
Highly Efficient Temperature Inducible CRISPR-Cas9 Gene Targeting in Drosophila suzukii

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
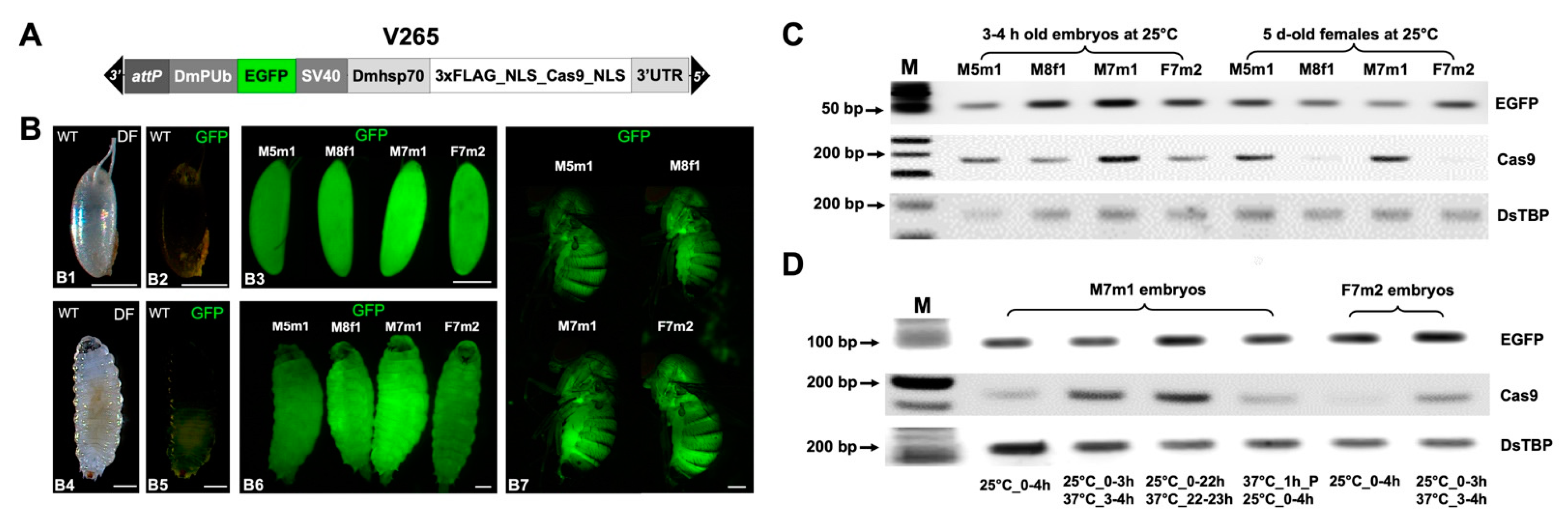
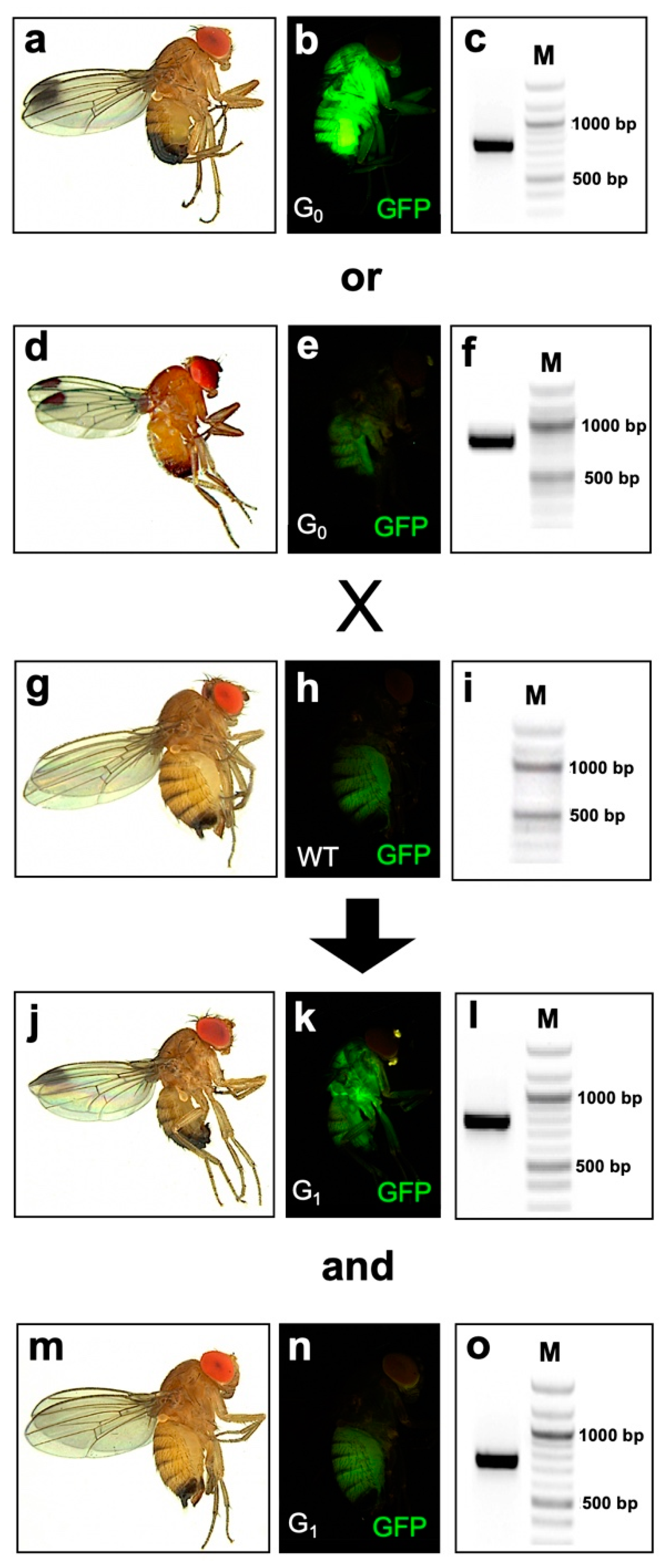
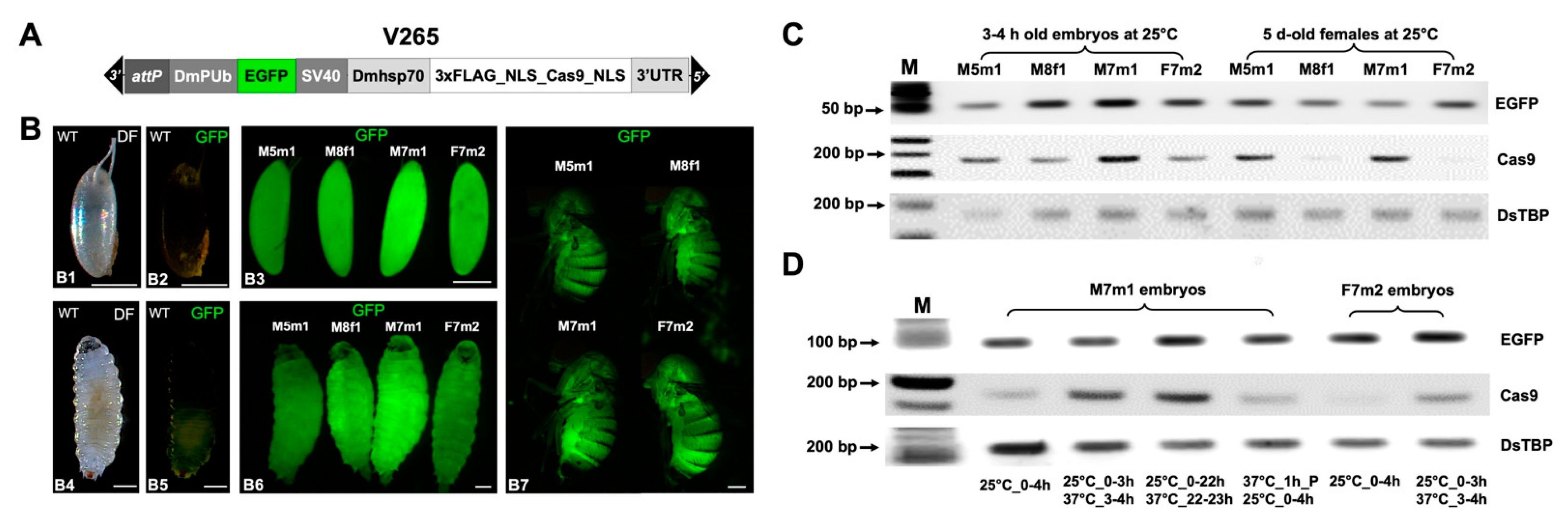
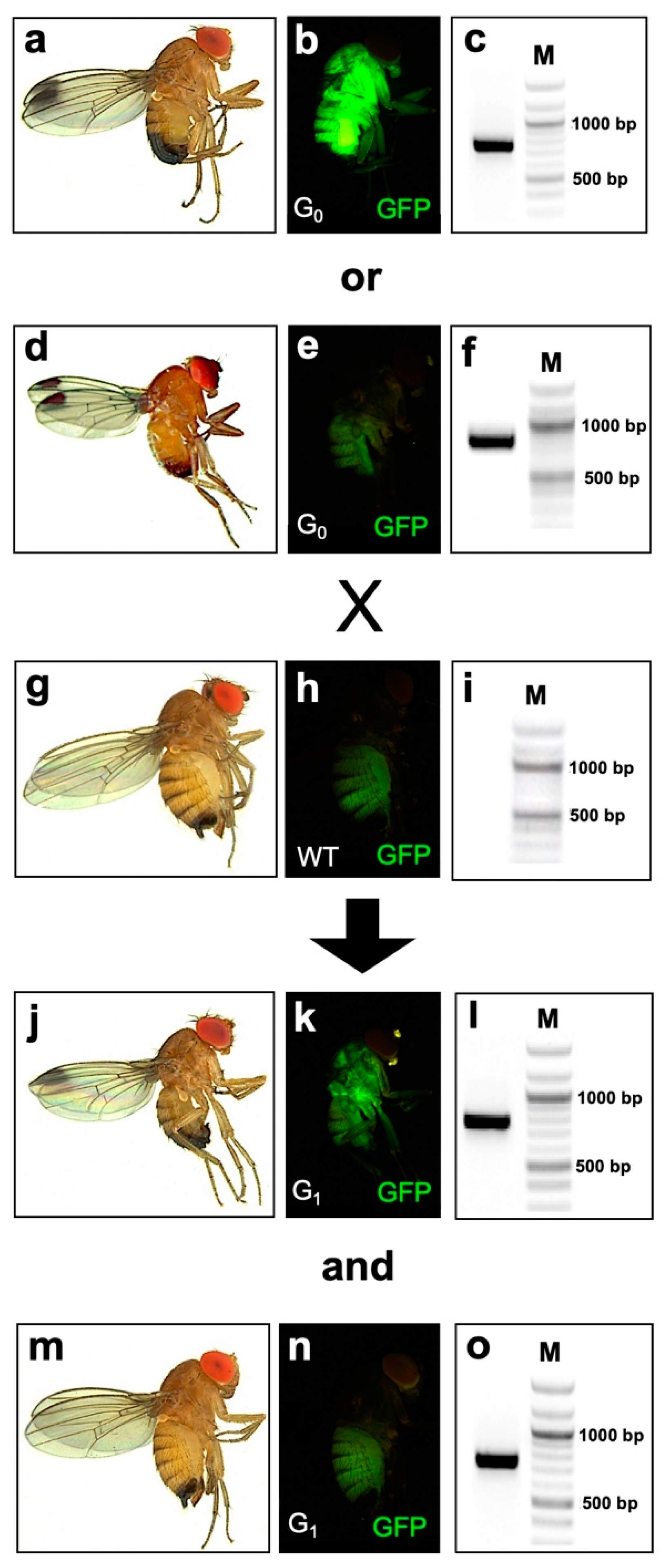
2.1. Generation and Characterization of D. suzukii Cas9-Expressing Lines
2.2. Efficient and Temperature-Dependent Mutagenesis of an EGFP Transgene
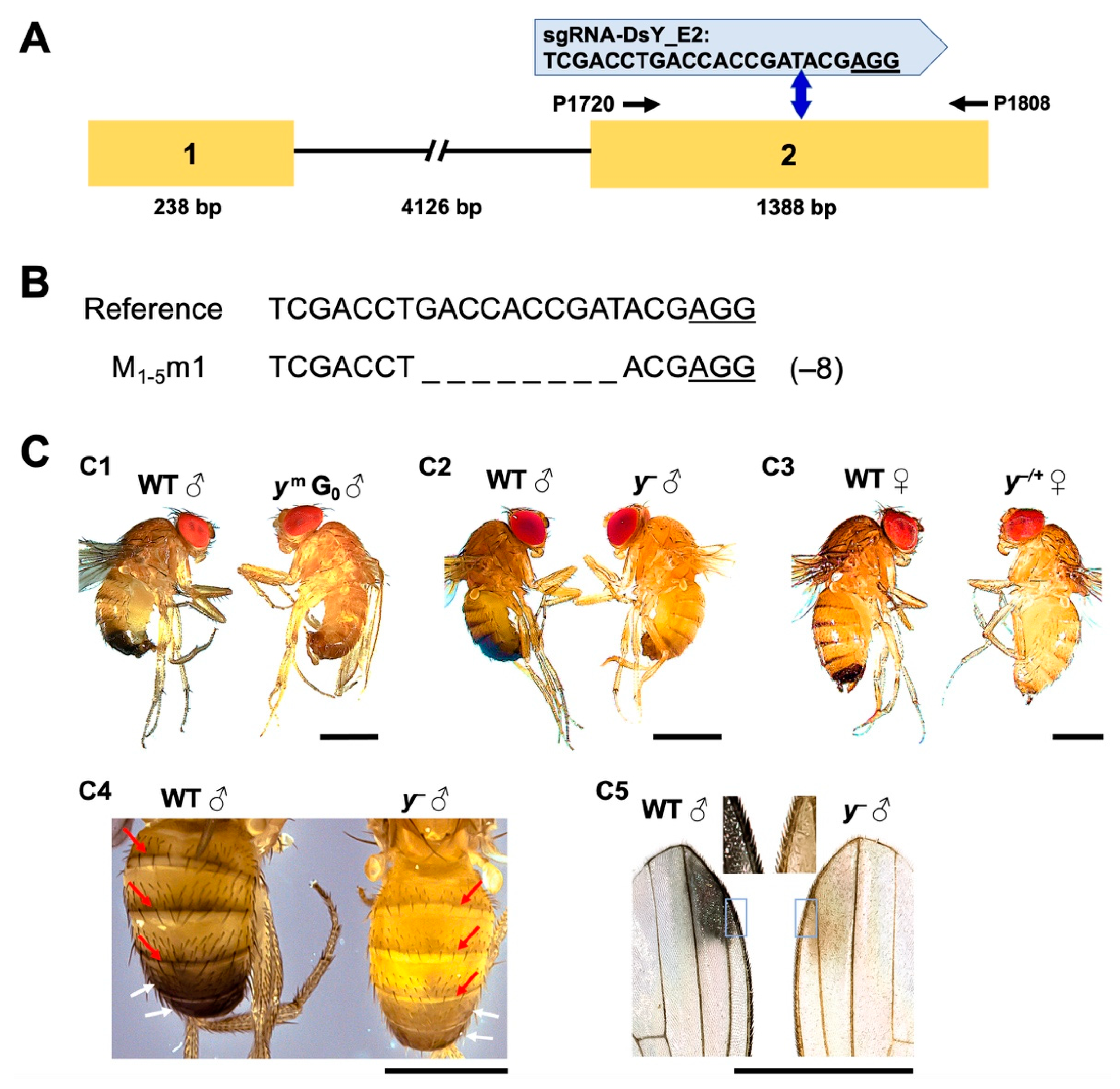
2.3. Disruption of the Endogenous Yellow Gene Causes Loss of Pigmentation and Male Lethality
3. Discussion
4. Methods and Materials
4.1. Insect Rearing and Germline Transformation
4.2. Plasmid Construction
4.3. Heat-Shock (HS) Treatments and Reverse Transcriptase (RT) PCR
4.4. Gene Sequence Isolation and Analysis
4.5. gRNA Design, Off-Target Analysis and Injections
4.6. Crosses, Stock Keeping and Lethality Tests
4.7. Imaging and Genotyping
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest. Manag. 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Cini, A.; Ioriatti, C.; Anforna, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Deprá, M.; Poppe, J.L.; Schmitz, H.J.; De Toni, D.C.; Valente, V.L. The first records of the invasive pest Drosophila suzukii in the South American continent. J. Pest Sci. 2014, 87, 379–383. [Google Scholar] [CrossRef]
- Kwadha, C.A.; Okwaro, L.A.; Kleman, I.; Rehermann, G.; Revadi, S.; Ndlela, S.; Khamis, F.M.; Nderitu, P.W.; Kasina, M.; George, M.K.; et al. Detection of the spotted wing drosophila, Drosophila suzukii, in continental sub-Saharan Africa. J. Pest Sci. 2021, 94, 251–259. [Google Scholar] [CrossRef]
- Maino, J.L.; Schouten, R.; Umina, P. Predicting the global invasion of Drosophila suzukii to improve Australian biosecurity preparedness. J. Appl. Ecol. 2021, 58, 789–800. [Google Scholar] [CrossRef]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Rogers, M.A.; Burkness, E.C.; Hutchison, W.D. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, L.A.; Mendes, M.F.; Kruger, A.P.; Blauth, M.L.; Gottschalk, M.S.; Garcia, F.R. Global potential distribution of Drosophila suzukii (Diptera, Drosophilidae). PLoS ONE 2017, 12, e0174318. [Google Scholar] [CrossRef] [Green Version]
- Chiu, J.C.; Jiang, X.; Zhao, L.; Hamm, C.A.; Cridland, J.M.; Saelao, P.; Hamby, K.A.; Lee, E.K.; Kwok, R.S.; Zhang, G.; et al. Genome of Drosophila suzukii, the spotted wing drosophila. G3 2013, 3, 2257–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ometto, L.; Cestaro, A.; Ramasamy, S.; Grassi, A.; Revadi, S.; Siozios, S.; Moretto, M.; Fontana, P.; Varotto, C.; Pisani, D.; et al. Linking genomics and ecology to investigate the complex evolution of an invasive Drosophila pest. Genome Biol. Evol. 2013, 5, 745–757. [Google Scholar] [CrossRef] [Green Version]
- Paris, M.; Boyer, R.; Jaenichen, R.; Wolf, J.; Karageorgi, M.; Green, J.; Cagnon, M.; Parinello, H.; Estoup, A.; Gautier, M.; et al. Near-chromosome level genome assembly of the fruit pest Drosophila suzukii using long-read sequencing. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Olazcuaga, L.; Loiseau, A.; Parrinello, H.; Paris, M.; Fraimout, A.; Guedot, C.; Diepenbrock, L.M.; Kenis, M.; Zhang, J.P.; Chen, X.; et al. A whole-genome scan for association with invasion success in the fruit fly Drosophila suzukii using contrasts of allele frequencies corrected for population structure. Mol. Biol. Evol. 2020, 37, 2369–2385. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Guo, Z.J.; Liu, Y.; Zhang, Y.J. Progress and Prospects of CRISPR/Cas Systems in Insects and Other Arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Van Eynde, B.; Yu, N.; Ma, S.Y.; Smagghe, G. CRISPR/Cas9 in insects: Applications, best practices and biosafety concerns. J. Insect Physiol. 2017, 98, 245–257. [Google Scholar] [CrossRef]
- Li, F.; Scott, M.J. CRISPR/Cas9-mediated mutagenesis of the white and Sex lethal loci in the invasive pest, Drosophila suzukii. Biochem. Biophys. Res. Commun. 2016, 469, 911–916. [Google Scholar] [CrossRef]
- Li, J.; Handler, A.M. Temperature-dependent sex-reversal by a transformer-2 gene-edited mutation in the spotted wing drosophila, Drosophila suzukii. Sci. Rep. 2017, 7, 12363. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.M.M.; Hildebrand, L.; Wimmer, E.A. Improvement and use of CRISPR/Cas9 to engineer a sperm-marking strain for the invasive fruit pest Drosophila suzukii. BMC Biotechnol. 2019, 19, 85. [Google Scholar] [CrossRef] [Green Version]
- Oberhofer, G.; Ivy, T.; Hay, B.A. Behavior of homing endonuclease gene drives targeting genes required for viability or female fertility with multiplexed guide RNAs. Proc. Natl. Acad. Sci. USA 2018, 115, E9343–E9352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoumani, K.T.; Meccariello, A.; Mathiopoulos, K.D.; Papathanos, P.A. Developing CRISPR-based sex-ratio distorters for the genetic control of fruit fly pests: A how to manual. Arch. Insect Biochem. Physiol. 2020, 103, e21652. [Google Scholar] [CrossRef]
- Kandul, N.P.; Liu, J.; Sanchez, C.H.; Wu, S.L.; Marshall, J.M.; Akbari, O.S. Reply to ‘Concerns about the feasibility of using “precision guided sterile males” to control insects’. Nat. Commun. 2019, 10, 3954. [Google Scholar] [CrossRef] [Green Version]
- Kandul, N.P.; Liu, J.; Sanchez, C.H.M.; Wu, S.L.; Marsha, J.M.; Akbari, O.S. Transforming insect population control with precision guided sterile males with demonstration in flies. Nat. Commun. 2019, 10, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratz, S.J.; Cummings, A.M.; Nguyen, J.N.; Hamm, D.C.; Donohue, L.K.; Harrison, M.M.; Wildonger, J.; O’Connor-Giles, K.M. Genome Engineering of Drosophila with the CRISPR RNA-Guided Cas9 Nuclease. Genetics 2013, 194, 1029–1035. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.L.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.B.; Jiang, W.Y.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aumann, R.A.; Schetelig, M.F.; Häcker, I. Highly efficient genome editing by homology-directed repair using Cas9 protein in Ceratitis capitata. Insect Biochem. Mol. Biol. 2018, 101, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.C.; Green, J.; Surridge, A.; Jiggins, C.D. Evolution of the insect yellow gene family. Mol. Biol. Evol. 2011, 28, 257–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittkopp, P.J.; Vaccaro, K.; Carroll, S.B. Evolution of yellow gene regulation and pigmentation in Drosophila. Curr. Biol. 2002, 12, 1547–1556. [Google Scholar] [CrossRef] [Green Version]
- Arnoult, L.; Su, K.F.Y.; Manoel, D.; Minervino, C.; Magrina, J.; Gompel, N.; Prud’homme, B. Emergence and diversification of fly pigmentation through evolution of a gene regulatory module. Science 2013, 339, 1423–1426. [Google Scholar] [CrossRef]
- Hinaux, H.; Bachem, K.; Battistara, M.; Rossi, M.; Xin, Y.Q.; Jaenichen, R.; Le Poul, Y.; Arnoult, L.; Kobler, J.M.; Kadow, I.C.G.; et al. Revisiting the developmental and cellular role of the pigmentation gene yellow in Drosophila using a tagged allele. Dev. Biol. 2018, 438, 111–123. [Google Scholar] [CrossRef]
- Gokcezade, J.; Sienski, G.; Duchek, P. Efficient CRISPR/Cas9 plasmids for rapid and versatile genome editing in Drosophila. G3 2014, 4, 2279–2282. [Google Scholar] [CrossRef] [Green Version]
- Kondo, S.; Ueda, R. Highly improved gene targeting by germline-specific Cas9 expression in Drosophila. Genetics 2013, 195, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.J.; Sun, J.; Housden, B.E.; Hu, Y.H.; Roesel, C.; Lin, S.L.; Liu, L.P.; Yang, Z.H.; Mao, D.C.; Sun, L.Z.; et al. Optimized gene editing technology for Drosophila melanogaster using germ line-specific Cas9. Proc. Natl. Acad. Sci. USA 2013, 110, 19012–19017. [Google Scholar] [CrossRef] [Green Version]
- Gratz, S.J.; Ukken, F.P.; Rubinstein, C.D.; Thiede, G.; Donohue, L.K.; Cummings, A.M.; O’Connor-Giles, K.M. Highly specific and efficient CRISPR/Cas9-catalyzed homology-directed repair in Drosophila. Genetics 2014, 196, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.Y.; Ren, M.D.; Wu, M.H.; Dai, J.B.; Rong, Y.K.S.; Gao, G.J. Efficient gene knock-out and knock-in with transgenic Cas9 in Drosophila. G3 2014, 4, 925–929. [Google Scholar] [CrossRef] [Green Version]
- Port, F.; Chen, H.M.; Lee, T.; Bullock, S.L. Optimized CRISPR/Cas tools for efficient germline and somatic genome engineering in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, E2967–E2976. [Google Scholar] [CrossRef] [Green Version]
- Akbari, O.S.; Oliver, D.; Eyer, K.; Pai, C.Y. An Entry/Gateway (R) cloning system for general expression of genes with molecular tags in Drosophila melanogaster. BMC Cell Biol. 2009, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwirz, J.; Yan, Y.; Franta, Z.; Schetelig, M.F. Bicistronic expression and differential localization of proteins in insect cells and Drosophila suzukii using picornaviral 2A peptides. Insect Biochem. Mol. Biol. 2020, 119, 103324. [Google Scholar] [CrossRef] [PubMed]
- Kalajdzic, P.; Schetelig, M.F. CRISPR/Cas-mediated gene editing using purified protein in Drosophila suzukii. Entomol. Exp. Appl. 2017, 164, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Ziemek, J.; Schetelig, M.F. CRISPR/Cas9 mediated disruption of the white gene leads to pigmentation deficiency and copulation failure in Drosophila suzukii. J. Insect Physiol. 2020, 126, 104091. [Google Scholar] [CrossRef]
- Karageorgi, M.; Braecker, L.B.; Lebreton, S.; Minervino, C.; Cavey, M.; Siju, K.P.; Kadow, I.C.G.; Gompel, N.; Prud’homme, B. Evolution of multiple sensory systems drives novel egg-laying behavior in the fruit pest Drosophila suzukii. Curr. Biol. 2017, 27, 847–853. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Beldade, P. Development and evolution of insect pigmentation: Genetic mechanisms and the potential consequences of pleiotropy. Semin. Cell Dev. Biol. 2009, 20, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Riedel, F.; Vorkel, D.; Eaton, S. Megalin-dependent Yellow endocytosis restricts melanization in the Drosophila cuticle. Development 2011, 138, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Wright, T.R.; Bewley, G.C.; Sherald, A.F. The genetics of dopa decarboxylase in Drosophila melanogaster. II. Isolation and characterization of dopa-decarboxylase-deficient mutants and their relationship to the alpha-methyl-dopa-hypersensitive mutants. Genetics 1976, 84, 287–310. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.R.; Hodgetts, R.B.; Sherald, A.F. The genetics of dopa decarboxylase in Drosophila melanogaster. I. Isolation and characterization of deficiencies that delete the dopa-decarboxylase-dosage-sensitive region and the alpha-methyl-dopa-hypersensitive locus. Genetics 1976, 84, 267–285. [Google Scholar] [CrossRef]
- Sterkel, M.; Ons, S.; Oliveira, P.L. DOPA decarboxylase is essential for cuticle tanning in Rhodnius prolixus (Hemiptera: Reduviidae), affecting ecdysis, survival and reproduction. Insect Biochem. Mol. Biol. 2019, 108, 24–31. [Google Scholar] [CrossRef]
- Drapeau, M.D.; Radovic, A.; Wittkopp, P.J.; Long, A.D. A gene necessary for normal male courtship, yellow, acts downstream of fruitless in the Drosophila melanogaster larval brain. J. Neurobiol. 2003, 55, 53–72. [Google Scholar] [CrossRef]
- Cobb, M. A gene mutation which changed animal behaviour: Margaret Bastock and the yellow fly. Anim. Behav. 2007, 74, 163–169. [Google Scholar] [CrossRef]
- Sitaraman, D.; Zars, M.; LaFerriere, H.; Chen, Y.C.; Sable-Smith, A.; Kitamoto, T.; Rottinghaus, G.E.; Zars, T. Serotonin is necessary for place memory in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 5579–5584. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.F.; Qiu, S.; Robertson, M. The white gene controls copulation success in Drosophila melanogaster. Sci. Rep. 2017, 7, 1–13. [Google Scholar]
- Dow, M.A. The genetic basis of receptivity of yellow mutant Drosophila melanogaster females. Behav. Genet. 1976, 6, 141–143. [Google Scholar] [CrossRef]
- Pirrotta, V.; Steller, H.; Bozzetti, M.P. Multiple upstream regulatory elements control the expression of the Drosophila white Gene. EMBO J. 1985, 4, 3501–3508. [Google Scholar] [CrossRef]
- Lucchesi, J.C.; Kuroda, M.I. Dosage Compensation in Drosophila. Cold Spring Harb. Perspect. Biol. 2015, 7, a019398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassett, A.R.; Tibbit, C.; Ponting, C.P.; Liu, J.L. Highly efficient targeted mutagenesis of Drosophila with the CRISPR/Cas9 system. Cell Rep. 2013, 4, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Massey, J.H.; Chung, D.; Siwanowicz, I.; Stern, D.L.; Wittkopp, P.J. The yellow gene influences Drosophila male mating success through sex comb melanization. eLife 2019, 8, e49388. [Google Scholar] [CrossRef]
- Meltzer, H.; Marom, E.; Alyagor, I.; Mayseless, O.; Berkun, V.; Segal-Gilboa, N.; Unger, T.; Luginbuhl, D.; Schuldiner, O. Tissue-specific (ts) CRISPR as an efficient strategy for in vivo screening in Drosophila. Nat. Commun. 2019, 10, 2113. [Google Scholar] [CrossRef] [Green Version]
- Port, F.; Strein, C.; Stricker, M.; Rauscher, B.; Heigwer, F.; Zhvou, J.; Beyersdorffer, C.; Frei, J.; Hess, A.; Kern, K.; et al. A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila. Elife 2020, 9, e53865. [Google Scholar] [CrossRef] [PubMed]
- Jaffri, S.A.; Yan, Y.; Scott, M.J.; Schetelig, M.F. Conditional expression systems for Drosophila suzukii pest control. In Drosophila Suzukii Management; Garcia, F.R.M., Ed.; Springer: Cham, Switzerland, 2021; pp. 199–215. [Google Scholar]
- Meccariello, A.; Krsticevic, F.; Colonna, R.; Del Corsano, G.; Fasulo, B.; Papathanos, P.A.; Windbichler, N. Engineered sex ratio distortion by X-shredding in the global agricultural pest Ceratitis capitata. BMC Biol. 2021, 19, 78. [Google Scholar] [CrossRef]
- Champer, J.; Chung, J.; Lee, Y.L.; Liu, C.; Yang, E.; Wen, Z.X.; Clark, A.G.; Messer, P.W. Molecular safeguarding of CRISPR gene drive experiments. Elife 2019, 8, e41439. [Google Scholar] [CrossRef]
- Akbari, O.S.; Bellen, H.J.; Bier, E.; Bullock, S.L.; Burt, A.; Church, G.M.; Cook, K.R.; Duchek, P.; Edwards, O.R.; Esvelt, K.M.; et al. Safeguarding gene drive experiments in the laboratory. Science 2015, 349, 927–929. [Google Scholar] [CrossRef] [Green Version]
- Schetelig, M.F.; Handler, A.M. Strategy for enhanced transgenic strain development for embryonic conditional lethality in Anastrepha suspensa. Proc. Natl. Acad. Sci. USA 2012, 109, 9348–9353. [Google Scholar] [CrossRef] [Green Version]
- Doench, J.G.; Hartenian, E.; Graham, D.B.; Tothova, Z.; Hegde, M.; Smith, I.; Sullender, M.; Ebert, B.L.; Xavier, R.J.; Root, D.E. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nat. Biotechnol. 2014, 32, 1262–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.Q.; Fine, E.J.; Wu, X.B.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Injection Mix a (ng/µL) | Heat-Shock (HS) Treatment b | Injected Embryos | Hatched Larvae c | G0 Adult Survivor c | Mosaic G0 Adults d | G0 Founders e | Inheritance from G0 to G1 f |
|---|---|---|---|---|---|---|---|
| V23 plasmid (500) | No HS | 697 | 16.4% (114/697) | 5.2% (36/697) | 2.7% (1/36) | 3.0% (1/33) | 1.0% |
| V24 plasmid (500) | No HS | 598 | 16.4% (98/598) | 3.9% (23/598) | 4.3% (1/23) | 0.0% (0/14) | 0.0% |
| EGFP-2b (200) | No HS | 546 | 8.1% (44/546) | 2.6% (14/546) | 50.0% (7/14) | 45.5% (5/11) | 39.4% (9.1–98.8%) |
| EGFP-2b (200) | HS (after 1 h) | 557 | 2.6% (12/557) | 0.5% (3/557) | 33.3% (1/3) | 0.0% (0/2) | 0.0% |
| EGFP-2b (200) | HS (after 20 h) | 516 | 18.0% (93/516) | 4.3% (22/516) | 95.4% (21/22) | 85.7% (6/7) | 78.3% (3.6–100%) |
| EGFP-2b (200) | HS (parents) | 509 | 10.0% (51/509) | 4.3% (22/509) | 40.9% (9/22) | 13.3% (2/15) | 92.0% (89.0–95.0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.; Kobayashi, Y.; Huang, C.; Liu, B.; Qian, W.; Wan, F.; Schetelig, M.F. Highly Efficient Temperature Inducible CRISPR-Cas9 Gene Targeting in Drosophila suzukii. Int. J. Mol. Sci. 2021, 22, 6724. https://doi.org/10.3390/ijms22136724
Yan Y, Kobayashi Y, Huang C, Liu B, Qian W, Wan F, Schetelig MF. Highly Efficient Temperature Inducible CRISPR-Cas9 Gene Targeting in Drosophila suzukii. International Journal of Molecular Sciences. 2021; 22(13):6724. https://doi.org/10.3390/ijms22136724
Chicago/Turabian StyleYan, Ying, Yukino Kobayashi, Cong Huang, Bo Liu, Wanqiang Qian, Fanghao Wan, and Marc F. Schetelig. 2021. "Highly Efficient Temperature Inducible CRISPR-Cas9 Gene Targeting in Drosophila suzukii" International Journal of Molecular Sciences 22, no. 13: 6724. https://doi.org/10.3390/ijms22136724
APA StyleYan, Y., Kobayashi, Y., Huang, C., Liu, B., Qian, W., Wan, F., & Schetelig, M. F. (2021). Highly Efficient Temperature Inducible CRISPR-Cas9 Gene Targeting in Drosophila suzukii. International Journal of Molecular Sciences, 22(13), 6724. https://doi.org/10.3390/ijms22136724