
Effects of Tyrosine and Tryptophan Supplements on the Vital Indicators in Mice Differently Prone to Diet-Induced Obesity

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Amino Acids Characterization
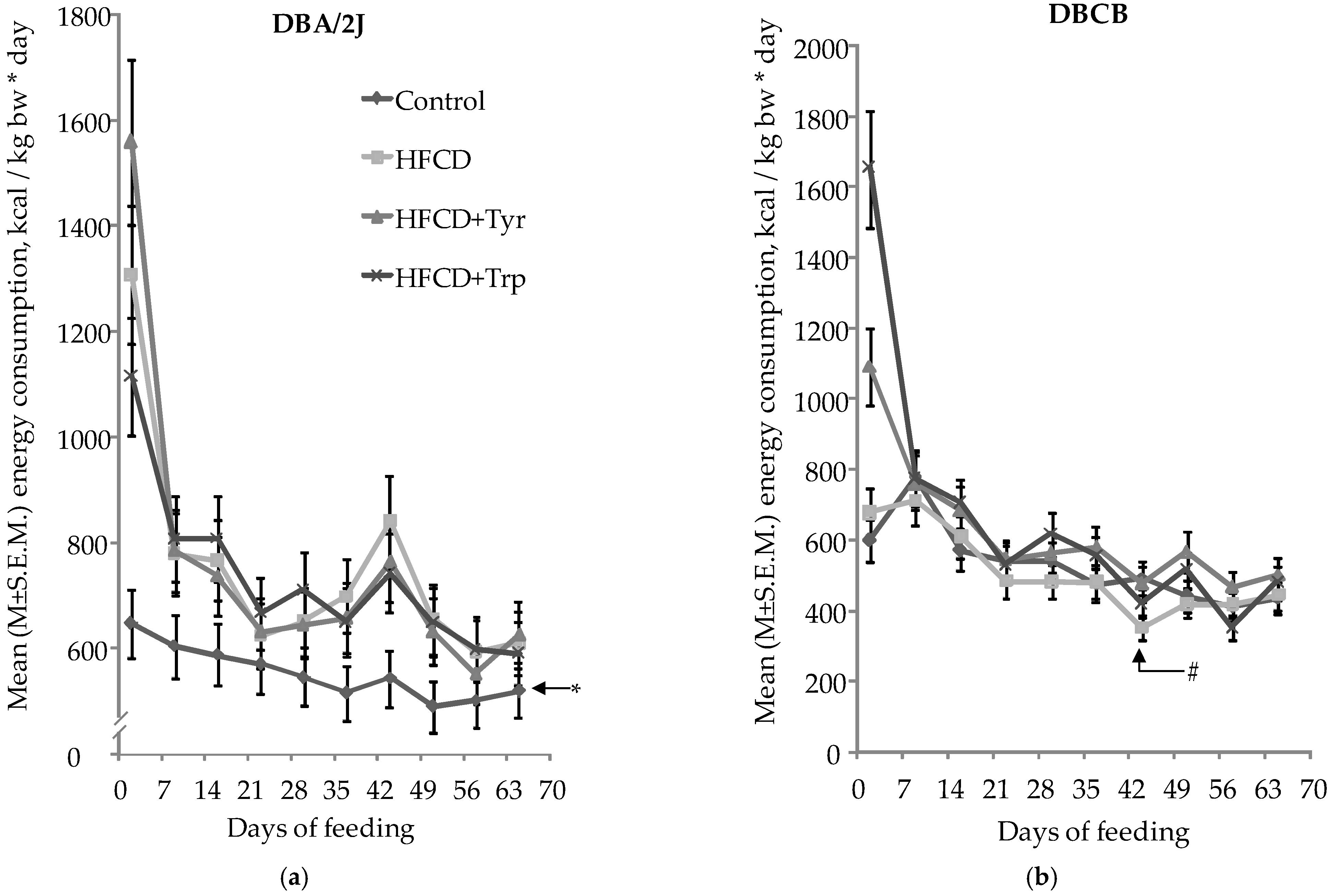
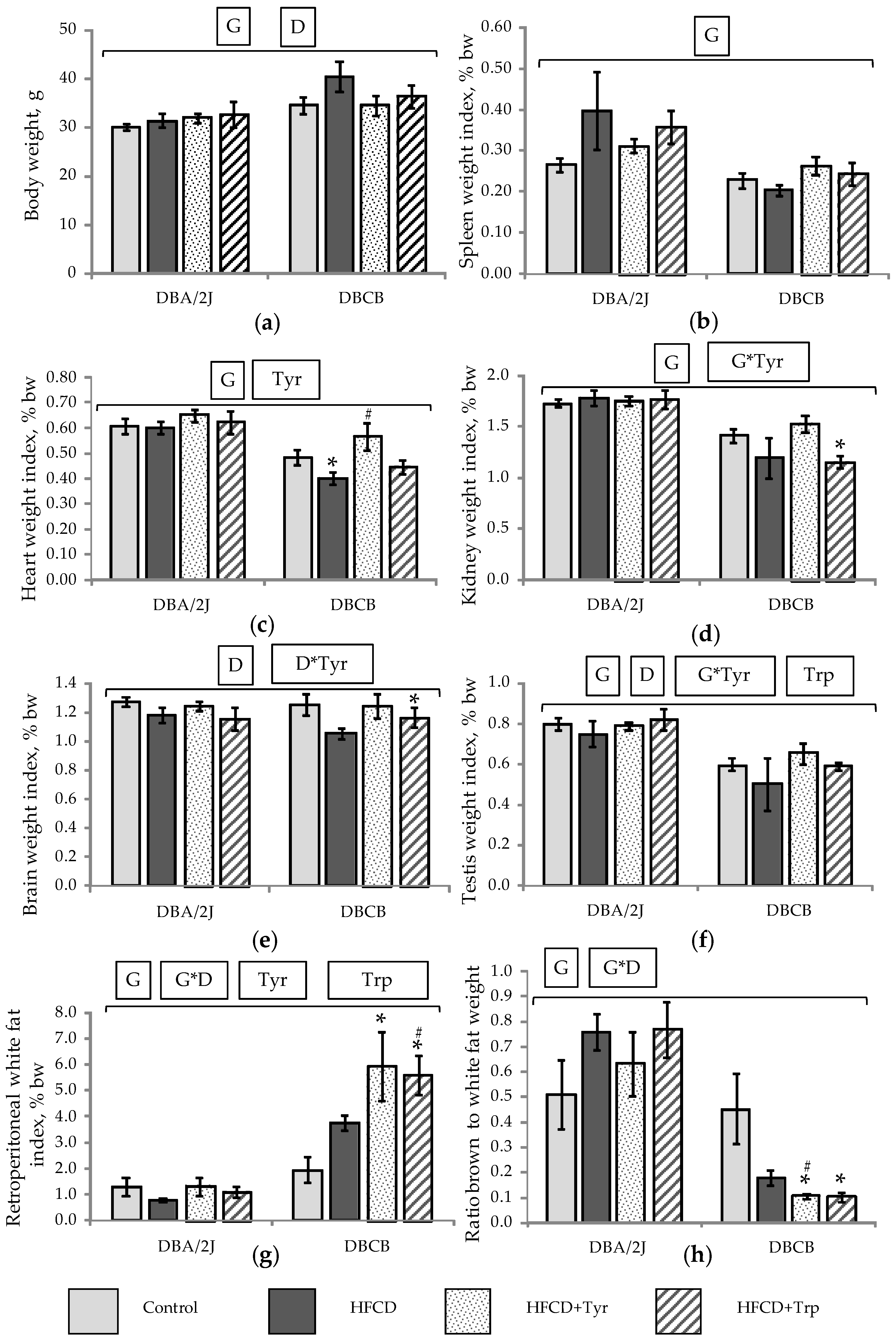
2.2. Mice Integral Indices
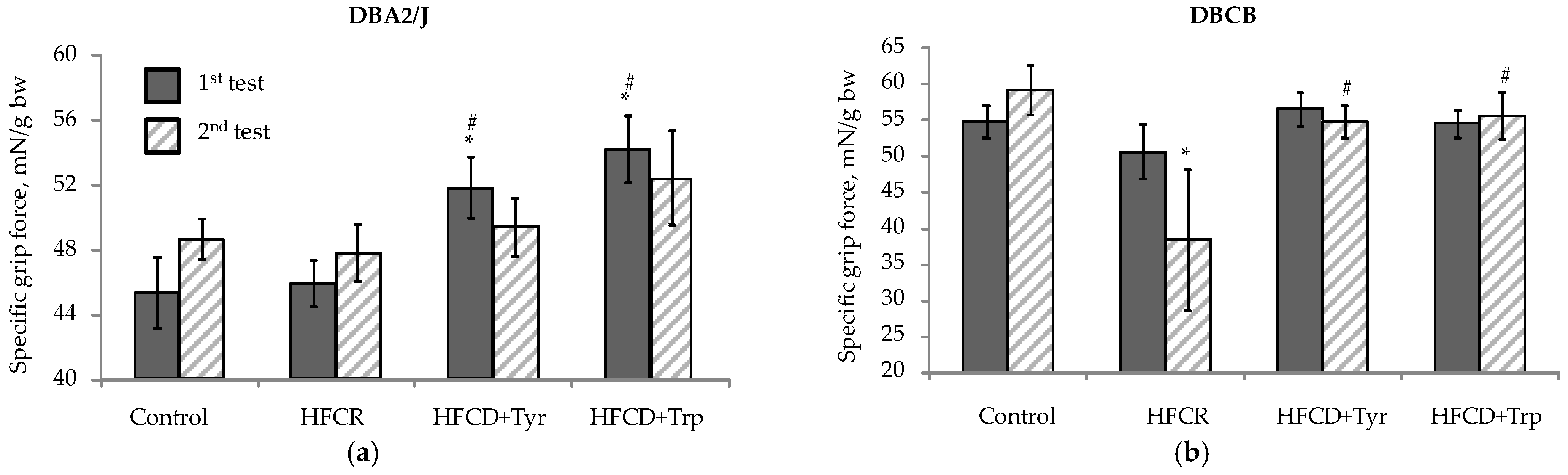
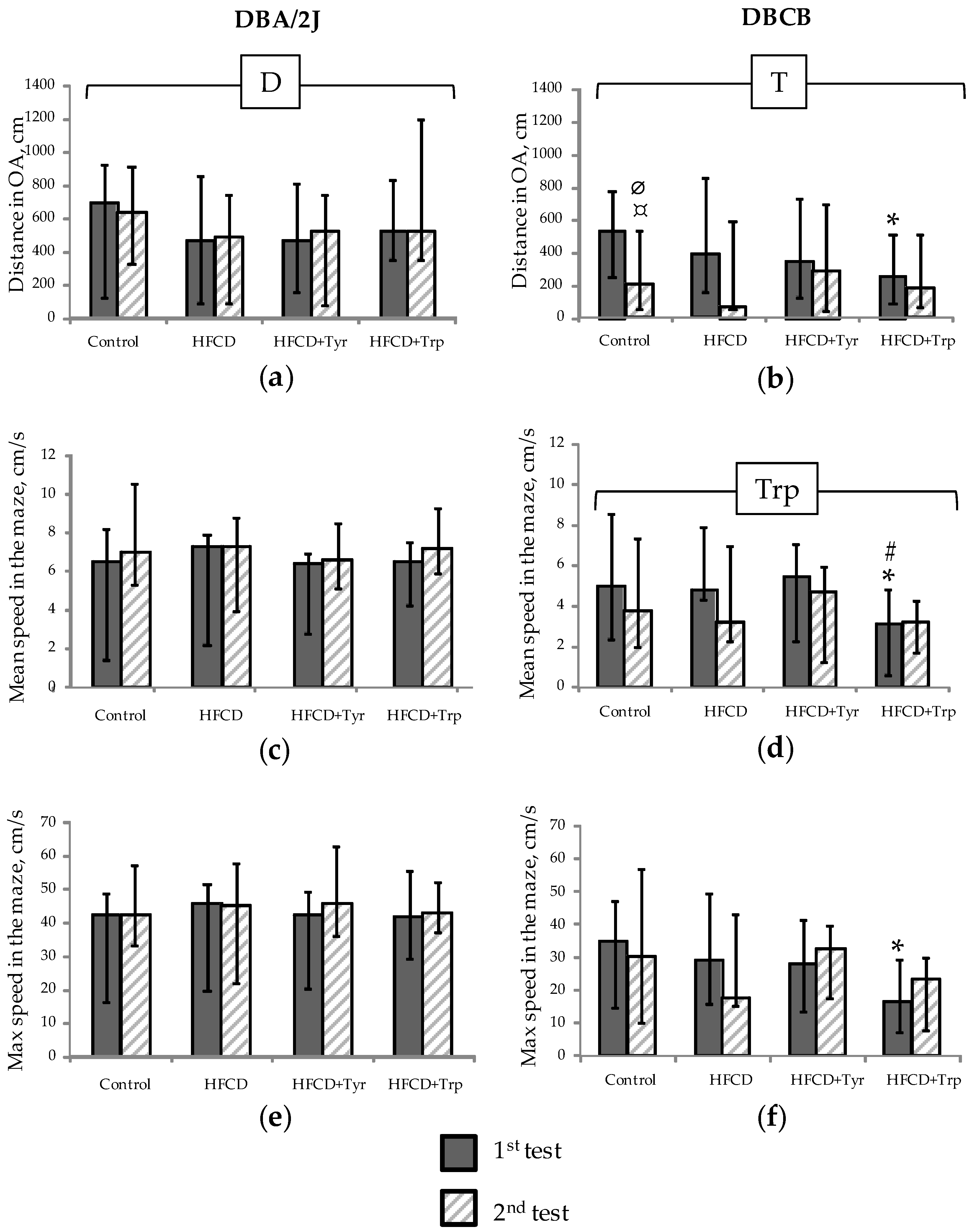
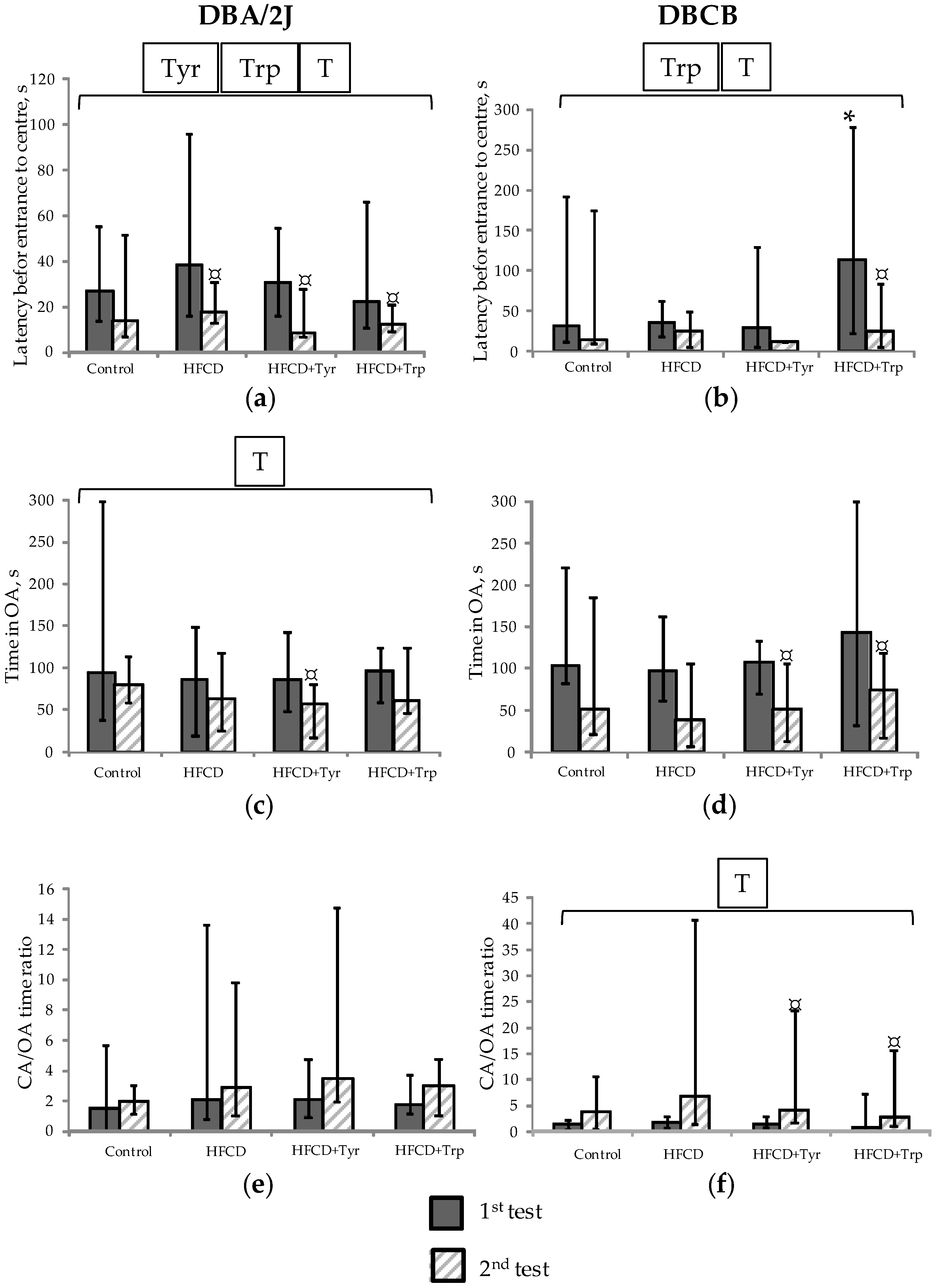
2.3. Muscle Tone, Search, and Anxiety-Like Behavioral Activity
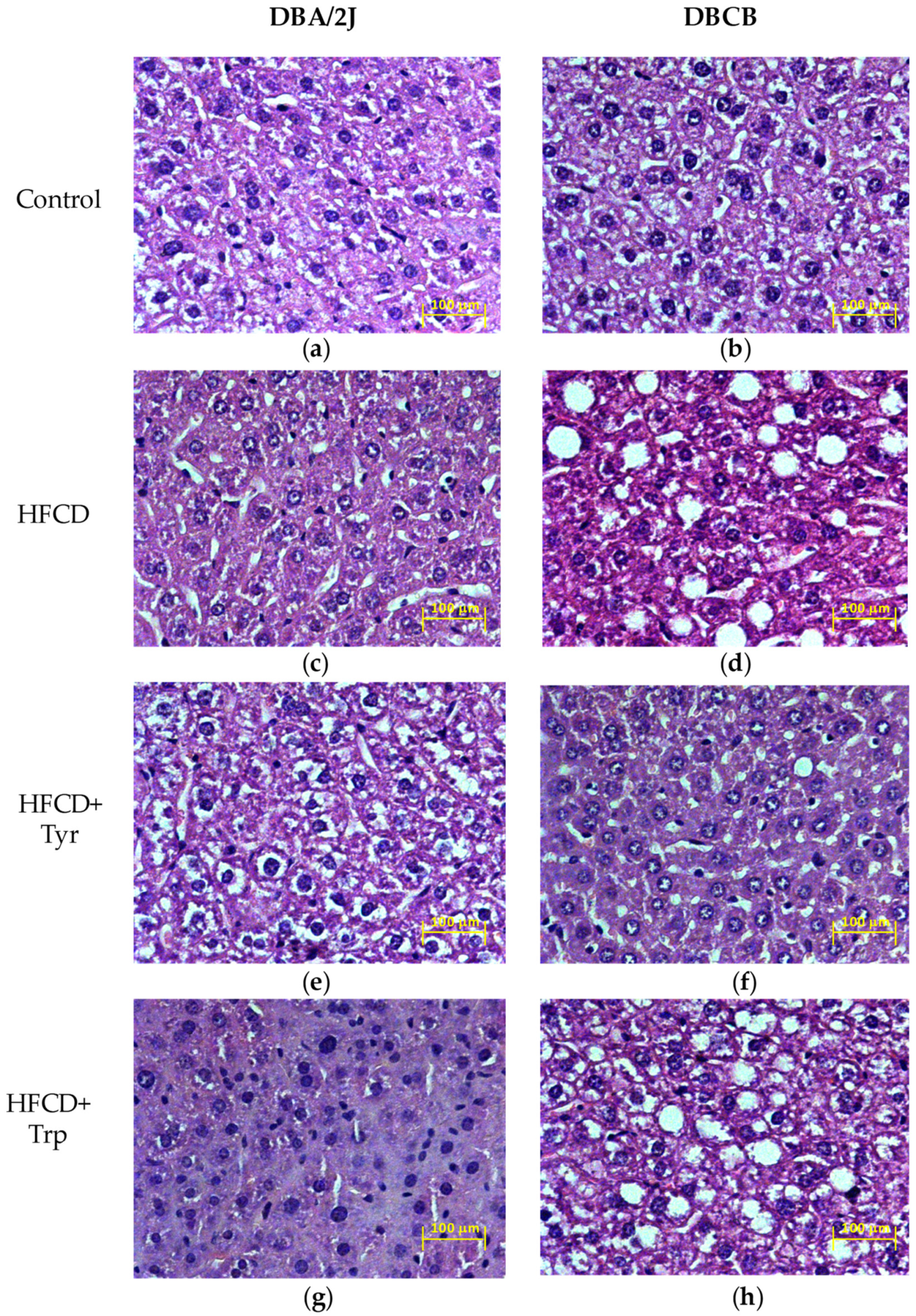
2.4. Liver Histology
2.5. Blood Biochemical Indices
2.6. Blood Cytokines and Adipokines Levels
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Assessment of Behavioral Responses, Anxiety, and Muscle Tone Indices
4.3. Assessment of Integral and Biochemical Indices
4.4. Cytokines and Adipokines Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheung, W.W.; Mao, P. Recent advances in obesity: Genetics and beyond. ISRN Endocrinol. 2012, 2012, 536905. [Google Scholar] [CrossRef]
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef]
- Bojanowska, E.; Ciosek, J. Can We Selectively Reduce Appetite for Energy-Dense Foods? An Overview of Pharmacological Strategies for Modification of Food Preference Behavior. Curr. Neuropharmacol. 2016, 14, 118–142. [Google Scholar] [CrossRef] [PubMed]
- Rønnestad, I.; Gomes, A.S.; Murashita, K.; Angotzi, R.; Jönsson, E.; Volkoff, H. Appetite-Controlling Endocrine Systems in Teleosts. Front. Endocrinol. 2017, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.J. Reward mechanisms in obesity: New insights and future directions. Neuron 2011, 69, 664–679. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wise, R.A. How can drug addiction help us understand obesity? Nat. Neurosci. 2005, 8, 555–560. [Google Scholar] [CrossRef]
- Vucetic, Z.; Carlin, J.L.; Totoki, K.; Reyes, T.M. Epigenetic dysregulation of the dopamine system in diet-induced obesity. J. Neurochem. 2012, 120, 891–898. [Google Scholar] [CrossRef]
- Burke, L.K.; Heisler, L.K. 5-hydroxytryptamine medications for the treatment of obesity. J. Neuroendocrinol. 2015, 27, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef]
- Hornykiewicz, O. A brief history of levodopa. J. Neurol. 2010, 257, S249–S252. [Google Scholar] [CrossRef]
- Wurtman, R.J.; Wurtman, J.J. Carbohydrate craving, obesity and brain serotonin. Appetite 1986, 7, 99–103. [Google Scholar] [CrossRef]
- Herrera, C.P.; Smith, K.; Atkinson, F.; Ruell, P.; Chow, C.M.; O’Connor, H.; Brand-Miller, J. High-glycaemic index and glycaemic load meals increase the availability of tryptophan in healthy volunteers. Br. J. Nutr. 2011, 105, 1601–1606. [Google Scholar] [CrossRef]
- Trusov, N.V.; Shipelin, V.A.; Mzhelskaya, K.V.; Shumakova, A.A.; Timonin, A.N.; Riger, N.A.; Apryatin, S.A.; Gmoshinski, I.V. Effect of resveratrol on behavioral, biochemical, and immunological parameters of DBA/2J and tetrahybrid DBCB mice receiving diet with excess fat and fructose. J. Nutr. Biochem. 2021, 88, 108527. [Google Scholar] [CrossRef]
- Allen, J.A.; Varga, J. Eosinophilia-Myalgia Syndrome. In Encyclopedia of Toxicology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 419–425. [Google Scholar] [CrossRef]
- Apryatin, S.A.; Shipelin, V.A.; Sidorova, Y.S.; Petrov, N.A.; Gmoshinskii, I.V.; Nikityuk, D.B. Interspecific Differences in Behavioral Responses and Neuromotorics between Laboratory Rodents Receiving Rations with Easily Digested Carbohydrates. Bull. Exp. Biol. Med. 2018, 165, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Matter, C.M.; Handschin, C. RANTES (regulated on activation, normal T cell expressed and secreted), inflammation, obesity, and the metabolic syndrome. Circulation 2007, 115, 946–948. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, M.; Iwata, M.; Inoue, T.; Aizawa, Y.; Yoshito, N.; Hayashi, K.; Suzuki, S. Decreased grip strength, muscle pain, and atrophy occur in rats following long-term exposure to excessive repetitive motion. FEBS Open Bio 2017, 7, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Botros, M.; Sikaris, K.A. The de ritis ratio: The test of time. Clin. Biochem. Rev. 2013, 34, 117–130. [Google Scholar] [PubMed]
- Nikniaz, L.; Nikniaz, Z.; Tabrizi, J.S.; Sadeghi-Bazargani, H.; Farahbakhsh, M. Is within-normal range liver enzymes associated with metabolic syndrome in adults? Clin. Res. Hepatol. Gastroenterol. 2018, 42, 92–98. [Google Scholar] [CrossRef]
- Esnault, S.; Kelly, E.A. Essential Mechanisms of Differential Activation of Eosinophils by IL-3 Compared to GM-CSF and IL-5. Crit. Rev. Immunol. 2016, 36, 429–444. [Google Scholar] [CrossRef]
- Gregory, B.; Kirchem, A.; Phipps, S.; Gevaert, P.; Pridgeon, C.; Rankin, S.M.; Robinson, D.S. Differential regulation of human eosinophil IL-3, IL-5, and GM-CSF receptor alpha-chain expression by cytokines: IL-3, IL-5, and GM-CSF down-regulate IL-5 receptor alpha expression with loss of IL-5 responsiveness, but up-regulate IL-3 receptor alpha exp. J. Immunol. 2003, 170, 5359–5366. [Google Scholar] [CrossRef]
- Williams, R.L.; Wood, L.G.; Collins, C.E.; Morgan, P.J.; Callister, R. Energy homeostasis and appetite regulating hormones as predictors of weight loss in men and women. Appetite 2016, 101, 1–7. [Google Scholar] [CrossRef]
- Apryatin, S.A.; Shipelin, V.A.; Trusov, N.V.; Mzhelskaya, K.V.; Evstratova, V.S.; Kirbaeva, N.V.; Soto, J.S.; Fesenko, Z.S.; Gainetdinov, R.R.; Gmoshinski, I.V. Comparative analysis of the influence of a high-fat/high-carbohydrate diet on the level of anxiety and neuromotor and cognitive functions in Wistar and DAT-KO rats. Physiol. Rep. 2019, 7, e13987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.S.; Davies, S.S. Microbial metabolism of dietary components to bioactive metabolites: Opportunities for new therapeutic interventions. Genome Med. 2016, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Jourde-Chiche, N.; Faure, V.; Cerini, C.; Berland, Y.; Dignat-George, F.; Brunet, P. The uremic solute indoxyl sulfate induces oxidative stress in endothelial cells. J. Thromb. Haemost. 2007, 5, 1302–1308. [Google Scholar] [CrossRef]
- Karbownik, M.; Stasiak, M.; Zygmunt, A.; Zasada, K.; Lewiński, A. Protective effects of melatonin and indole-3-propionic acid against lipid peroxidation, caused by potassium bromate in the rat kidney. Cell Biochem. Funct. 2006, 24, 483–489. [Google Scholar] [CrossRef]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; Deng, Y.; Blennerhassett, P.; Macri, J.; McCoy, K.D.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609.e1–3. [Google Scholar] [CrossRef] [PubMed]
- Bartolomeo, M.P.; Maisano, F. Validation of a reversed-phase HPLC method for quantitative amino acid analysis. J. Biomol. Tech. 2006, 17, 131–137. [Google Scholar] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Watson, R.R. Trace Elements in Laboratory Rodents; CRC Press: Boca Raton, FL, USA; London, UK, 1996; p. 416. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | DBA/2J | DBCB | Factor *** | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Group No | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| Diet | Control | HFCD | HFCD + Tyr | HRCD + Trp | Control | HFCD | HFCD + Tyr | HRCD + Trp | |
| Number of Animals | 8 | 7 | 8 | 8 | 7 | 4 | 7 | 7 | |
| Glucose, mmol/L | 12.8 ± 0.7 2–8* | 16.2 ± 0.9 1 | 17.8 ± 0.8 1 | 19.8 ± 1.9 1 | 20.6 ± 1.8 1 | 22.1 ± 2.5 1 | 20.2 ± 1.8 1 | 20.2 ± 0.7 1 | G |
| Protein, g/L | 52.0 ± 2.5 3,4 | 52.9 ± 2.0 3,4 | 61.1 ± 1.2 1,2 | 59.8 ± 2.4 1,2 | 58.0 ± 3.6 | 56.1 ± 4.3 | 61.5 ± 1.2 | 60.1 ± 1.0 | Tyr |
| Albumin, g/L | 25.7 ± 1.2 3,4 | 25.0 ± 1.9 3 | 30.1 ± 0.7 1,2 | 29.6 ± 1.4 1 | 31.0 ± 1.2 | 29.7 ± 2.0 | 32.1 ± 0.6 | 32.7 ± 0.5 | Tyr, Trp |
| Urea, mmol/L | 5.15 ± 0.45 3,4 | 5.18 ± 0.34 3,4 | 7.27 ± 0.43 1,2 | 6.54 ± 0.32 1,2 | 6.63 ± 0.42 | 5.57 ± 0.32 7,8 | 7.03 ± 0.46 6 | 6.59 ± 0.22 6 | G ****, Tyr, Trp |
| Calcium, mmol/L | 1.86 ± 0.09 3,4 | 1.86 ± 0.10 3,4 | 2.35 ± 0.12 1,2 | 2.34 ± 0.14 1,2 | 2.15 ± 0.06 | 1.95 ± 0.19 8 | 2.30 ± 0.08 | 2.26 ± 0.06 6 | Tyr, Trp |
| Phosphorus, mmol/L | 2.51 ± 0.19 4 | 2.95 ± 0.19 | 3.06 ± 0.26 | 3.72 ± 0.32 1 | 3.30 ± 0.20 | 3.03 ± 0.36 | 3.14 ± 0.23 | 3.37 ± 0.19 | Trp |
| AlAT, U/mL | 22.0 ± 3.8 3 | 27.5 ± 11.1 | 50.9 ± 13.4 1 | 35.3 ± 8.6 | 19.6 ± 4.9 6 | 62.8 ± 20.5 5 | 26.9 ± 6.2 | 47.7 ± 16.2 | D, G × Tyr |
| AsAT, U/mL | 220 ± 43 | 201 ± 39 | 205 ± 29 | 258 ± 66 | 180 ± 35 | 454 ± 253 | 191 ± 39 | 220 ± 38 | G × D |
| AsAT/AlAT | 11.2 ± 2.0 3 | 10.6 ± 2.5 3 | 5.0 ± 0.8 1,2 | 9.9 ± 2.6 | 12.5 ± 5.1 | 10.1 ± 4.6 | 5.1 ± 0.3 | 12.3 ± 4.7 | Tyr |
| KPK kU/mL | 5.7 ± 1.7 | 5.9 ± 1.6 | 5.9 ± 0.9 | 7.1 ± 1.9 | 5.1 ± 0.7 | 10.7 ± 3.7 7 | 3.6 ± 1.0 6 | 5.6 ± 1.4 | Tyr |
| Lipase U/mL | 129 ± 13 | 110 ± 4 4 | 120 ± 3 | 138 ± 9 2 | 119 ± 15 | 111 ± 7 8 | 148 ± 20 | 140 ± 8 6 | Trp |
| GPO mmol/min/ mg protein ** | 0.71 ± 0.02 2,3,4 | 0.39 ± 0.03 1 | 0.40 ± 0.03 1 | 0.46 ± 0.01 1 | 0.68 ± 0.03 6,7 | 0.44 ± 0.03 5,8 | 0.49 ± 0.06 5 | 0.60 ± 0.02 6 | G, D, Trp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gmoshinski, I.V.; Shipelin, V.A.; Trusov, N.V.; Apryatin, S.A.; Mzhelskaya, K.V.; Shumakova, A.A.; Timonin, A.N.; Riger, N.A.; Nikityuk, D.B. Effects of Tyrosine and Tryptophan Supplements on the Vital Indicators in Mice Differently Prone to Diet-Induced Obesity. Int. J. Mol. Sci. 2021, 22, 5956. https://doi.org/10.3390/ijms22115956
Gmoshinski IV, Shipelin VA, Trusov NV, Apryatin SA, Mzhelskaya KV, Shumakova AA, Timonin AN, Riger NA, Nikityuk DB. Effects of Tyrosine and Tryptophan Supplements on the Vital Indicators in Mice Differently Prone to Diet-Induced Obesity. International Journal of Molecular Sciences. 2021; 22(11):5956. https://doi.org/10.3390/ijms22115956
Chicago/Turabian StyleGmoshinski, Ivan V., Vladimir A. Shipelin, Nikita V. Trusov, Sergey A. Apryatin, Kristina V. Mzhelskaya, Antonina A. Shumakova, Andrey N. Timonin, Nikolay A. Riger, and Dmitry B. Nikityuk. 2021. "Effects of Tyrosine and Tryptophan Supplements on the Vital Indicators in Mice Differently Prone to Diet-Induced Obesity" International Journal of Molecular Sciences 22, no. 11: 5956. https://doi.org/10.3390/ijms22115956
APA StyleGmoshinski, I. V., Shipelin, V. A., Trusov, N. V., Apryatin, S. A., Mzhelskaya, K. V., Shumakova, A. A., Timonin, A. N., Riger, N. A., & Nikityuk, D. B. (2021). Effects of Tyrosine and Tryptophan Supplements on the Vital Indicators in Mice Differently Prone to Diet-Induced Obesity. International Journal of Molecular Sciences, 22(11), 5956. https://doi.org/10.3390/ijms22115956