
Exogenous Brassinosteroid Facilitates Xylem Development in Pinus massoniana Seedlings


Abstract
:1. Introduction
2. Results
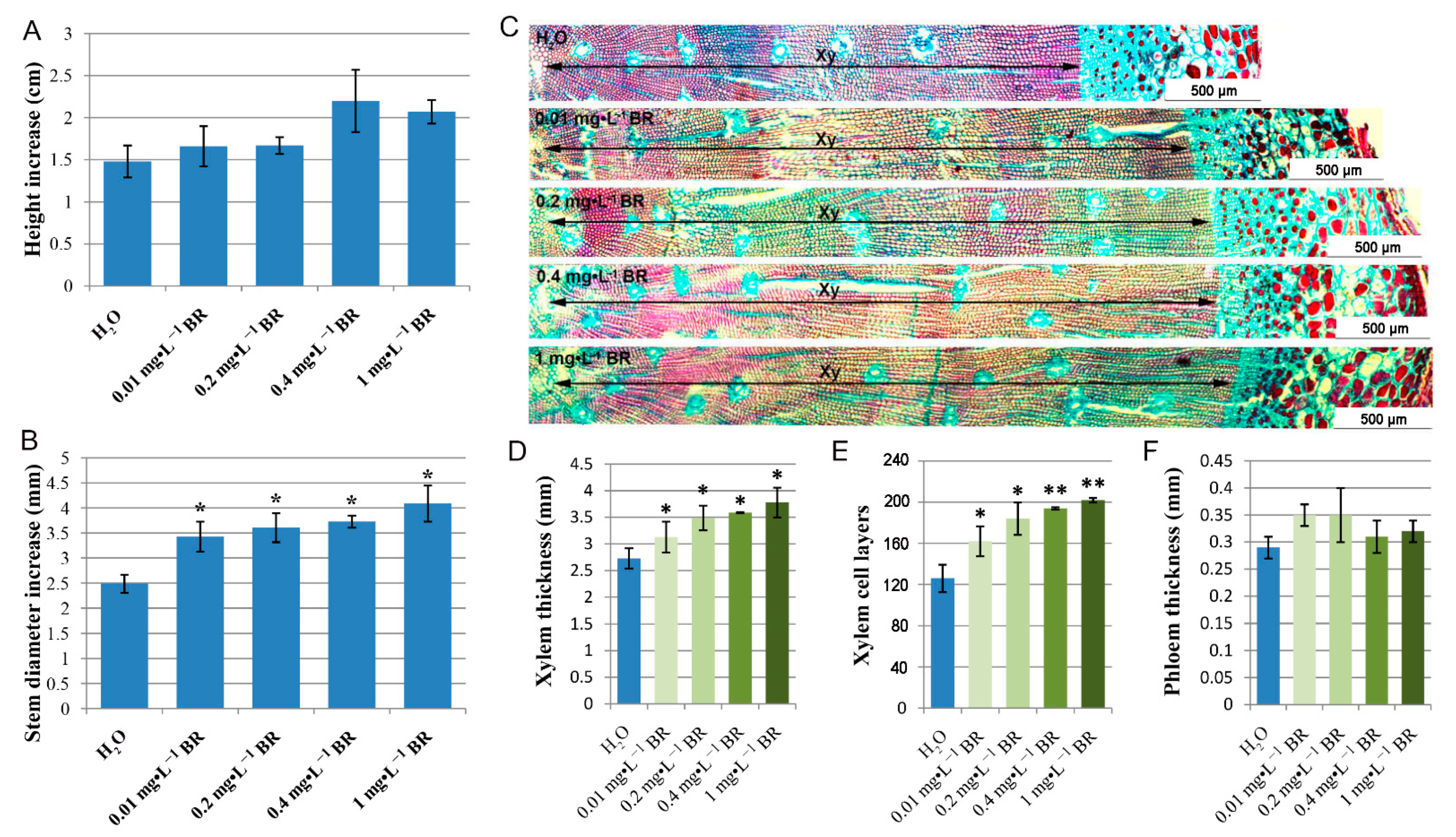
2.1. BR Treatment Facilitates Stem Growth and Xylem Development during Secondary Growth
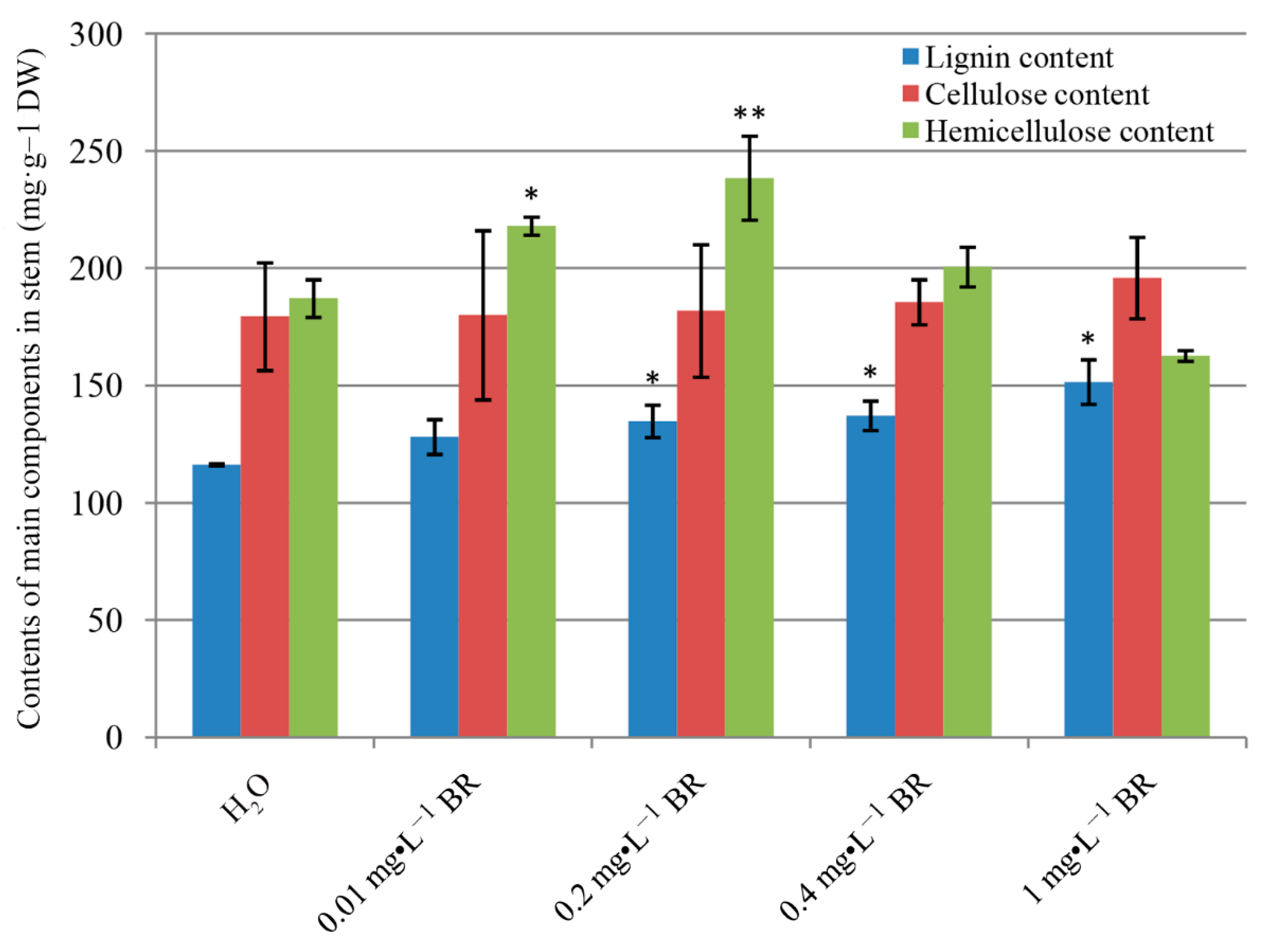
2.2. Effect of Exogenous Application of BR on the Contents of Lignin, Cellulose and Hemicellulose
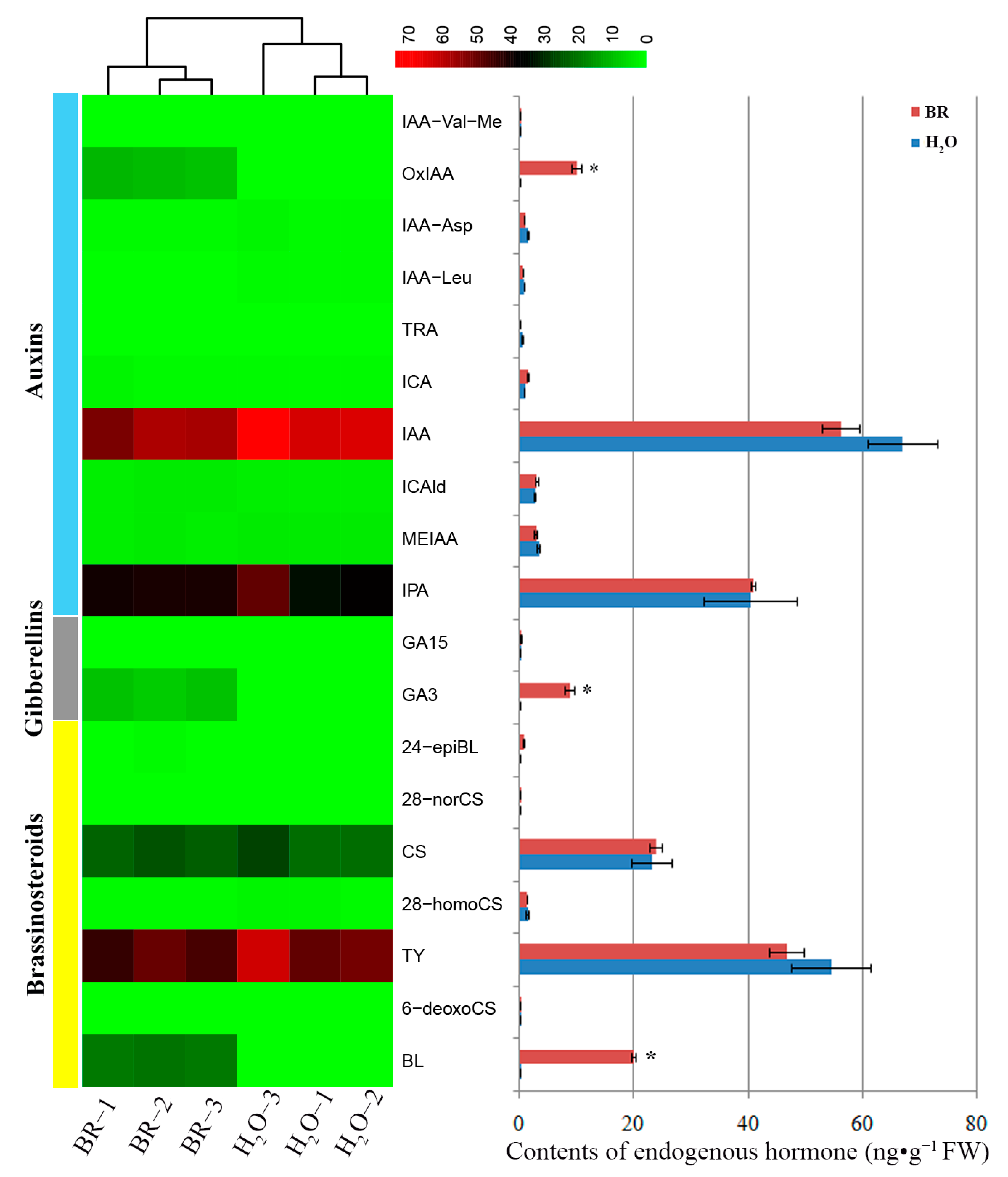
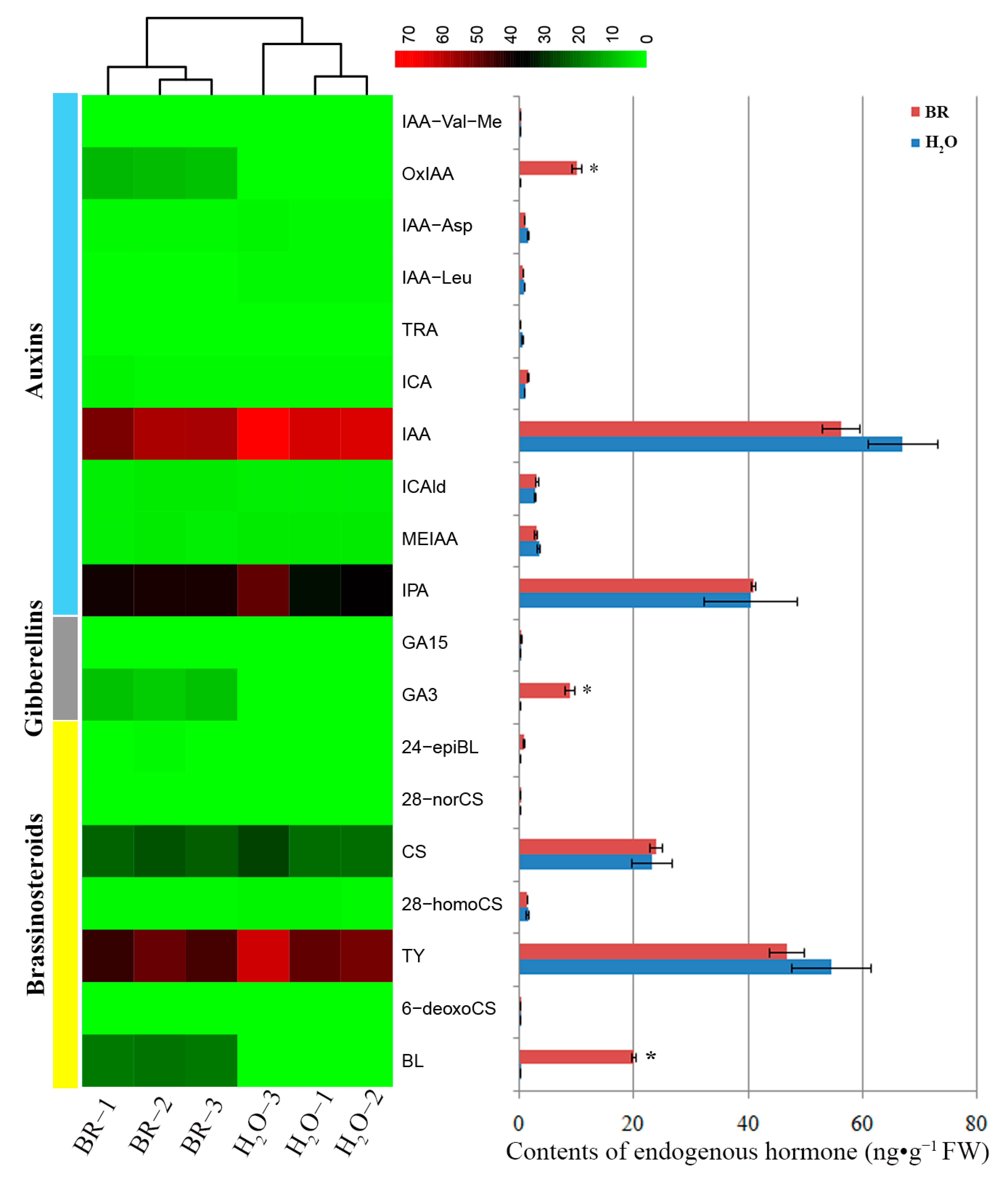
2.3. Changes of Endogenous Phytohormone Levels in Seedling Stems in Response to BR Treatment
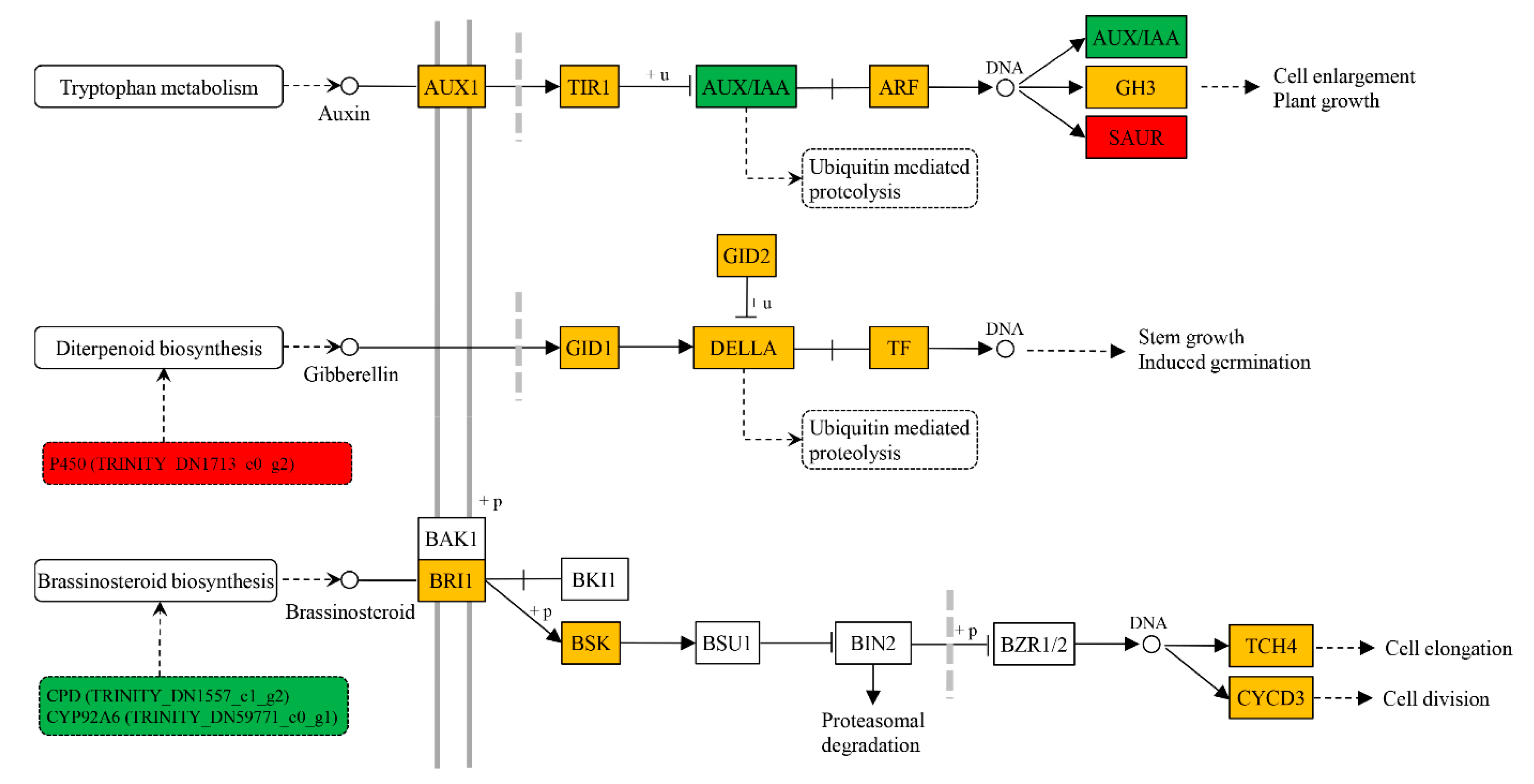
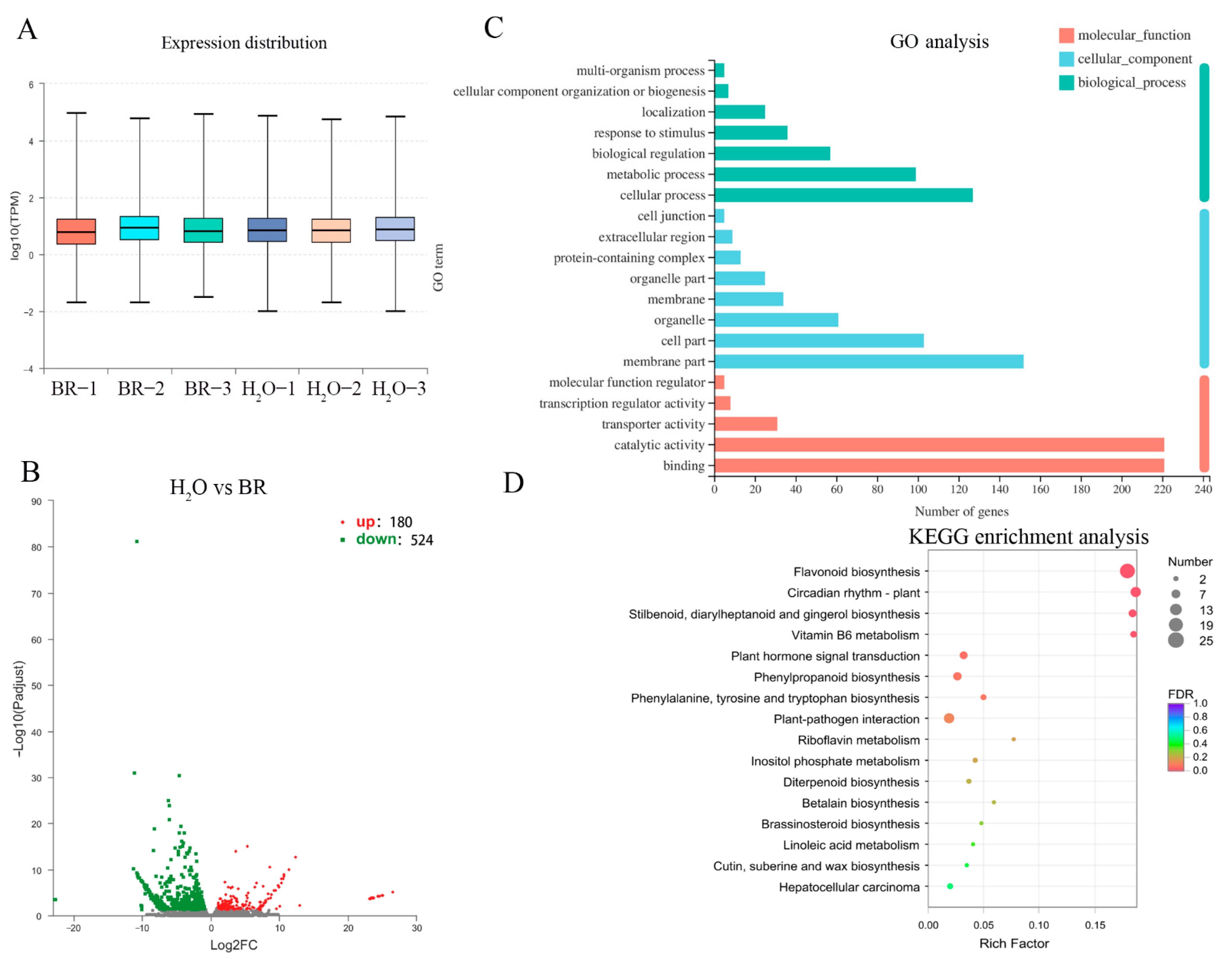
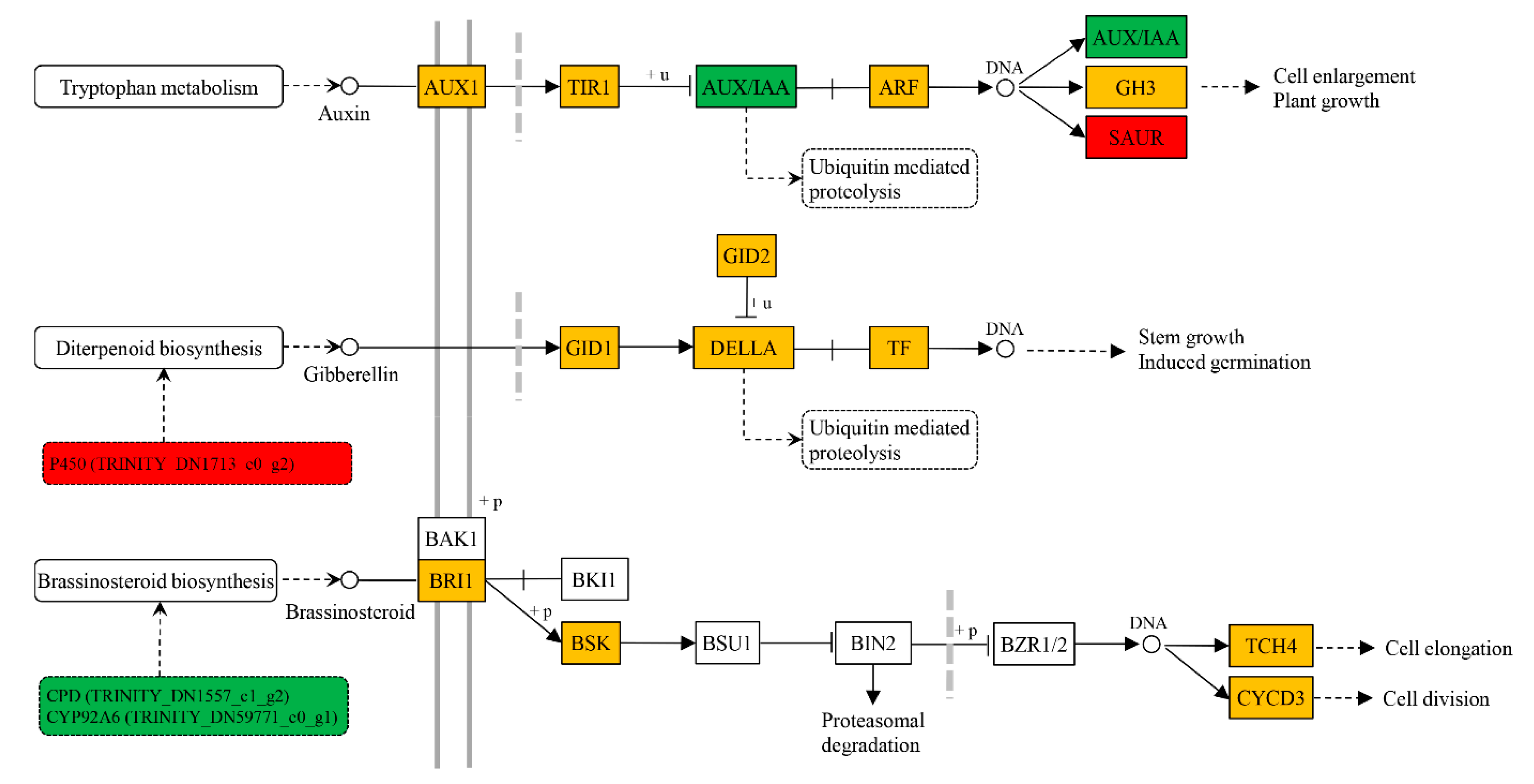
2.4. Transcription Level of Genes Associated with Xylem Development in Response to BR Treatment
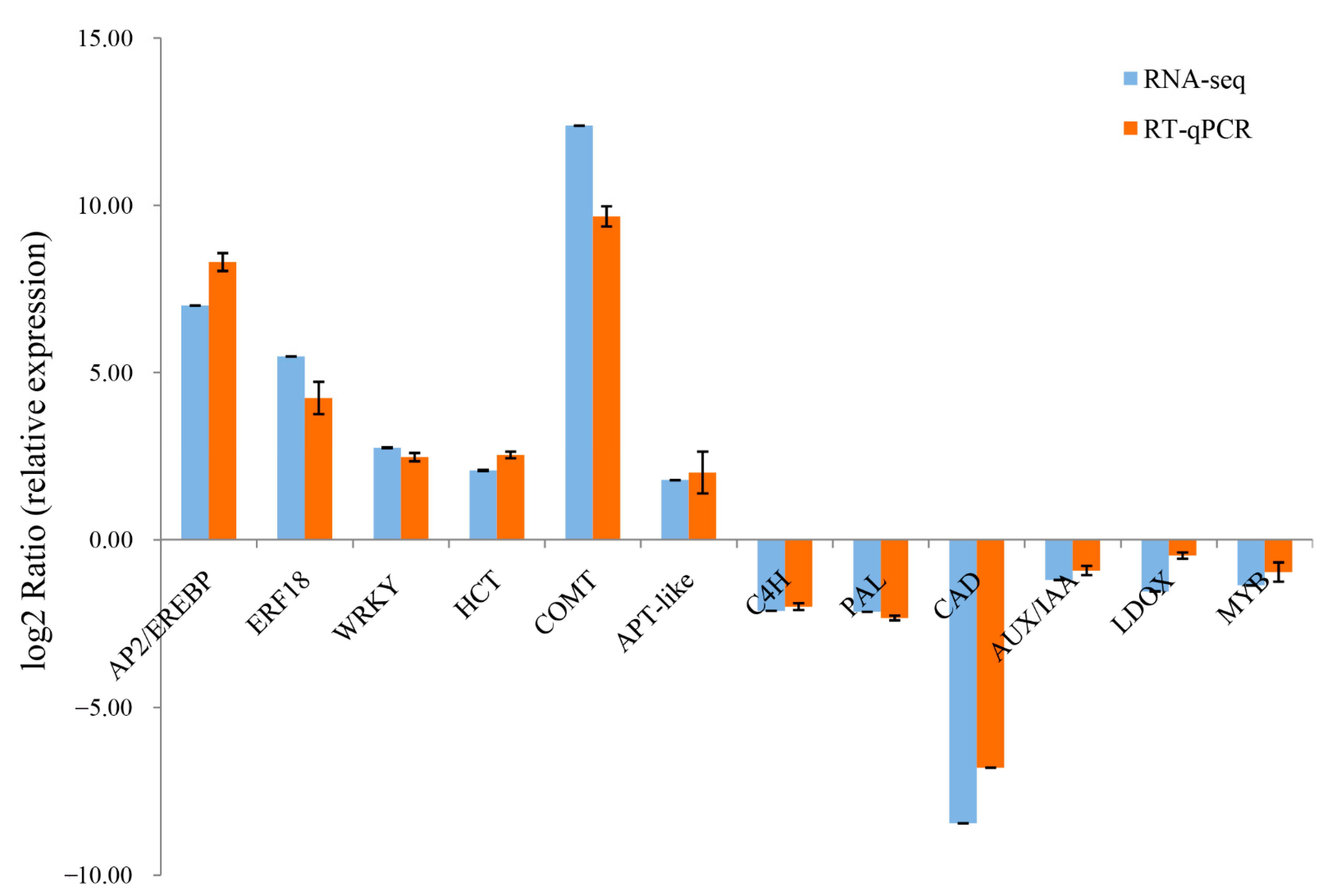
2.5. RT-qPCR Validation of DEGs from the RNA-Seq
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Morphology, Histology and Imaging
4.3. Quantitative Determination of Lignin, Cellulose and Hemicellulose
4.4. Analysis of Phytohormones Levels by LC-MS/MS
4.5. RNA Isolation, cDNA Library Construction and RNA-Seq
4.6. Data Processing and Bioinformatics Analysis
4.7. Validation of Gene Expression Profiles via RT-qPCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nieminen, K.M.; Kauppinen, L.; Helariutta, Y. A weed for wood? Arabidopsis as a genetic model for xylem development. Plant Physiol. 2004, 135, 6536–6559. [Google Scholar] [CrossRef] [Green Version]
- Milhinhos, A.; Miguel, C.M. Hormone interactions in xylem development: A matter of signals. Plant Cell Rep. 2013, 32, 867–883. [Google Scholar] [CrossRef]
- Yuan, H.; Zhao, L.; Guo, W.; Yu, Y.; Tan, L.; Zhang, L.; Song, X.; Huang, W.; Cheng, L.; Chen, J.; et al. Exogenous application of phytohormones promotes growth and regulates expression of wood formation-related genes in Populus simonii × P. nigra. Int. J. Mol. Sci. 2019, 20, 792. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Gerttula, S.; Li, Z.; Zhao, S.T.; Liu, Y.L.; Liu, Y.; Lu, M.Z.; Groover, A.T. Brassinosteroid regulation of wood formation in poplar. New Phytol. 2020, 225, 1516–1530. [Google Scholar] [CrossRef]
- Du, S.; Yamamoto, F. An overview of the biology of reaction wood formation. J. Integr. Plant Biol. 2007, 49, 131–143. [Google Scholar] [CrossRef]
- Dettmer, J.; Elo, A.; Helariutta, Y. Hormone interactions during vascular development. Plant Mol. Biol. 2009, 69, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Liu, K.; Liu, S.; Chen, H.; Zhou, L.; Liu, Y. Effect of exogenous IAA on tension wood formation by facilitating polar auxin transport and cellulose biosynthesis in hybrid poplar (Populus deltoids × Populus nigra) wood. Holzforschung 2017, 71, 179–188. [Google Scholar] [CrossRef]
- Xu, C.; Shen, Y.; He, F.; Fu, X.; Yu, H.; Lu, W.; Li, Y.; Li, C.; Fan, D.; Wang, H.C.; et al. Auxin-mediated Aux/IAA-ARF-HB signaling cascade regulates secondary xylem development in Populus. New Phytol. 2019, 222, 752–767. [Google Scholar] [CrossRef]
- Björklund, S.; Antti, H.; Uddestrand, I.; Moritz, T.; Sundberg, B. Cross-talk between gibberellin and auxin in development of Populus wood: Gibberellin stimulates polar auxin transport and has a common transcriptome with auxin. Plant J. 2007, 52, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Gerttula, S.; Zinkgraf, M.; Muday, G.K.; Lewis, D.R.; Ibatullin, F.M.; Brumer, H.; Hart, F.; Mansfield, S.D.; Filkov, V.; Groover, A. Transcriptional and hormonal regulation of gravitropism of woody stems in Populus. Plant Cell 2015, 27, 2800–2813. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Han, S.; Lee, H.Y.; Jeong, B.; Heo, T.Y.; Hyun, T.K.; Kim, K.; Je, B.; Lee, H.; Shim, D.; et al. Brassinosteroids facilitate xylem differentiation and wood formation in tomato. Planta 2019, 249, 1391–1403. [Google Scholar] [CrossRef]
- Ibañes, M.; Fàbregas, N.; Chory, J.; Caño-Delgado, A.I. Brassinosteroid signaling and auxin transport are required to establish the periodic pattern of Arabidopsis shoot vascular bundles. Proc. Natl. Acad. Sci. USA 2009, 106, 13630–13635. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Lv, B.; Ding, T.; Bai, M.; Ding, Z. Auxin-BR interaction regulates plant growth and development. Front. Plant Sci. 2018, 8, 2256. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Yu, M.; Zhu, S.; Zhou, L.; Liu, S. Effects of exogenous 24-epibrassinolide and brassinazole on negative gravitropism and tension wood formation in hybrid poplar (Populus deltoids × Populus nigra). Planta 2019, 249, 1449–1463. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Yokota, T. Biosynthesis and metabolism of brassinosteroids. Annu. Rev. Plant Biol. 2003, 54, 137–164. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Seto, H.; Fujioka, S.; Yoshida, S.; Chory, J. BRI1 is a critical component of a plasma-membrane receptor for plant steroids. Nature 2001, 410, 380–383. [Google Scholar] [CrossRef]
- Wang, X.; Chory, J. Brassinosteroids regulate dissociation of BKI1, a negative regulator of BRI1 signaling, from the plasma membrane. Science 2006, 313, 1118–1122. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell. 2020, 32, 295–318. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wang, Z.Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef] [Green Version]
- He, J.X.; Gendron, J.M.; Sun, Y.; Gampala, S.S.L.; Gendron, N.; Sun, C.Q.; Wang, Z.Y. BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science 2005, 307, 1634–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Cui, D.; Ye, Z.H. Secondary cell wall biosynthesis. New Phytol. 2019, 221, 1703–1723. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Han, X.; Yang, Z.; Xu, M.; Feng, Y.; Chen, Y.; Xu, L. Integrative analysis of wood biomass and developing xylem transcriptome provide insights into mechanisms of lignin biosynthesis in wood formation of Pinus massoniana. Int. J. Biol. Macromol. 2020, 163, 1926–1937. [Google Scholar] [CrossRef]
- Ohtani, M.; Demura, T. The quest for transcriptional hubs of lignin biosynthesis: Beyond the NAC-MYB-gene regulatory network model. Curr. Opin. Biotechnol. 2019, 56, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Guillaumie, S.; Mzid, R.; Méchin, V.; Léon, C.; Hichri, I.; Destrac-Irvine, A.; Trossat-Magnin, C.; Delrot, S.; Lauvergeat, V. The grapevine transcription factor WRKY2 influences the lignin pathway and xylem development in tobacco. Plant Mol. Biol. 2010, 72, 215–234. [Google Scholar] [CrossRef]
- Ohashi-Ito, K.; Matsukawa, M.; Fukuda, H. An atypical bHLH transcription factor regulated early xylem development downstream of auxin. Plant Cell Physiol. 2013, 54, 398–405. [Google Scholar] [CrossRef] [Green Version]
- Wessels, B.; Seyfferth, C.; Escamez, S.; Vain, T.; Antos, K.; Vahala, J.; Delhomme, N.; Kangasjärvi, J.; Eder, M.; Felten, J.; et al. An AP2/ERF transcription factor ERF139 coordinates xylem cell expansion and secondary cell wall deposition. New Phytol. 2019, 224, 1585–1599. [Google Scholar] [CrossRef]
- Yamamoto, R.; Fujioka, S.; Demura, T.; Takatsuto, S.; Yoshida, S.; Fukuda, H. Brassinosteroid levels increase drastically prior to morphogenesis of tracheary elements. Plant Physiol. 2001, 125, 556–563. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Wang, C.; Liu, Y.; He, G.; Ma, N.; Chai, G.; Li, S.; Xu, H.; Zhou, G. Brassinosteroid signaling converges with auxin-mediated C3H17 to regulate xylem formation in Populus. Front. Plant Sci. 2020, 11, 586014. [Google Scholar] [CrossRef]
- Xu, W.; Purugganan, M.M.; Polisensky, D.H.; Antosiewicz, D.M.; Fry, S.C.; Braam, J. Arabidopsis TCH4 regulated by hormones and the environment, encodes a xyloglucan endotransglycosylase. Plant Cell 1995, 7, 1555–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Bao, F.; Li, J. Promotive effect of brassinosteroids on cell division involves a distinct CycD3-induction pathway. Plant J. 2000, 24, 693–701. [Google Scholar] [CrossRef]
- Nemhauser, J.L.; Mockler, T.C.; Chory, J. Interdependency of brassinosteroid and auxin signaling in Arabidopsis. PLoS Biol. 2004, 2, e258. [Google Scholar] [CrossRef]
- Nakamura, A.; Nakajima, N.; Goda, H.; Shimada, Y.; Hayashi, K.; Nozaki, H.; Asami, T.; Yoshida, S.; Fujioka, S. Arabidopsis Aux/IAA genes are involved in brassinosteroid-mediated growth responses in a manner dependent on organ type. Plant J. 2006, 45, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Stortenbeker, N.; Bemer, M. The SAUR gene family: The plant’s toolbox for adaptation of growth and development. J. Exp. Bot. 2019, 70, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, H.; Xiao, Y.; Liu, D.; Gao, S.; Liu, L.; Yin, Y.; Jin, Y.; Qian, Q.; Chu, C. Brassinosteroid regulates cell elongation by modulating gibberelin metabolism in rice. Plant Cell 2014, 26, 3276–4393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unterholzner, S.J.; Rozhon, W.; Papacek, M.; Ciomas, J.; Lange, T.; Kugler, K.G.; Mayer, K.F.; Sieberer, T.; Poppenberger, B. Brassinosteroids are master regulators of gibberellin biosynthesis in Arabidopsis. Plant Cell 2015, 27, 2261–2272. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Chen, H.; Yang, H.; He, Y.; Tian, Z.; Li, J. A brassinosteroid responsive miRNA-target module regulates gibberellins biosynthesis and plant development. New Phytol. 2018, 220, 488–501. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Do, J.; Shin, S.J.; Choi, J.W.; Choi, Y.I.; Kim, W.; Kwon, M. Exogenously applied 24-epi brassinolide reduces lignifications and alters cell wall carbohydrate biosynthesis in the secondary xylem of Liriodendron tulipifera. Phytochemistry 2014, 101, 40–51. [Google Scholar] [CrossRef]
- Xie, L.; Yang, C.; Wang, X. Brassinosteroids can regulate cellulose biosynthesis by controlling the expression of CESA genes in Arabidopsis. J. Exp. Bot. 2011, 62, 4495–4506. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Mazarei, M.; Hisano, H.; Escamilla-Trevino, L.; Fu, C.; Pu, Y.; Rudis, M.R.; Tang, Y.; Xiao, X.; Jackson, L.; et al. A genomics approach to deciphering lignin biosynthesis in switchgrass. Plant Cell 2013, 25, 4342–4361. [Google Scholar] [CrossRef] [Green Version]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, R.A.; Barros, J. Lignin biosynthesis: Old roads revisited and new roads explored. Open Biol. 2019, 9, 190215. [Google Scholar] [CrossRef] [Green Version]
- Pu, Y.; Chen, F.; Ziebell, A.; Davison, B.H.; Ragauskas, A.J. NMR characterization of C3H and HCT down-regulated alfalfa lignin. BioEnergy Res. 2009, 2, 198. [Google Scholar] [CrossRef]
- Barros, J.; Escamilla-Trevino, L.; Song, L.; Ran, X.; Serrani-Yarce, J.C.; Palacios, M.D.; Engle, N.; Choudhury, F.K.; Tschaplinski, T.J.; Venables, B.J.; et al. 4-Coumarate 3-hydroxylase in the lignin biosynthesis pathway is a cytosolic ascorbate peroxidase. Nat. Commun. 2019, 10, 1994. [Google Scholar] [CrossRef] [Green Version]
- Keegstra, K. Plant cell walls. Plant Physiol. 2010, 154, 483–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyfferth, C.; Wessels, B.; Jokipii-Lukkari, S.; Sundberg, B.; Delhomme, N.; Felten, J.; Tuominen, H. Ethylene-related gene expression networks in wood formation. Front. Plant Sci. 2018, 9, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahala, J.; Felten, J.; Love, J.; Gorzsás, A.; Gerber, L.; Lamminmäki, A.; Kangasjärvi, J.; Sundberg, B. A genome-wide screen for ethylene-induced Ethylene Response Factors (ERFs) in hybrid aspen stem identifies ERF genes that modify stem growth and wood properties. New Phytol. 2013, 200, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Avci, U.; Nakashima, J.; Hahn, M.G.; Chen, F.; Dixon, R.A. Mutation of WRKY transcription factors initiates pith secondary wall formation and increases stem biomass in dicotyledonous plants. Proc. Natl. Acad. Sci. USA 2010, 107, 22338–22343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Tian, Z.; Yu, D. WRKY13 acts in stem development in Arabidopsis thaliana. Plant Sci. 2015, 236, 205–213. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Classification | Gene Name | Accession Number | Description | Fold Change Log2 (BR/Control) |
|---|---|---|---|---|
| Phytohormone biosynthesis | CPD | TRINITY_DN1557_c1_g2 | Cytochrome P450, oxidoreductase activity | −2.35 |
| CYP92A6 | TRINITY_DN59771_c0_g1 | Cytochrome P450, monooxygenase activity | −7.13 | |
| Phytohormone metabolism and signal transduction | GRP | TRINITY_DN7994_c0_g1 | Gibberellin-regulated protein, posttranslational modification, protein turnover, chaperones | 4.69 |
| DAO | TRINITY_DN9023_c0_g1 | Gibberellin 3-beta-dioxygenase activity | −2.81 | |
| LOB | TRINITY_DN142222_c0_g1 | Lateral organ boundaries (LOB) domain-containing protein, response to gibberellin | −3.73 | |
| APT-like | TRINITY_DN68002_c0_g2 | Regulation of auxin polar transport | 1.79 | |
| SAUR | TRINITY_DN8347_c0_g1 | Auxin responsive family-like protein | 2.21 | |
| 5NG4 | TRINITY_DN370_c0_g1 | Auxin-induced protein | −1.31 | |
| AUX/IAA | TRINITY_DN1260_c0_g1 | Auxin-responsive protein IAA7 | −1.19 |
| Functional Classification | Gene Name | Accession Number | Description | Fold Change Log2 (BR/Control) |
|---|---|---|---|---|
| Phenylpropanoid biosynthesis | HCT | TRINITY_DN10676_c0_g1 | Agmatine coumaroyltransferase | 2.07 |
| Tyrosine metabolism | COMT | TRINITY_DN48043_c1_g2 | Catechol O-methyltransferase A | 12.38 |
| Glycerophospholipid metabolism | CHK | TRINITY_DN1519_c0_g1 | Choline/ethanolamine kinase | 1.89 |
| Carbohydrate transport and metabolism | 3Beta-HSD | TRINITY_DN10086_c0_g1 | 3-beta hydroxysteroid dehydrogenase/isomerase | 1.47 |
| UDPGT | TRINITY_DN18407_c0_g2 | UDP-glucoronosyl and UDP-glucosyl transferase | 5.18 | |
| PIP | TRINITY_DN8514_c0_g1 | Probable aquaporin PIP2-6 | 3.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, F.; Zhou, Z.; Qin, H.; Tan, J.; Ding, G. Exogenous Brassinosteroid Facilitates Xylem Development in Pinus massoniana Seedlings. Int. J. Mol. Sci. 2021, 22, 7615. https://doi.org/10.3390/ijms22147615
Fan F, Zhou Z, Qin H, Tan J, Ding G. Exogenous Brassinosteroid Facilitates Xylem Development in Pinus massoniana Seedlings. International Journal of Molecular Sciences. 2021; 22(14):7615. https://doi.org/10.3390/ijms22147615
Chicago/Turabian StyleFan, Fuhua, Zijing Zhou, Huijuan Qin, Jianhui Tan, and Guijie Ding. 2021. "Exogenous Brassinosteroid Facilitates Xylem Development in Pinus massoniana Seedlings" International Journal of Molecular Sciences 22, no. 14: 7615. https://doi.org/10.3390/ijms22147615
APA StyleFan, F., Zhou, Z., Qin, H., Tan, J., & Ding, G. (2021). Exogenous Brassinosteroid Facilitates Xylem Development in Pinus massoniana Seedlings. International Journal of Molecular Sciences, 22(14), 7615. https://doi.org/10.3390/ijms22147615