
Linking Fibrotic Remodeling and Ultrastructural Alterations of Alveolar Epithelial Cells after Deletion of Nedd4-2

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
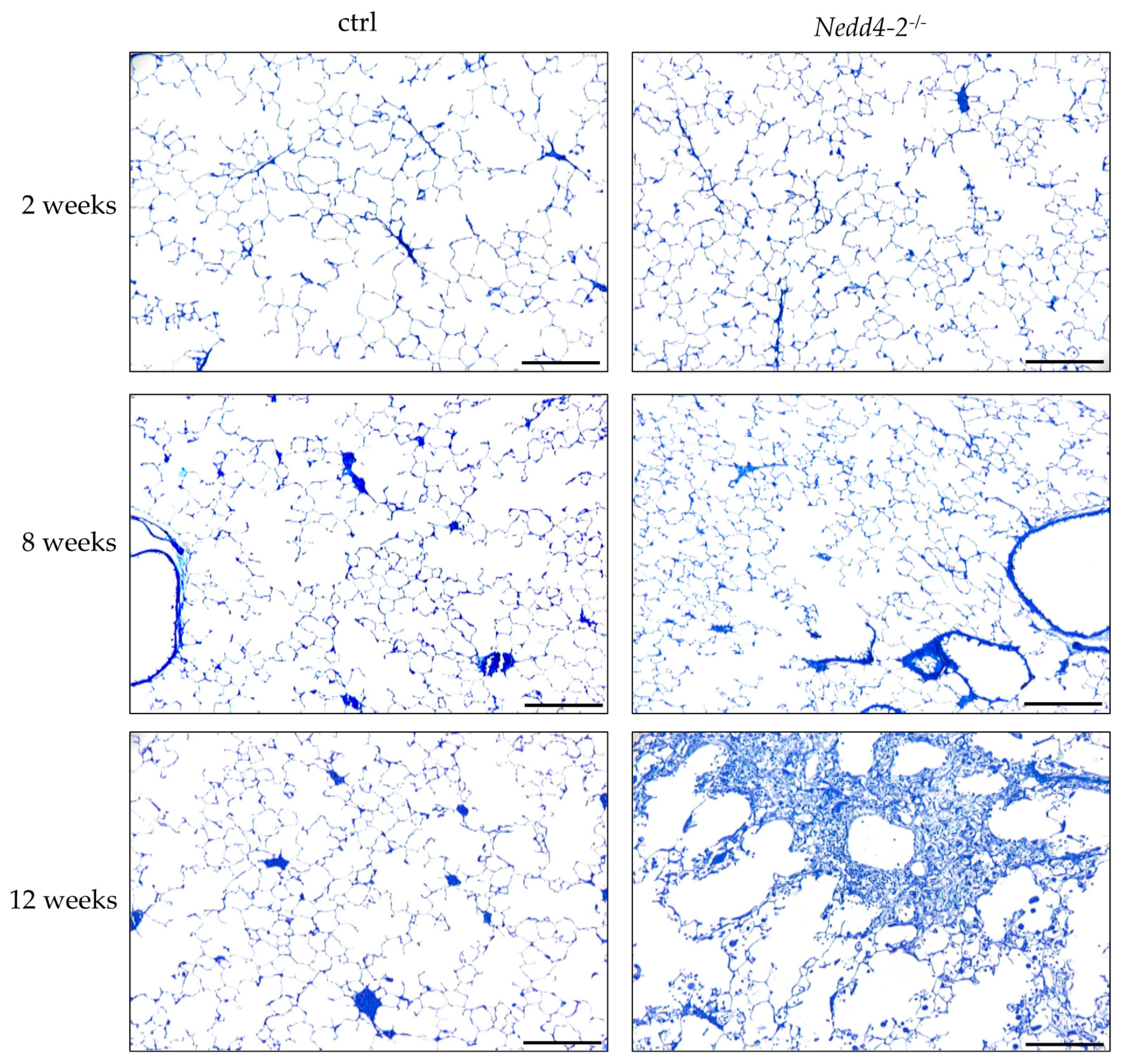
2.1. Progressive Fibrotic Remodeling in Nedd4-2-Deficient Lungs
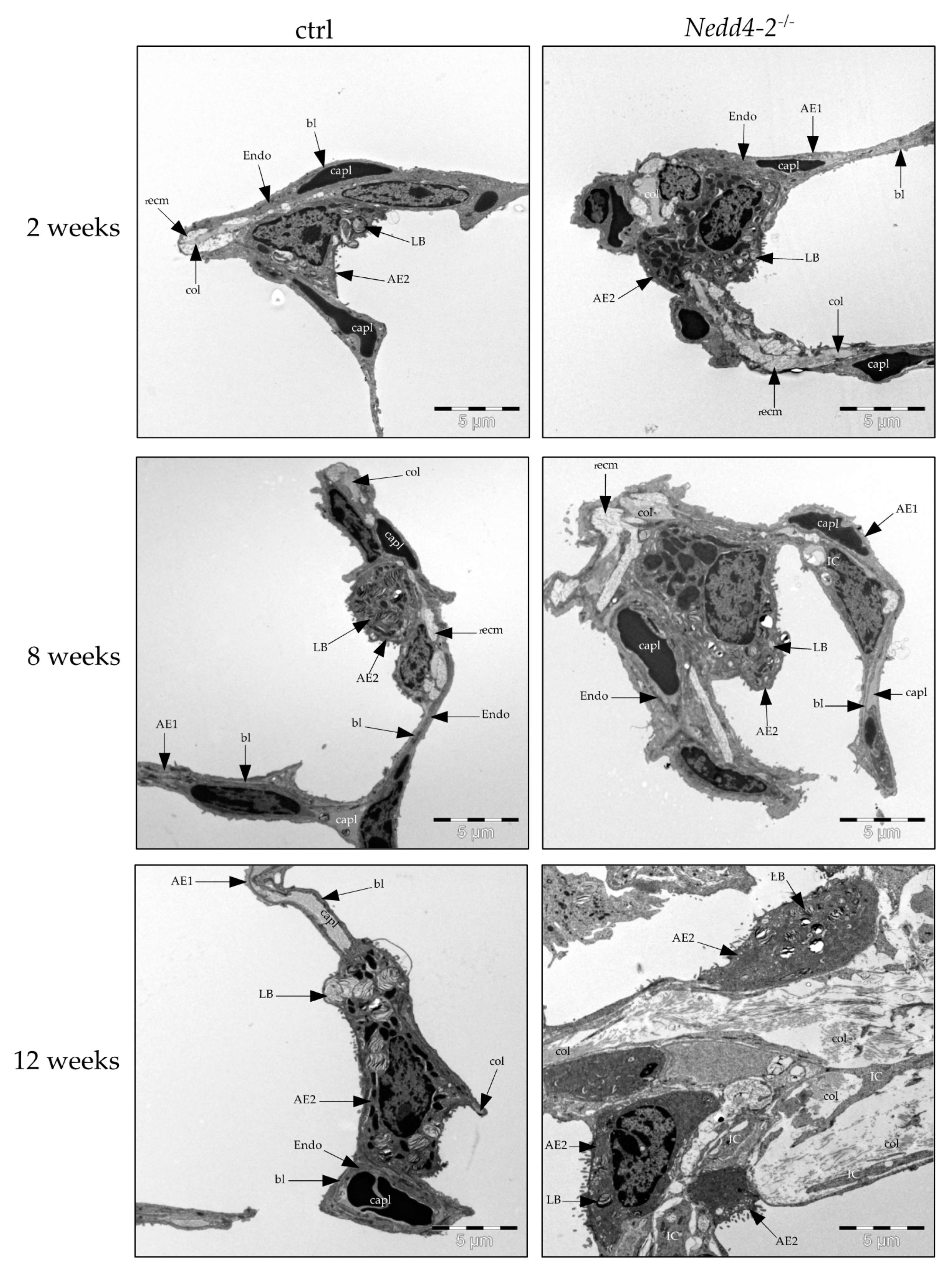
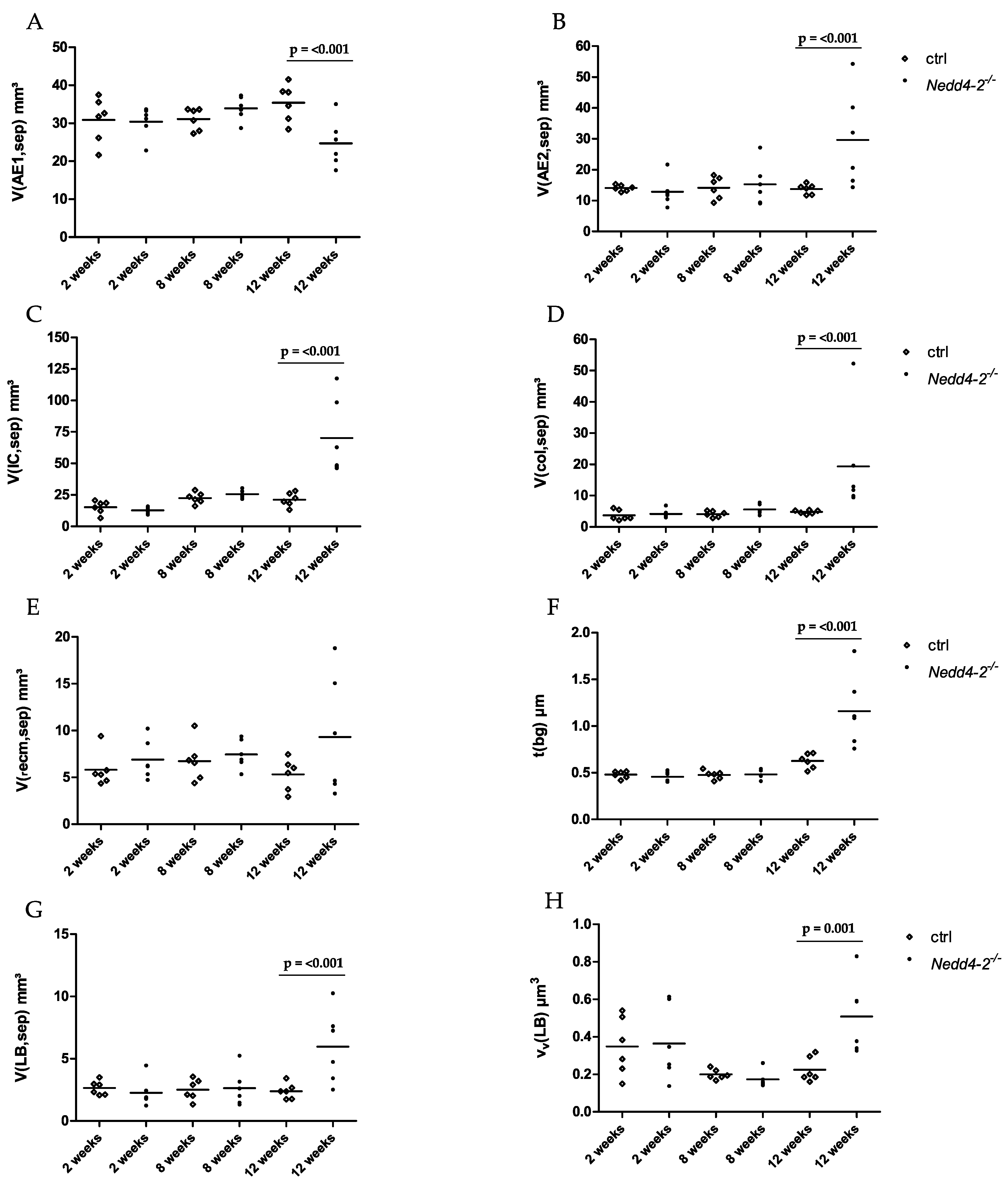
2.2. Composition of the Interalveolar Septa
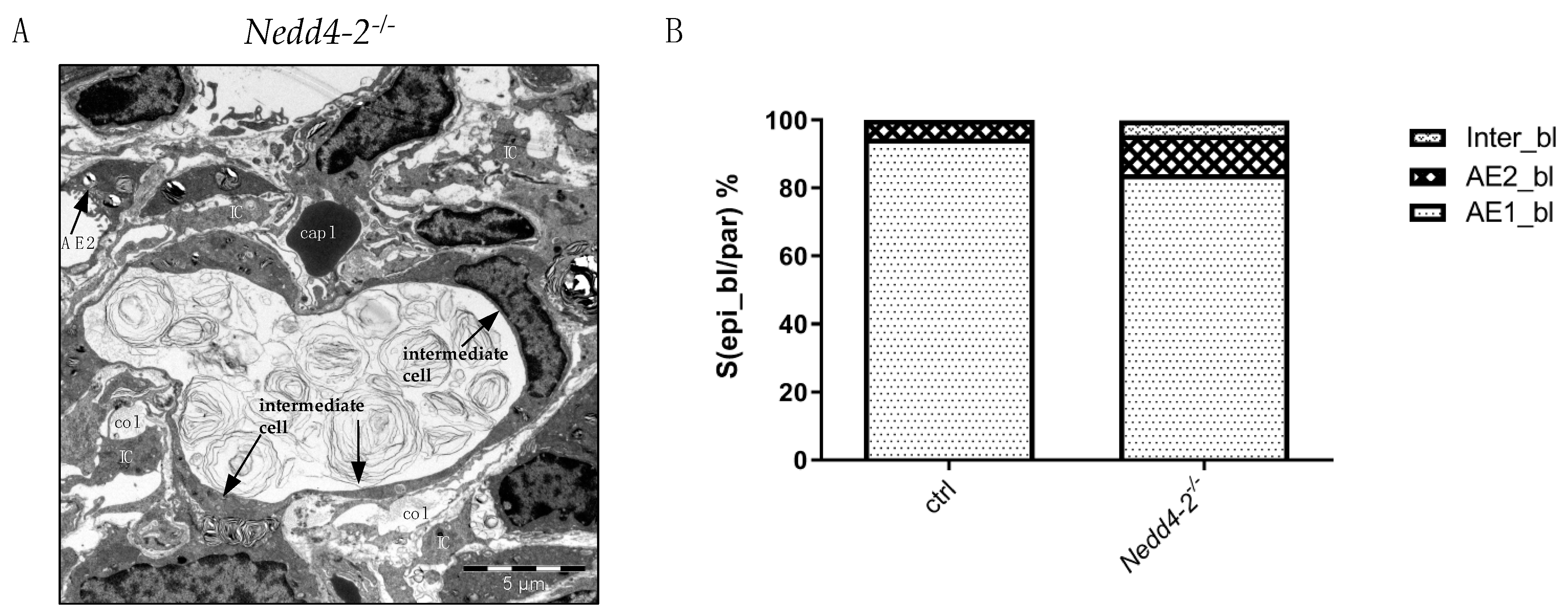
2.3. Alveolar Epithelial Cell Injury
2.4. Correlation Analyses
3. Discussion
4. Materials and Methods
4.1. Animal Model, Study Groups, Sampling and Embedding for Light and Electron Microscopy
4.2. Design-Based Stereology
4.3. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.F.; Flaherty, K.R.; Lasky, J.A.; et al. An official ATS/ERS/JRS/ALAT statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 2011, 183, 788–824. [Google Scholar] [CrossRef]
- Ley, B.; Collard, H.R.; King, T.E. Clinical course and prediction of survival in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2011, 183, 431–440. [Google Scholar] [CrossRef]
- Knudsen, L.; Ruppert, C.; Ochs, M. Tissue remodelling in pulmonary fibrosis. Cell Tissue Res. 2017, 367, 607–626. [Google Scholar] [CrossRef] [PubMed]
- Fingerlin, T.E.; Murphy, E.; Zhang, W.; Peljto, A.L.; Brown, K.K.; Steele, M.P.; Loyd, J.E.; Cosgrove, G.P.; Lynch, D.; Groshong, S.; et al. Genome-wide association study identifies multiple susceptibility loci for pulmonary fibrosis. Nat. Genet. 2013, 45, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Scharm, S.C.; Vogel-Claussen, J.; Schaefer-Prokop, C.; Dettmer, S.; Knudsen, L.; Jonigk, D.; Fuge, J.; Apel, R.M.; Welte, T.; Wacker, F.; et al. Quantification of dual-energy CT-derived functional parameters as potential imaging markers for progression of idiopathic pulmonary fibrosis. Eur. Radiol. 2021. [CrossRef]
- Knudsen, L.; Ochs, M. The micromechanics of lung alveoli: Structure and function of surfactant and tissue components. Histochem. Cell Biol. 2018, 150, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Albert, R.K.; Smith, B.; Perlman, C.E.; Schwartz, D.A. Is Progression of Pulmonary Fibrosis due to Ventilation-induced Lung Injury? Am. J. Respir. Crit. Care Med. 2019, 200, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Fehrenbach, H. Alveolar epithelial type II cell: Defender of the alveolus revisited. Respir. Res. 2001, 2, 33–46. [Google Scholar] [CrossRef]
- Selman, M.; Pardo, A. Role of epithelial cells in idiopathic pulmonary fibrosis: From innocent targets to serial killers. Proc. Am. Thorac. Soc. 2006, 3, 364–372. [Google Scholar] [CrossRef]
- Parimon, T.; Yao, C.; Stripp, B.R.; Noble, P.W.; Chen, P. Alveolar Epithelial Type II Cells as Drivers of Lung Fibrosis in Idiopathic Pulmonary Fibrosis. Int. J. Mol. Sci. 2020, 21, 2269. [Google Scholar] [CrossRef] [Green Version]
- Uhal, B.D.; Joshi, I.; Hughes, W.F.; Ramos, C.; Pardo, A.; Selman, M. Alveolar epithelial cell death adjacent to underlying myofibroblasts in advanced fibrotic human lung. Am. J. Physiol. 1998, 275, L1192–L1199. [Google Scholar] [CrossRef] [PubMed]
- Barbas-Filho, J.V.; Ferreira, M.A.; Sesso, A.; Kairalla, R.A.; Carvalho, C.R.; Capelozzi, V.L. Evidence of type II pneumocyte apoptosis in the pathogenesis of idiopathic pulmonary fibrosis (IFP)/usual interstitial pneumonia (UIP). J. Clin. Pathol. 2001, 54, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Haschek, W.M.; Witschi, H. Pulmonary fibrosis--a possible mechanism. Toxicol. Appl. Pharmacol. 1979, 51, 475–487. [Google Scholar] [CrossRef]
- Adamson, I.Y.; Young, L.; Bowden, D.H. Relationship of alveolar epithelial injury and repair to the induction of pulmonary fibrosis. Am. J. Pathol. 1988, 130, 377–383. [Google Scholar]
- Duerr, J.; Leitz, D.H.W.; Szczygiel, M.; Dvornikov, D.; Fraumann, S.G.; Kreutz, C.; Zadora, P.K.; Seyhan Agircan, A.; Konietzke, P.; Engelmann, T.A.; et al. Conditional deletion of Nedd4-2 in lung epithelial cells causes progressive pulmonary fibrosis in adult mice. Nat. Commun. 2020, 11, 2012. [Google Scholar] [CrossRef] [PubMed]
- Hancock, L.A.; Hennessy, C.E.; Solomon, G.M.; Dobrinskikh, E.; Estrella, A.; Hara, N.; Hill, D.B.; Kissner, W.J.; Markovetz, M.R.; Grove Villalon, D.E.; et al. Muc5b overexpression causes mucociliary dysfunction and enhances lung fibrosis in mice. Nat. Commun. 2018, 9, 5363. [Google Scholar] [CrossRef]
- Rotin, D.; Staub, O. Nedd4-2 and the regulation of epithelial sodium transport. Front. Physiol. 2012, 3, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mall, M.; Grubb, B.R.; Harkema, J.R.; O’Neal, W.K.; Boucher, R.C. Increased airway epithelial Na+ absorption produces cystic fibrosis-like lung disease in mice. Nat. Med. 2004, 10, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Mall, M.A.; Button, B.; Johannesson, B.; Zhou, Z.; Livraghi, A.; Caldwell, R.A.; Schubert, S.C.; Schultz, C.; O’Neal, W.K.; Pradervand, S.; et al. Airway surface liquid volume regulation determines different airway phenotypes in liddle compared with betaENaC-overexpressing mice. J. Biol. Chem. 2010, 285, 26945–26955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitz, D.H.; Duerr, J.; Mulugeta, S.; Agircan, A.S.; Zimmermann, S.; Kawabe, H.; Dalpke, A.H.; Beers, M.F.; Mall, M.A. Congenital deletion of Nedd4-2 in lung epithelial cells causes progressive alveolitis and pulmonary fibrosis in neonatal mice. Int. J. Mol. Sci. 2021, 22, 6146. [Google Scholar] [CrossRef]
- Gao, S.; Alarcón, C.; Sapkota, G.; Rahman, S.; Chen, P.Y.; Goerner, N.; Macias, M.J.; Erdjument-Bromage, H.; Tempst, P.; Massagué, J. Ubiquitin ligase Nedd4L targets activated Smad2/3 to limit TGF-beta signaling. Mol. Cell 2009, 36, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotorashvili, A.; Russo, S.J.; Mulugeta, S.; Guttentag, S.; Beers, M.F. Anterograde transport of surfactant protein C proprotein to distal processing compartments requires PPDY-mediated association with Nedd4 ubiquitin ligases. J. Biol. Chem. 2009, 284, 16667–16678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conkright, J.J.; Apsley, K.S.; Martin, E.P.; Ridsdale, R.; Rice, W.R.; Na, C.L.; Yang, B.; Weaver, T.E. Nedd4-2-mediated ubiquitination facilitates processing of surfactant protein-C. Am. J. Respir. Cell Mol. Biol. 2010, 42, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Bonniaud, P.; Fabre, A.; Frossard, N.; Guignabert, C.; Inman, M.; Kuebler, W.M.; Maes, T.; Shi, W.; Stampfli, M.; Uhlig, S.; et al. Optimising experimental research in respiratory diseases: An ERS statement. Eur. Respir. J. 2018, 51. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, R.G.; Moore, B.B.; Chambers, R.C.; Eickelberg, O.; Königshoff, M.; Kolb, M.; Laurent, G.J.; Nanthakumar, C.B.; Olman, M.A.; Pardo, A.; et al. An Official American Thoracic Society Workshop Report: Use of Animal Models for the Preclinical Assessment of Potential Therapies for Pulmonary Fibrosis. Am. J. Respir. Cell Mol. Biol. 2017, 56, 667–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, Y.; Yang, I.V.; Walts, A.D.; Watson, A.M.; Helling, B.A.; Fletcher, A.A.; Lara, A.R.; Schwarz, M.I.; Evans, C.M.; Schwartz, D.A. MUC5B Promoter Variant rs35705950 Affects MUC5B Expression in the Distal Airways in Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2016, 193, 464–466. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.V.; Coldren, C.D.; Leach, S.M.; Seibold, M.A.; Murphy, E.; Lin, J.; Rosen, R.; Neidermyer, A.J.; McKean, D.F.; Groshong, S.D.; et al. Expression of cilium-associated genes defines novel molecular subtypes of idiopathic pulmonary fibrosis. Thorax 2013, 68, 1114–1121. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.M.; Fingerlin, T.E.; Schwarz, M.I.; Lynch, D.; Kurche, J.; Warg, L.; Yang, I.V.; Schwartz, D.A. Idiopathic Pulmonary Fibrosis: A Genetic Disease That Involves Mucociliary Dysfunction of the Peripheral Airways. Physiol. Rev. 2016, 96, 1567–1591. [Google Scholar] [CrossRef]
- Hsia, C.; Hyde, D.; Ochs, M.; Weibel, E. An official research policy statement of the American Thoracic Society/European Respiratory Society: Standards for quantitative assessment of lung structure. Am. J. Respir. Crit. Care Med. 2010, 181, 394–418. [Google Scholar] [CrossRef] [Green Version]
- Perez-Gil, J.; Weaver, T. Pulmonary surfactant pathophysiology: Current models and open questions. Physiology 2010, 25, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krischer, J.M.; Albert, K.; Pfaffenroth, A.; Lopez-Rodriguez, E.; Ruppert, C.; Smith, B.J.; Knudsen, L. Mechanical ventilation-induced alterations of intracellular surfactant pool and blood-gas barrier in healthy and pre-injured lungs. Histochem. Cell Biol. 2021, 155, 183–202. [Google Scholar] [CrossRef]
- Beike, L.; Wrede, C.; Hegermann, J.; Lopez-Rodriguez, E.; Kloth, C.; Gauldie, J.; Kolb, M.; Maus, U.A.; Ochs, M.; Knudsen, L. Surfactant dysfunction and alveolar collapse are linked with fibrotic septal wall remodeling in the TGF-β1-induced mouse model of pulmonary fibrosis. Lab. Investig. 2019, 99, 830–852. [Google Scholar] [CrossRef]
- Lopez-Rodriguez, E.; Boden, C.; Echaide, M.; Perez-Gil, J.; Kolb, M.; Gauldie, J.; Maus, U.A.; Ochs, M.; Knudsen, L. Surfactant dysfunction during overexpression of TGF-beta 1 precedes profibrotic lung remodeling in vivo. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L1260–L1271. [Google Scholar] [CrossRef] [PubMed]
- Birkelbach, B.; Lutz, D.; Ruppert, C.; Henneke, I.; Lopez-Rodriguez, E.; Guenther, A.; Ochs, M.; Mahavadi, P.; Knudsen, L. Linking progression of fibrotic lung remodeling and ultrastructural alterations of alveolar epithelial type II cells in the amiodarone mouse model. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L63–L75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozarska, A.; Rodríguez-Castillo, J.A.; Surate Solaligue, D.E.; Ntokou, A.; Rath, P.; Mižíková, I.; Madurga, A.; Mayer, K.; Vadász, I.; Herold, S.; et al. Stereological monitoring of mouse lung alveolarization from the early post-natal period to adulthood. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, D.; Gazdhar, A.; Lopez-Rodriguez, E.; Ruppert, C.; Mahavadi, P.; Gunther, A.; Klepetko, W.; Bates, J.H.; Smith, B.; Geiser, T.; et al. Alveolar Derecruitment and Collapse Induration as Crucial Mechanisms in Lung Injury and Fibrosis. Am. J. Respir. Cell Mol. Biol. 2015, 52, 232–243. [Google Scholar] [CrossRef]
- Ochs, M.; Timm, S.; Elezkurtaj, S.; Horst, D.; Meinhardt, J.; Heppner, F.L.; Weber-Carstens, S.; Hocke, A.C.; Witzenrath, M. Collapse induration of alveoli is an ultrastructural finding in a COVID-19 patient. Eur. Respir. J. 2021, 57. [Google Scholar] [CrossRef] [PubMed]
- Katzenstein, A. Pathogenesis of “fibrosis” in interstitial pneumonia: An electron microscopic study. Hum. Pathol. 1985, 16, 1015–1024. [Google Scholar] [CrossRef]
- Myers, J.; Katzenstein, A. Epithelial necrosis and alveolar collapse in the pathogenesis of usual interstitial pneumonia. Chest 1988, 94, 1309–1311. [Google Scholar] [CrossRef]
- Coxson, H.O.; Hogg, J.C.; Mayo, J.R.; Behzad, H.; Whittall, K.P.; Schwartz, D.A.; Hartley, P.G.; Galvin, J.R.; Wilson, J.S.; Hunninghake, G.W. Quantification of idiopathic pulmonary fibrosis using computed tomography and histology. Am. J. Respir. Crit. Care Med. 1997, 155, 1649–1656. [Google Scholar] [CrossRef]
- Jansing, N.L.; McClendon, J.; Henson, P.M.; Tuder, R.M.; Hyde, D.M.; Zemans, R.L. Unbiased Quantitation of Alveolar Type II to Alveolar Type I Cell Transdifferentiation during Repair after Lung Injury in Mice. Am. J. Respir. Cell Mol. Biol. 2017, 57, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.J.; Brownfield, D.G.; Krasnow, M.A. Alveolar progenitor and stem cells in lung development, renewal and cancer. Nature 2014, 507, 190–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ware, L.B.; Matthay, M.A. Keratinocyte and hepatocyte growth factors in the lung: Roles in lung development, inflammation, and repair. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, L924–L940. [Google Scholar] [CrossRef] [Green Version]
- Bachofen, M.; Weibel, E.R. Structural alterations of lung parenchyma in the adult respiratory distress syndrome. Clin. Chest. Med. 1982, 3, 35–56. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Cronce, M.J.; Rackley, C.R.; Bowie, E.J.; Keene, D.R.; Stripp, B.R.; Randell, S.H.; Noble, P.W.; Hogan, B.L. Type 2 alveolar cells are stem cells in adult lung. J. Clin. Investig. 2013, 123, 3025–3036. [Google Scholar] [CrossRef]
- Rock, J.R.; Barkauskas, C.E.; Cronce, M.J.; Xue, Y.; Harris, J.R.; Liang, J.; Noble, P.W.; Hogan, B.L. Multiple stromal populations contribute to pulmonary fibrosis without evidence for epithelial to mesenchymal transition. Proc. Natl. Acad. Sci. USA 2011, 108, E1475–E1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Zhang, Y.; Xie, T.; Liu, N.; Chen, H.; Geng, Y.; Kurkciyan, A.; Mena, J.M.; Stripp, B.R.; Jiang, D.; et al. Hyaluronan and TLR4 promote surfactant-protein-C-positive alveolar progenitor cell renewal and prevent severe pulmonary fibrosis in mice. Nat. Med. 2016. [Google Scholar] [CrossRef]
- Aspal, M.; Zemans, R.L. Mechanisms of ATII-to-ATI Cell Differentiation during Lung Regeneration. Int. J. Mol. Sci. 2020, 21, 3188. [Google Scholar] [CrossRef]
- Riemondy, K.A.; Jansing, N.L.; Jiang, P.; Redente, E.F.; Gillen, A.E.; Fu, R.; Miller, A.J.; Spence, J.R.; Gerber, A.N.; Hesselberth, J.R.; et al. Single cell RNA sequencing identifies TGFβ as a key regenerative cue following LPS-induced lung injury. JCI Insight 2019, 5. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Gil de Rubio, R.; Hrycaj, S.M.; Gurczynski, S.J.; Riemondy, K.A.; Moore, B.B.; Omary, M.B.; Ridge, K.M.; Zemans, R.L. Ineffectual Type 2-to-Type 1 Alveolar Epithelial Cell Differentiation in Idiopathic Pulmonary Fibrosis: Persistence of the KRT8. Am. J. Respir. Crit. Care Med. 2020, 201, 1443–1447. [Google Scholar] [CrossRef]
- Mahavadi, P.; Henneke, I.; Ruppert, C.; Knudsen, L.; Venkatesan, S.; Liebisch, G.; Chambers, R.C.; Ochs, M.; Schmitz, G.; Vancheri, C.; et al. Altered surfactant homeostasis and alveolar epithelial cell stress in amiodarone-induced lung fibrosis. Toxicol. Sci. 2014, 142, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Mahavadi, P.; Korfei, M.; Henneke, I.; Liebisch, G.; Schmitz, G.; Gochuico, B.R.; Markart, P.; Bellusci, S.; Seeger, W.; Ruppert, C.; et al. Epithelial stress and apoptosis underlie Hermansky-Pudlak syndrome-associated interstitial pneumonia. Am. J. Respir. Crit. Care Med. 2010, 182, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, L.; Lopez-Rodriguez, E.; Berndt, L.; Steffen, L.; Ruppert, C.; Bates, J.H.T.; Ochs, M.; Smith, B.J. Alveolar micromechanics in bleomycin-induced lung injury. Am. J. Respir. Cell Mol. Biol. 2018, 59, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Kawanami, O.; Ferrans, V.J.; Crystal, R.G. Structure of alveolar epithelial cells in patients with fibrotic lung disorders. Lab. Investig. 1982, 46, 39–53. [Google Scholar] [PubMed]
- Seibold, M.A.; Smith, R.W.; Urbanek, C.; Groshong, S.D.; Cosgrove, G.P.; Brown, K.K.; Schwarz, M.I.; Schwartz, D.A.; Reynolds, S.D. The idiopathic pulmonary fibrosis honeycomb cyst contains a mucocilary pseudostratified epithelium. PLoS ONE 2013, 8, e58658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou-Suckow, Z.; Duerr, J.; Hagner, M.; Agrawal, R.; Mall, M.A. Airway mucus, inflammation and remodeling: Emerging links in the pathogenesis of chronic lung diseases. Cell Tissue Res. 2017, 367, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ribeiro, C.M.P.; Sun, L.; Okuda, K.; Kato, T.; Gilmore, R.C.; Martino, M.B.; Dang, H.; Abzhanova, A.; Lin, J.M.; et al. XBP1S Regulates MUC5B in a Promoter Variant-Dependent Pathway in Idiopathic Pulmonary Fibrosis Airway Epithelia. Am. J. Respir. Crit. Care Med. 2019, 200, 220–234. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Perlman, C.E. Tracheal acid or surfactant instillation raises alveolar surface tension. J. Appl. Physiol. 2018, 125, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Perlman, C.E. Sulforhodamine B and exogenous surfactant effects on alveolar surface tension under acute respiratory distress syndrome conditions. J. Appl. Physiol. 2020, 129, 1505–1513. [Google Scholar] [CrossRef]
- Perlman, C.E. The Contribution of Surface Tension-Dependent Alveolar Septal Stress Concentrations to Ventilation-Induced Lung Injury in the Acute Respiratory Distress Syndrome. Front. Physiol. 2020, 11, 388. [Google Scholar] [CrossRef]
- Rühl, N.; Lopez-Rodriguez, E.; Albert, K.; Smith, B.J.; Weaver, T.E.; Ochs, M.; Knudsen, L. Surfactant Protein B Deficiency Induced High Surface Tension: Relationship between Alveolar Micromechanics, Alveolar Fluid Properties and Alveolar Epithelial Cell Injury. Int. J. Mol. Sci. 2019, 20, 4243. [Google Scholar] [CrossRef] [Green Version]
- Gil, J.; Bachofen, H.; Gehr, P.; Weibel, E. Alveolar volume-surface area relation in air- and saline-filled lungs fixed by vascular perfusion. J. Appl. Physiol. 1979, 47, 990–1001. [Google Scholar] [CrossRef]
- Kimura, T.; Kawabe, H.; Jiang, C.; Zhang, W.; Xiang, Y.-Y.; Lu, C.; Salter, M.W.; Brose, N.; Lu, W.-Y.; Rotin, D. Deletion of the ubiquitin ligase Nedd4L in lung epithelia causes cystic fibrosis-like disease. Proc. Natl. Acad. Sci. USA 2011, 108, 3216–3221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duerr, J.; Gruner, M.; Schubert, S.C.; Haberkorn, U.; Bujard, H.; Mall, M.A. Use of a New-Generation Reverse Tetracycline Transactivator System for Quantitative Control of Conditional Gene Expression in the Murine Lung. Am. J. Respir. Cell Mol. Biol. 2011, 44, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Baron, U.; Freundlieb, S.; Gossen, M.; Bujard, H. Co-regulation of two gene activities by tetracycline via a bidirectional promoter. Nucleic Acids Res. 1995, 23, 3605–3606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenig, K.; Schwenk, F.; Rajewsky, K.; Hermann, B. Stringent doxycycline dependent control of CRE recombinase in vivo. Nucleic Acids Res. 2002, 30, e134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherle, W. A simple method for volumetry of organs in quantitative stereology. Mikroskopie 1970, 26, 57–60. [Google Scholar]
- Tschanz, S.; Schneider, J.P.; Knudsen, L. Design-based stereology: Planning, volumetry and sampling are crucial steps for a successful study. Ann. Anat. 2014, 196, 3–11. [Google Scholar] [CrossRef]
- Schneider, J.P.; Ochs, M. Alterations of mouse lung tissue dimensions during processing for morphometry: A comparison of methods. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L341–L350. [Google Scholar] [CrossRef] [Green Version]
- Mühlfeld, C.; Knudsen, L.; Ochs, M. Stereology and morphometry of lung tissue. Methods Mol. Biol. 2013, 931, 367–390. [Google Scholar] [CrossRef]
- Braendgaard, H.; Gundersen, H.J. The impact of recent stereological advances on quantitative studies of the nervous system. J. Neurosci. Methods 1986, 18, 39–78. [Google Scholar] [CrossRef]
- Ochs, M.; Mühlfeld, C. Quantitative microscopy of the lung: A problem-based approach. Part 1: Basic principles of lung stereology. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L15–L22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlfeld, C.; Ochs, M. Quantitative microscopy of the lung: A problem-based approach. Part 2: Stereological parameters and study designs in various diseases of the respiratory tract. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L205–L221. [Google Scholar] [CrossRef] [Green Version]
- Tschanz, S.A.; Burri, P.H.; Weibel, E.R. A simple tool for stereological assessment of digital images: The STEPanizer. J. Microsc. 2011, 243, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J.; Jensen, E.B. Stereological estimation of the volume-weighted mean volume of arbitrary particles observed on random sections. J. Microsc. 1985, 138, 127–142. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | 2 Weeks | 8 Weeks | 12 Weeks | Two-Way ANOVA | |||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | ctrl | Nedd4-2−/− | ctrl | Nedd4-2−/− | ctrl | Nedd4-2−/− | G | T | Inter |
| Females/males | 4/2 | 3/3 | 3/3 | 3/3 | 4/2 | 3/3 | |||
| V(lung) cm³ | 0.59 (0.03) | 0.60 (0.05) | 0.64 (0.04) | 0.61 (0.05) | 0.80 (0.03) | 0.85 (0.06) | n.s. | <0.001 | 0.028 |
| V(par,lung) cm³ | 0.51 (0.04) | 0.51 (0.04) | 0.54 (0.03) | 0.51 (0.04) | 0.65 (0.03) | 0.68 (0.07) | n.s. | <0.001 | n.s. |
| V(sep,par) cm³ | 0.13 (0.02) | 0.12 (0.01) | 0.13 (0.01) | 0.16 (0.01) | 0.16 (0.02) | 0.22 (0.05) | 0.001 | <0.001 | 0.011 |
| V(air,par) cm³ | 0.38 (0.05) | 0.39 (0.03) | 0.40 (0.02) | 0.36 (0.04) | 0.49 (0.02) | 0.40 (0.09) | 0.016 | 0.004 | n.s. |
| V(cons,lung) cm³ | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) | 0.05 (0.03) | ––– | ––– | <0.001 |
| Group | 2 Weeks | 8 Weeks | 12 Weeks | Two-Way ANOVA | |||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | ctrl | Nedd4-2−/− | ctrl | Nedd4-2−/− | ctrl | Nedd4-2−/− | G | T | Inter |
| V(AE1,sep) mm³ | 30.88 (5.96) | 30.43 (4.02) | 31.16 (2.89) | 33.93 (3.15) | 35.40 (4.92) | 24.73 (6.24) | n.s. | 0.004 | n.s. |
| V(AE2,sep) mm³ | 14.07 (0.98) | 12.85 (4.71) | 14.19 (3.60) | 15.29 (6.74) | 13.75 (1.62) | 29.64 (15.59) | n.s. | 0.014 | 0.020 |
| V(Endo,sep) mm³ | 36.78 (10.80) | 34.80 (6.63) | 34.50 (4.83) | 46.72 (3.35) | 42.61 (10.07) | 35.88 (8.54) | n.s. | 0.016 | n.s. |
| V(capl,sep) mm³ | 21.49 (2.50) | 23.05 (5.61) | 20.72 (3.92) | 24.18 (5.21) | 35.42 (5.74) | 26.94 (6.98) | n.s. | 0.018 | <0.001 |
| V(recm,sep) mm³ | 5.81 (1.84) | 6.89 (2.10) | 6.75 (2.15) | 7.46 (1.53) | 5.33 (1.70) | 9.30 (6.43) | n.s. | n.s. | n.s. |
| V(col,sep) mm³ | 3.68 (1.64) | 4.13 (1.42) | 4.10 (0.98) | 5.57 (1.59) | 4.76 (0.60) | 19.33 (16.56) | <0.001 | <0.001 | 0.004 |
| V(IC,sep) mm³ | 15.29 (5.14) | 12.78 (2.67) | 22.58 (4.42) | 25.67 (3.11) | 21.35 (5.44) | 70.19 (30.47) | <0.001 | <0.001 | <0.001 |
| V(LB,sep) mm³ | 2.65 (0.57) | 2.27 (1.14) | 2.52 (0.84) | 2.63 (1.45) | 2.40 (0.63) | 5.97 (2.92) | 0.034 | 0.013 | 0.005 |
| νV(LB) µm3 | 0.35 (0.16) | 0.36 (0.20) | 0.20 (0.03) | 0.17 (0.04) | 0.22 (0.07) | 0.51 (0.20) | n.s. | 0.004 | 0.018 |
| S(epi_air,sep) cm² | 1322 (181) | 1305 (205) | 1386 (210) | 1622 (150) | 2232 (191) | 1518 (501) | <0.001 | n.s. | <0.001 |
| τ(bg) µm | 0.48 (0.04) | 0.46 (0.05) | 0.48 (0.05) | 0.48 (0.05) | 0.83 (0.10) | 1.29 (0.41) | 0.0027 | <0.001 | 0.002 |
| Group | 12 Weeks | ||
|---|---|---|---|
| Parameter | Ctrl | Nedd4-2−/− | t-Test |
| S(AE1_bl,sep) cm² | 1702 (156) | 1221 (370) | 0.015 |
| S(AE2_bl,sep) cm² | 98.0 (16.7) | 160.8 (60.5) | 0.034 |
| S(Inter_bl,sep) cm² | 0.00 (0.00) | 59.6 (16.8) | <0.01 |
| S(epi_bl,sep) cm² | 1799 (159] | 1442 (381) | 0.060 |
| S(AE1_air,par) cm² | 2085 (174] | 1302 (418) | <0.01 |
| S(AE2_air,par) cm² | 147 (27.1) | 216 (119) | n.s. |
| S(Inter_air,par) cm² | 0.00 (0.00) | 67.1 (25.1) | <0.01 |
| S(epi_air,par) cm² | 2232 (501) | 1518 [191) | <0.01 |
| S(endo,sep) cm² | 1707 (203) | 1963 (370) | n.s. |
| Stereological Parameters (Light Microscopy) | ||
|---|---|---|
| Parameter | Test System | Primary Magnification |
| Volume of parenchyma, V(par,lung) | Point counting | 5× |
| Volume of non-parenchyma, V(non-par,lung) | Point counting | 5× |
| Volume of (ductal and alveoli) airspace, V(air,par) | Point counting | 20× |
| Volume of septal wall, V(sep,par) | Point counting | 20× |
| Volume of consolidated, destroyed tissue areas, V(cons,lung) | Point counting | 20× |
| Stereological Parameters (Electron Microscopy) | ||
|---|---|---|
| Parameter | Test System | Primary Magnification |
| Volume of alveolar epithelial type 1 cells in septal walls, V(AE1,sep) | Point counting | 8900× |
| Volume of alveolar epithelial type 2 cells in septal walls, V(AE2,sep) | Point counting | 8900× |
| Volume of endothelial cells within septal walls, V(Endo,sep) | Point counting | 8900× |
| Volume of interstitial cells within septal walls, V(IC,sep) | Point counting | 8900× |
| Volume of collagen fibrils within septal walls, V(col,sep) | Point counting | 8900× |
| Volume of the extracellular matrix without collagen fibrils within septal walls, V(recm,sep) | Point counting | 8900× |
| Volume of capillary lumen within septal walls, V(capl,sep) | Point counting | 8900× |
| Surface area of endothelium within septal walls, S(endo,sep) | Point and intersection counting | 8900× |
| Surface area of of basal lamina covered by alveolar epithelium within septal walls, S(epi_bl,sep) (12 weeks) | Point and intersection counting | 8900× |
| Surface area of the basal lamina covered by AE1, S(AE1_bl,sep) (12 weeks) | Intersection counting | 8900× |
| Surface area of the basal lamina covered by AE2, S(AE2_bl,sep) (12 weeks) | Intersection counting | 8900× |
| Surface area of the basal lamina covered by intermediate cells, S(Inter_bl,sep) (12 weeks) | Intersection counting | 8900× |
| Surface area of alveolar epithelium covered by air within parenchyma, S(epi_air,par) | Intersection counting | 8900× |
| Surface area of the AE1 covered by air within parenchyma, S(AE1_air,par) (12 weeks) | Intersection counting | 8900× |
| Surface area of the AE2 covererd by air within parenchyma, S(AE2_air,par) (12weeks) | Intersection counting | 8900× |
| Surface area of the intermediate cells covered by air within parenchyma, S(Inter_air, par) (12 weeks) | Intersection counting | 8900× |
| Arithmetic mean thickness of the blood–gas barrier, τ(bg) | Volume-to-surface ratio | 8900× |
| Volume of lamellar bodies within the septal walls V(LB,sep) | Point counting | 7100× |
| Volume-weighted mean volume of the LB’s, νv(LB) | Point sampling intercept method | 7100× |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Engelmann, T.A.; Knudsen, L.; Leitz, D.H.W.; Duerr, J.; Beers, M.F.; Mall, M.A.; Ochs, M. Linking Fibrotic Remodeling and Ultrastructural Alterations of Alveolar Epithelial Cells after Deletion of Nedd4-2. Int. J. Mol. Sci. 2021, 22, 7607. https://doi.org/10.3390/ijms22147607
Engelmann TA, Knudsen L, Leitz DHW, Duerr J, Beers MF, Mall MA, Ochs M. Linking Fibrotic Remodeling and Ultrastructural Alterations of Alveolar Epithelial Cells after Deletion of Nedd4-2. International Journal of Molecular Sciences. 2021; 22(14):7607. https://doi.org/10.3390/ijms22147607
Chicago/Turabian StyleEngelmann, Theresa A., Lars Knudsen, Dominik H. W. Leitz, Julia Duerr, Michael F. Beers, Marcus A. Mall, and Matthias Ochs. 2021. "Linking Fibrotic Remodeling and Ultrastructural Alterations of Alveolar Epithelial Cells after Deletion of Nedd4-2" International Journal of Molecular Sciences 22, no. 14: 7607. https://doi.org/10.3390/ijms22147607
APA StyleEngelmann, T. A., Knudsen, L., Leitz, D. H. W., Duerr, J., Beers, M. F., Mall, M. A., & Ochs, M. (2021). Linking Fibrotic Remodeling and Ultrastructural Alterations of Alveolar Epithelial Cells after Deletion of Nedd4-2. International Journal of Molecular Sciences, 22(14), 7607. https://doi.org/10.3390/ijms22147607