1. Introduction
Sea urchins are long-living marine invertebrates (estimated life-span of ~15 years on average for
P. lividus [
1]) that are constantly exposed in nature to pathogens in the marine environment through direct contact with marine substrates, seawater, and consumed food. For protection, they have evolved a very sophisticated and robust innate immune system with multiple effector arms and a wide genome representation of immune genes [
2]. The echinoid immune gene repertoire includes several expanded immune gene families that encode receptors and hemolymph proteins that participate in either cellular responses (e.g., Toll-like receptors, NOD-like receptors, scavenger receptor) or humoral responses (e.g., C-type lectins, complement components such as C3 and factor B) [
3,
4,
5]. Despite the absence of vertebrate-like adaptive immunity, the large numbers of the germline-encoded immune proteins in sea urchins suggest that they may specifically recognize and act against a variety of pathogen-associated molecular patterns (PAMPs). One of the central immune gene families in sea urchin’s immune response encodes the Transformer (Trf) proteins.
The Trf protein family (formerly termed 185/333) was first identified in the California purple sea urchin
Strongylocentrotus purpuratus [
6]. Since then, more than 30 studies about the family in this species were published, and only one study in a different sea urchin species—
Heliocidaris erythrogramma [
7]. To date, no
Trf genes or transcript sequences have been identified outside of the echinoid lineage, suggesting that Trf-based immunity is unique to sea urchins. In
S. purpuratus, a robust Trf-mediated response was documented upon challenge with bacteria including
Escherichia coli and
Vibrio diazotrophicus [
8,
9] and with PAMPs including lipopolysaccharide (LPS), β-1,3-glucan and double-stranded RNA [
10]. The response was characterized by a significant increase in
SpTrf transcript and protein quantities [
8,
10,
11,
12,
13] and in increasing Trf-expressing cell ratios of the whole coelomocytes population [
14,
15,
16]. The SpTrf proteins are capable of binding to subsets of different antigens [
17] and can augment phagocytosis and retard bacterial growth [
18]. A recombinant SpTrf protein undergoes a shape transformation upon binding to antigens [
16]. Consequently, the family name was changed from 185/333 to Transformer.
The
SpTrf genes are 1.2–2 kb long and consist of two exons; a short leader and a second exon which encodes the functional protein [
11]. The structure of the second exon is composed of a mosaic of conserved blocks of sequences, termed elements, that are either present or absent in different members of the family [
11,
12,
19]. The defined combinations of the elements in a second exon are the primary source of diversity in gene sequence and size. However, additional diversification processes occur downstream, including post-transcription [
20] and post-translation [
6], resulting in a diversity of up to 260 different SpTrf proteins per individual as deduced from 2DE Western blot analysis [
8,
13]. Fifteen
SpTrf genes were identified within two loci in the sequenced sea urchin [
21]. However, the
SpTrf loci are predicted to be highly unstable due to gene clustering and tandem sequence repeats [
22,
23] and the
SpTrf gene repertoire shows high variability in the sea urchin population [
21]. Furthermore,
SpTrf genes may be subjected to regulated genomic diversification processes leading to gene deletions, duplications, and single point mutations (SNPs) in single cells [
15] and that each cell may express a unique single
SpTrf gene [
9].
The Trf proteins are intrinsically unstructured in their native form and are predicted to adopt an α helical structure upon binding to a target [
24]. Their predicted molecular weight based on the size of the messages ranged from 4 to 55 kDa in
S. purpuratus [
10] and from 8 to 39 kDa in
Heliocidaris erythrogramma [
7], while the actual protein range as observed on western blots is 30 kDa to >200 kDa in both sea urchin species [
7,
13,
16]. This difference in size is attributed to the multimerization of the Trf proteins [
16]. The primary structure of the Trf proteins consists of a glycine-rich region, a histidine-rich region and a C-terminal region [
11]. The Gly-rich region contains the protein multimerization motif in its C-terminus, which mediates the Trf protein multimerization [
17]. Recombinant peptides of the Gly-rich and His-rich Trf regions have both different and overlapping antigen-binding specificities [
17]. The Trf proteins are expressed in a subset of the phagocytic class of sea urchin coelomocytes in at least two different expression patterns. In small phagocytes, Trf proteins are localized to the cell surface and in cytoplasmatic vesicles, whereas in large polygonal and discoidal phagocytes, the proteins were localized to perinuclear vesicles [
7,
16]. On the other hand, no Trf proteins were identified in the other cell types, including red and colorless spherule cells and vibratile cells [
25]. Immunolocalization of SpTrf proteins in
S. purpuratus histological sections identified cellular SpTrf protein expression in all major sea urchin organs, including axial organ, gut, esophagus, gonad, and pharynx [
14], which is attributed to infiltrated coelomocytes [
25].
Based on multiple studies in
S. purpuratus, a rough model for Trf-mediated immune response was proposed [
18,
25]. According to the model, the Trf protein isoforms that are stored within the perinuclear vesicles of phagocytes are secreted to the coelomic fluid following the detection of bacterial challenge. The secreted Trf protein isoforms multimerize with the same/other isoforms upon binding to bacteria and may adapt an α helical shape. The proteins are also predicted to be involved in bacterial growth retardation [
18]. It is not known how Trf-bound bacteria are recognized by the phagocytes; however, it was suggested that it may be facilitated through either an unknown receptor, multimerization with surface Trf proteins or through phosphatidic acid (PA) located in the plasma membrane. Trf protein binding to PA may cause membrane deformation and therefore may participate in phagocytosis [
26].
To date, the
Trf gene/protein family was characterized into only two sea urchin species—
S. purpuratus and
H. erythrogramma. Here, we characterize the
Trf genes and proteins and the Trf-mediated immune response in a third sea urchin species—the Mediterranean Sea urchin
Paracentrotus lividus.
P. lividus is abundant in shallow subtidal areas and in tidal pools on rocks or seagrass meadows in the Mediterranean Sea and in the North-East Atlantic Ocean [
27]. It is widely used in the food industry (e.g., [
28]) as well as a model in scientific research, including the fields of developmental biology (e.g., [
29]) and comparative immunology (e.g., [
30]).
We report here that, similar to
S. purpuratus and
H. erythrogramma,
P. lividus utilizes a diverse PlTrf protein repertoire as part of its innate immune response to pathogens. The
PlTrf genes and the translated protein sequences, although phylogenetically distinct from
S. purpuratus and
H. erythrogramma, bear the same basic structure as
SpTrf and
HeTrf genes and proteins. In this study, we used three anti-SpTrf antibodies previously raised against the peptides within the N terminus, the central part and the C terminus of the mature Trf proteins [
16]. Using three different approaches, we verified the cross-reactivity of these antibodies with PlTrf proteins. Western blot analyses show high PlTrf protein diversity among different
P. lividus individuals. Furthermore, each of the sea urchins tested expressed a different Trf protein profile in the coelomocytes vs. the cell-free coelomic fluid (CF). Using fluorescence-activated cell sorting (FACS), we identify five different coelomocyte sub-populations with detectable membranal PlTrf expression. We also show that a challenge with heat-killed
E. coli is followed by a sharp increase in the ratios of Trf-positive coelomocytes and that the
P. lividus response to
E. coli challenge is likely mediated by a specific set of PlTrf proteins. Lastly, we demonstrate that
P. lividus cell-free CF is important for phagocytosis of
E. coli, a process that can be inhibited by blocking membranal PlTrf function with the anti-Trf antibodies.
3. Discussion
Immune systems are one of the most diverse systems in multicellular organisms, both among species as well as within the population of a species and even among different cells of an individual. This is especially true for invertebrates that use many different ways to achieve protection against a great variety of pathogens in their surrounding environment (e.g., [
39]). The Trfs are an example of unique proteins that participate in the effector arm of the innate immune response in sea urchins. To date, this family has been studied in
S. purpuratus [
25] as well as in
H. erythrogramma [
7]. Here, we characterize a family of Trf genes and proteins in the Mediterranean Sea urchin
P. lividus, an echinoid model organism. Our results suggest that despite the differences among PlTrf, SpTrf, and HeTrf gene and protein sequences, the basic features and the mechanisms of action appear to be conserved. The PlTrf protein sequences vary in size and include typical Trf regions such as the characteristic hydrophobic leader, glycine-rich and histidine-rich regions, identifiable element patterns, and repeats that were characterized for the two other species. On the other hand, the RGD motif, which is located in the multimerization region, is absent from both
P. lividus and
H. erythrogramma and therefore may be unique to
S. purpuratus and may not have a role in multimerization. To study the Trf-mediated response in
P. lividus, we established a strong molecular tool, which is sometimes a challenging task in non-classical invertebrate models. We invested resources to validate the cross-reactivity of a potent anti-SpTrf antibody mix that was developed by the Smith group against conserved areas of the SpTrf proteins [
16]. We used the validated antibody mix to characterize the PlTrf protein repertoire in cells and CF of different individual sea urchins to detect PlTrf-expressing coelomocytes, FACS analysis, and for the blocking the cell surface PlTrf activity in functional phagocytosis assays.
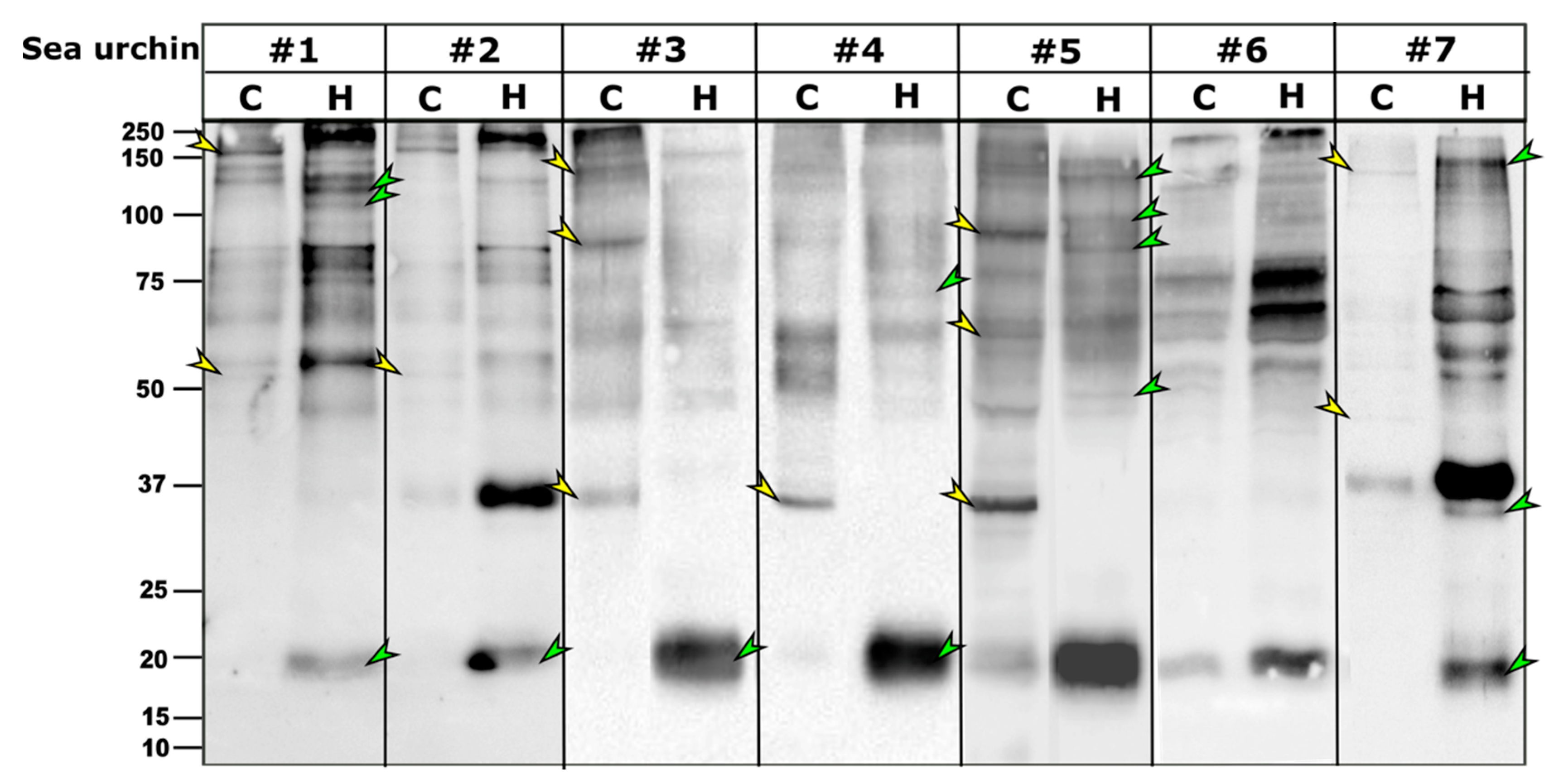
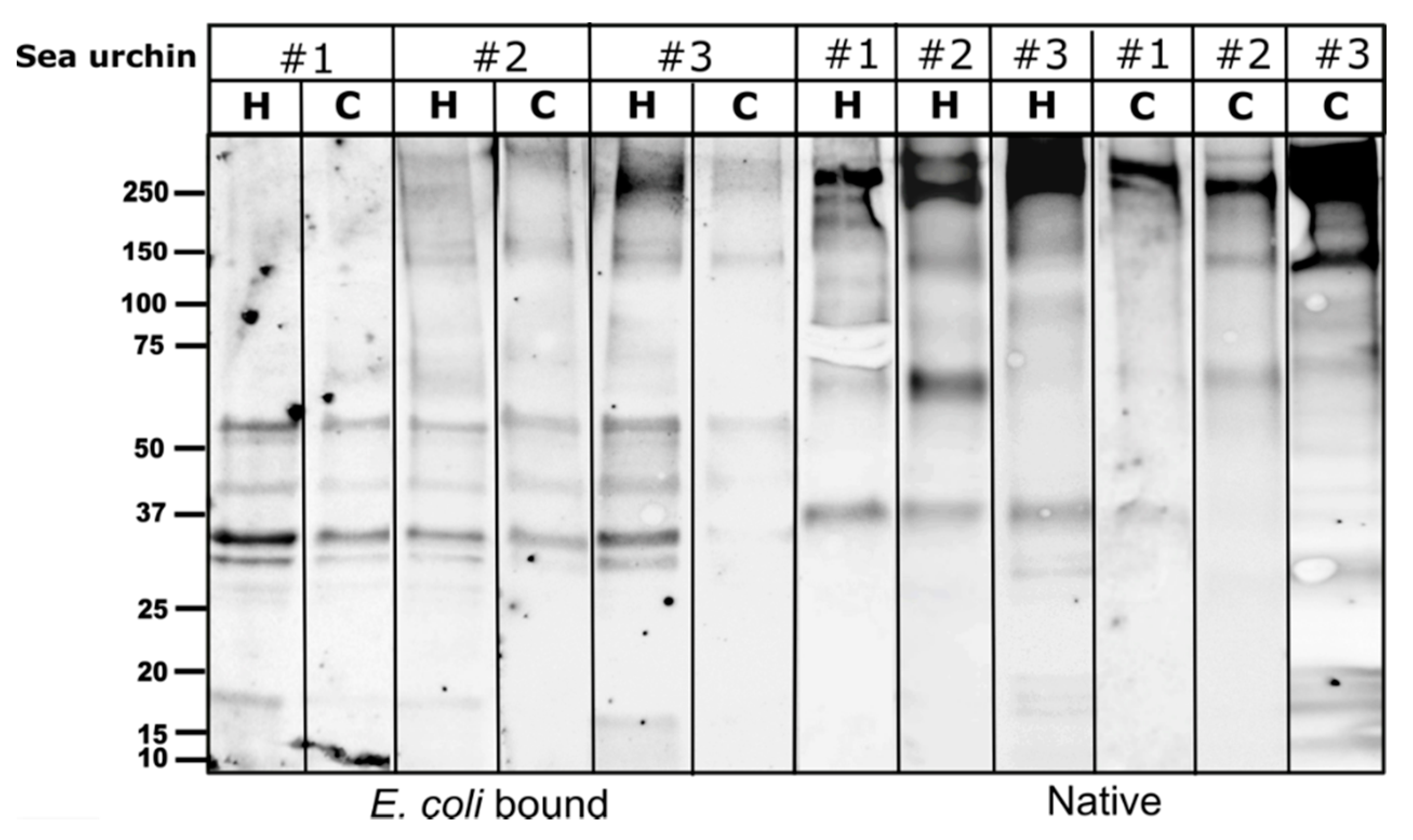
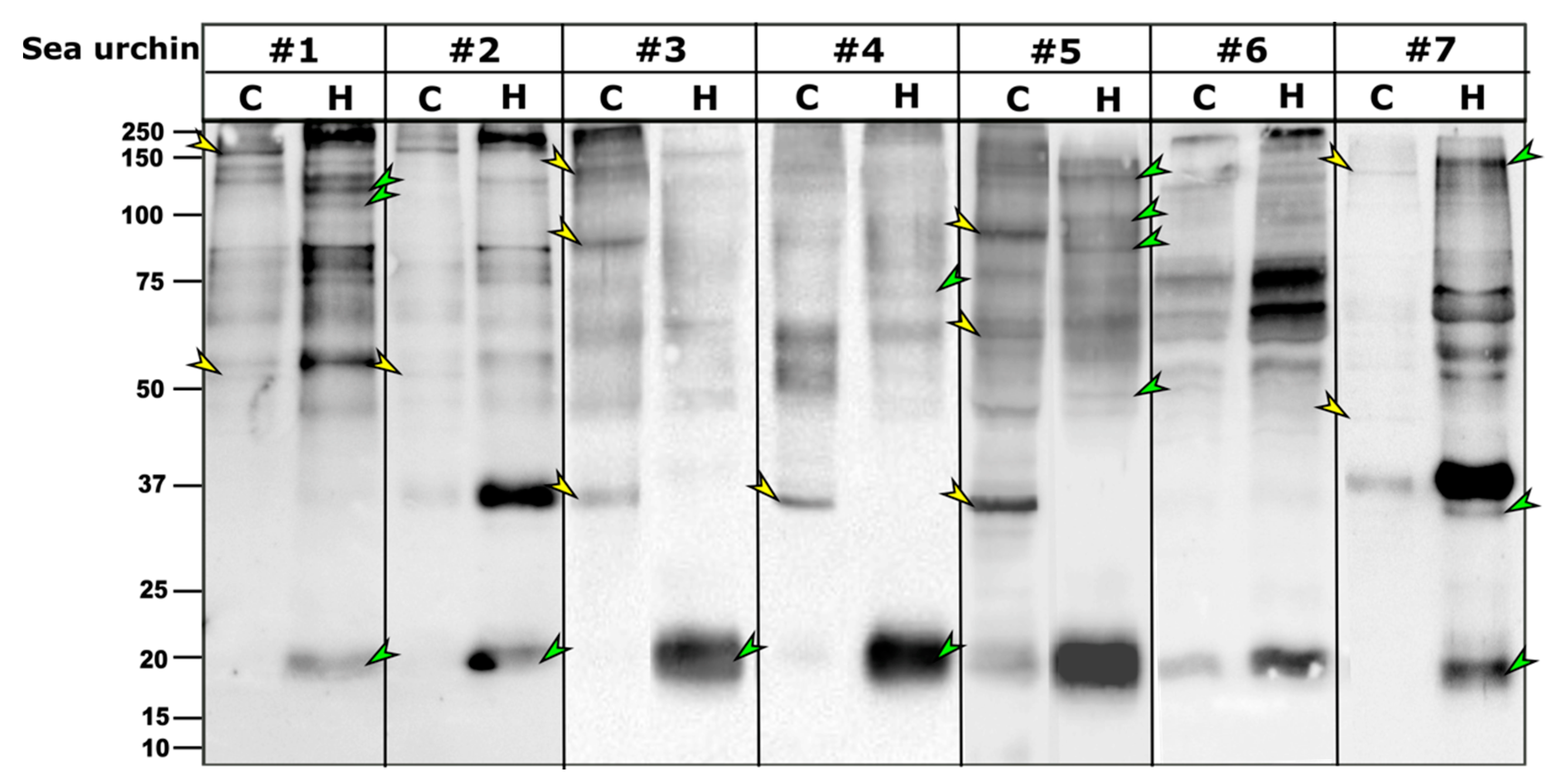
As in other sea urchin species, the PlTrf proteins vary among individuals, although they often share subsets of proteins of the same molecular weight. It was previously hypothesized that some SpTrf proteins multimerize through the glycine-rich multimerization region but that only monomers bind, lyse, or alter the shape of liposomes containing phosphatidic acid [
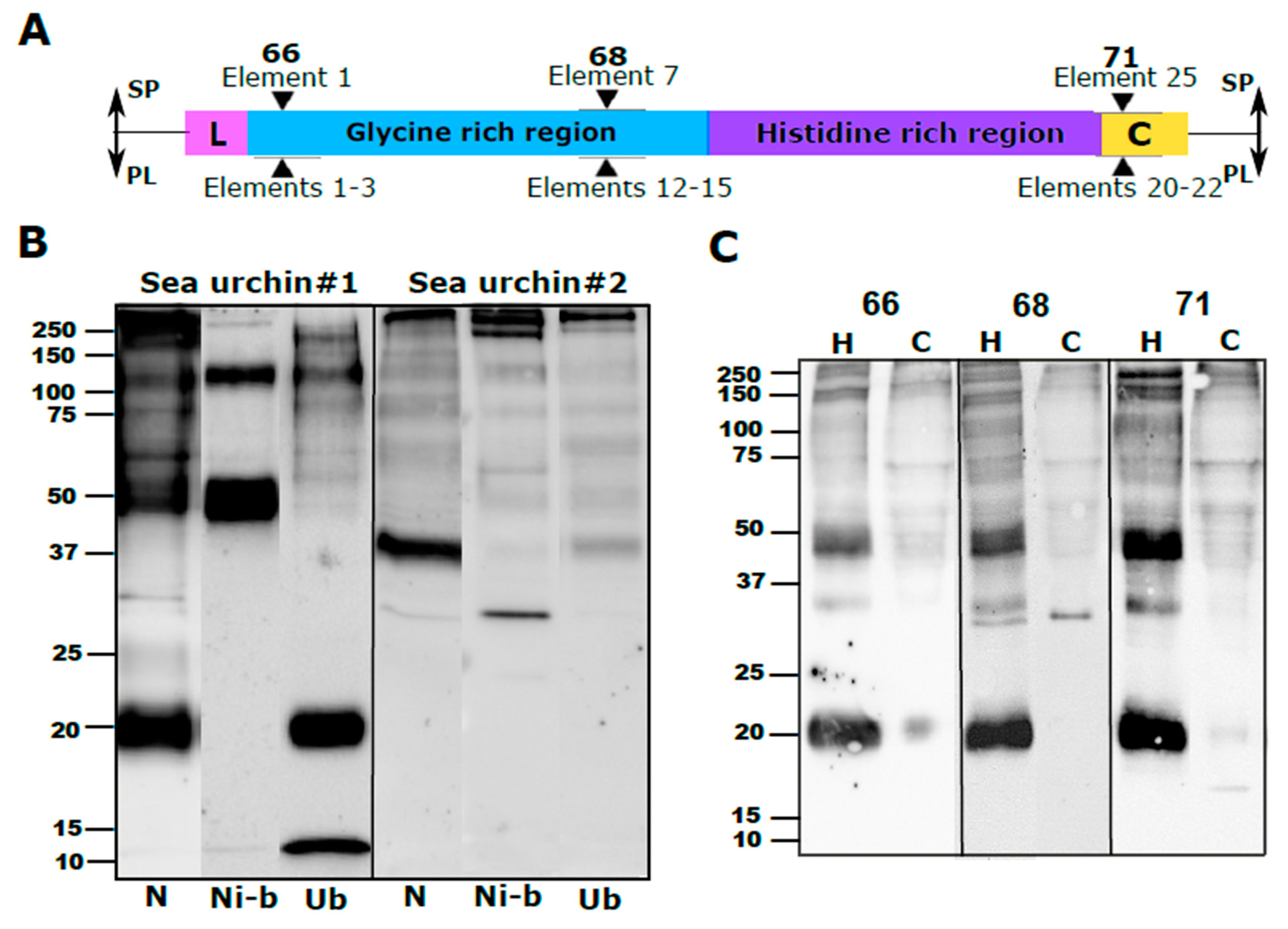
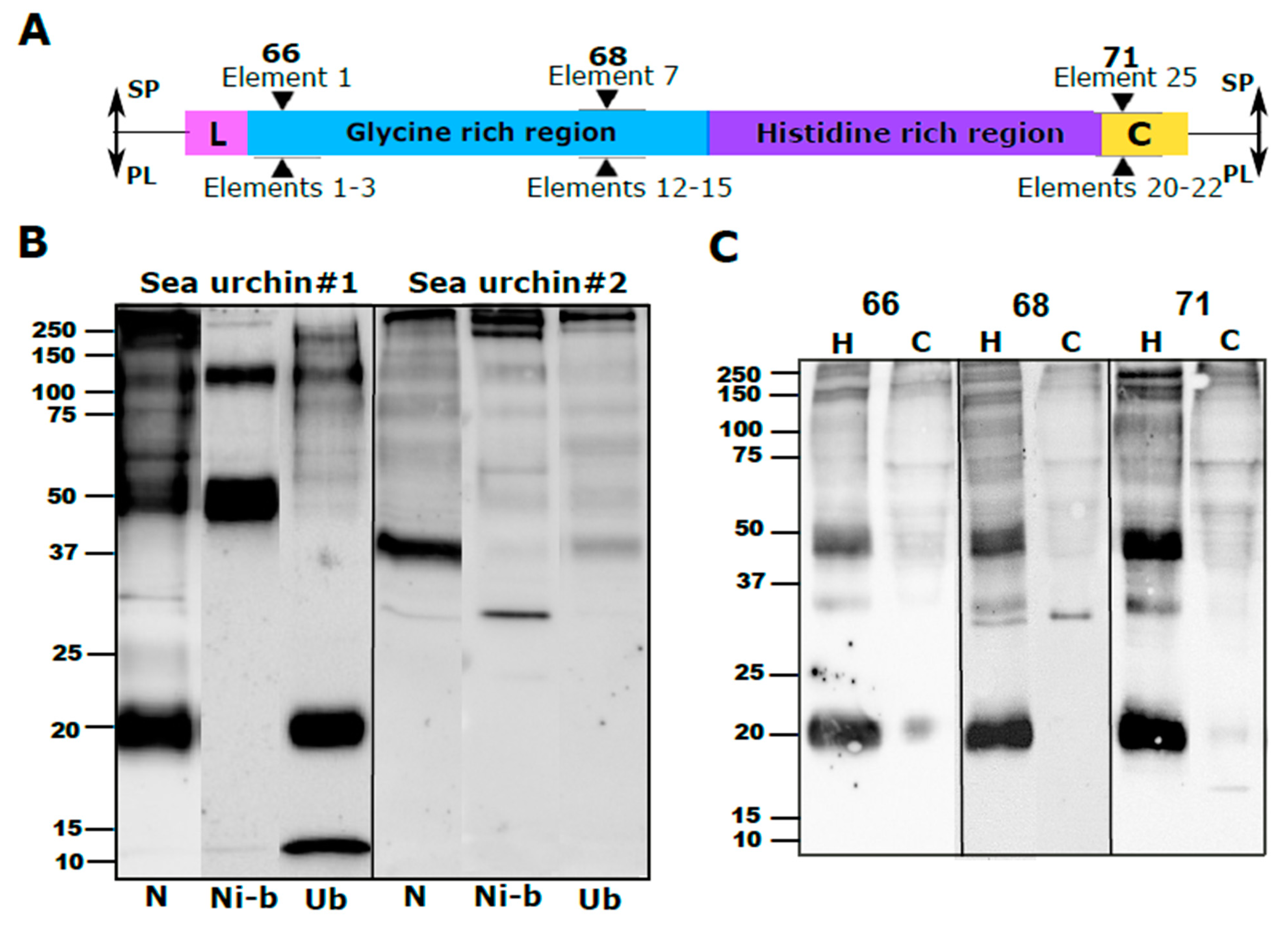
26]. The PlTrf protein multimers vary greatly in size, suggesting several levels of multimerization. The Trf protein multimerization is irreversible and stable at high temperature in 2-mercaptoethanol, and SDS treatments, which is why multimers are observed on WBs. We identified two size categories of PlTrf protein monomers of approximately 20 and 34 kDa that are similar to the sizes of the SpTrf and HeTrf protein monomers. While the ~34 kDa monomer size was predicted based on the CDS sequences, the ~20 kDa monomer could not be predicted based on the available full-length primary sequences and may represent truncated proteins. In
S. purpuratus, truncated Trf sequences that lack the histidine-rich region may serve as potential surveillance proteins [
8]. This may also the case with the PlTrf as supported by the Ni-column isolation in which the ~20 kDa proteins were found in the unbound fraction from one of the sea urchins (
Figure 2A).
Recent reports on the function of the SpTrf proteins suggest that they act through both cellular and humoral effector arms to augment phagocytosis [
18,
26]. Here, we identified significant differences between the PlTrf protein repertoires associated with the coelomocytes (cellular) compared to those in the cell-free CF (humoral) fraction of the coelomic fluid in each of the tested sea urchins. Differences are observed in the monomer and the multimer content of the two fractions. The smaller PlTrf monomers of ~20 kDa, which are consistently present in the cell-free CF, are either absent or almost absent from coelomocytes. On the other hand, the ~35 kDa monomers were present in either coelomocytes, cell-free CF, or both, depending on the sea urchin. Significant variability was observed in the PlTrf multimer content in all tested sea urchins. To better understand the functions of each of these Trf-mediated effector routes and the possible interplay between them, we studied each of the two fractions separately and tested how the absence or inhibition of the fractions affects the phagocytosis process.
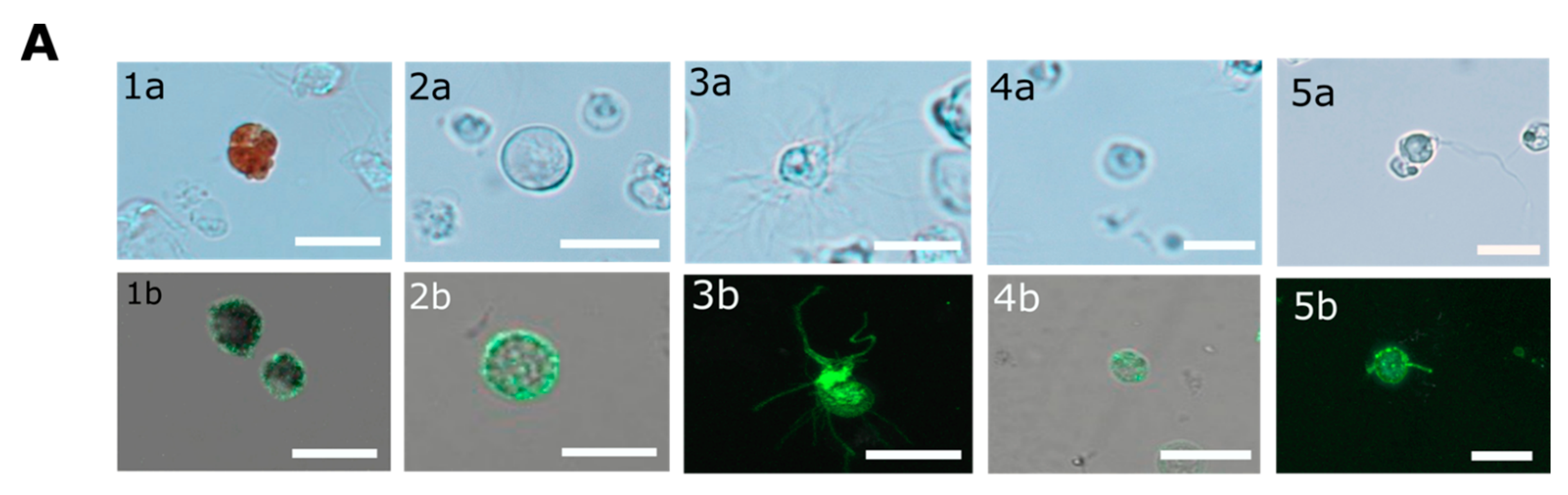
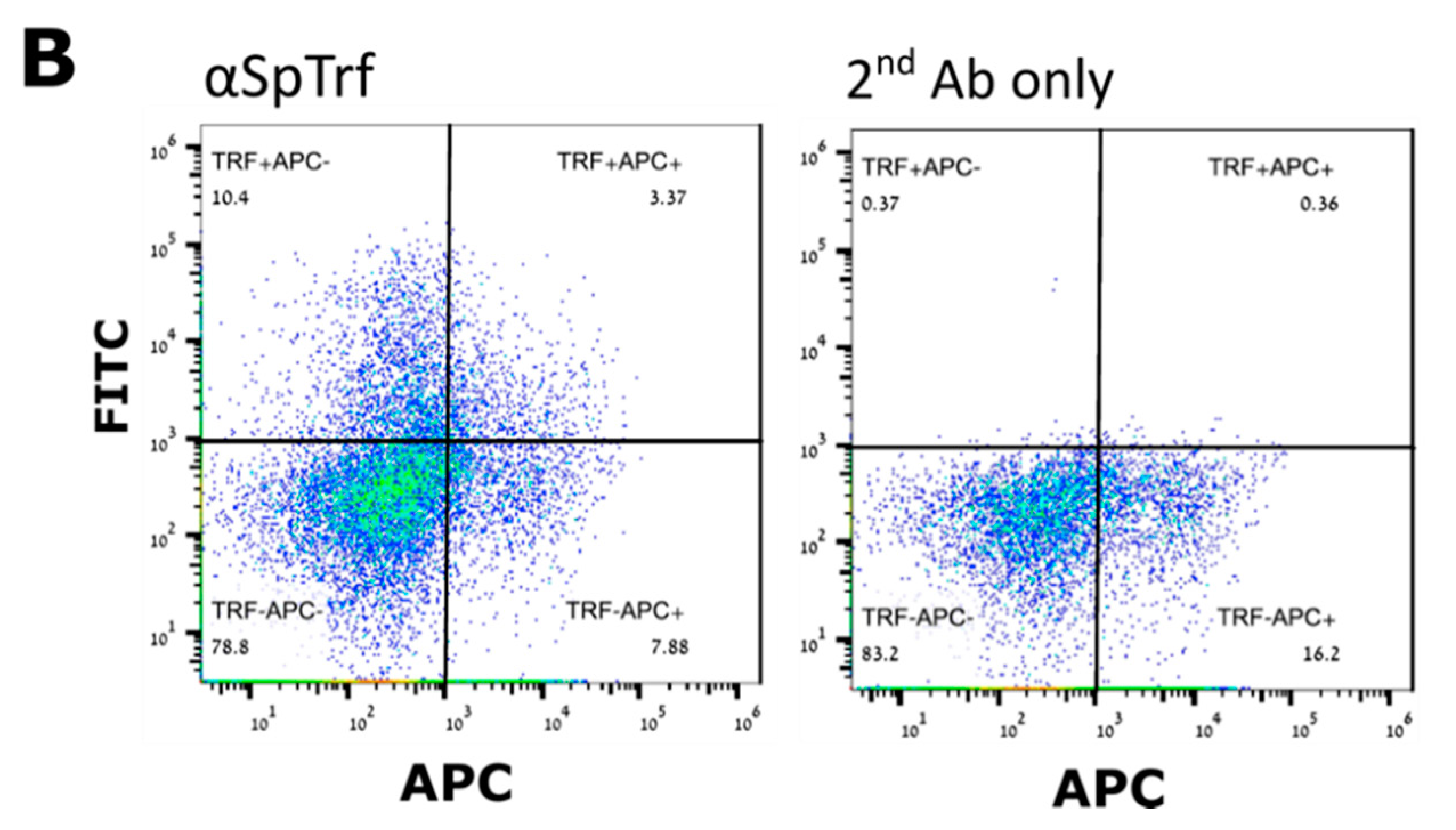
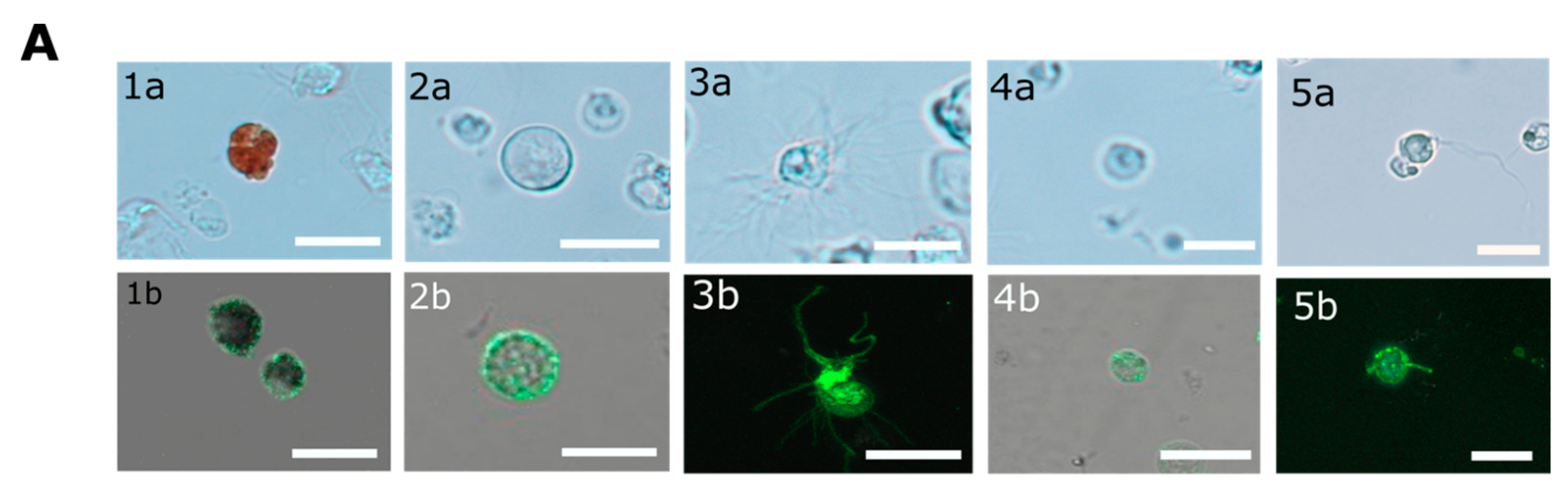
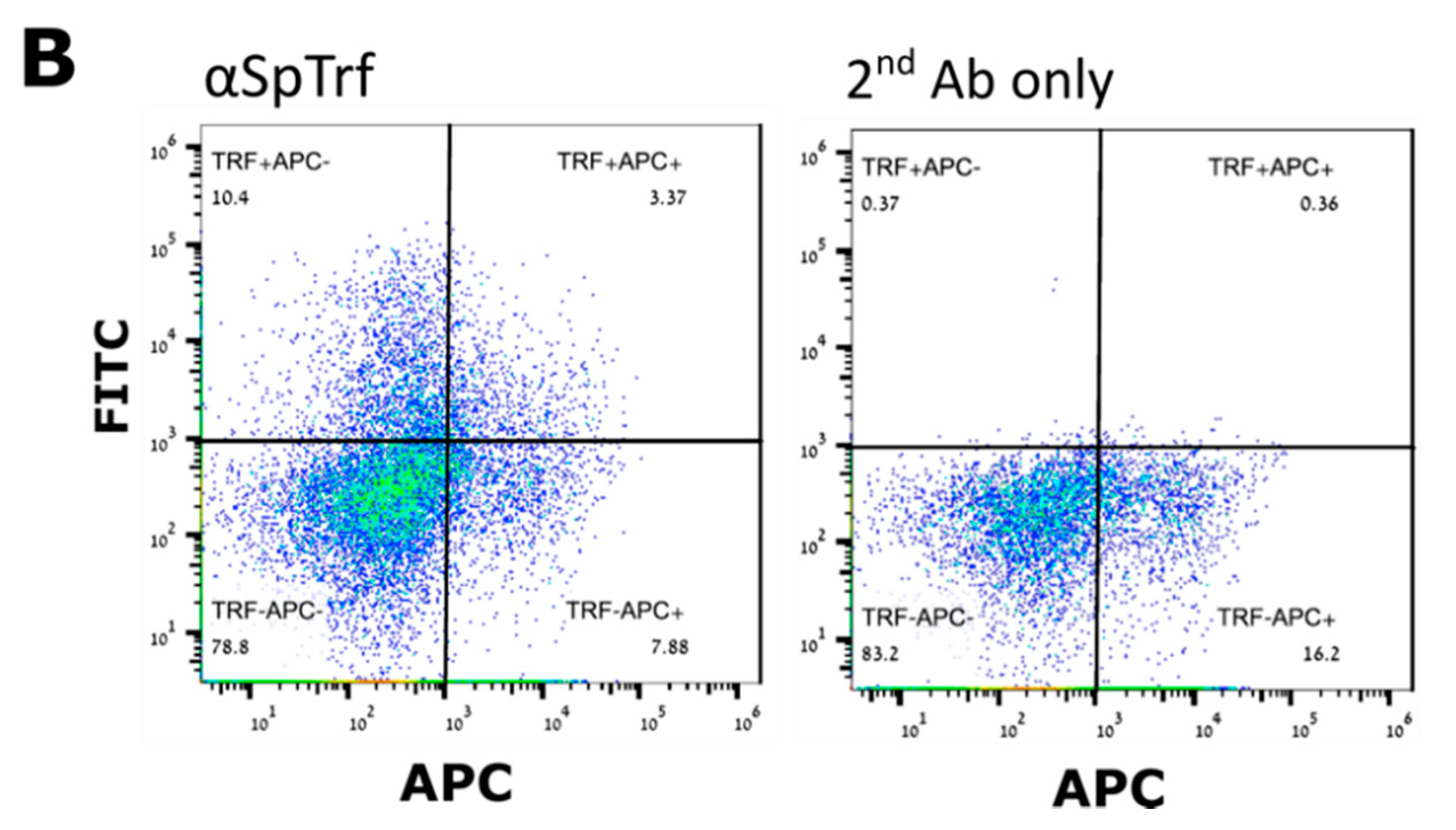
In contrary to the cell surface localization of Trf proteins, which was restricted to small phagocytes (petaloid coelomocytes) in
S. purpuratus [
25], we identified at least five morphological subpopulations of coelomocytes with cell surface-bound PlTrf proteins. These differences between
S. purpuratus and
P. lividus may be due to differences in Trf activity between the species. However, they may also be due to the use of FACS to sort live coelomocytes, which enabled sensitive detection prior to imaging. An increase in membranal Trf-expressing coelomocyte percentages was recorded in
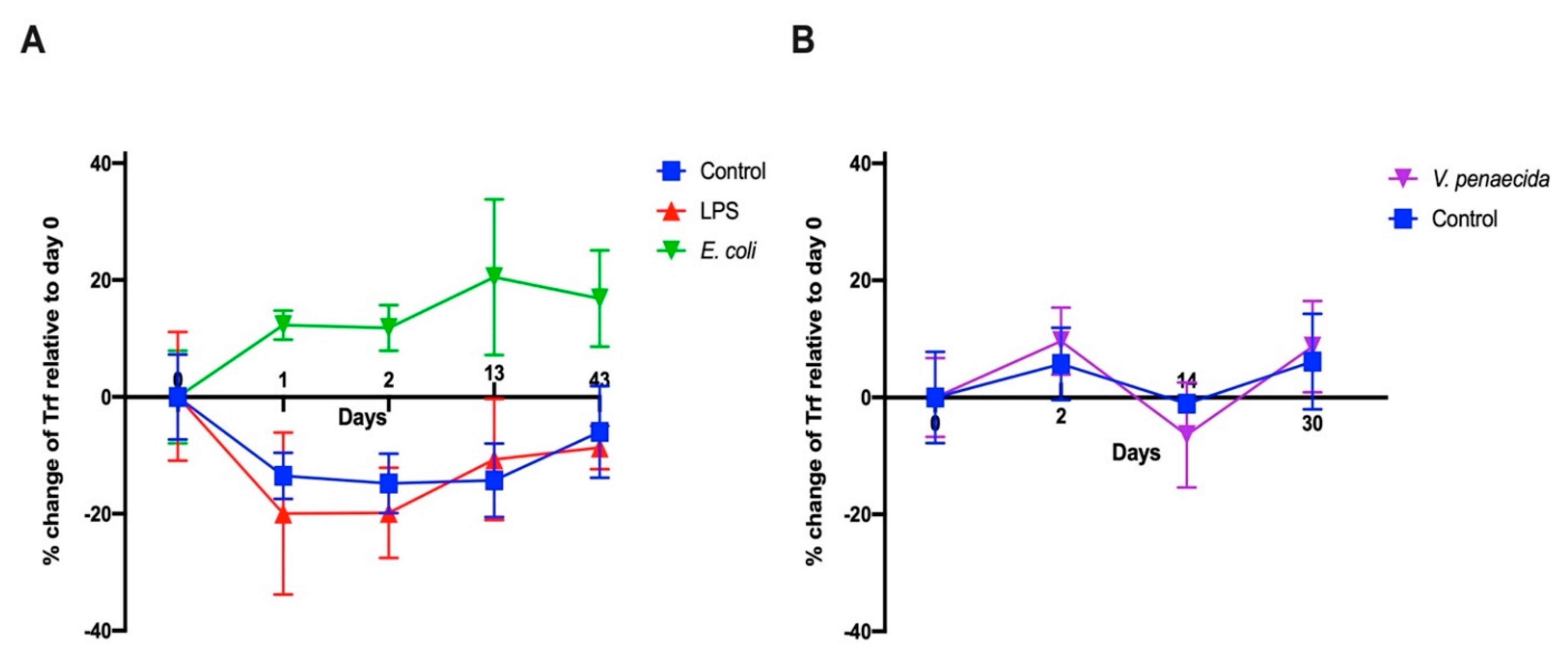
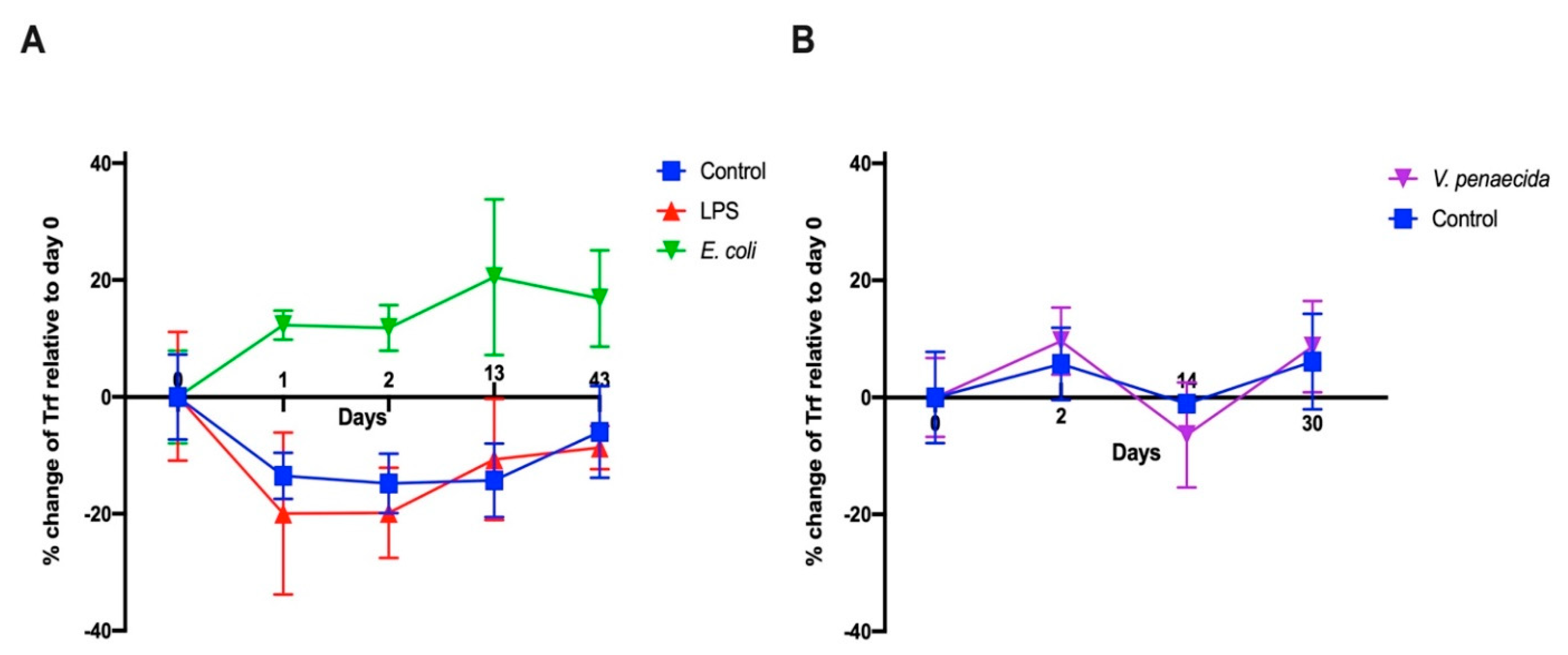
S. purpuratus in response to LPS [
16] and
V. diazotrophicus [
9,
15]. We followed a similar experimental design to test first the effect of LPS and heat-killed
E. coli. We were surprised to see that the results obtained were different from
S. purpurtus. LPS does not influence the percentage of PlTrf-expressing coelomocytes, whereas there is a sharp increase in PlTrf-expressing cells in response to
E. coli. This suggests that LPS might not be the main target of the PlTrf proteins but rather different molecules on the
E. coli surface. We also used heat-killed
V. penaeicida, a putative
P. lividus pathogen, which is similar to
V. diazotrophicus, that was used to challenge
S. purpurtus. Unexpectedly,
V. penaeicida does not have an effect on the PlTrf-expressing coelomocytes, suggesting that this species may escape the surveillance by the PlTrf proteins.
Specificity is a key feature in immune systems because it enables optimal recognition of pathogens within a variable environment and prevents self-destruction through autoimmunity [
40]. Although this attribute is classically linked with adaptive immunity, a high degree of specificity may characterize certain types of innate immune systems and innate immune receptors. The great protein diversity of the Trf family within sea urchin populations [
13] and the unique single-gene expression in individual coelomocytes [
9] allow a high degree of specificity. Indeed, SpTrf specificity was demonstrated in the specific gene-expression response to different elicitors [
8,
9,
10,
18], the semi-specific binding of a recombinant SpTrf protein [
17] and the selective PlTrf-mediated cellular response to different types of immune challenges, as shown here. By incubating live
E. coli with coelomocytes or cell-free CF, we profiled the specific
E. coli bound PlTrf protein profile. The profiles included the same specific sets of PlTrf proteins, regardless of the genotype or the coelomic fluid fraction from which they were identified.
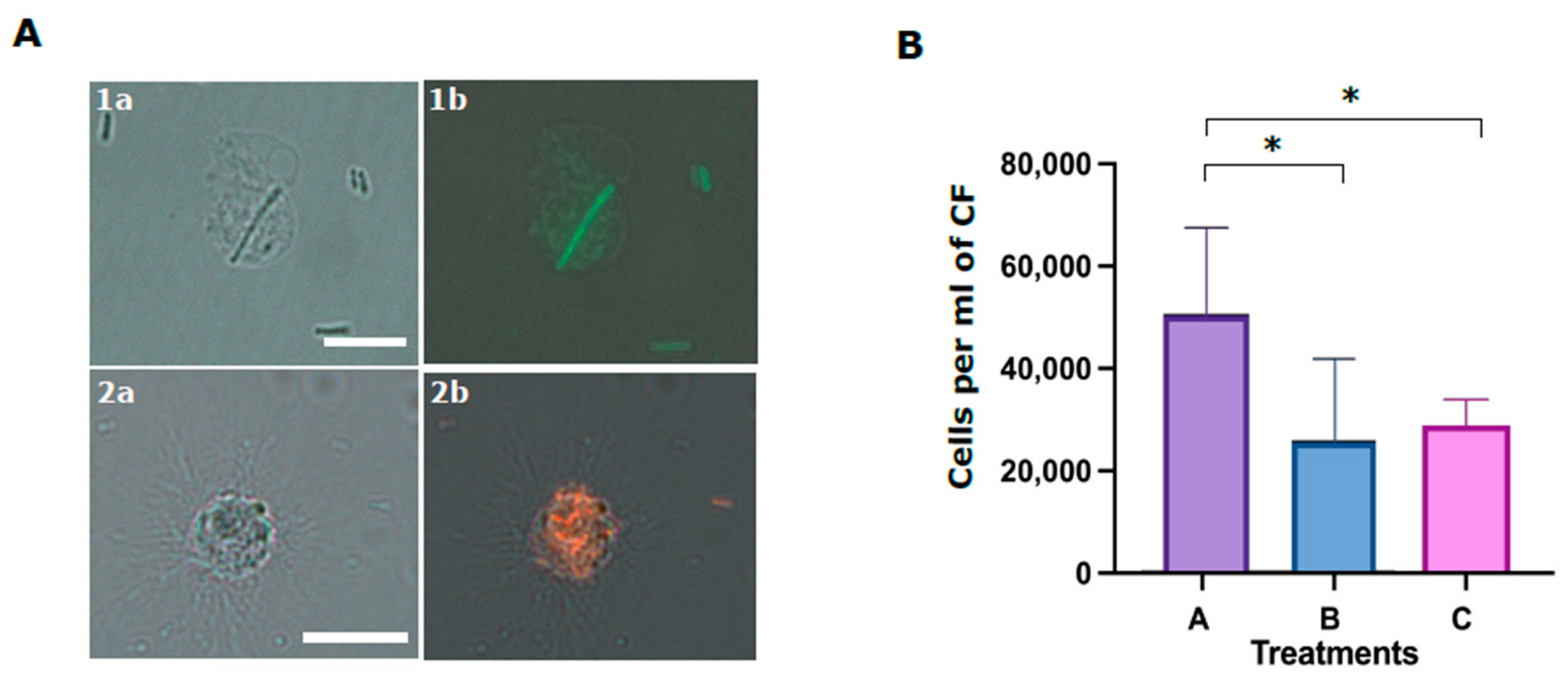
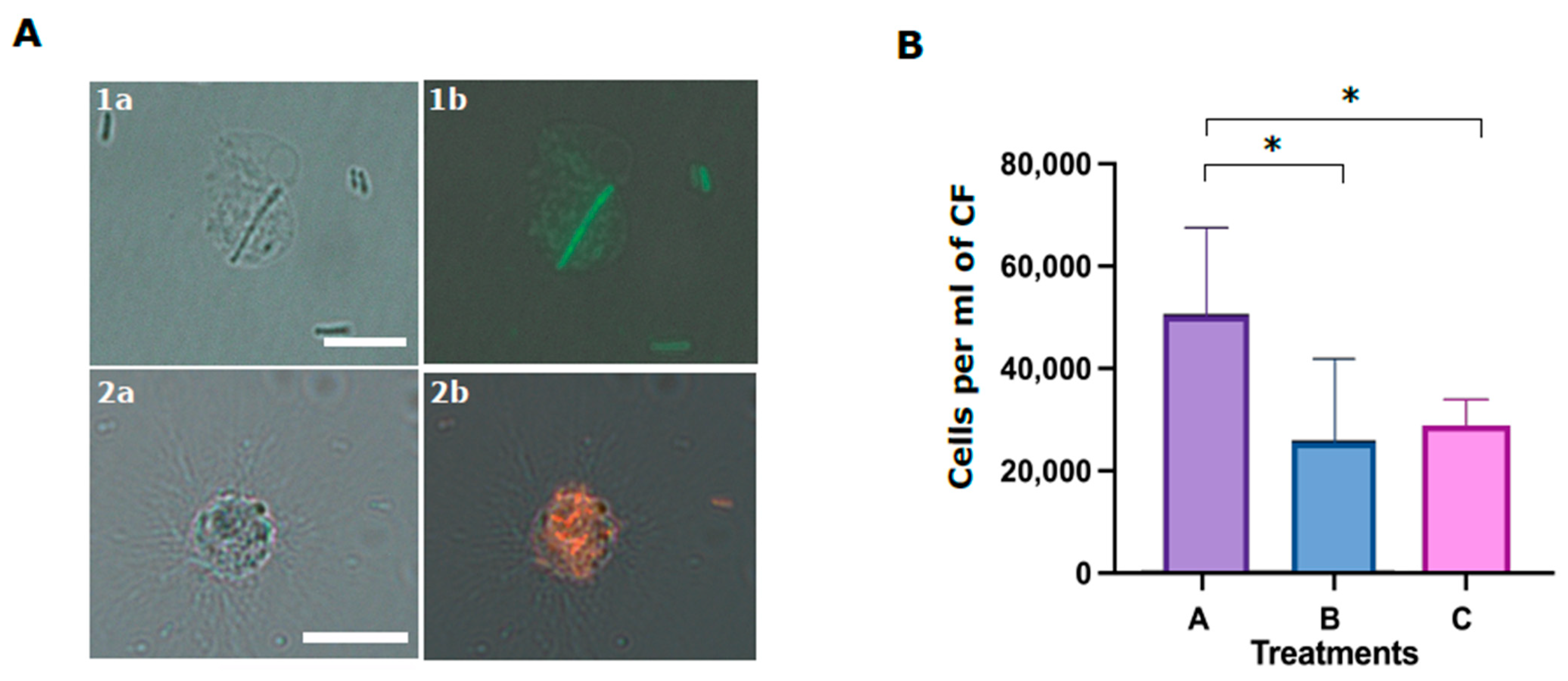
In the last experiment, we studied the role of the Trf-mediated cellular and humoral fractions in the phagocytosis of
E. coli. Although it was demonstrated that SpTrf proteins augment phagocytosis [
18], the function of the membranal SpTrf proteins in this process was not addressed. We found that although the coelomocytes alone (without CF) are sufficient for phagocytosis, the absence of CF (humoral fraction) reduces phagocytosis efficiency. We also used the anti-SpTrf antibody mix to block the cell surface PlTrf proteins to determine whether this affected phagocytosis efficiency. Our results demonstrate that the inhibition of cell-surface PlTrf activity results in significant inhibition of phagocytosis. Following both results, we hypothesis that the membrane-bound cellular PlTrfs interact with bacteria-bound humoral PlTrf as part of the phagocytosis process.
Taken together, this study characterizes a diverse PlTrf gene/protein family in the Mediterranean Sea urchin P. lividus. It demonstrates the collaboration of distinct cellular and humoral PlTrf-mediated effector arms in mounting specific P. lividus immune responses, opsonization of bacteria, and phagocytosis.
4. Materials and Methods
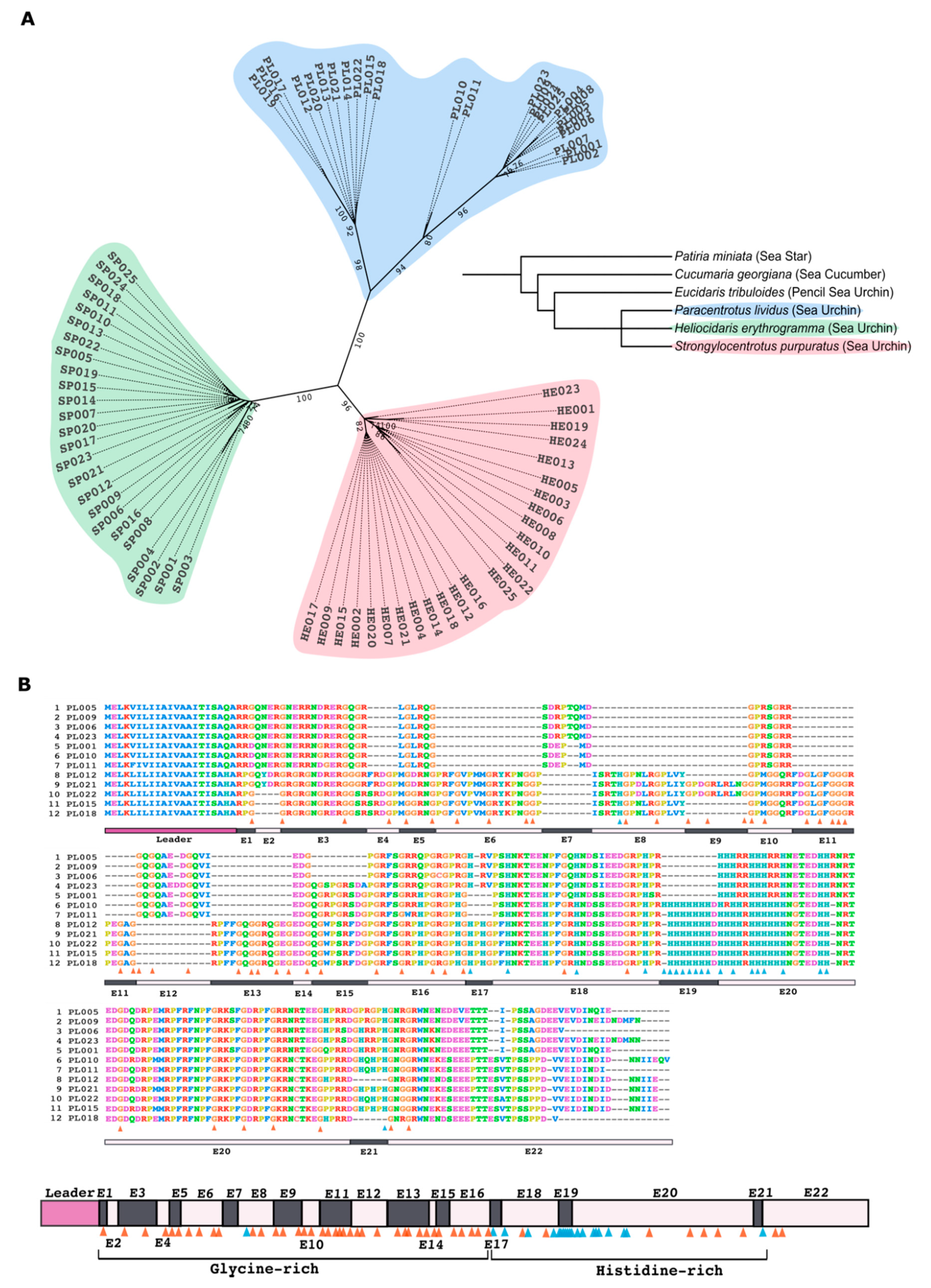
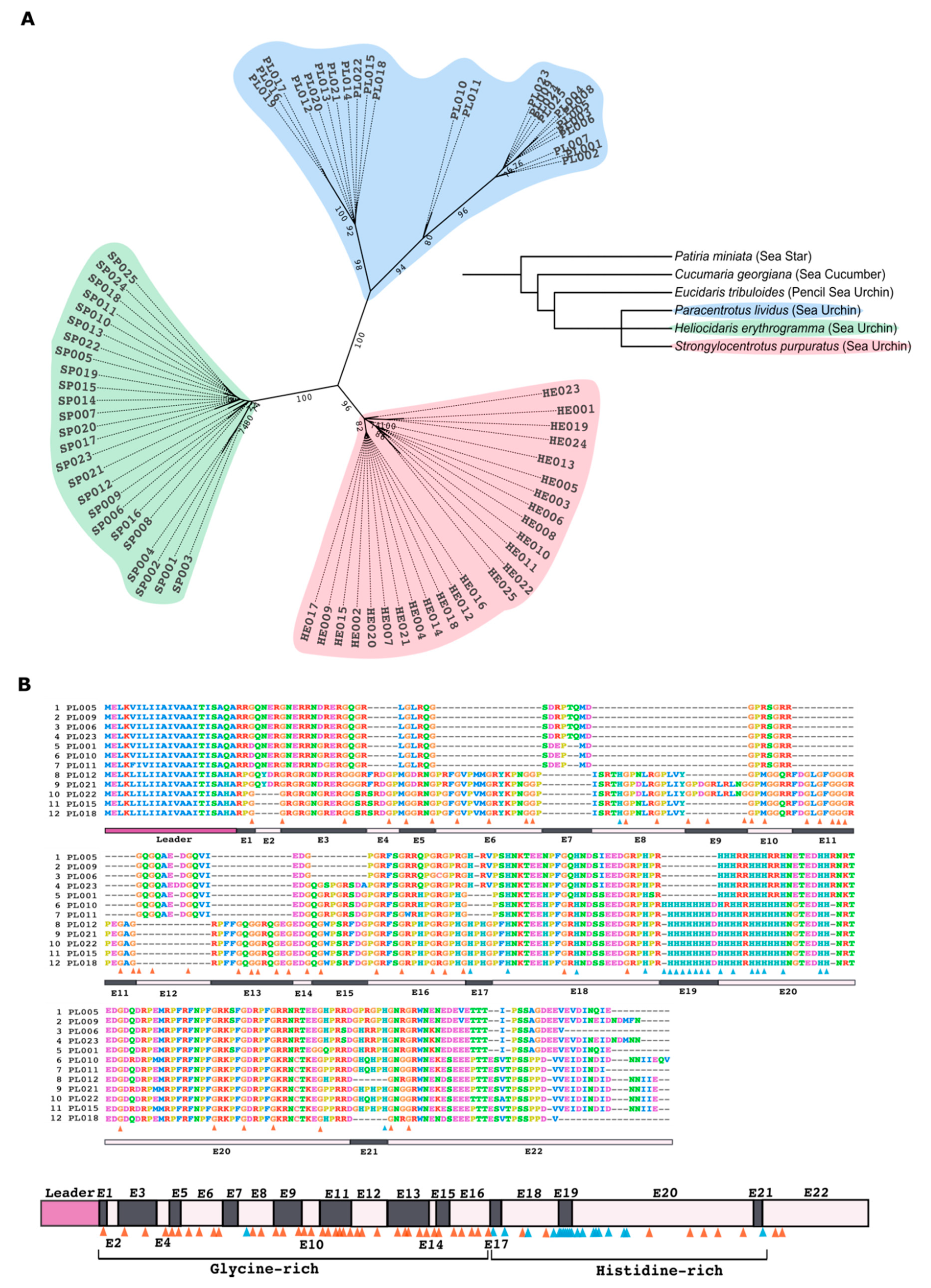
4.1. Alignment and Phylogenetic Analyses of Trf Sequences
The search for
PlTrf sequences was performed in four TSA databases: HACU01, GCZS01, GEDS01, GFRN01 and in the Octopus database. Available online on request:
http://octopus.obs-vlfr.fr/blast/oursin/blast_oursin.php (accessed on 15 March 2021).
S. purpuratus and
H. erythrogramma Trf sequences were compared to the databases using the local BLAST algorithm. Twenty-five sequences from each of the three species were aligned using ClustalW multiple alignments with default parameters and further manual refinement using MEGA X [
33]. Phylogenetic analyses of PlTrf sequences were performed using Maximum Parsimony, Neighbor-joining, and Maximum Likelihood analyses with bootstrapping of 1000 repetitions. The best algorithm combination was inferred by MEGAX as Maximum Likelihood with Hasegawa–Kishino–Yano model [
32]. Initial trees for the heuristic search were obtained by applying the Neighbor-Joining method to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach. A discrete Gamma distribution was used to model evolutionary rate differences among sites (3 categories (+
G, parameter = 2.7099)). The tree was visualized as unrooted with the bootstrap value displayed in ITOL program [
41]. Twelve translated
PlTrf sequences were chosen based on their unique aa sequence to generate alignments and to represent all available element patterns.
4.2. Sea Urchins
Sea urchins were obtained from the Israeli National Center for Mariculture in Eilat. Urchins were kept in artificial seawater (Red Sea Fish Pharm Ltd., Herzliya, Israel) in a 165-L aquarium in the Molecular Ecology Laboratory at the Ariel University, of which 1/10 volume was replaced every week. The water salinity was kept around 40 ppt and the temperature between 20 and 22 °C. The animals were fed once a week with either fresh or frozen Ulva lactuca algae. Unhealthy-looking or dying sea urchins are isolated in a different aquarium to avoid contamination of healthy individuals. Experimental animals were isolated in individual floating plastic cages in a separate compartment in the aquarium.
4.3. Bacteria Cultures
Escherichia coli (25,922 strain) and mCherry-expressing
E. coli were kindly provided by Dr. Shiri Navon-Venezia and Ms. Helena Tuchinsky,
Vibrio penaeicida (51,842 strain) were obtained from ATCC bioresource center (Manassas, VA, USA), commercial lipopolysaccharide (LPS) from
E. coli was purchased from Sigma-Aldrich (Israel).
E. coli were cultured in liquid Luria Bertani (LB) medium overnight at 37 °C.
V. penaeicida bacteria were cultured in Marine Broth media (MB) from Sigma-Aldrich (Israel) overnight at 26 °C. The concentration of bacteria was either calculated based on OD values measured in a Nanodrop NP80 spectrophotometer from Implen (Westlake Village, CA, USA) or manually counted with a hemocytometer. All bacteria were heat-killed for 5 min at 95 °C, washed, and resuspended in aCF (10 mM CaCl
2, 14 mM KCl, 50 mM MgCl
2, 398 mM NaCl, 1.7 mM Na
2HCO
3, 25 mM Na
2SO
4 [
12]) and used immediately in experiments.
4.4. Preparation of Coelomocytes and Cell-Free CF Samples
CF (~300 μL) was collected from the coelomic cavity of the sea urchin with a 23 G needle and a 1 mL syringe filled with an equal volume of ice-cold calcium-magnesium free seawater (CMFSW-EH; 460 mM NaCl, 10.73 mM KCl, 7.06 mM Na2SO4, 2.38 mM NaHCO3, 70 mM EDTA, 20 mM HEPES; pH 7.4; Terwilliger et al. 2006) and kept on ice until processing. The CF was centrifuged at 500× g at 4 °C for 5 min. The supernatant was carefully collected without disturbing the cell pellet and transferred to a separate tube. The cells were carefully resuspended and washed in CMFSW-EH. The supernatant was centrifuged again at 2000× g at 4 °C for 5 min to obtain the cell-free CF. The cell-free CF and the washed coelomocytes were used separately for WB and phagocytosis assays.
4.5. Protein Extraction
Whole CF, cell-free CF, and washed coelomocytes were lysed using RIPA buffer (Thermo Fisher Scientific, Waltham, MA, USA). The lysates were incubated for 30 min at room temperature with protease inhibitor cocktail III (Calbiochem) with constant vortexing. Alternatively, cells were sonicated for 5 min. Cell debris was pelleted by centrifugation for 10 min at 10,000× g at 4 °C. The supernatant was stored in aliquots at −80 °C until use. To equalize the protein quantities before loading on the protein gels, protein concentrations were measured using the Bradford assay and with an NP80 spectrophotometer (Implen, Westlake Village, CA, USA).
4.6. His60 Ni-Column Binding Assay
The His60 Ni-column binding assay was performed according to [
8]. The whole CF was mixed with 2 mL of lysis buffer (50 mM NaH
2PO
4, 300 mM NaCl, 10 mM imidazole, adjusted pH to 8.0 with NaOH, supplemented with 2 mg lysozyme). The mixture was sonicated for 5 min and incubated at room temperature for 10 min. The lysate was centrifuged for 30 min at 10,000×
g at 4 °C, and the supernatant was collected. His60 Ni Superflow Resin (2 mL; Takara Bio, Japan) was loaded into a gravity column, followed by 10 min incubation with 4 mL lysis buffer. The lysate supernatant was added to the column and incubated with resin for 1 h at 4 °C with gentle shaking after which the flow-through was discarded. Unbound proteins were collected with 4 volumes of lysis buffer. Ni-bound proteins were eluted with 5 volumes of elution buffer (50 mM NaH
2PO
4, 300 mM NaCl, 500 mM imidazole, water, adjusted pH to 8.0 with NaOH). The flow-through and eluted proteins were used for subsequent analysis by WB and MS.
4.7. Western Blots
Lysate samples with equal quantities of proteins were mixed with 4× Laemmli sample buffer (Bio-Rad), heated at 90 °C for 5 min, and 10–30 μg of protein was loaded per lane on 10% or 12% SDS-PAGE gels. Gel electrophoresis was performed in a fresh running buffer (25 mM Tris, 192 mM glycine, 0.1% SDS, pH 8.3). Gels were subjected to semi-dry transfer with Trans-Blot Turbo™ System (Bio-Rad) onto PVDF membranes (Bio-Rad) with the preprogrammed Bio-Rad 1.5 mm gel transfer program. The membranes were incubated for 1 h in 5% blocking solution (BSA-TBST; 5% bovine serum albumin in 20 mM Tris, 150 mM NaCl, 0.1% Tween20). The blocking solution was replaced with 2.5% BSA-TBST solution containing 1:10,000 dilution of primary Rabbit anti-SpTrf antibodies mixture and gently agitated overnight at 4 °C. The membranes were washed 5 times for 5 min each with TBST and then soaked for 1 h with gentle shaking in TBST containing 1:20,000 dilution of HRP labeled Goat anti-Rabbit-Ig antibody (Abcam, Cambridge, UK). Images were obtained by Quantity One or with ChemiDoc imaging systems (Bio-Rad, Haifa, Israel) and processed with ImageJ2 (FIJI) [
42] or the Image Lab 6.1 (Bio-Rad, Hercules, CA, USA) software.
4.8. Mass Spectrometry
The eluted and washed protein fractions from the nickel affinity column were separated on an SDS-PAGE gel in a duplicate loading series. The gel was cut according to the duplicated lanes. One gel half was used to identify the band sizes by WB. The other gel half was used to cut gel slices that corresponded to high molecular Trf-positive bands that were identified in the WB. An empty gel slice from the gel corner was used as a background control. Gel slices were processed for MS as follows. The bands were subjected to in-gel slice tryptic digestion, followed by a desalting step. The resulting peptides were analyzed using Waters HSS-T3 column on nanoflow liquid chromatography (nanoAcquity) coupled to high resolution, high mass accuracy mass spectrometry (Q Exactive Plus). The samples were analyzed by discovery mode. The data were processed using Proteome Discoverer version 2.4.1.15, searched against custom databases, to which a list of common lab contaminants was added. The search was carried out with the Byonic search algorithms using the fixed modification of cysteine carbamidomethylation, and the variable modification of methionine oxidation, asparagine- or glutamine-deamidation, and protein N-terminal acetylation. One combined database was generated from the custom-tailored subjected databases of Euechinoidea_proteins_collective (53221 protein entries of all identified to the date proteins for Euechinoidea subclass uploaded from NCBI), P_lividus_translated_EMBOSS (raw translated by all 6 frames peptides untreated while removing all X and * indicate stop codons; 468 entries), Trf_P_lividus_cleaned (53 entries of cleaned manually confirmed sequences).
4.9. Bacterial Binding Assay
Assays for PlTrf protein binding to E. coli employed 109 live bacteria incubated with either live P. lividus coelomocytes or cell-free CF for 30 min at room temperature with gentle shaking. After the incubation, coelomocytes were washed 3 times for 5 min each with CMFSW-EH. Bacteria that were incubated with cell-free CF were pelleted at 10,000× g for 5 min and washed 2 times for 5 min each. Both coelomocytes and bacteria were lysed and used for Trf protein profiling by Western blot as described above.
4.10. Immune Challenge
100 μL of aCF (10 mM CaCl
2, 14 mM KCl, 50 mM MgCl
2, 398 mM NaCl, 1.7 mM NaHCO
3, 25 mM Na
2SO
4, and 10 mM HEPES, pH 7.4) [
16] containing 10
6 heat-killed (95 °C for 5 min)
E. coli or
V. penaeicida or 1 μg of LPS per mL of sea urchin CF were injected into 3 or 5 sea urchins in each of the experimental groups. The CF volumes was calculated according to (weight of animal (g) × 0.22 = mL whole coelomic fluid (wCF)) [
43]. Equivalent volumes of aCF were injected into control animals (
n = 3 or 5). Before injections, CF from all animals was used to measure the base level of Trf expression.
4.11. FCM and FACS
The CF from challenged and unchallenged animals were collected on days 0 and days 1, 2, 14, 30, 43 post-challenge as described above and evaluated by flow cytometry (FCM). All steps in preparation for FCM were performed on ice. Cells were counted and adjusted to 106 cells/mL. Coelomocytes were washed as described above and pelleted by centrifugation at 500× g for 5 min at 4 °C. The pellet was gently resuspended in 100 μL of a staining medium (3.3× PBS with 20 mM HEPES, 2% fetal calf serum, 0.09% sodium azide, pH 7.4) plus a mixture of a 1:300 dilution each of three anti-SpTrf polyclonal antibodies provided by L. Courtney Smith. Control samples were resuspended in a staining medium without the primary antibodies. Samples were incubated for 30 min on ice in the dark, followed by cell pelleting and washing in a 500 μL staining medium. The washed cells were resuspended in 30 μL of staining medium with the 1:250 dilution of Goat anti-Rabbit-Ig secondary antibody conjugated to Alexa Fluor 488 (Thermo Fisher Pierce) and incubated on ice for 30 min in the dark. Cells were pelleted and resuspended in 500 μL staining medium containing 1 μg/mL DAPI for the exclusion of dead cells and subjected to the FCM evaluation or by FACS. The evaluation was performed on the Calibur flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA) and analyzed by FlowJo 10.6 software (FlowJo LLC, Ashland, OR, USA). Sorting was carried out using MA900 cell sorter (Sony, Tokyo, Japan). Sorted live Trf-positive cells were imaged with Nikon Eclipse fluorescent microscope and with ZEISS LSM900 confocal microscope (Thornwood, NY, USA).
4.12. Phagocytosis Assay
Cells and cell-free CF from three sea urchins were used in the phagocytosis assay. The cells were washed two times with cold staining medium and pelleted for 3 min at 500 g at 4 °C. Washed cells from each animal were divided into three aliquots and kept on ice until use. Either FITC-stained or mCherry expressing E. coli were used for the phagocytosis assays. Bacteria were diluted to a concentration of 107 per ml and divided into three groups. Groups A and C were incubated for 40 min with sea urchin CF at 16 °C in the dark. Group B was incubated in the absence of cells. Group C bacteria were subsequently incubated with antiSpTrf polyclonal antibodies mixture at 1:100 dilution in 100 μL for 30 min in the dark. All bacteria samples (A, B, C) were mixed with the corresponding coelomocyte aliquots of each of three sea urchins and incubated in the dark for 70 min. After the incubation, cells were pelleted for 3 min at 500× g and loaded to the hemocytometer for the visual evaluation of phagocytosis using fluorescent microscope Nikon Eclipse with standard fluorescent lamp and relevant filters (model IX81, with Nikon filters). Phagocytosis events were counted blindly (without knowing the treatment type), using the 16 middle squares (n = 3) of the hemocytometer for A, B and C phagocytosis treatments. Differences in the percentages of cells with phagocyted bacteria in different treatments were assessed by Anova two-factor and one-factor analyses.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}