
Evolution of Terpene Synthases in Orchidaceae

 and
and
Abstract
:1. Introduction
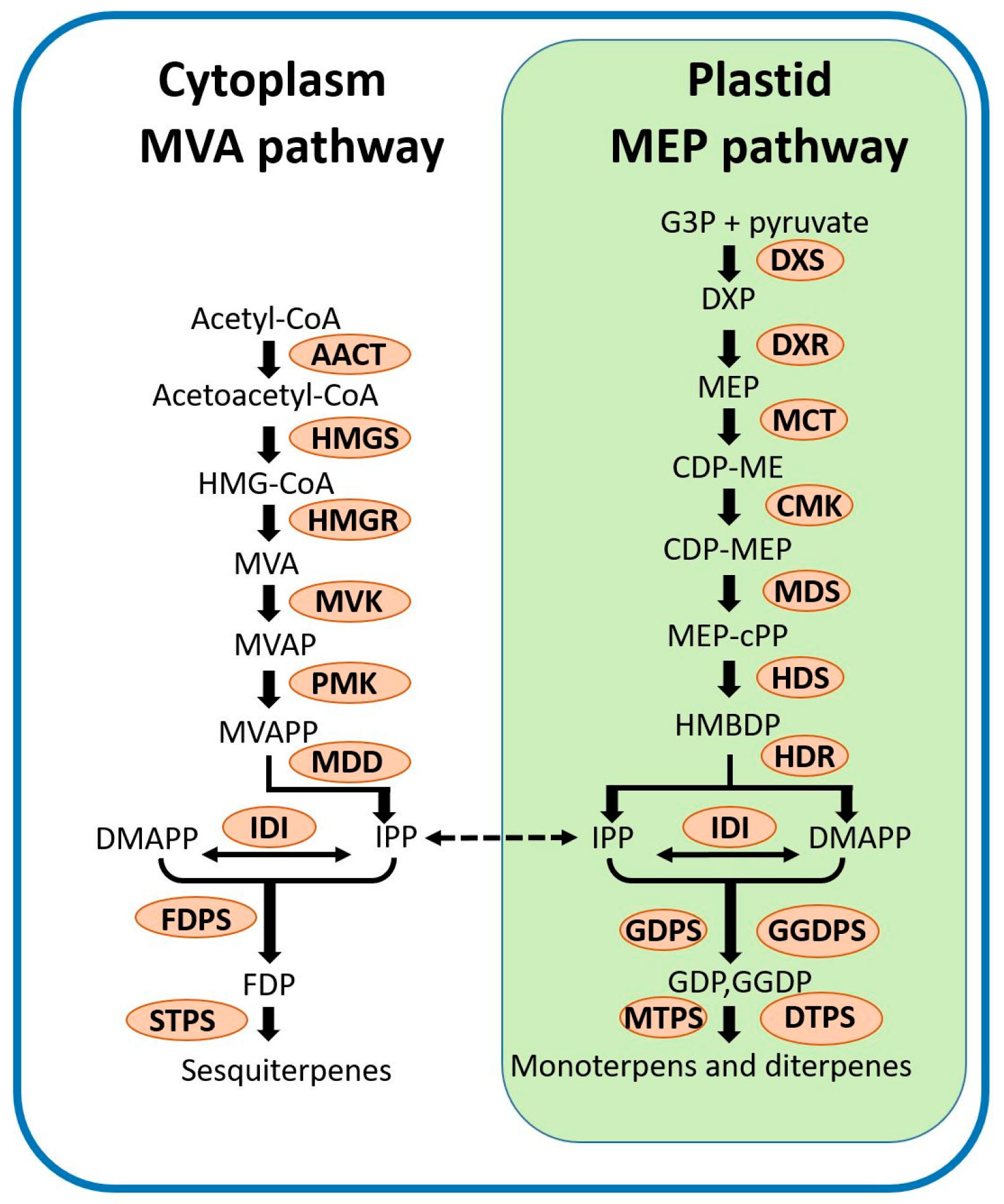
2. Terpenoids and Their Biosynthesis in Plants
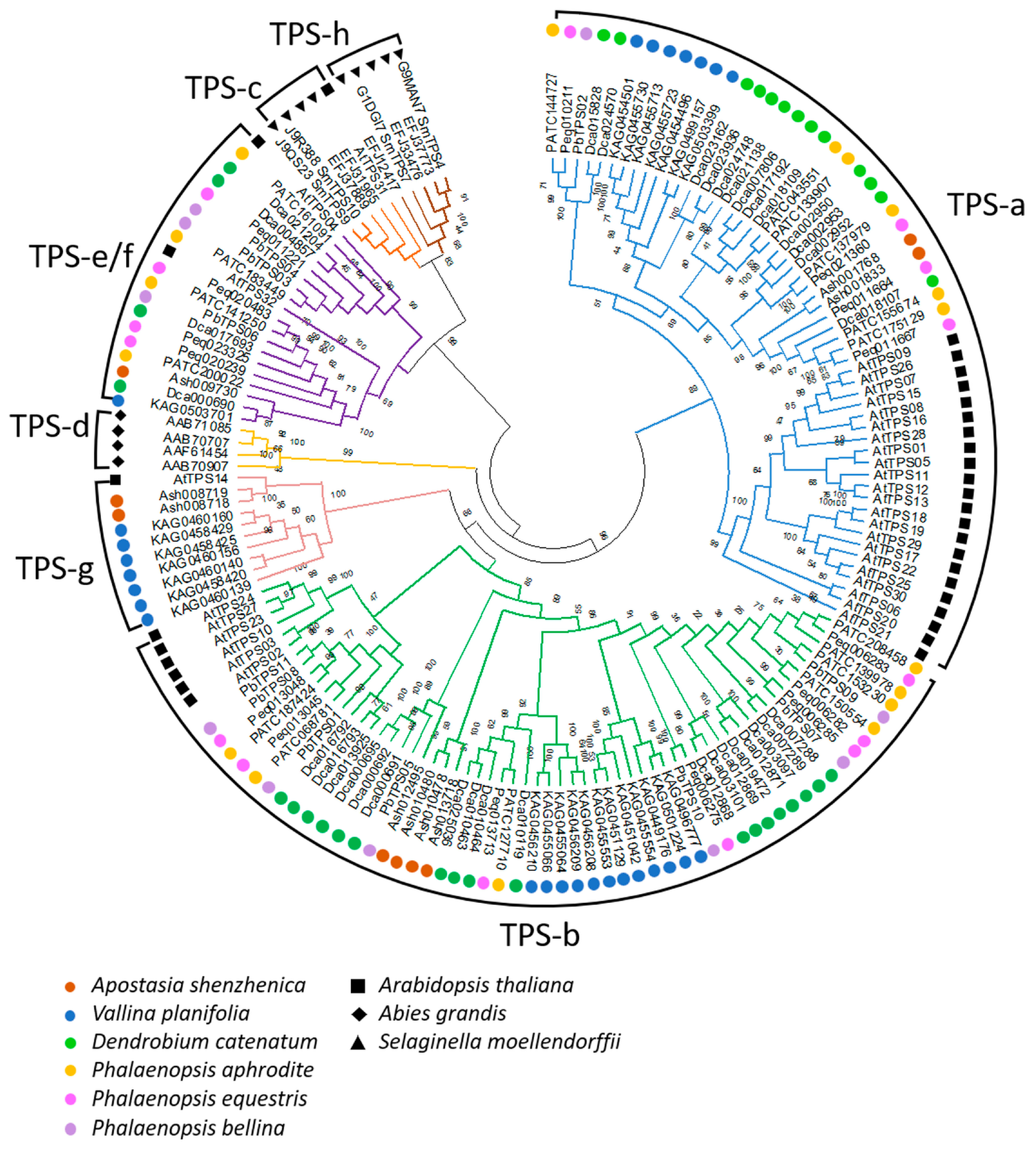
3. The Evolution of TPS Genes in Orchidaceae Species
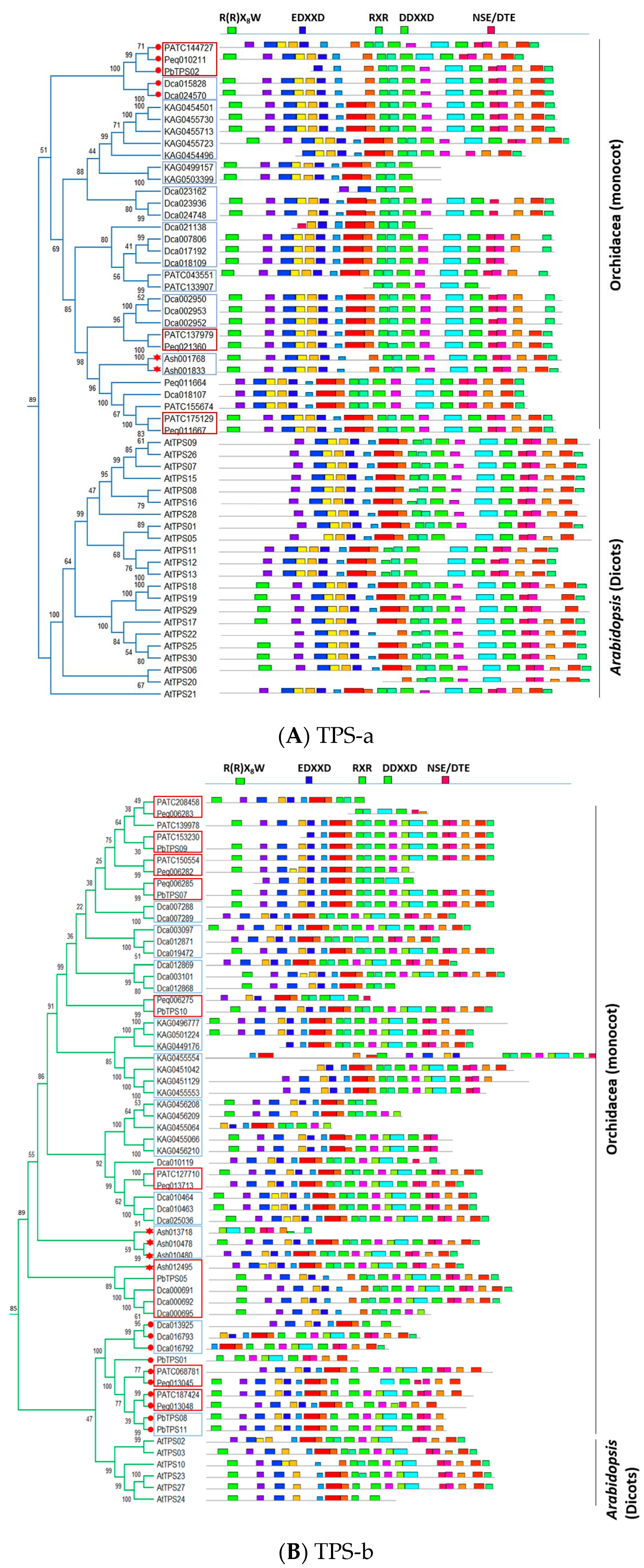
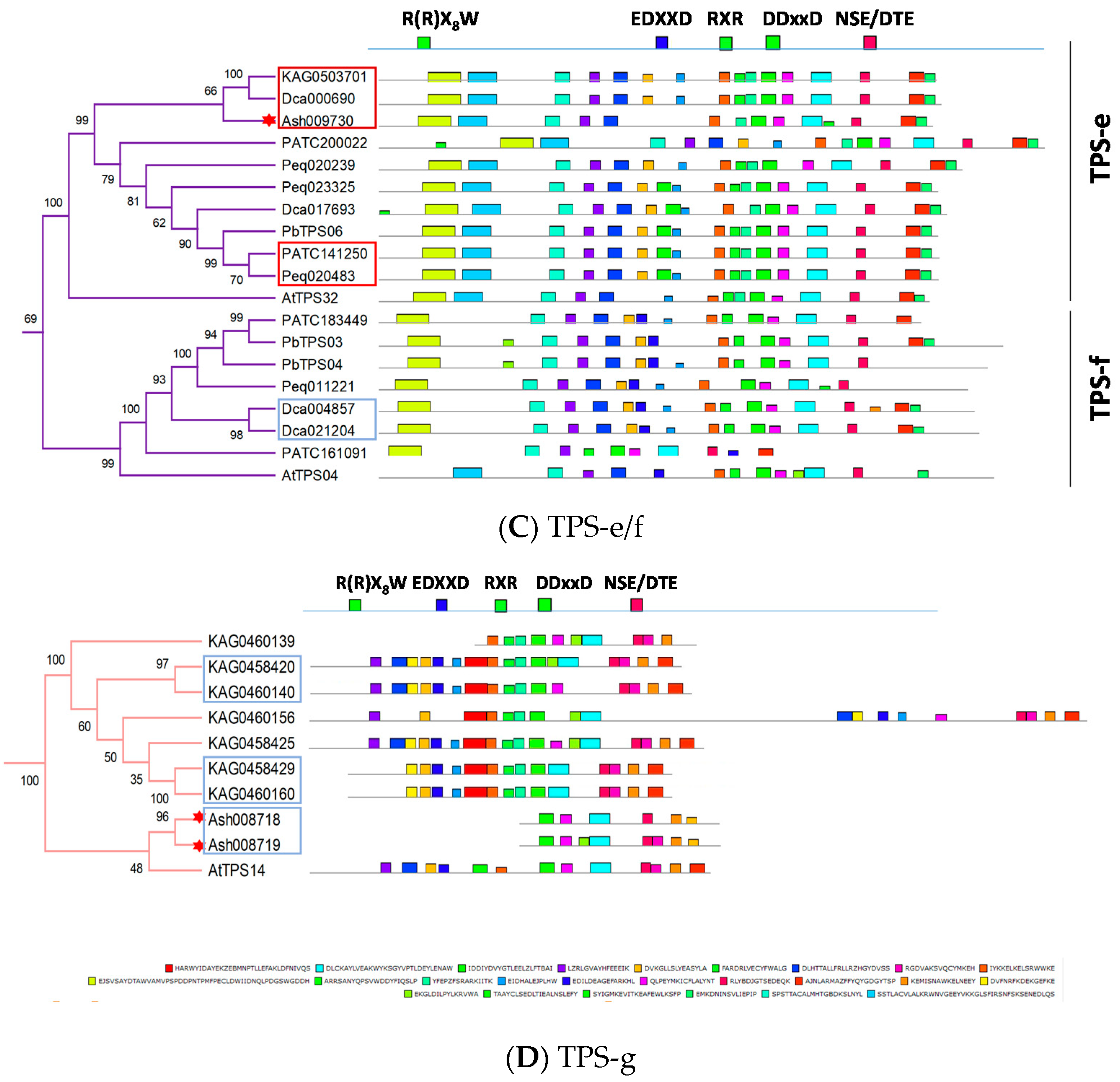
4. The Arrangement of TPS
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene ID | Accession Number of TPS Gene |
|---|---|---|
| Apostasia shenzhenica1 | Ash001768 | Ash001768 |
| Ash001833 | Ash001833 | |
| Ash008718 | Ash008718 | |
| Ash008719 | Ash008719 | |
| Ash009730 | Ash009730 | |
| Ash010478 | Ash010478 | |
| Ash010480 | Ash010480 | |
| Ash012495 | Ash012495 | |
| Ash013718 | Ash013718 | |
| Vallina planifolia2 | KAG0449176 | KAG0449176 |
| KAG0451042 | KAG0451042 | |
| KAG0451129 | KAG0451129 | |
| KAG0454496 | KAG0454496 | |
| KAG0454501 | KAG0454501 | |
| KAG0455064 | KAG0455064 | |
| KAG0455066 | KAG0455066 | |
| KAG0455553 | KAG0455553 | |
| KAG0455554 | KAG0455554 | |
| KAG0455713 | KAG0455713 | |
| KAG0455723 | KAG0455723 | |
| KAG0455730 | KAG0455730 | |
| KAG0456208 | KAG0456208 | |
| KAG0456209 | KAG0456209 | |
| KAG0456210 | KAG0456210 | |
| KAG0458420 | KAG0458420 | |
| KAG0458425 | KAG0458425 | |
| KAG0458429 | KAG0458429 | |
| KAG0460139 | KAG0460139 | |
| KAG0460140 | KAG0460140 | |
| KAG0460156 | KAG0460156 | |
| KAG0460160 | KAG0460160 | |
| KAG0496777 | KAG0496777 | |
| KAG0499157 | KAG0499157 | |
| KAG0501224 | KAG0501224 | |
| KAG0503399 | KAG0503399 | |
| KAG0503701 | KAG0503701 | |
| Dendrobium catenatum1 | Dca000690 | Dca000690 |
| Dca000691 | Dca000691 | |
| Dca000692 | Dca000692 | |
| Dca000695 | Dca000695 | |
| Dca002950 | Dca002950 | |
| Dca002952 | Dca002952 | |
| Dca002953 | Dca002953 | |
| Dca003097 | Dca003097 | |
| Dca003101 | Dca003101 | |
| Dca004857 | Dca004857 | |
| Dca007288 | Dca007288 | |
| Dca007289 | Dca007289 | |
| Dca007806 | Dca007806 | |
| Dca010119 | Dca010119 | |
| Dca010463 | Dca010463 | |
| Dca010464 | Dca010464 | |
| Dca012868 | Dca012868 | |
| Dca012869 | Dca012869 | |
| Dca012871 | Dca012871 | |
| Dca013925 | Dca013925 | |
| Dca015828 | Dca015828 | |
| Dca016792 | Dca016792 | |
| Dca016793 | Dca016793 | |
| Dca017192 | Dca017192 | |
| Dca017693 | Dca017693 | |
| Dca018107 | Dca018107 | |
| Dca018109 | Dca018109 | |
| Dca019472 | Dca019472 | |
| Dca021138 | Dca021138 | |
| Dca021204 | Dca021204 | |
| Dca023162 | Dca023162 | |
| Dca023936 | Dca023936 | |
| Dca024570 | Dca024570 | |
| Dca024748 | Dca024748 | |
| Dca025036 | Dca025036 | |
| Phalaenopsis aphrodite3 | PATC043551 | PATC043551 |
| PATC068781 | PATC068781 | |
| PATC127710 | PATC127710 | |
| PATC133907 | PATC133907 | |
| PATC137979 | PATC137979 | |
| PATC139978 | PATC139978 | |
| PATC141250 | PATC141250 | |
| PATC144727 | PATC144727 | |
| PATC150554 | PATC150554 | |
| PATC153230 | PATC153230 | |
| PATC155674 | PATC155674 | |
| PATC161091 | PATC161091 | |
| PATC175129 | PATC175129 | |
| PATC183449 | PATC183449 | |
| PATC187424 | PATC187424 | |
| PATC200022 | PATC200022 | |
| PATC208458 | PATC208458 | |
| Phalaenopsis equestris1 | Peq006275 | Peq006275 |
| Peq006282 | Peq006282 | |
| Peq006283 | Peq006283 | |
| Peq006285 | Peq006285 | |
| Peq010211 | Peq010211 | |
| Peq011221 | Peq011221 | |
| Peq011664 | Peq011664 | |
| Peq011667 | Peq011667 | |
| Peq013045 | Peq013045 | |
| Peq013048 | Peq013048 | |
| Peq013713 | Peq013713 | |
| Peq020239 | Peq020239 | |
| Peq020483 | Peq020483 | |
| Peq021360 | Peq021360 | |
| Peq023325 | Peq023325 | |
| Phalaenopsis bellina4 | PbTPS01 | CL86.Contig1 |
| PbTPS02 | CL214.Contig2 | |
| PbTPS03 | CL376.Contig6 | |
| PbTPS04 | CL376.Contig8 | |
| PbTPS05 | CL1323.Contig1 | |
| PbTPS06 | CL2295.Contig2 | |
| PbTPS07 | CL2800.Contig3 | |
| PbTPS08 | CL4514.Contig2 | |
| PbTPS09 | CL6288.Contig1 | |
| PbTPS10 | CL6288.Contig7 | |
| PbTPS11 | Unigene4722 | |
| Arabidopsis thaliana2 | AtTPS1 | At4g15870 |
| AtTPS2 | At4g16730 | |
| AtTPS3 | At4g16740 | |
| AtTPS4 | At1g61120 | |
| AtTPS5 | At4g20230 | |
| AtTPS6 | At1g70080 | |
| AtTPS7 | At4g20200 | |
| AtTPS8 | At4g20210 | |
| AtTPS9 | At2g23230 | |
| AtTPS10 | At2g24210 | |
| AtTPS11 | At5g44630 | |
| AtTPS12 | At4g13280 | |
| AtTPS13 | At4g13300 | |
| AtTPS14 | At1g61680 | |
| AtTPS15 | At3g29190 | |
| AtTPS16 | At3g29110 | |
| AtTPS17 | At3g14490 | |
| AtTPS18 | At3g14520 | |
| AtTPS19 | At3g14540 | |
| AtTPS20 | At5g48110 | |
| AtTPS21 | At5g23960 | |
| AtTPS22 | At1g33750 | |
| AtTPS23 | At3g25830 | |
| AtTPS24 | At3g25810 | |
| AtTPS25 | At3g29410 | |
| AtTPS26 | At1g66020 | |
| AtTPS27 | At1g48820 | |
| AtTPS28 | At1g48800 | |
| AtTPS29 | At1g31950 | |
| AtTPS30 | At3g32030 | |
| AtTPS31 | At4g02780 | |
| AtTPS32 | At1g79460 | |
| Abies grandis2 | AAB70707 | AGU87910 |
| AAB70907 | AF006193 | |
| AAB71085 | U87909 | |
| AAF61454 | AF139206 | |
| Selaginella moellendorffii2 | EFJ31965 | GL377573 |
| EFJ37889 | GL377565 | |
| J9QS23_SmTPS9 | XM_002960304 | |
| |J9R388_SmTPS10 | XM_024672072 | |
| G9MAN7_SmTPS4 | XM_024672355. | |
| G1DGI7_SmTPS7 | XM_024689660 | |
| EFJ12417 | GL377639 | |
| EFJ37773 | GL377565 | |
| EFJ33476 | GL377571 |
References
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: Nature’s diversity and ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.; Sun, T.; Kawaide, H.; Kamiya, Y. The GA2 locus of Arabidopsis thaliana encodes ent-kaurene synthase of gibberellin biosynthesis. Plant Physiol. 1998, 116, 1271–1278. [Google Scholar] [CrossRef] [Green Version]
- El Tamer, M.K.; Lücker, J.; Bosch, D.; Verhoeven, H.A.; Verstappen, F.W.; Schwab, W.; van Tunen, A.J.; Voragen, A.G.; De Maagd, R.A.; Bouwmeester, H.J. Domain swapping of Citrus limon monoterpene synthases: Impact on enzymatic activity and product specificity. Arch. Biochem. Biophys. 2003, 411, 196–203. [Google Scholar] [CrossRef]
- Lange, B.M.; Severin, K.; Bechthold, A.; Heide, L. Regulatory role of microsomal 3-hydroxy-3-methylglutaryl-coenzyme A reductase for shikonin biosynthesis in Lithospermum erythrorhizon cell suspension cultures. Planta 1998, 204, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Aubourg, S.; Lecharny, A.; Bohlmann, J. Genomic analysis of the terpenoid synthase (AtTPS) gene family of Arabidopsis thaliana. Mol. Genet. Genom. 2002, 267, 730–745. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Kawaide, H.; Notomi, M.; Sakigi, Y.; Matsuo, A.; Nozaki, H. Identification and functional analysis of bifunctional ent-kaurene synthase from the moss Physcomitrella patens. FEBS Lett. 2006, 580, 6175–6181. [Google Scholar] [CrossRef] [Green Version]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Aubourg, S.; Schouwey, M.B.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.T.; Bohlmann, J. Functional annotation, genome organization and phylogeny of the grapevine (Vitis vinifera) terpene synthase gene family based on genome assembly, FLcDNA cloning, and enzyme assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falara, V.; Akhtar, T.A.; Nguyen, T.T.; Spyropoulou, E.A.; Bleeker, P.M.; Schauvinhold, I.; Matsuba, Y.; Bonini, M.E.; Schilmiller, A.L.; Last, R.L.; et al. The tomato terpene synthase gene family. Plant Physiol. 2011, 157, 770–789. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Kollner, T.G.; Yin, Y.; Jiang, Y.; Chen, H.; Xu, Y.; Gershenzon, J.; Pichersky, E.; Chen, F. Nonseed plant Selaginella moellendorffi has both seed plant and microbial types of terpene synthases. Proc. Natl. Acad. Sci. USA 2012, 109, 14711–14715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Huang, F.; Wang, X.; Zhang, M.; Zheng, R.; Wang, J.; Yu, D. Genome-wide analysis of terpene synthases in soybean: Functional characterization of GmTPS3. Gene 2014, 544, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Irmisch, S.; Jiang, Y.; Chen, F.; Gershenzon, J.; Köllner, T.G. Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa). BMC Plant Biol. 2014, 14, 270. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Li, G.; Kollner, T.G.; Jia, Q.; Gershenzon, J.; Chen, F. Positive Darwinian selection is a driving force for the diversification of terpenoid biosynthesis in the genus Oryza. BMC Plant Biol. 2014, 14, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Zhao, C.; Zhang, G.; Teixeira da Silva, J.A.; Duan, J. Genome-Wide Identification and Expression Profile of TPS Gene Family in Dendrobium officinale and the Role of DoTPS10 in Linalool Biosynthesis. Int. J. Mol. Sci. 2020, 21, 5419. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Starks, C.M.; Back, K.; Chappell, J.; Noel, J.P. Structural basis for cyclic terpene biosynthesis by tobacco 5-epi-aristolochene synthase. Science 1997, 277, 1815–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rynkiewicz, M.J.; Cane, D.E.; Christianson, D.W. Structure of trichodiene synthase from Fusarium sporotrichioides provides mechanistic inferences on the terpene cyclization cascade. Proc. Natl. Acad. Sci. USA 2001, 98, 13543–13548. [Google Scholar] [CrossRef] [Green Version]
- Whittington, D.A.; Wise, M.L.; Urbansky, M.; Coates, R.M.; Croteau, R.B.; Christianson, D.W. Bornyl diphosphate synthase: Structure and strategy for carbocation manipulation by a terpenoid cyclase. Proc. Natl. Acad. Sci. USA 2002, 99, 15375–15380. [Google Scholar] [CrossRef] [Green Version]
- Christianson, D.W. Structural biology and chemistry of the terpenoid cyclases. Chem. Rev. 2006, 106, 3412–3442. [Google Scholar] [CrossRef]
- Degenhardt, J.; Kollner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef]
- Wise, M.L.; Croteau, R. Monoterpene biosynthesis. In Comprehensive Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 1999; Volume 2, pp. 97–159. [Google Scholar]
- MacMillan, J.; Beale, M.H. Diterpene biosynthesis. In Comprehensive Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 1999; Volume 2, pp. 217–243. [Google Scholar]
- Taniguchi, S.; Hosokawa-Shinonaga, Y.; Tamaoki, D.; Yamada, S.; Akimitsu, K.; Gomi, K. Jasmonate induction of the monoterpene linalool confers resistance to rice bacterial blight and its biosynthesis is regulated by JAZ protein in rice. Plant Cell Environ. 2014, 37, 451–461. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, J.; Li, K.; Yu, D. Identification and characterization of a novel monoterpene synthase from soybean restricted to neryl diphosphate precursor. PLoS ONE 2013, 8, e75972. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, A.; Giri, A.P.; Verstappen, F.W.; Bertea, C.M.; Sevenier, R.; Sun, Z.; Jongsma, M.A.; Schwab, W.; Bouwmeester, H.J. Gain and loss of fruit flavor compounds produced by wild and cultivated strawberry species. Plant Cell 2004, 16, 3110–3131. [Google Scholar] [CrossRef] [Green Version]
- Bohlmann, J.; Crock, J.; Jetter, R.; Croteau, R. Terpenoid-based defenses in conifers: CDNA cloning, characterization, and functional expression of wound-inducible (E)-alpha-bisabolene synthase from grand fir (Abies grandis). Proc. Natl. Acad. Sci. USA 1998, 95, 6756–6761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeling, C.I.; Weisshaar, S.; Lin, R.P.; Bohlmann, J. Functional plasticity of paralogous diterpene synthases involved in conifer defense. Proc. Natl. Acad. Sci. USA 2008, 105, 1085–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Unsicker, S.B.; Kunert, G.; Gershenzon, J. Protective perfumes: The role of vegetative volatiles in plant defense against herbivores. Curr. Opin. Plant Biol. 2009, 12, 479–485. [Google Scholar] [CrossRef]
- Weitzel, C.; Simonsen, H.T. Cytochrome P450-enzymes involved in the biosynthesis of mono- and sesquiterpenes. Phytochem. Rev. 2015, 14, 7–24. [Google Scholar] [CrossRef]
- Fay, M.F. Orchid conservation: How can we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Doucette, A.; Caro, G.G.; McDaniel, J.; Clements, M.A.; et al. Orchid historical biogeography, diversification, Antarctica and the paradox of orchid dispersal. J. Biogeogr. 2016, 43, 1905–1916. [Google Scholar] [CrossRef]
- Chase, M.W.; Cameron, K.M.; Barrett, R.L.; Freudenstein, J.V. DNA data and Orchidaceae systematics: A new phylogenetic classification. In Orchid Conservation; Natural History Publications: Kota Kinabalu, Malaysia, 2003; pp. 69–89. [Google Scholar]
- Raguso, R.A.; Levin, R.A.; Foose, S.E.; Holmberg, M.W.; McDade, L.A. Fragrance chemistry, nocturnal rhythms and pollination “syndromes” in Nicotiana. Phytochemistry 2003, 63, 265–284. [Google Scholar] [CrossRef]
- Gregg, K.B. Variation in floral fragrances and morphology: Incipient speciation in Cycnoches? Bot. Gaz. 1983, 144, 566–576. [Google Scholar] [CrossRef]
- Tan, K.H.; Tan, L.T.; Nishida, R. Floral phenylpropanoid cocktail and architecture of Bulbophyllum vinaceum orchid in attracting fruit flies for pollination. J. Chem. Ecol. 2006, 32, 2429–2441. [Google Scholar] [CrossRef] [PubMed]
- Cancino, A.D.M.; Damon, A. Fragrance analysis of euglossine bee pollinated orchids from Soconusco, south-east Mexico. Plant Species Biol. 2007, 22, 127–132. [Google Scholar]
- Salzmann, C.C.; Schiestl, F.P. Odour and colour polymorphism in the food-deceptive orchid Dactylorhiza romana. Plant Syst. Evol. 2007, 267, 37–45. [Google Scholar] [CrossRef]
- Brodmann, J.; Twele, R.; Francke, W.; Luo, Y.B.; Song, X.Q.; Ayasse, M. Orchid Mimics Honey Bee Alarm Pheromone in Order to Attract Hornets for Pollination. Curr. Biol. 2009, 19, 1368–1372. [Google Scholar] [CrossRef] [Green Version]
- Delle-Vedove, R.; Juillet, N.; Bessiere, J.M.; Grison, C.; Barthes, N.; Pailler, T.; Dormont, L.; Schatz, B. Colour-scent associations in a tropical orchid: Three colours but two odours. Phytochemistry 2011, 72, 735–742. [Google Scholar] [CrossRef]
- Wiemer, A.P.; More, M.; Benitez-Vieyra, S.; Cocucci, A.A.; Raguso, R.A.; Sersic, A.N. A simple floral fragrance and unusual osmophore structure in Cyclopogon elatus (Orchidaceae). Plant Biol. 2009, 11, 506–514. [Google Scholar] [CrossRef]
- Nunes, C.E.; Gerlach, G.; Bandeira, K.D.; Gobbo-Neto, L.; Pansarin, E.R.; Sazima, M. Two orchids, one scent? Floral volatiles of Catasetum cernuum and Gongora bufonia suggest convergent evolution to a unique pollination niche. Flora 2017, 232, 207–216. [Google Scholar] [CrossRef]
- Ramya, M.; Park, P.H.; Chuang, Y.-C.; Kwon, O.K.; An, H.R.; Park, P.M.; Baek, Y.S.; Kang, B.-C.; Tsai, W.-C.; Chen, H.-H. RNA sequencing analysis of Cymbidium goeringii identifies floral scent biosynthesis related genes. BMC Plant Biol. 2019, 19, 337. [Google Scholar] [CrossRef]
- Kaiser, R.A. On the Scent of Orchids; ACS Publications: Washington, DC, USA, 1993; pp. 240–268. [Google Scholar]
- Hsiao, Y.Y.; Pan, Z.J.; Hsu, C.C.; Yang, Y.P.; Hsu, Y.C.; Chuang, Y.C.; Shih, H.H.; Chen, W.H.; Tsai, W.C.; Chen, H.H. Research on orchid biology and biotechnology. Plant Cell Physiol. 2011, 52, 1467–1486. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, Y.Y.; Tsai, W.C.; Kuoh, C.S.; Huang, T.H.; Wang, H.C.; Wu, T.S.; Leu, Y.L.; Chen, W.H.; Chen, H.H. Comparison of transcripts in Phalaenopsis bellina and Phalaenopsis equestris (Orchidaceae) flowers to deduce monoterpene biosynthesis pathway. BMC Plant Biol. 2006, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Awano, K.; Ichikawa, Y.; Tokuda, K.; Kuraoka, M. Volatile Components of the Flowers of Two Calanthe Species. Flavour Fragr. J. 1997, 12, 327–333. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.D.; Chappell, J. Isoprenoid biosynthesis in plants: Carbon partitioning within the cytoplasmic pathway. Crit. Rev. Biochem. Mol. Biol. 1999, 34, 95–106. [Google Scholar] [CrossRef]
- Sapir-Mir, M.; Mett, A.; Belausov, E.; Tal-Meshulam, S.; Frydman, A.; Gidoni, D.; Eyal, Y. Peroxisomal Localization of Arabidopsis Isopentenyl Diphosphate Isomerases Suggests That Part of the Plant Isoprenoid Mevalonic Acid Pathway Is Compartmentalized to Peroxisomes. Plant Physiol. 2008, 148, 1219–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashour, M.; Wink, M.; Gershenzon, J. Biochemistry of terpenoids: Monoterpenes, sesquiterpenes and diterpenes. In Annual Plant Reviews: Biochemistry of Plant Secondary Metabolism; Wiley-Blackwell: Chichester, UK, 2010; pp. 258–303. [Google Scholar]
- Zhang, G.Q.; Liu, K.W.; Li, Z.; Lohaus, R.; Hsiao, Y.Y.; Niu, S.C.; Wang, J.Y.; Lin, Y.C.; Xu, Q.; Chen, L.J.; et al. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasing, T.; Tang, H.; Brym, M.; Khazi, F.; Huang, T.; Chambers, A.H. A phased Vanilla planifolia genome enables genetic improvement of flavour and production. Nat. Food 2020, 1, 811–819. [Google Scholar] [CrossRef]
- Zhang, G.-Q.; Xu, Q.; Bian, C.; Tsai, W.-C.; Yeh, C.-M.; Liu, K.-W.; Yoshida, K.; Zhang, L.-S.; Chang, S.-B.; Chen, F.; et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 19029. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Liu, X.; Vanneste, K.; Proost, S.; Tsai, W.C.; Liu, K.W.; Chen, L.J.; He, Y.; Xu, Q.; Bian, C.; et al. The genome sequence of the orchid Phalaenopsis equestris. Nat. Genet. 2015, 47, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.C.; Fu, C.H.; Hsiao, Y.Y.; Huang, Y.M.; Chen, L.J.; Wang, M.; Liu, Z.J.; Chen, H.H. OrchidBase 2.0: Comprehensive collection of Orchidaceae floral transcriptomes. Plant Cell Physiol. 2013, 54, e7. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.P.; Kamiya, Y. The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell 1994, 6, 1509–1518. [Google Scholar] [PubMed] [Green Version]
- Tsai, W.C.; Dievart, A.; Hsu, C.C.; Hsiao, Y.Y.; Chiou, S.Y.; Huang, H.; Chen, H.H. Post genomics era for orchid research. Bot. Stud. 2017, 58, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Trapp, S.C.; Croteau, R.B. Genomic organization of plant terpene synthases and molecular evolutionary implications. Genetics 2001, 158, 811–832. [Google Scholar] [CrossRef] [PubMed]
- Shalev, T.J.; Yuen, M.M.S.; Gesell, A.; Yuen, A.; Russell, J.H.; Bohlmann, J. An annotated transcriptome of highly inbred Thuja plicata (Cupressaceae) and its utility for gene discovery of terpenoid biosynthesis and conifer defense. Tree Genet. Genomes 2018, 14, 35. [Google Scholar] [CrossRef]
- Williams, D.C.; McGarvey, D.J.; Katahira, E.J.; Croteau, R. Truncation of limonene synthase preprotein provides a fully active ‘pseudomature’ form of this monoterpene cyclase and reveals the function of the amino-terminal arginine pair. Biochemistry 1998, 37, 12213–12220. [Google Scholar] [CrossRef]
- Hyatt, D.C.; Youn, B.; Zhao, Y.; Santhamma, B.; Coates, R.M.; Croteau, R.B.; Kang, C. Structure of limonene synthase, a simple model for terpenoid cyclase catalysis. Proc. Natl. Acad. Sci. USA 2007, 104, 5360–5365. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Zhang, Y.; Mann, F.M.; Huang, C.; Mukkamala, D.; Hudock, M.P.; Mead, M.E.; Prisic, S.; Wang, K.; Lin, F.Y.; et al. Diterpene cyclases and the nature of the isoprene fold. Proteins Struct. Funct. Bioinform. 2010, 78, 2417–2432. [Google Scholar] [CrossRef] [Green Version]
- Keeling, C.I.; Dullat, H.K.; Yuen, M.; Ralph, S.G.; Jancsik, S.; Bohlmann, J. Identification and Functional Characterization of Monofunctional ent-Copalyl Diphosphate and ent-Kaurene Synthases in White Spruce Reveal Different Patterns for Diterpene Synthase Evolution for Primary and Secondary Metabolism in Gymnosperms. Plant Physiol. 2010, 152, 1197–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicandri, E.; Paolacci, A.R.; Osadolor, S.; Sorgona, A.; Badiani, M.; Ciaffi, M. On the Evolution and Functional Diversity of Terpene Synthases in the Pinus Species: A Review. J. Mol. Evol. 2020, 88, 253–283. [Google Scholar] [CrossRef]
- Chen, X.; Kollner, T.G.; Shaulsky, G.; Jia, Q.; Dickschat, J.S.; Gershenzon, J.; Chen, F. Diversity and Functional Evolution of Terpene Synthases in Dictyostelid Social Amoebae. Sci. Rep. 2018, 8, 14361. [Google Scholar] [CrossRef]
- Ker, D.S.; Pang, S.L.; Othman, N.F.; Kumaran, S.; Tan, E.F.; Krishnan, T.; Chan, K.G.; Othman, R.; Hassan, M.; Ng, C.L. Purification and biochemical characterization of recombinant Persicaria minor beta-sesquiphellandrene synthase. Peer J. 2017, 5, e2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Zhao, T.; Sheng, Y.; Zheng, T.; Fu, L.; Zhang, Y. Dendrobium officinale Kimura et Migo: A Review on Its Ethnopharmacology, Phytochemistry, Pharmacology, and Industrialization. Evid. Based Complementary Altern. Med. 2017, 2017, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, Y.C.; Hung, Y.C.; Tsai, W.C.; Chen, W.H.; Chen, H.H. PbbHLH4 regulates floral monoterpene biosynthesis in Phalaenopsis orchids. J. Exp. Bot. 2018, 69, 4363–4377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuizen, N.J.; Wang, M.Y.; Matich, A.J.; Green, S.A.; Chen, X.; Yauk, Y.K.; Beuning, L.L.; Nagegowda, D.A.; Dudareva, N.; Atkinson, R.G. Two terpene synthases are responsible for the major sesquiterpenes emitted from the flowers of kiwifruit (Actinidia deliciosa). J. Exp. Bot. 2009, 60, 3203–3219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yauk, Y.-K.; Nieuwenhuizen, N.J.; Matich, A.J.; Wang, M.Y.; Perez, R.L.; Atkinson, R.G.; Beuning, L.L. Characterisation of an (S)-linalool synthase from kiwifruit (Actinidia arguta) that catalyses the first committed step in the production of floral lilac compounds. Funct. Plant Biol. 2010, 37, 232–243. [Google Scholar] [CrossRef]
- Krokida, A.; Delis, C.; Geisler, K.; Garagounis, C.; Tsikou, D.; Peña-Rodríguez, L.M.; Katsarou, D.; Field, B.; Osbourn, A.E.; Papadopoulou, K.K. A metabolic gene cluster in Lotus japonicus discloses novel enzyme functions and products in triterpene biosynthesis. New Phytol. 2013, 200, 675–690. [Google Scholar] [CrossRef]
- Jiang, S.Y.; Jin, J.; Sarojam, R.; Ramachandran, S. A Comprehensive Survey on the Terpene Synthase Gene Family Provides New Insight into Its Evolutionary Patterns. Genome Biol. Evol. 2019, 11, 2078–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuba, Y.; Nguyen, T.T.; Wiegert, K.; Falara, V.; Gonzales-Vigil, E.; Leong, B.; Schafer, P.; Kudrna, D.; Wing, R.A.; Bolger, A.M.; et al. Evolution of a complex locus for terpene biosynthesis in solanum. Plant Cell 2013, 25, 2022–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Külheim, C.; Padovan, A.; Hefer, C.; Krause, S.T.; Köllner, T.G.; Myburg, A.A.; Degenhardt, J.; Foley, W.J. The Eucalyptus terpene synthase gene family. BMC Genom. 2015, 16, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimura, K.; Okada, A.; Okada, K.; Jikumaru, Y.; Ko, K.-W.; Toyomasu, T.; Sassa, T.; Hasegawa, M.; Kodama, O.; Shibuya, N.; et al. Identification of a Biosynthetic Gene Cluster in Rice for Momilactones. J. Biol. Chem. 2007, 282, 34013–34018. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Kollner, T.G.; Li, G.; Wei, G.; Chen, X.; Zeng, D.; Qian, Q.; Chen, F. Combinatorial Evolution of a Terpene Synthase Gene Cluster Explains Terpene Variations in Oryza. Plant Physiol. 2020, 182, 480–492. [Google Scholar] [CrossRef] [Green Version]
- Boutanaev, A.M.; Moses, T.; Zi, J.; Nelson, D.R.; Mugford, S.T.; Peters, R.J.; Osbourn, A. Investigation of terpene diversification across multiple sequenced plant genomes. Proc. Natl. Acad. Sci. USA 2015, 112, E81–E88. [Google Scholar] [CrossRef] [Green Version]


| TPS Subfamily | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Species | a | b | c | d | e/f | g | h | Total | Reference |
| Apostasia shenzhenica | 2 | 4 | 0 | 0 | 1 | 2 | 0 | 9 | This research |
| Vallina planifolia | 7 | 12 | 0 | 0 | 1 | 7 | 0 | 27 | This research |
| Dendrobium catenatum | 13 | 18 | 0 | 0 | 4 | 0 | 0 | 35 | This research |
| Phalaenopsis equestris | 4 | 7 | 0 | 0 | 4 | 0 | 0 | 15 | This research |
| Phalaenopsis aphrodite | 6 | 7 | 0 | 0 | 4 | 0 | 0 | 17 | This research |
| Phalaenopsis bellina | 1 | 7 | 0 | 0 | 3 | 0 | 0 | 11 | This research |
| Arabidopsis thaliana | 22 | 6 | 1 | 0 | 2 | 1 | 0 | 32 | Aubourg et al. (2002) [6] |
| Solanum lycopersicum | 12 | 8 | 2 | 0 | 5 | 2 | 0 | 29 | Falara et al. (2011) [10] |
| Oryza sativa | 18 | 0 | 3 | 0 | 9 | 2 | 0 | 32 | Chen et al. (2014) [14] |
| Sorghum bicolor | 15 | 2 | 1 | 0 | 3 | 3 | 0 | 24 | Paterson et al. (2009) [8] |
| Vitis vinifera | 30 | 19 | 2 | 0 | 1 | 17 | 0 | 69 | Martin et al. (2010) [9] |
| Populus trichocarpa | 16 | 14 | 2 | 0 | 3 | 3 | 0 | 38 | Irmisch et al., (2014) [13] |
| Selaginella moellendorffii | 0 | 0 | 3 | 0 | 3 | 0 | 8 | 14 | Li et al., (2012) [11] |
| Species | Number of Clusters | Number of Scaffolds | Number of Clustered TPSs | Number of Total TPSs | Percentage of Clustered TPSs (%) |
|---|---|---|---|---|---|
| Apostasia shenzhenica | 3 | 3 | 6 | 9 | 66.7 |
| Vallina planifolia | 7 | 5 | 22 | 27 | 81.5 |
| Dendrobium catenatum | 8 | 7 | 20 | 35 | 57.1 |
| Phalaenopsis equestris | 3 | 3 | 8 | 15 | 53.3 |
| Arabidopsis thaliana [6] | 5 | 5 | 13 | 32 | 40.6 |
| Species | Genome Size (Mb) | Cluster Length of TPSs (Kb) | Total Length of TPSs (Kb) | Cluster Density of TPSs (%) |
|---|---|---|---|---|
| Apostasia shenzhenica | 349 | 26 | 56 | 47.3 |
| Vallina planifolia | 744 | 595 | 758 | 78.6 |
| Dendrobium catenatum | 1104 | 125 | 248 | 50.5 |
| Phalaenopsis equestris | 1064 | 62 | 158 | 38.9 |
| Arabidopsis thaliana | 120 | 43 | 109 | 39.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.-M.; Huang, H.; Chuang, Y.-C.; Chen, W.-H.; Wang, C.-N.; Chen, H.-H. Evolution of Terpene Synthases in Orchidaceae. Int. J. Mol. Sci. 2021, 22, 6947. https://doi.org/10.3390/ijms22136947
Huang L-M, Huang H, Chuang Y-C, Chen W-H, Wang C-N, Chen H-H. Evolution of Terpene Synthases in Orchidaceae. International Journal of Molecular Sciences. 2021; 22(13):6947. https://doi.org/10.3390/ijms22136947
Chicago/Turabian StyleHuang, Li-Min, Hsin Huang, Yu-Chen Chuang, Wen-Huei Chen, Chun-Neng Wang, and Hong-Hwa Chen. 2021. "Evolution of Terpene Synthases in Orchidaceae" International Journal of Molecular Sciences 22, no. 13: 6947. https://doi.org/10.3390/ijms22136947
APA StyleHuang, L.-M., Huang, H., Chuang, Y.-C., Chen, W.-H., Wang, C.-N., & Chen, H.-H. (2021). Evolution of Terpene Synthases in Orchidaceae. International Journal of Molecular Sciences, 22(13), 6947. https://doi.org/10.3390/ijms22136947