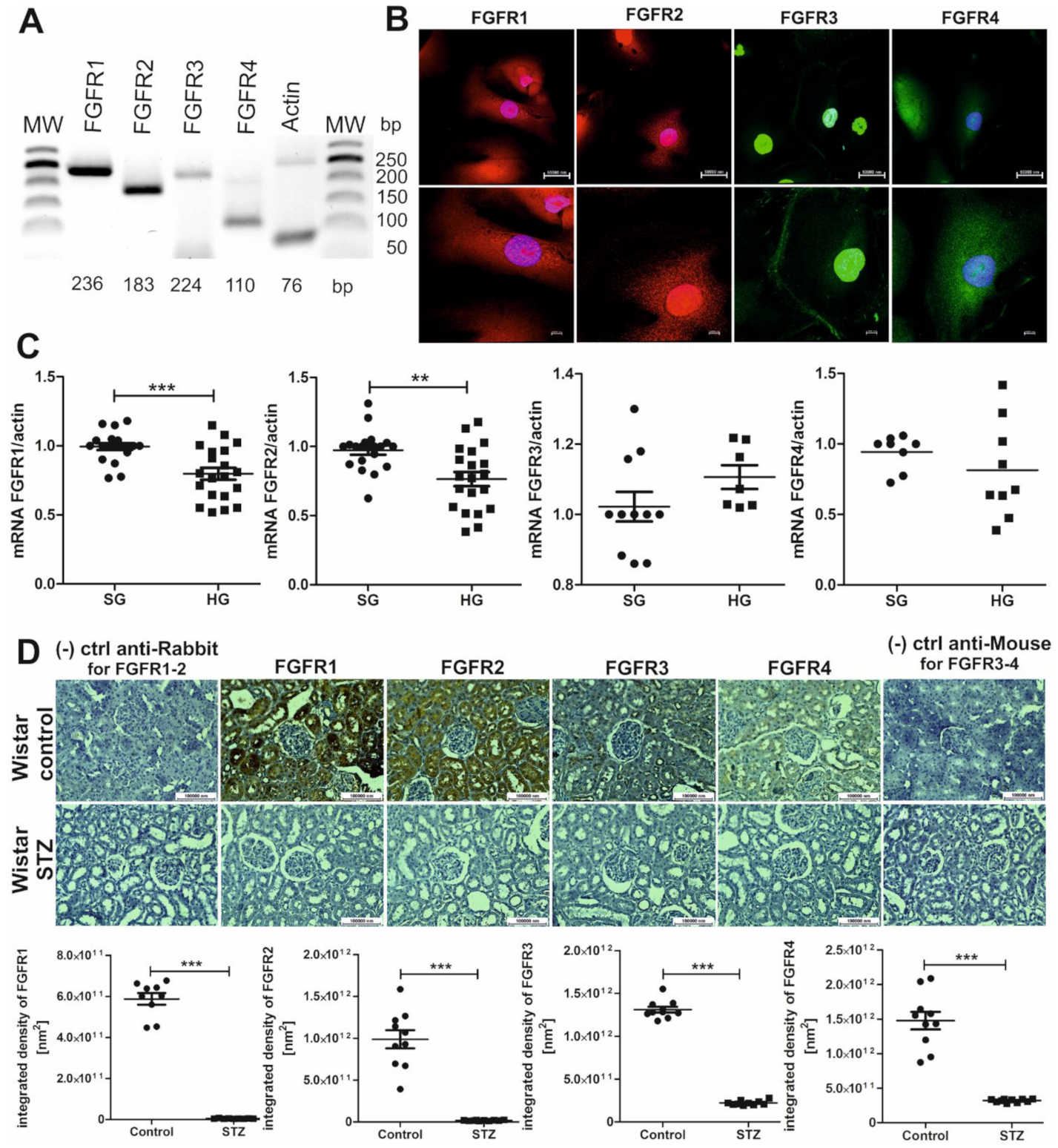
2.3. FGFRs Are Present on Podocytes and Cells That Form Tubules of the Nephron, and Their Expression Decreases in Hyperglycemia
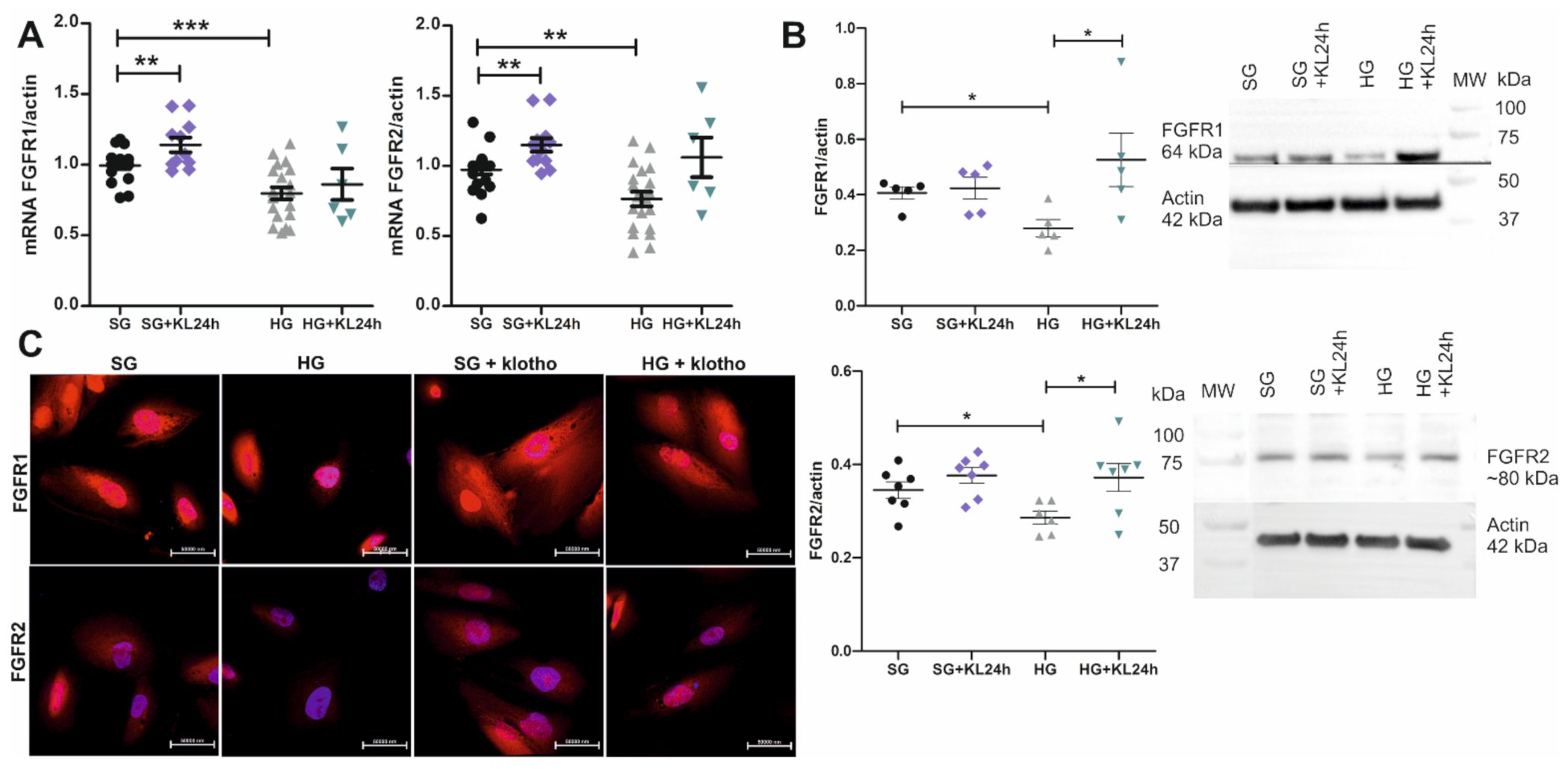
2.4. The Addition of Klotho Increases FGFR1 and FGFR2 Expression
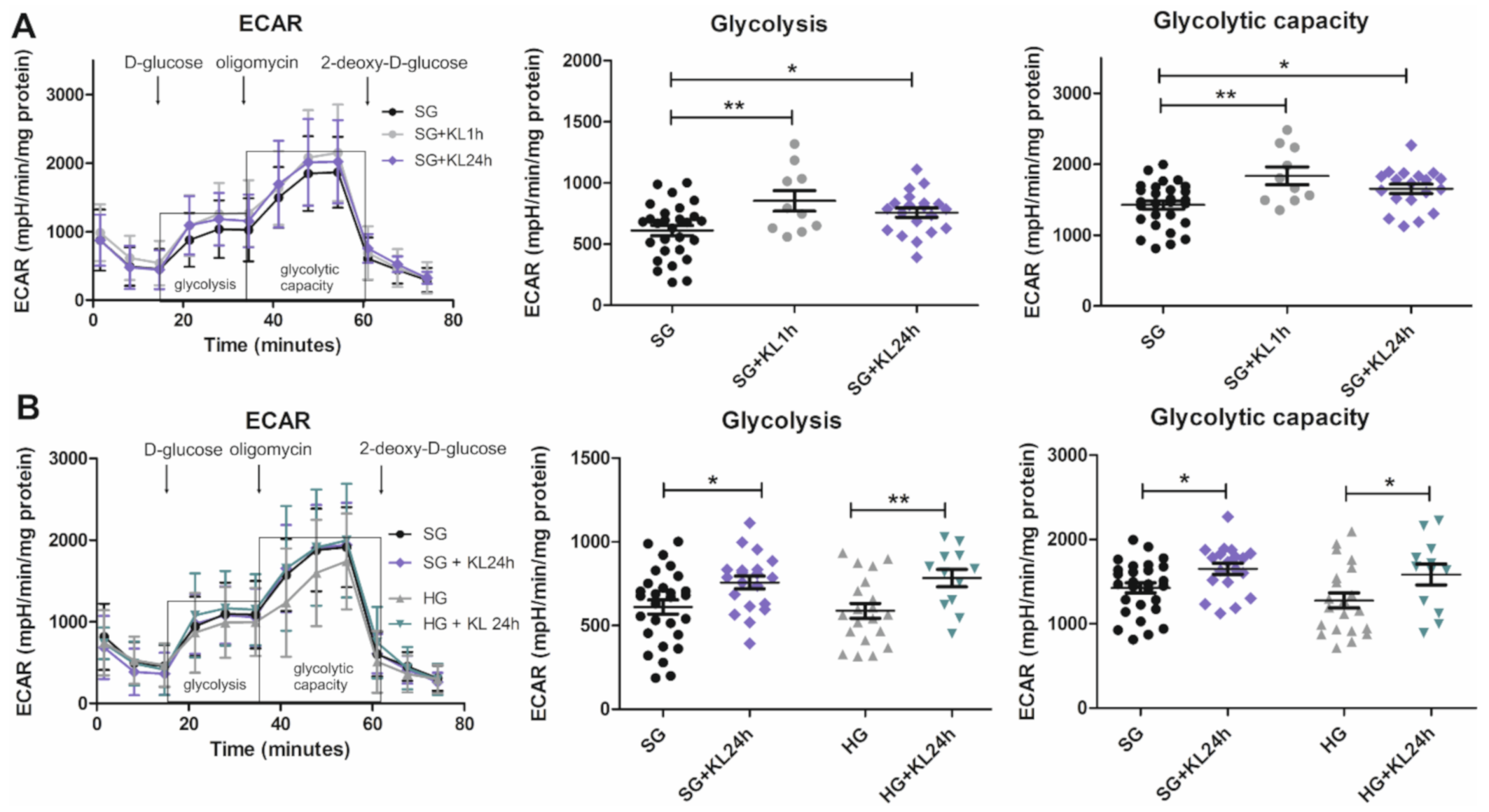
2.5. Klotho Increases Glycolysis and Glycolytic Capacity Levels in Podocytes That Are Grown under Both SG and HG Conditions
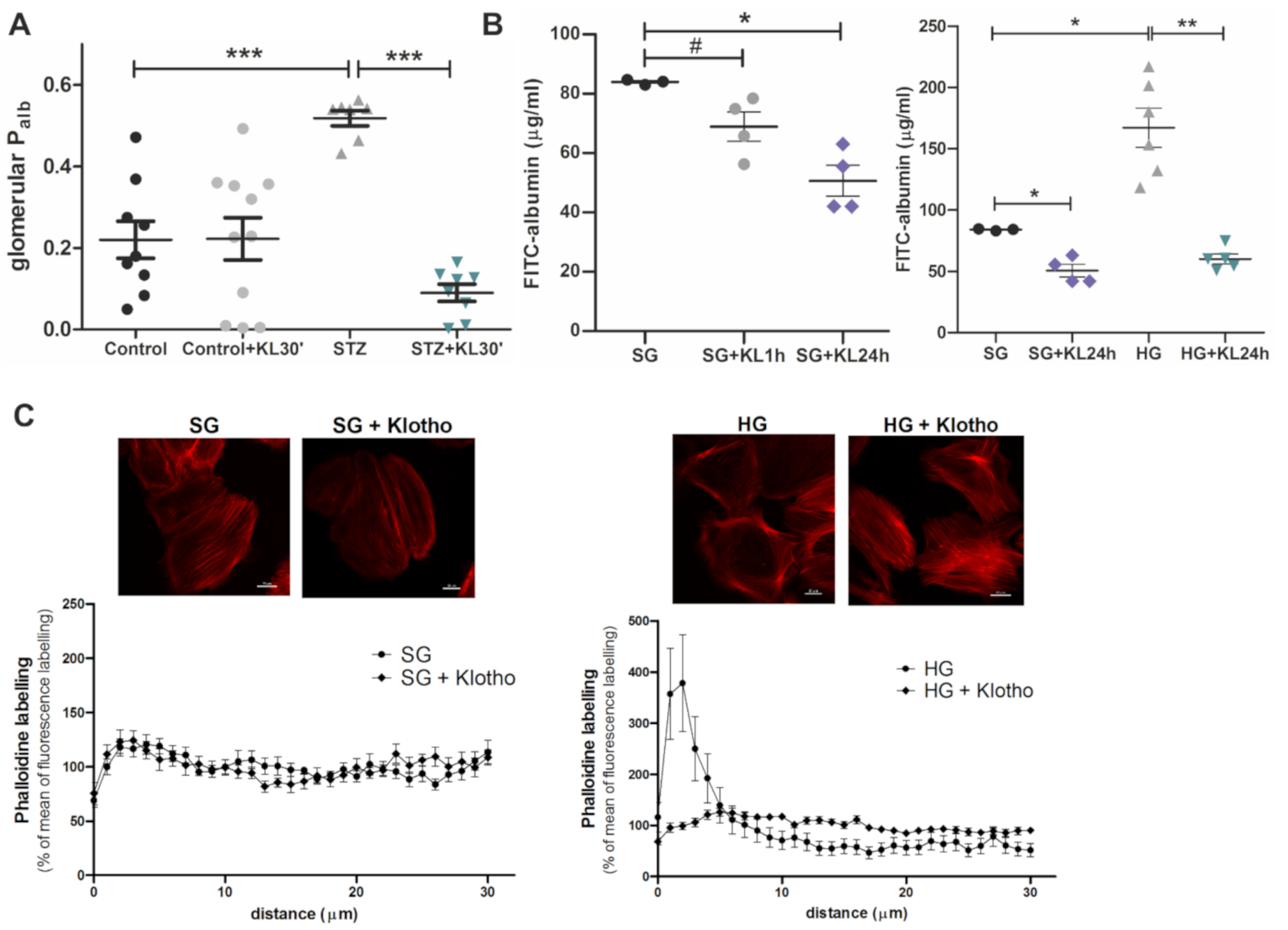
2.6. Klotho Improves Function of the GFB
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Experimental Animals and Metabolic Cage Studies
4.3. Isolation of Rat Glomeruli
4.4. Human Podocyte Cell Culture
4.5. Biotinylation
4.6. Western Blot
4.7. Immunohistochemistry
4.8. ELISA
4.9. Real-Time Polymerase Chain Reaction
4.10. Immunofluorescent Staining
4.11. Glycolysis Stress Test
4.12. Glomerular Permeability to Albumin In Vitro
4.13. Albumin Permeability of Human Podocytes
4.14. F-Actin Network
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pavenstädt, H.; Kriz, W.; Kretzler, M. Cell biology of the glomerular podocyte. Physiol. Rev. 2003, 83, 253–307. [Google Scholar] [CrossRef]
- Sarwar, N.; Gao, P.; Seshasai, S.R.; Gobin, R.; Kaptoge, S.; Di Angelantonio, E.; Ingelsson, E.; Lawlor, D.A.; Selvin, E.; Stampfer, M.; et al. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: A collaborative meta-analysis of 102 prospective studies. Lancet 2010, 375, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- Henning, R.J. Type-2 diabetes mellitus and cardiovascular disease. Future Cardiol. 2018, 14, 491–509. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.Y.; Mei Wong, J.L.; Sim, Y.J.; Wong, S.S.; Mohamed Elhassan, S.A.; Tan, S.H.; Ling Lim, G.P.; Rong Tay, N.W.; Annan, N.C.; Bhattamisra, S.K.; et al. Type 1 and 2 diabetes mellitus: A review on current treatment approach and gene therapy as potential intervention. Diabetes Metab. Syndr. 2019, 13, 364–372. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Status Report on Noncommunicable Diseases; World Health Organisation: Geneva, Switzerland, 2014. [Google Scholar]
- Susztak, K.; Raff, A.C.; Schiffer, M.; Böttinger, E.P. Glucose-induced reactive oxygen species cause apoptosis of podocytes and podocyte depletion at the onset of diabetic nephropathy. Diabetes 2006, 55, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, S.; Ueda, S.; Imamura, H.; Mori, K.; Asanuma, K.; Yanagita, M.; Nakagawa, T. Glycolysis, but not mitochondria, responsible for intracellular atp distribution in cortical area of podocytes. Sci. Rep. 2015, 5, 18575. [Google Scholar] [CrossRef] [PubMed]
- Imasawa, T.; Obre, E.; Bellance, N.; Lavie, J.; Rigothier, C.; Delmas, Y.; Combe, C.; Lacombe, D.; Benard, G.; Claverol, S.; et al. High glucose repatterns human podocyte energy metabolism during differentiation and diabetic nephropathy. FASEB J. 2017, 31, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Kwak, S.J.; Jung, D.S.; Kim, J.J.; Yoo, T.H.; Ryu, D.R.; Han, S.H.; Choi, H.Y.; Lee, J.E.; Moon, S.J.; et al. Podocyte biology in diabetic nephropathy. Kidney Int. 2007, 72, S36–S42. [Google Scholar] [CrossRef] [PubMed]
- Lerco, M.M.; Macedo, C.S.; Silva, R.J.; Pinheiro Dde, O.; Spadella, C.T. The number of podocyte and slit diaphragm is decreased in experimental diabetic nephropathy. Acta Cir. Bras. 2006, 21, 87–91. [Google Scholar] [CrossRef][Green Version]
- Kasztan, M.; Piwkowska, A.; Kreft, E.; Rogacka, D.; Audzeyenka, I.; Szczepanska-Konkel, M.; Jankowski, M. Extracellular purines’ action on glomerular albumin permeability in isolated rat glomeruli: Insights into the pathogenesis of albuminuria. Am. J. Physiol. Renal. Physiol. 2016, 311, F103–F111. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhou, L. The Signaling of Cellular Senescence in Diabetic Nephropathy. Oxidative Med. Cell. Longev. 2019, 2019, 7495629-16. [Google Scholar] [CrossRef] [PubMed]
- Buendía, P.; Ramírez, R.; Aljama, P.; Carracedo, J. Klotho Prevents Translocation of NFκB. Vitam. Horm. 2016, 101, 119–150. [Google Scholar] [CrossRef] [PubMed]
- Mencke, R.; Harms, G.; Moser, J.; Van Meurs, M.; Diepstra, A.; Leuvenink, H.; Hillebrands, J.-L. Human alternative Klotho mRNA is a nonsense-mediated mRNA decay target inefficiently spliced in renal disease. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Xie, J.; Hwang, K.-H.; Wu, Y.-L.; Oliver, N.; Eom, M.; Park, K.-S.; Barrezueta, N.; Kong, I.-D.; Fracasso, R.P.; et al. Klotho May Ameliorate Proteinuria by Targeting TRPC6 Channels in Podocytes. J. Am. Soc. Nephrol. 2016, 28, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.C.; Shi, M.; Zhang, J.; Pastor, J.; Nakatani, T.; Lanske, B.; Razzaque, M.S.; Rosenblatt, K.P.; Baum, M.G.; Kuro-O, M.; et al. Klotho: A novel phosphaturic substance acting as an autocrine enzyme in the renal proximal tubule. FASEB J. 2010, 24, 3438–3450. [Google Scholar] [CrossRef]
- Richter, B.; Faul, C. FGF23 Actions on Target Tissues—With and Without Klotho. Front. Endocrinol. 2018, 9, 189. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Song, S.H.; Kim, I.J.; Lee, E.Y.; Lee, S.M.; Chung, C.; Kwak, I.S.; Kim, Y.K. Decreased plasma α-klotho predict progression of nephropathy with type 2 diabetic patients. J. Diabetes Complicat. 2016, 30, 887–892. [Google Scholar] [CrossRef]
- Typiak, M.; Piwkowska, A. Antiinflammatory Actions of Klotho: Implications for Therapy of Diabetic Nephropathy. Int. J. Mol. Sci. 2021, 22, 956. [Google Scholar] [CrossRef]
- Guo, Y.; Zhuang, X.-D.; Zou, J.; Yang, D.; Hu, X.; Du, Z.; Wang, L.; Liao, X. Klotho protects the heart from hyperglycemia-induced injury by inactivating ROS and NF-κB-mediated inflammation both in vitro and in vivo. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 238–251. [Google Scholar] [CrossRef]
- Oh, H.J.; Nam, B.Y.; Wu, M.; Kim, S.; Park, J.; Kang, S.; Park, J.T.; Yoo, T.-H.; Kang, S.-W.; Han, S.H. Klotho plays a protective role against glomerular hypertrophy in a cell cycle-dependent manner in diabetic nephropathy. Am. J. Physiol. Physiol. 2018, 315, F791–F805. [Google Scholar] [CrossRef]
- Jiang, W.; Xiao, T.; Han, W.; Xiong, J.; He, T.; Liu, Y.; Huang, Y.; Yang, K.; Bi, X.; Xu, X.; et al. Klotho inhibits PKCα/p66SHC-mediated podocyte injury in diabetic nephropathy. Mol. Cell. Endocrinol. 2019, 494, 110490. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Su, N.; Yang, J.; Tan, Q.; Huang, S.; Jin, M.; Ni, Z.; Zhang, B.; Zhang, D.; Luo, F.; et al. FGF/FGFR signaling in health and disease. Signal Transduct. Target. Ther. 2020, 5, 1–38. [Google Scholar] [CrossRef]
- Cancilla, B.; Davies, A.; Cauchi, J.A.; Risbridger, G.; Bertram, J. Fibroblast growth factor receptors and their ligands in the adult rat kidney. Kidney Int. 2001, 60, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Vierthaler, L.; Tang, W.; Zhou, J.; Quarles, L.D. FGFR3 and FGFR4 Do not Mediate Renal Effects of FGF23. J. Am. Soc. Nephrol. 2008, 19, 2342–2350. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef]
- Davidson, G.; Dono, R.; Zeller, R. FGF signalling is required for differentiation-induced cytoskeletal reorganisation and formation of actin-based processes by podocytes. J. Cell Sci. 2001, 114, 3359–3366. [Google Scholar] [CrossRef]
- Reiser, J.; Sever, S.; Faul, C. Signal transduction in podocytes—Spotlight on receptor tyrosine kinases. Nat. Rev. Nephrol. 2014, 10, 104–115. [Google Scholar] [CrossRef]
- Hu, M.C.; Kuro-O, M.; Moe, O.W. Secreted Klotho and Chronic Kidney Disease. In Endocrine FGFs and Klothos; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2012; Volume 728, pp. 126–157. [Google Scholar] [CrossRef]
- Kacso, I.M.; Bondor, C.I.; Kacso, G. Soluble serum Klotho in diabetic nephropathy: Relationship to VEGF-A. Clin. Biochem. 2012, 45, 1415–1420. [Google Scholar] [CrossRef]
- Bob, F.; Schiller, A.; Timar, R.; Lighezan, D.; Schiller, O.; Timar, B.; Bujor, C.G.; Munteanu, M.; Gadalean, F.; Mihaescu, A.; et al. Rapid decline of kidney function in diabetic kidney disease is associated with high soluble Klotho levels. Nefrologia 2019, 39, 250–257. [Google Scholar] [CrossRef]
- Van Ark, J.; Hammes, H.-P.; Van Dijk, M.C.R.F.; Vervloet, M.G.; Wolffenbuttel, B.H.; Van Goor, H.; Hillebrands, J.-L. Circulating alpha-klotho levels are not disturbed in patients with type 2 diabetes with and without macrovascular disease in the absence of nephropathy. Cardiovasc. Diabetol. 2013, 12, 116. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.-J.; Han, D.-J.; Lee, J.-H.; Jang, S.-H.; Kang, J.S.; Gil, H.-W.; Park, S.; Lee, E.Y. Soluble klotho as a marker of renal fibrosis and podocyte injuries in human kidneys. PLoS ONE 2018, 13, e0194617. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.H.; Lam, J.K.Y.; Shiu, S.W.M.; Wong, Y.; Betteridge, D.J.; Tan, K.C.B. Serum Level of Soluble Receptor for Advanced Glycation End Products Is Associated with A Disintegrin And Metalloproteinase 10 in Type 1 Diabetes. PLoS ONE 2015, 10, e0137330. [Google Scholar] [CrossRef]
- Palau, V.; Pascual, J.; Soler, M.J.; Riera, M. Role of ADAM17 in kidney disease. Am. J. Physiol. Physiol. 2019, 317, F333–F342. [Google Scholar] [CrossRef]
- Asai, O.; Nakatani, K.; Tanaka, T.; Sakan, H.; Imura, A.; Yoshimoto, S.; Samejima, K.-I.; Yamaguchi, Y.; Matsui, M.; Akai, Y.; et al. Decreased renal α-Klotho expression in early diabetic nephropathy in humans and mice and its possible role in urinary calcium excretion. Kidney Int. 2012, 81, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.; Yanucil, C.; Musgrove, J.; Shi, M.; Ide, S.; Souma, T.; Faul, C.; Wolf, M.; Grabner, A. FGFR4 does not contribute to progression of chronic kidney disease. Sci. Rep. 2019, 9, 14023. [Google Scholar] [CrossRef]
- Cheng, M.-F.; Chen, L.-J.; Wang, M.-C.; Hsu, C.-T.; Cheng, J.-T. Decrease of FGF Receptor (FGFR) and Interstitial Fibrosis in the Kidney of Streptozotocin-induced Diabetic Rats. Horm. Metab. Res. 2013, 46, 1–7. [Google Scholar] [CrossRef]
- Dixit, G.; Schanz, W.; Pappas, B.; Maretzky, T. Members of the Fibroblast Growth Factor Receptor Superfamily Are Proteolytically Cleaved by Two Differently Activated Metalloproteases. Int. J. Mol. Sci. 2021, 22, 3165. [Google Scholar] [CrossRef]
- Wu, A.-L.; Kolumam, G.; Stawicki, S.; Chen, Y.; Li, J.; Zavala-Solorio, J.; Phamluong, K.; Feng, B.; Li, L.; Marsters, S.; et al. Amelioration of Type 2 Diabetes by Antibody-Mediated Activation of Fibroblast Growth Factor Receptor 1. Sci. Transl. Med. 2011, 3, 113ra126. [Google Scholar] [CrossRef]
- Xu, Z.; Zhu, X.; Wang, M.; Lu, Y.; Dai, C. FGF/FGFR2 Protects against Tubular Cell Death and Acute Kidney Injury Involving Erk1/2 Signaling Activation. Kidney Dis. 2020, 6, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Mazucanti, C.H.; Kawamoto, E.; Mattson, M.P.; Scavone, C.; Camandola, S. Activity-dependent neuronal Klotho enhances astrocytic aerobic glycolysis. Br. J. Pharmacol. 2018, 39, 1544–1556. [Google Scholar] [CrossRef] [PubMed]
- Brinkkoetter, P.; Bork, T.; Salou, S.; Liang, W.; Mizi, A.; Özel, C.; Koehler, S.; Hagmann, H.H.; Ising, C.; Kuczkowski, A.; et al. Anaerobic Glycolysis Maintains the Glomerular Filtration Barrier Independent of Mitochondrial Metabolism and Dynamics. Cell Rep. 2019, 27, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Agilent, Technologies and Inc. Seahorse XF Glycolysis Stress Test Kit User Guide; Agilent, Technologies and Inc.: Wilmington, NC, USA, 2019; pp. 5–8. [Google Scholar]
- Bender, D.A.; Mayes, P.A. Glycolysis & the Oxidation of Pyruvate. In Harper’s Illustrated Biochemistry 29; Bender, D.A., Botham, K.M., Weil, P.A., Kennelly, P.J., Murray, R.K., Rodwell, V.W., Eds.; McGraw-Hill: New York, NY, USA, 2012. [Google Scholar]
- Guo, X.; Li, H.; Xu, H.; Woo, S.; Dong, H.; Lu, F.; Lange, A.J.; Wu, C. Glycolysis in the control of blood glucose homeostasis. Acta Pharm. Sin. B 2012, 2, 358–367. [Google Scholar] [CrossRef]
- Piwkowska, A.; Rogacka, D.; Audzeyenka, I.; Angielski, S.; Jankowski, M. Combined effect of insulin and high glucose concentration on albumin permeability in cultured rat podocytes. Biochem. Biophys. Res. Commun. 2015, 461, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Audzeyenka, I.; Rachubik, P.; Rogacka, D.; Typiak, M.; Kulesza, T.; Angielski, S.; Rychłowski, M.; Wysocka, M.; Gruba, N.; Lesner, A.; et al. Cathepsin C is a novel mediator of podocyte and renal injury induced by hyperglycemia. Biochim. Biophys. Acta Bioenerg. 2020, 1867, 118723. [Google Scholar] [CrossRef] [PubMed]
- Szrejder, M.; Rachubik, P.; Rogacka, D.; Audzeyenka, I.; Rychłowski, M.; Kreft, E.; Angielski, S.; Piwkowska, A. Metformin reduces TRPC6 expression through AMPK activation and modulates cytoskeleton dynamics in podocytes under diabetic conditions. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165610. [Google Scholar] [CrossRef]
- Olejnik, A.; Franczak, A.; Krzywonos-Zawadzka, A.; Kałużna-Oleksy, M.; Bil-Lula, I. The Biological Role of Klotho Protein in the Development of Cardiovascular Diseases. BioMed Res. Int. 2018, 2018, 1–17. [Google Scholar] [CrossRef]
- Saleem, M.A.; O’Hare, M.J.; Reiser, J.; Coward, R.J.; Inward, C.D.; Farren, T.; Xing, C.Y.; Ni, L.; Mathieson, P.W.; Mundel, P. A conditionally immortalized human podocyte cell line demonstrating nephrin and podocin expression. J. Am. Soc. Nephrol. 2002, 13, 630–638. [Google Scholar] [CrossRef]
- Piwkowska, A.; Rogacka, D.; Jankowski, M.; Kocbuch, K.; Angielski, S. Hydrogen peroxide induces dimerization of protein kinase G type Iα subunits and increases albumin permeability in cultured rat podocytes. J. Cell. Physiol. 2011, 227, 1004–1016. [Google Scholar] [CrossRef]
- Divakaruni, A.S.; Paradyse, A.; Ferrick, D.A.; Murphy, A.N.; Jastroch, M. Analysis and Interpretation of Microplate-Based Oxygen Consumption and pH Data. Methods Enzymol. 2014, 547, 309–354. [Google Scholar] [CrossRef] [PubMed]
- Savin, V.J.; Sharma, R.; Lovell, H.B.; Welling, D.J. Measurement of albumin reflection coefficient with isolated rat glomeruli. J. Am. Soc. Nephrol. 1992, 3, 1260–1269. [Google Scholar] [CrossRef] [PubMed]
- Oshimaa, T.; Larouxa, F.S.; Coe, L.L.; Morise, Z.; Kawachia, S.; Bauera, P.; Grisham, M.B.; Specian, R.D.; Cartera, P.; Jenningsb, S.; et al. Interferon-γ and Interleukin-10 Reciprocally Regulate Endothelial Junction Integrity and Barrier Function. Microvasc. Res. 2001, 61, 130–143. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control n = 4 | STZ n = 4 |
|---|---|---|
| Body weight (g) | 286 ± 23.98 | 210.25 ± 19.80 |
| Blood glucose (mg/dl) | 112.75 ± 4.87 | 422 ± 65.40 * |
| Urine volume (ml/24 h) | 9.5 ± 1.51 | 166.5 ± 10.87 * |
| Urinary albumin excretion (mg/24 h) | 0.15 ± 0.03 | 2.57 ± 0.67 * |
| Antibody | Clonality | Dilution | Source |
|---|---|---|---|
| Klotho | monoclonal | 1:50 (IF); 1:400 (IHC); 1:445 (WB) | Sigma Aldrich, SAB3500604 |
| Soluble Klotho | monoclonal | 1:500 (WB) | Cusabio Technology, CSB-PA552336 |
| FGFR1 | polyclonal | 1:50 (IF); 1:30 (IHC); 1:2000 (WB) | Cusabio Technology, CSB-PA008642LA01HU |
| FGFR2 | polyclonal | 1:25 (IF); 1:25 (IHC); 1:500 (WB) | Cusabio Technology, CSB-PA000992 |
| FGFR3 | monoclonal | 1:15 (IF); 1:20 (IHC) | Santa Cruz Biotechnology, sc-390423 (IF), sc-13121 (IHC) |
| FGFR4 | monoclonal | 1:15 (IF); 1:15 (IHC) | Santa Cruz Biotechnology, sc-136988 |
| Actin | monoclonal | 1:5000 (WB) | Sigma Aldrich, A5441 |
| Gene Name | Accession no. for mRNA Sequence | Primer Sequence | Probe Sequence | Product Length |
|---|---|---|---|---|
| KL | NM_004795.4 | F: GCTCAACTCCCCCAGTCAGG R: TGTGGGCTTTGAGAGCTTCG | CCAGGGCA | 193 |
| FGFR1 | NM_001174063.2 | F: CTTAGGCAAACCCCTGGGAG R: ACAAGGGACCATCCTGCG | CTGCTGGG | 236 |
| FGFR2 | NM_000141.5 | F: CCGTGAAGATGTTGAAAGATGATGC R: GGTATTCTCGGAGGTTGCCT | CTTGGAGC | 183 |
| FGFR3 | NM_000142.5 | F: TGCTGAAAGACGATGCCACTG R: CTTGCAGGTGTCGAAGGAGT | ACCTGCTG | 224 |
| FGFR4 | NM_001291980.2 | F: TTGCCAGCTTCCTACCTGAG R: GCTGGAGGTCAAGGAGTCAC | CTCTGCCT | 110 |
| ACTB | NM_001101.5 | F: ATTGGCAATGAGCGGTTC R: GGATGCCACAGGACTCCA | CTTCCAGC | 76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Typiak, M.; Kulesza, T.; Rachubik, P.; Rogacka, D.; Audzeyenka, I.; Angielski, S.; Saleem, M.A.; Piwkowska, A. Role of Klotho in Hyperglycemia: Its Levels and Effects on Fibroblast Growth Factor Receptors, Glycolysis, and Glomerular Filtration. Int. J. Mol. Sci. 2021, 22, 7867. https://doi.org/10.3390/ijms22157867
Typiak M, Kulesza T, Rachubik P, Rogacka D, Audzeyenka I, Angielski S, Saleem MA, Piwkowska A. Role of Klotho in Hyperglycemia: Its Levels and Effects on Fibroblast Growth Factor Receptors, Glycolysis, and Glomerular Filtration. International Journal of Molecular Sciences. 2021; 22(15):7867. https://doi.org/10.3390/ijms22157867
Chicago/Turabian StyleTypiak, Marlena, Tomasz Kulesza, Patrycja Rachubik, Dorota Rogacka, Irena Audzeyenka, Stefan Angielski, Moin A. Saleem, and Agnieszka Piwkowska. 2021. "Role of Klotho in Hyperglycemia: Its Levels and Effects on Fibroblast Growth Factor Receptors, Glycolysis, and Glomerular Filtration" International Journal of Molecular Sciences 22, no. 15: 7867. https://doi.org/10.3390/ijms22157867
APA StyleTypiak, M., Kulesza, T., Rachubik, P., Rogacka, D., Audzeyenka, I., Angielski, S., Saleem, M. A., & Piwkowska, A. (2021). Role of Klotho in Hyperglycemia: Its Levels and Effects on Fibroblast Growth Factor Receptors, Glycolysis, and Glomerular Filtration. International Journal of Molecular Sciences, 22(15), 7867. https://doi.org/10.3390/ijms22157867