
Skin Mast Cell-Driven Ceramides Drive Early Apoptosis in Pre-Symptomatic Eczema in Mice


 , and
, and
Abstract
:1. Introduction
2. Results
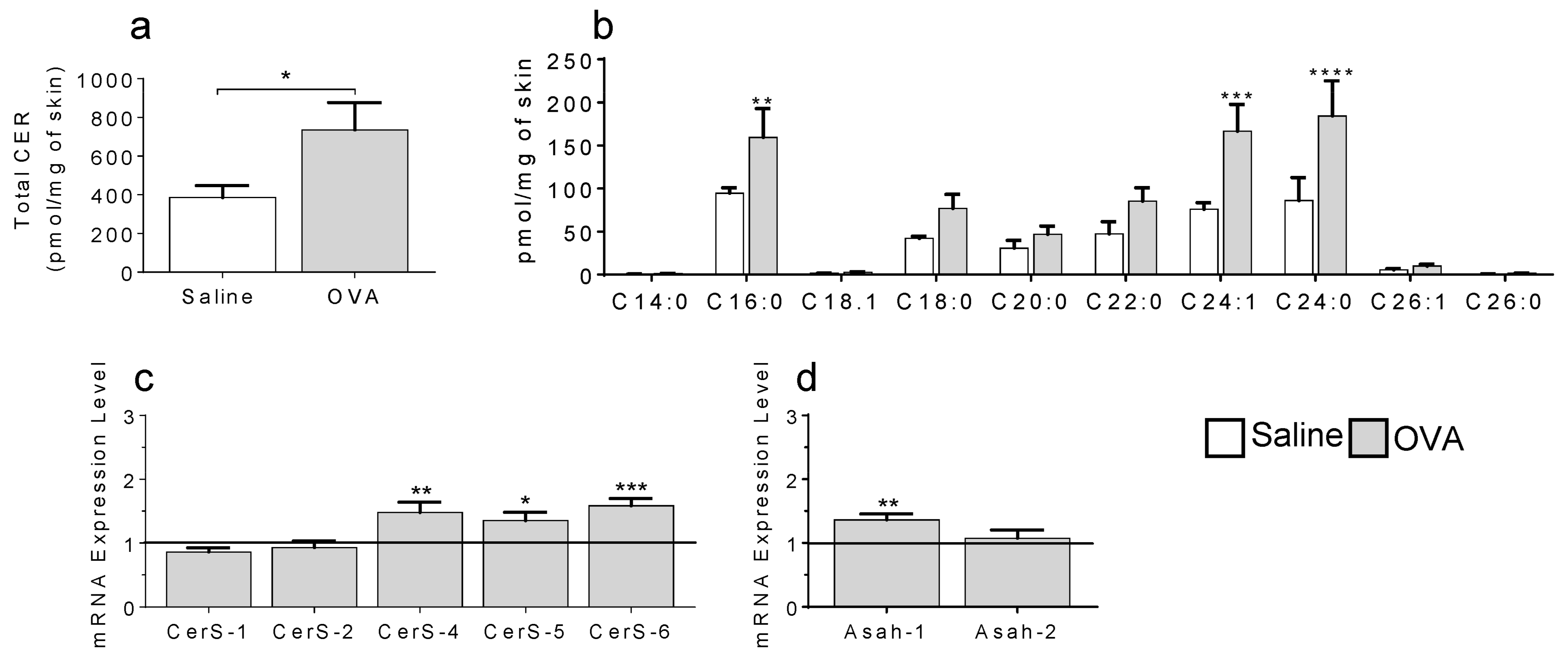
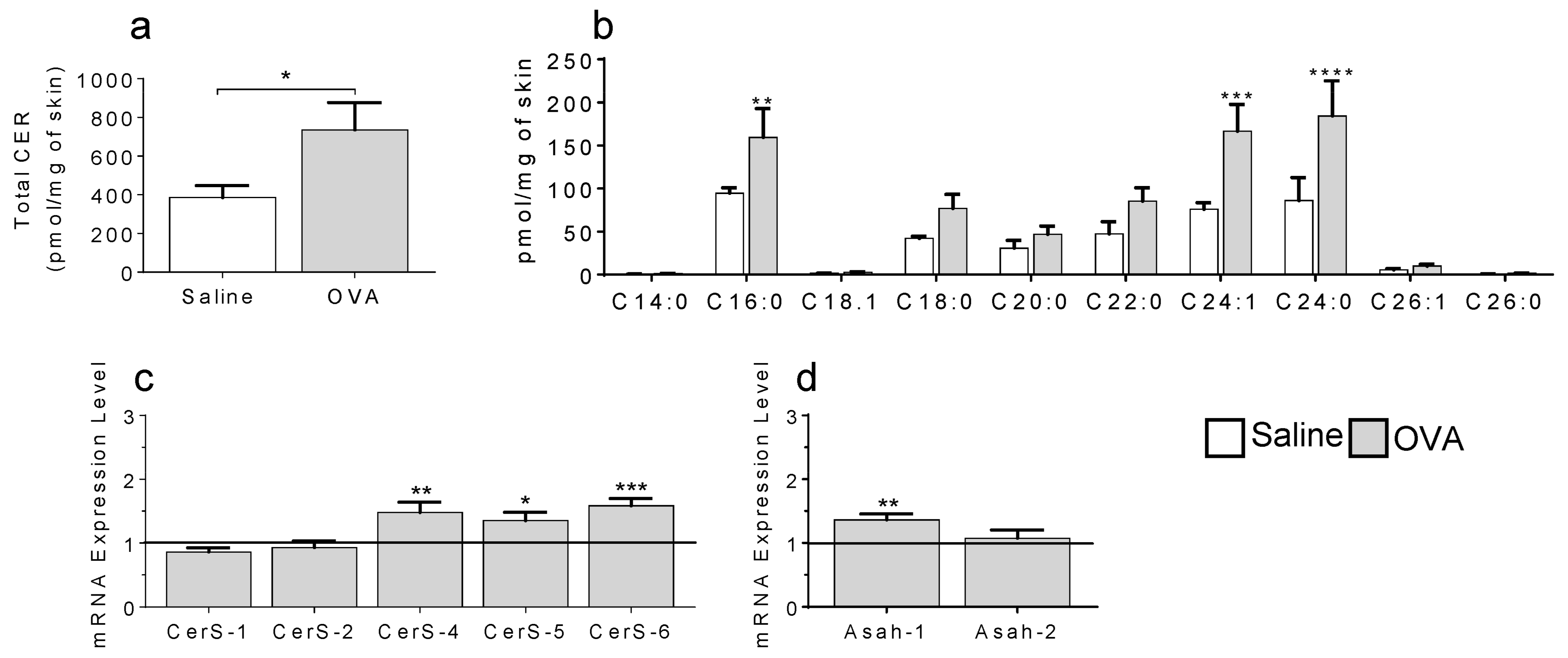
2.1. In a Model of Preclinical Atopic Dermatitis, a Single OVA Exposure Triggers an Increase in Skin Ceramides
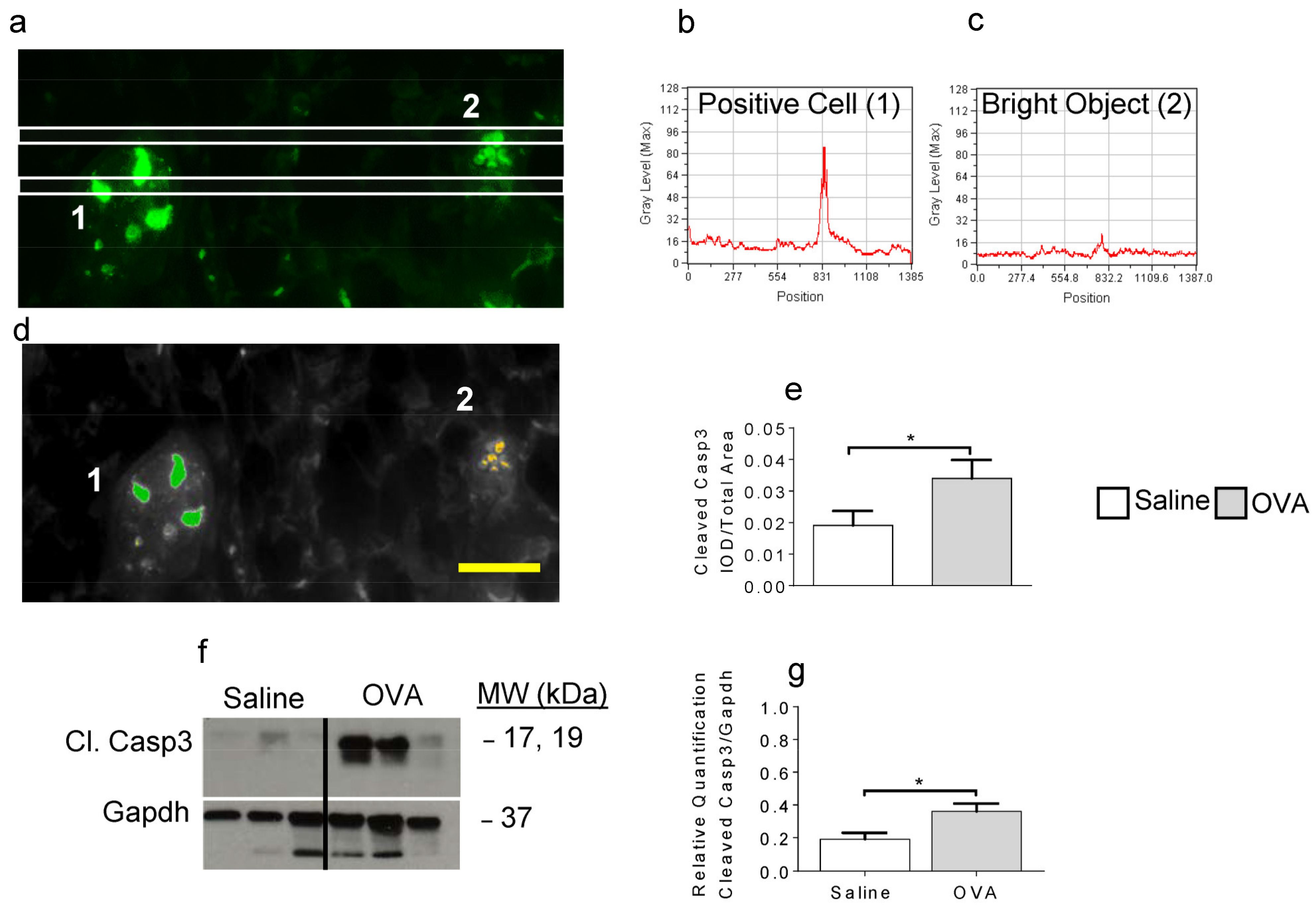
2.2. OVA Exposure Leads to Increased Detection of Cleaved Caspase-3
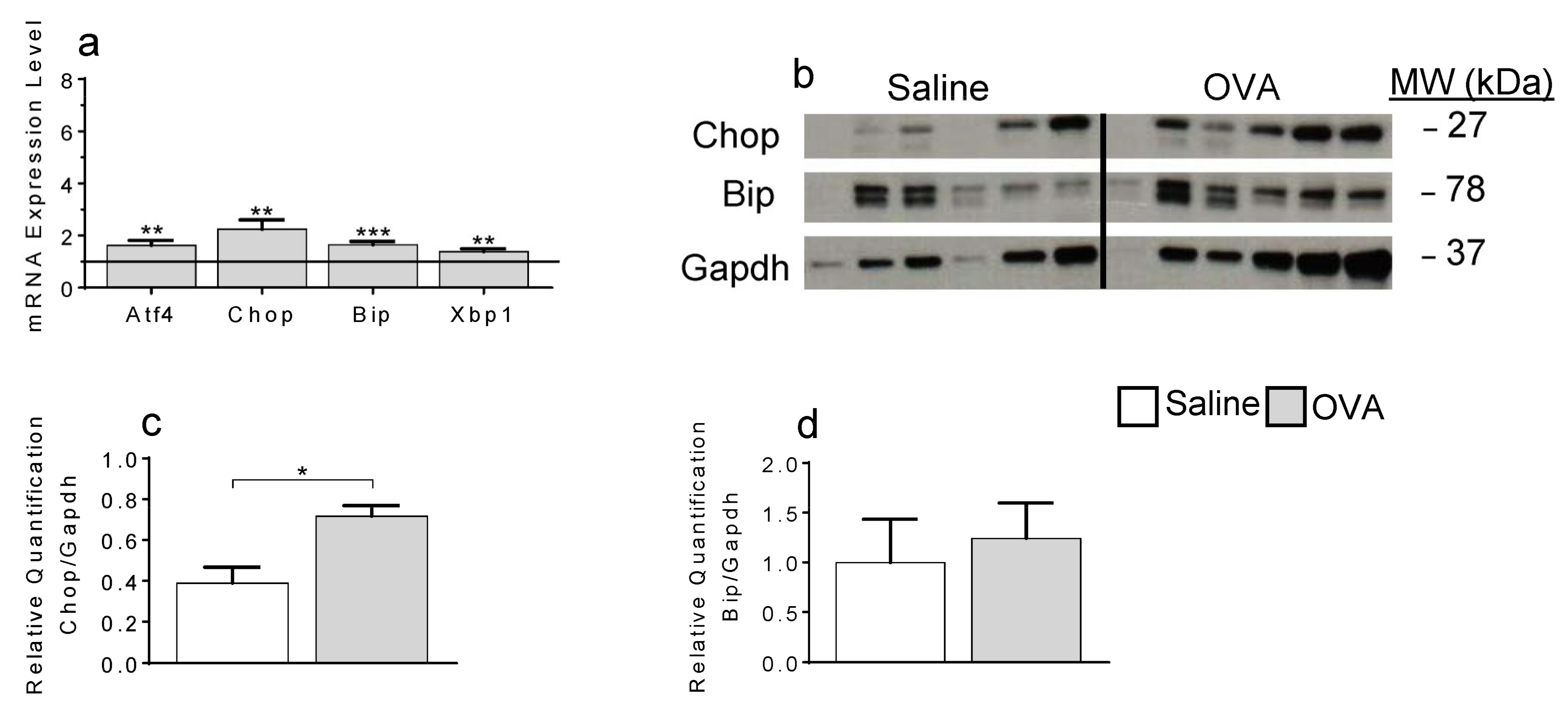
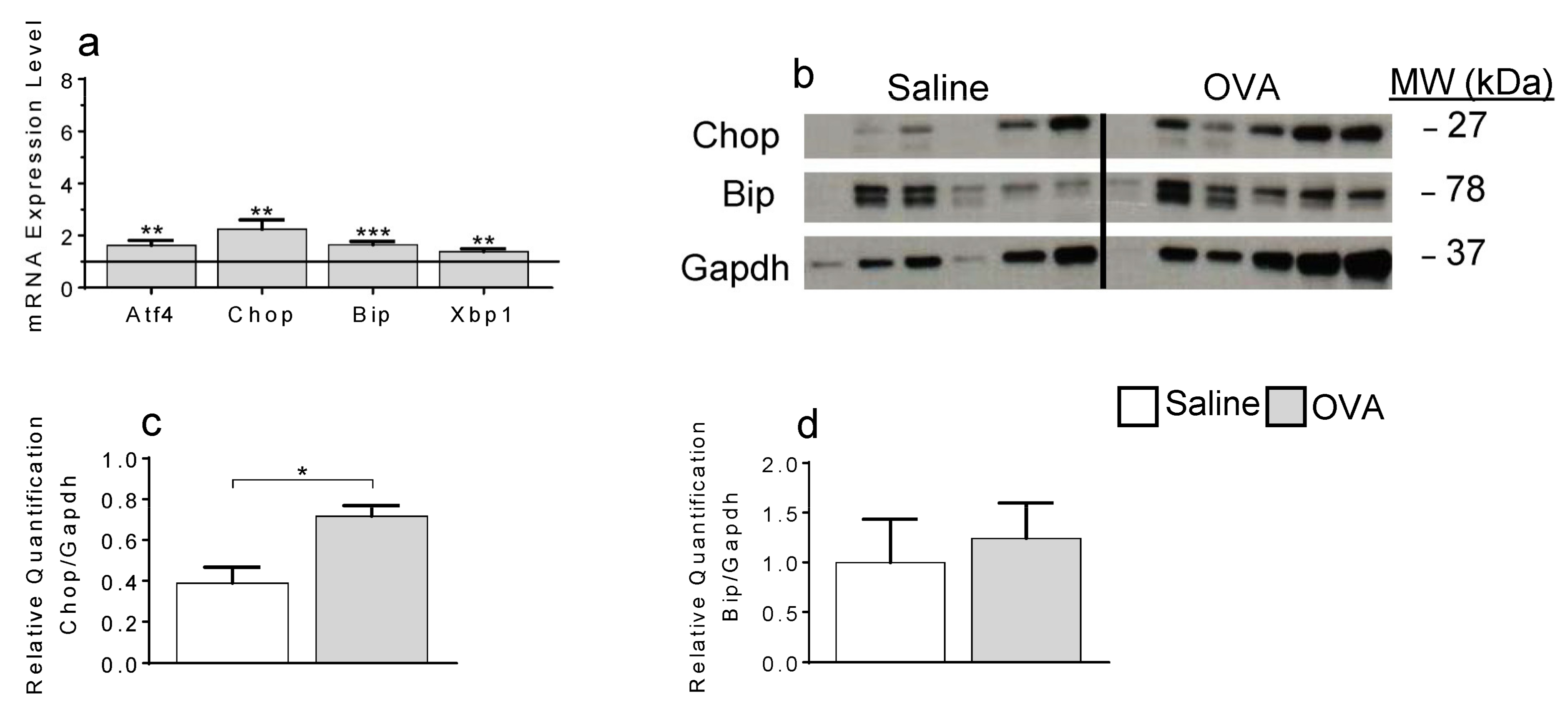
2.3. ER-Stress Contributes to Local Apoptosis Following OVA Exposure
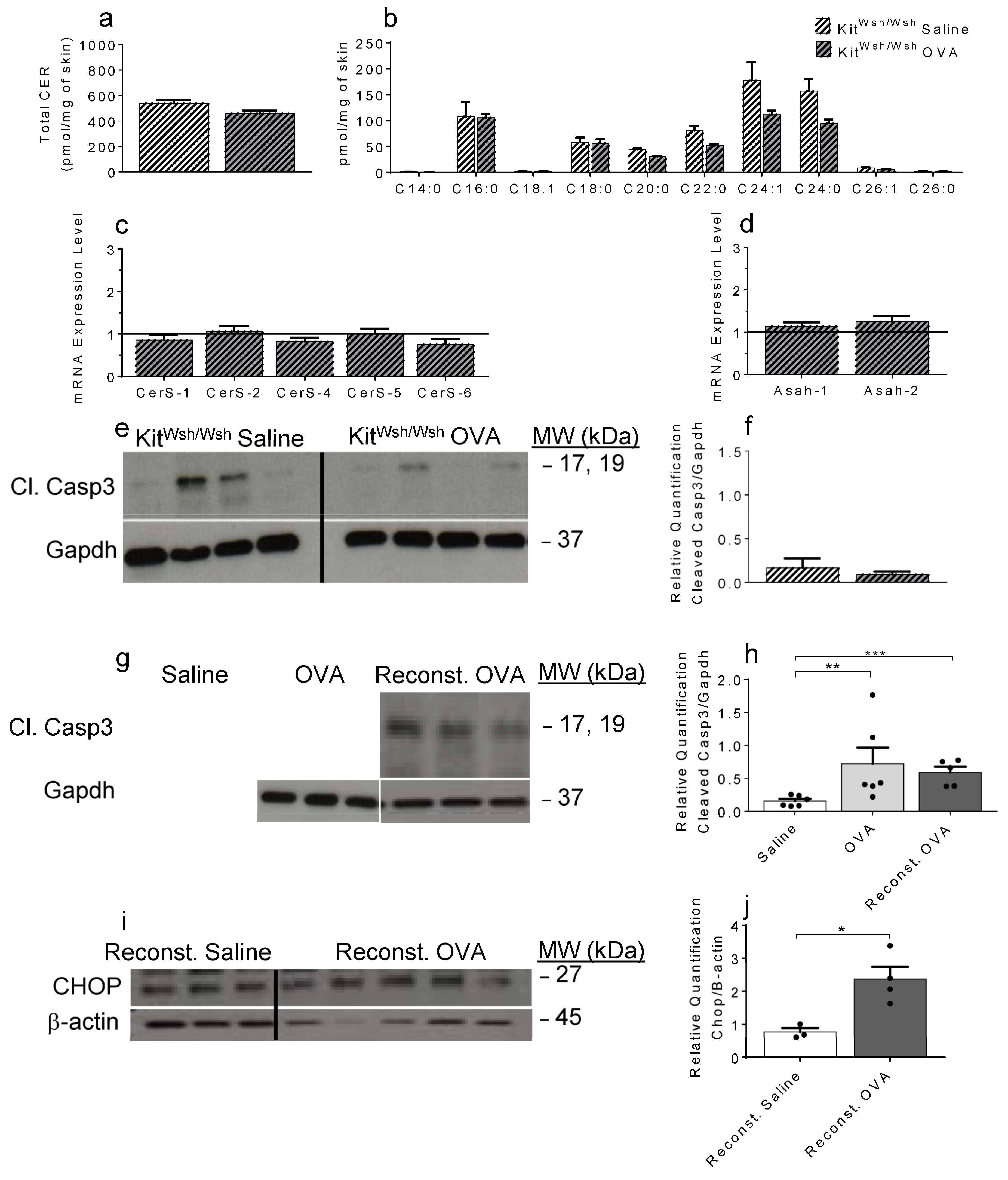
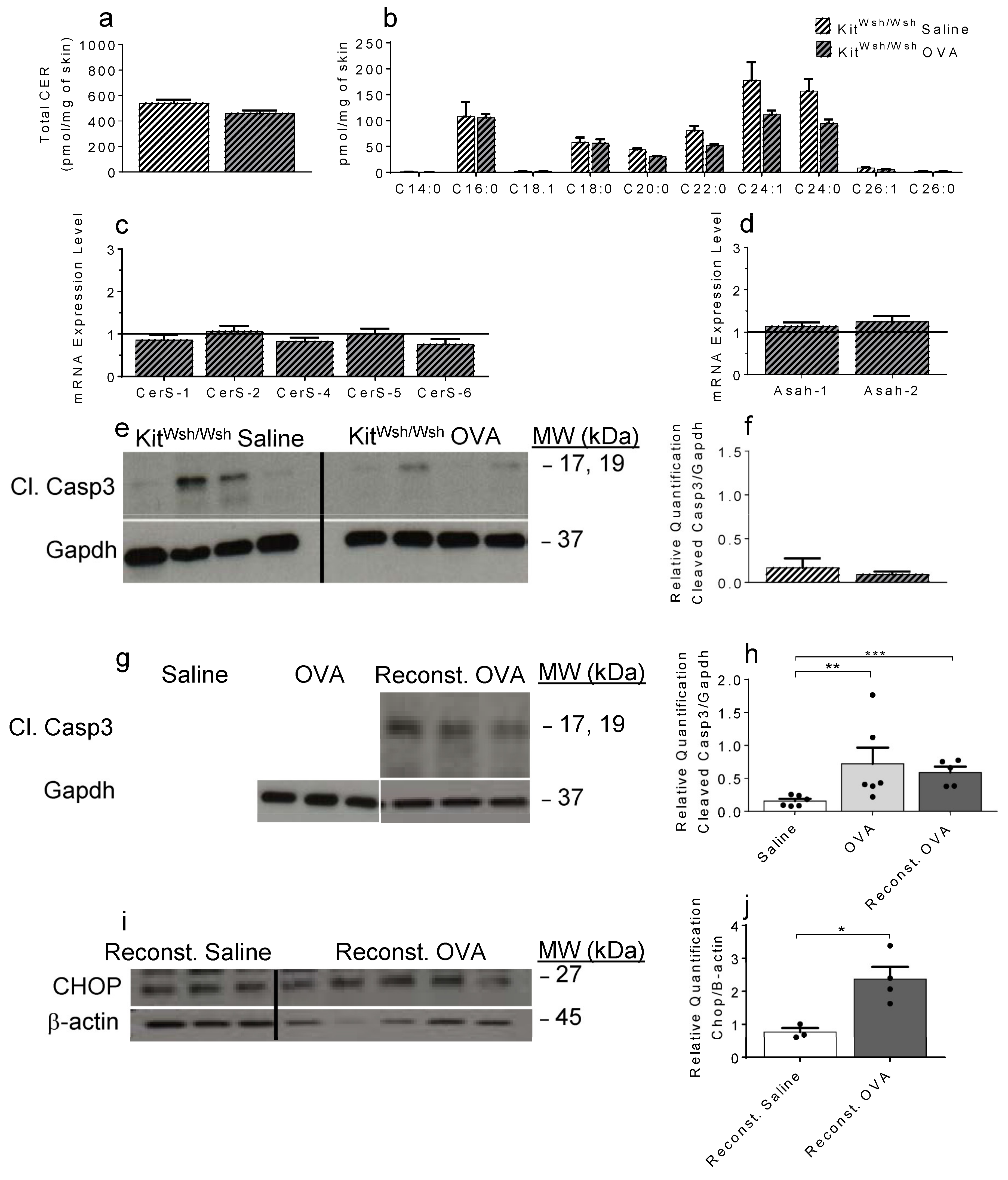
2.4. OVA-Induced Increase in Cer and the Resulting Apoptosis Do Not Occur in the Absence of Mast Cells
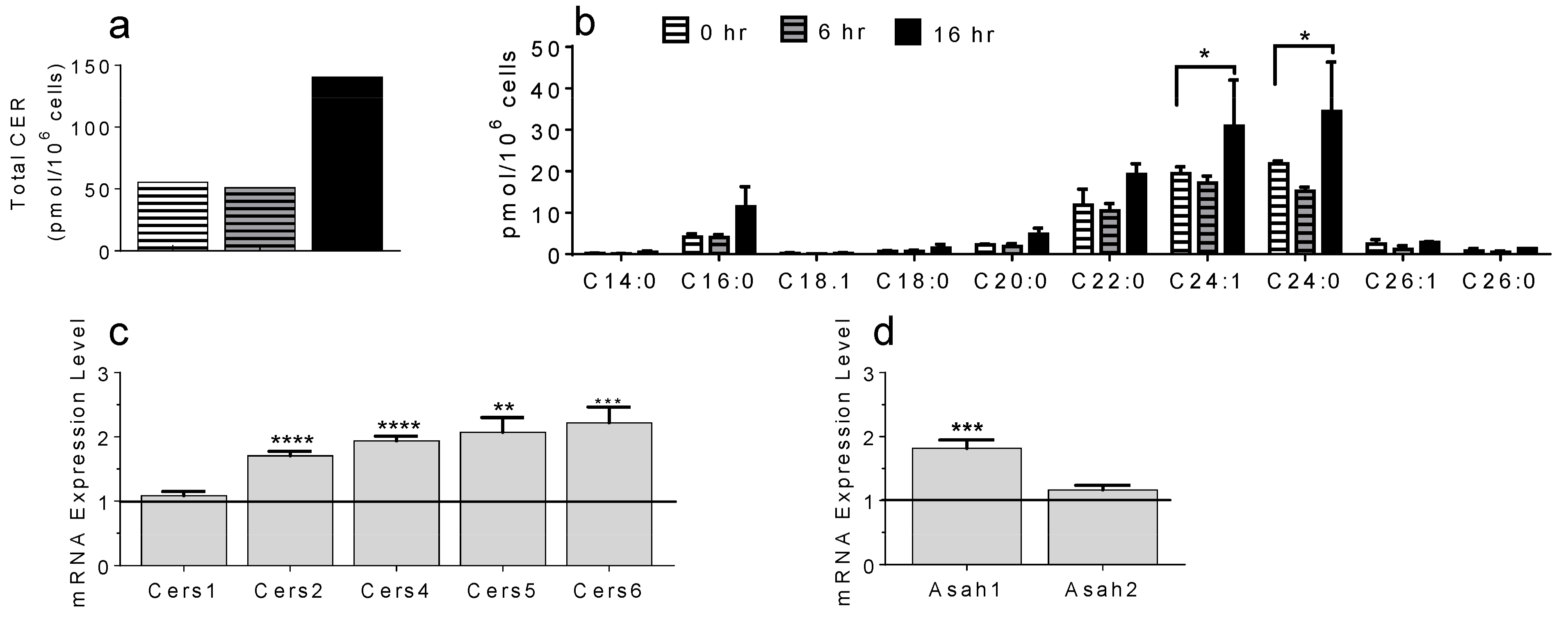
2.5. Bone Marrow-Derived MC, Activated In Vitro by Sphingosine-1-Phosphate, Recapitulate the Ceramide Profile Observed in OVA-Treated WT Mouse Skin Samples
3. Discussion
4. Materials and Methods
4.1. Atopic Dermatitis Model
4.2. Mass Spectrometric Measurements of Cer
4.3. QRTPCR
4.4. Immunofluorescence Quantification of Cleaved Caspase-3 in Skin Sections
4.5. Western Blots
4.6. Culture and Activation of Bone Marrow-Derived Mast Cells
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Skolová, B.; Hudská, K.; Pullmannová, P.; Kováčik, A.; Palát, K.; Roh, J.; Fleddermann, J.; Estrela-Lopis, I.; Vávrová, K. Different phase behavior and packing of ceramides with long (C16) and very long (C24) acyls in model membranes: Infrared spectroscopy using deuterated lipids. J. Phys. Chem. B 2014, 118, 10460–10470. [Google Scholar] [CrossRef]
- Elias, P.M. Lipid abnormalities and lipid-based repair strategies in atopic dermatitis. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2014, 1841, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Janssens, M.; van Smeden, J.; Gooris, G.S.; Bras, W.; Portale, G.; Caspers, P.J.; Vreeken, R.; Hankemeier, T.; Kezic, S.; Wolterbeek, R.; et al. Increase in short-chain ceramides correlates with an altered lipid organization and decreased barrier function in atopic eczema patients. J. Lipid Res. 2012, 53, 2755–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Smeden, J.; Hoppel, L.; van der Heijden, R.; Hankemeier, T.; Vreeken, R.; Bouwstra, J.A. LC/MS analysis of stratum corneum lipids: Ceramide profiling and discovery. J. Lipid Res. 2011, 52, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Aflaki, E.; Doddapattar, P.; Radović, B.; Povoden, S.; Kolb, D.; Vujic, N.; Wegscheider, M.; Koefeler, H.; Hornemann, T.; Graier, W.; et al. C16 ceramide is crucial for triacylglycerol-induced apoptosis in macrophages. Cell Death Dis. 2012, 3, e280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seumois, G.; Fillet, M.; Gillet, L.; Faccinetto, C.; Desmet, C.; François, C.; Dewals, B.; Oury, C.; Vanderplasschen, A.; Lekeux, P.; et al. De novo C16- and C24-ceramide generation contributes to spontaneous neutrophil apoptosis. J. Leukoc. Biol. 2007, 81, 1477–1486. [Google Scholar] [CrossRef]
- Grösch, S.; Schiffmann, S.; Geisslinger, G. Chain length-specific properties of ceramides. Prog. Lipid Res. 2012, 51, 50–62. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Riley, R.T.; Merrill, A.H.J. Ceramide synthase inhibition by fumonisins: A perfect storm of perturbed sphingolipid metabolism, signaling, and disease. J. Lipid Res. 2019, 60, 1183–1189. [Google Scholar] [CrossRef] [Green Version]
- Custodia, A.; Aramburu-Núñez, M.; Correa-Paz, C.; Posado-Fernández, A.; Gómez-Larrauri, A.; Castillo, J.; Gómez-Muñoz, A.; Sobrino, T.; Ouro, A. Ceramide metabolism and Parkinson’s disease—Therapeutic targets. Biomolecules 2021, 11, 945. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-H.; Jang, W.-H.; Seo, J.A.; Park, M.; Lee, T.R.; Park, Y.-H.; Kim, D.K.; Lim, K.-M. Decrease of ceramides with very long–chain fatty acids and downregulation of elongases in a murine atopic dermatitis model. J. Investig. Dermatol. 2012, 132, 476–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdyshev, E.; Goleva, E.; Bronova, I.; Dyjack, N.; Rios, C.; Jung, J.; Taylor, P.; Jeong, M.; Hall, C.F.; Richers, B.N.; et al. Lipid abnormalities in atopic skin are driven by type 2 cytokines. JCI Insight 2018, 3, e98006. [Google Scholar] [CrossRef] [Green Version]
- Spergel, J.; Mizoguchi, E.; Brewer, J.P.; Martin, T.R.; Bhan, A.K.; Geha, R.S. Epicutaneous sensitization with protein antigen induces localized allergic dermatitis and hyperresponsiveness to methacholine after single exposure to aerosolized antigen in mice. J. Clin. Investig. 1998, 101, 1614–1622. [Google Scholar] [CrossRef] [PubMed]
- Wedman, P.A.; Aladhami, A.; Chumanevich, A.P.; Fuseler, J.W.; Oskeritzian, C.A. Mast cells and sphingosine-1-phosphate underlie prelesional remodeling in a mouse model of eczema. Allergy 2018, 73, 405–415. [Google Scholar] [CrossRef]
- Thomas, R.L.; Matsko, C.M.; Lotze, M.T.; Amoscato, A.A. Mass spectrometric identification of increased C16 ceramide levels during apoptosis. J. Biol. Chem. 1999, 274, 30580–30588. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Larrauri, A.; Presa, N.; Dominguez-Herrera, A.; Ouro, A.; Trueba, M.; Gomez-Muñoz, A. Role of bioactive sphingolipids in physiology and pathology. Essays Biochem. 2020, 64, 579–589. [Google Scholar] [CrossRef]
- Duan, W.R.; Garner, D.S.; Williams, S.D.; Funckes-Shippy, C.L.; Spath, I.S.; Blomme, E.A.G. Comparison of immunohistochemistry for activated caspase-3 and cleaved cytokeratin 18 with the TUNEL method for quantification of apoptosis in histological sections of PC-3 subcutaneous xenografts. J. Pathol. 2003, 199, 221–228. [Google Scholar] [CrossRef]
- Rao, R.V.; Ellerby, H.M.; Bredesen, D.E.; Rao, R.V.; Ellerby, H.M. Coupling endoplasmic reticulum stress to the cell death program. Cell Death Differ. 2004, 11, 372–380. [Google Scholar] [CrossRef] [Green Version]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oskeritzian, C.A.; Alvarez, S.E.; Hait, N.C.; Price, M.M.; Milstien, S.; Spiegel, S. Distinct roles of sphingosine kinases 1 and 2 in human mast-cell functions. Blood 2008, 111, 4193–4200. [Google Scholar] [CrossRef]
- Oskeritzian, C.A.; Price, M.M.; Hait, N.C.; Kapitonov, D.; Falanga, Y.T.; Morales, J.K.; Ryan, J.J.; Milstien, S.; Spiegel, S. Essential roles of sphingosine-1–phosphate receptor 2 in human mast cell activation, anaphylaxis, and pulmonary edema. J. Exp. Med. 2010, 207, 465–474. [Google Scholar] [CrossRef]
- Oskeritzian, C.A.; Hait, N.C.; Wedman, P.; Chumanevich, A.; Kolawole, E.M.; Price, M.M.; Falanga, Y.T.; Harikumar, K.; Ryan, J.J.; Milstien, S.; et al. The sphingosine-1-phosphate/sphingosine-1-phosphate receptor 2 axis regulates early airway T-cell infiltration in murine mast cell–dependent acute allergic responses. J. Allergy Clin. Immunol. 2015, 135, 1008–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaess, M.; Deigner, H.-P. Derailed ceramide metabolism in Atopic Dermatitis (AD): A causal starting point for a personalized (basic) therapy. Int. J. Mol. Sci. 2019, 20, 3967. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Lee, S.; Lee, Y.-M. Sphingolipids and antimicrobial peptides: Function and roles in atopic dermatitis. Biomol. Ther. 2013, 21, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedman, P.; Aladhami, A.; Beste, M.; Edwards, M.K.; Chumanevich, A.; Fuseler, J.W.; Oskeritzian, C.A. A new image analysis method based on morphometric and fractal parameters for rapid evaluation of in situ mammalian mast cell status. Microsc. Microanal. 2015, 21, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Futerman, A.H.; Riezman, H. The ins and outs of sphingolipid synthesis. Trends Cell Biol. 2005, 15, 312–318. [Google Scholar] [CrossRef]
- Longato, L.; Ripp, K.; Setshedi, M.; Dostalek, M.; Akhlaghi, F.; Branda, M.; Wands, J.R.; De La Monte, S.M. Insulin resistance, ceramide accumulation, and endoplasmic reticulum stress in human chronic alcohol-related liver disease. Oxidative Med. Cell. Longev. 2012, 2012, 479348. [Google Scholar] [CrossRef] [Green Version]
- Oyadomari, S.; Mori, M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2003, 11, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Grimbaldeston, M.A.; Chen, C.-C.; Piliponsky, A.M.; Tsai, M.; Tam, S.-Y.; Galli, S.J. Mast cell-deficient W-sash c-kit mutant Kitw-sh/W-sh mice as a model for investigating mast cell biology in vivo. Am. J. Pathol. 2005, 167, 835–848. [Google Scholar] [CrossRef] [Green Version]
- Mullen, T.D.; Jenkins, R.W.; Clarke, C.J.; Bielawski, J.; Hannun, Y.A.; Obeid, L.M. Ceramide synthase-dependent ceramide generation and programmed cell death: Involvement of salvage pathway in regulating postmitochondrial events. J. Biol. Chem. 2011, 286, 15929–15942. [Google Scholar] [CrossRef] [Green Version]
- Karahatay, S.; Thomas, K.; Koybasi, S.; Senkal, C.E.; ElOjeimy, S.; Liu, X.; Bielawski, J.; Day, T.A.; Gillespie, M.B.; Sinha, D.; et al. Clinical relevance of ceramide metabolism in the pathogenesis of human head and neck squamous cell carcinoma (HNSCC): Attenuation of C18-ceramide in HNSCC tumors correlates with lymphovascular invasion and nodal metastasis. Cancer Lett. 2007, 256, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Schiffmann, S.; Sandner, J.; Birod, K.; Wobst, I.; Angioni, C.; Ruckhäberle, E.; Kaufmann, M.; Ackermann, H.; Lotsch, J.; Schmidt, H.; et al. Ceramide synthases and ceramide levels are increased in breast cancer tissue. Carcinogenesis 2009, 30, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Jung, H.; Kim, M.; Moon, J.; Ban, G.; Kim, S.J.; Yoo, H.; Park, H. Ceramide/sphingosine-1-phosphate imbalance is associated with distinct inflammatory phenotypes of uncontrolled asthma. Allergy 2020, 75, 1987–2000. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Mitsutake, S.; Tsuji, K.; Kihara, A.; Igarashi, Y. Ceramide biosynthesis in keratinocyte and its role in skin function. Biochimie 2009, 91, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Tirodkar, T.S.; Lu, P.; Bai, A.; Scheffel, M.J.; Gencer, S.; Garrett-Mayer, E.; Bielawska, A.; Ogretmen, B.; Voelkel-Johnson, C. Expression of ceramide synthase 6 transcriptionally activates acid ceramidase in a c-Jun N-terminal Kinase (JNK)-dependent manner. J. Biol. Chem. 2015, 290, 13157–13167. [Google Scholar] [CrossRef] [Green Version]
- Kanto, T.; Kalinski, P.; Hunter, O.C.; Lotze, M.T.; Amoscato, A. Ceramide mediates tumor-induced dendritic cell apoptosis. J. Immunol. 2001, 167, 3773–3784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-C.; Wang, J.D.; Svoboda, K.K.; Casillas, R.P.; Laskin, J.D.; Gordon, M.K.; Gerecke, N.R. Sulfur mustard induces an endoplasmic reticulum stress response in the mouse ear vesicant model. Toxicol. Appl. Pharmacol. 2013, 268, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Oyeniran, C.; Sturgill, J.L.; Hait, N.C.; Huang, W.-C.; Avni, D.; Maceyka, M.; Newton, J.; Allegood, J.C.; Montpetit, A.; Conrad, D.H.; et al. Aberrant ORM (yeast)–like protein isoform 3 (ORMDL3) expression dysregulates ceramide homeostasis in cells and ceramide exacerbates allergic asthma in mice. J. Allergy Clin. Immunol. 2015, 136, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chumanevich, A.P.; Wedman, P.A.; Oskeritzian, C.A. Methods for analyzing sphingosine-1-phosphate signaling in human and mouse primary mast cells. Adv. Struct. Saf. Stud. 2017, 1697, 21–30. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward Sequence | Reverse Sequence |
|---|---|---|
| Mu Gapdh | CAG AAG GGG CGG AGA TGA T | AGG CCG GTG CTG CTG AGT ATG TC |
| Mu Cers1 | CCA CCA CAC ACA TCT TTC GG | GGA GCA GGT AAG CGC AGT AG |
| Mu Cers2 | ATG CTC CAG ACC TTG TAT GAC T | CTG AGG CTT TGG CAT AGA CAC |
| MuCers4 | TAC CCA CAT CAG ACC CTG AAT | TGA AGT CCT TGC GTT TGA CAT C |
| Mu Cers5 | CGG GGA AAG GTG TCT AAG GAT | GTT CAT GCA GTT GGC ACC ATT |
| Mu Cers6 | ATT CAA CGC TGG TTT CGA CAA | TTC AAG AAC CGG ACT CCG TAG |
| Mu Asah1 | CAC CAG CGT TGA GGA TTT TAG T | TAC CAG GCA GCT TTT GAT CCA |
| Mu Asah2 | GCA AAG CGA ACC TTC TCC AC | ACT GGT AAC AAA CAA GAG GGT GA |
| Mu Casp3 | ATG GAG AAC AAC AAA ACC TCA GT | TTG CTC CCA TGT ATG GTC TTT AC |
| Mu Atf4 | GGG TTC TGT CTT CCA CTC CA | AAG CAG CAG AGT CAG GCT TTC |
| Mu Chop | CCA CCA CAC CTG AAA GCA GAA | AGG TGA AAG GCA GGG ACT CA |
| Mu Bip | TTC AGC CAA TTA TCA GCA AAC TCT | TTT TCT GAT GTA TCC TCT TCA CCA GT |
| Mu Xbp1 | GAA CCA GGA GTT AAG AAC ACG | AGG CAA CAG TGT CAG AGT CC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robida, P.A.; Chumanevich, A.P.; Gandy, A.O.; Fuseler, J.W.; Nagarkatti, P.; Nagarkatti, M.; Oskeritzian, C.A. Skin Mast Cell-Driven Ceramides Drive Early Apoptosis in Pre-Symptomatic Eczema in Mice. Int. J. Mol. Sci. 2021, 22, 7851. https://doi.org/10.3390/ijms22157851
Robida PA, Chumanevich AP, Gandy AO, Fuseler JW, Nagarkatti P, Nagarkatti M, Oskeritzian CA. Skin Mast Cell-Driven Ceramides Drive Early Apoptosis in Pre-Symptomatic Eczema in Mice. International Journal of Molecular Sciences. 2021; 22(15):7851. https://doi.org/10.3390/ijms22157851
Chicago/Turabian StyleRobida, Piper A., Alena P. Chumanevich, Alexa Orr Gandy, John W. Fuseler, Prakash Nagarkatti, Mitzi Nagarkatti, and Carole A. Oskeritzian. 2021. "Skin Mast Cell-Driven Ceramides Drive Early Apoptosis in Pre-Symptomatic Eczema in Mice" International Journal of Molecular Sciences 22, no. 15: 7851. https://doi.org/10.3390/ijms22157851
APA StyleRobida, P. A., Chumanevich, A. P., Gandy, A. O., Fuseler, J. W., Nagarkatti, P., Nagarkatti, M., & Oskeritzian, C. A. (2021). Skin Mast Cell-Driven Ceramides Drive Early Apoptosis in Pre-Symptomatic Eczema in Mice. International Journal of Molecular Sciences, 22(15), 7851. https://doi.org/10.3390/ijms22157851