
Mialostatin, a Novel Midgut Cystatin from Ixodes ricinus Ticks: Crystal Structure and Regulation of Host Blood Digestion

 , , ,
, , , 
Abstract
1. Introduction
2. Results
2.1. Mialostatin Transcript Predominantly Accumulates in the Tick Midgut
2.2. Mialostatin Is a Broad-Spectrum Inhibitor of Cysteine Cathepsins and Is Highly Effective against Cathepsin L
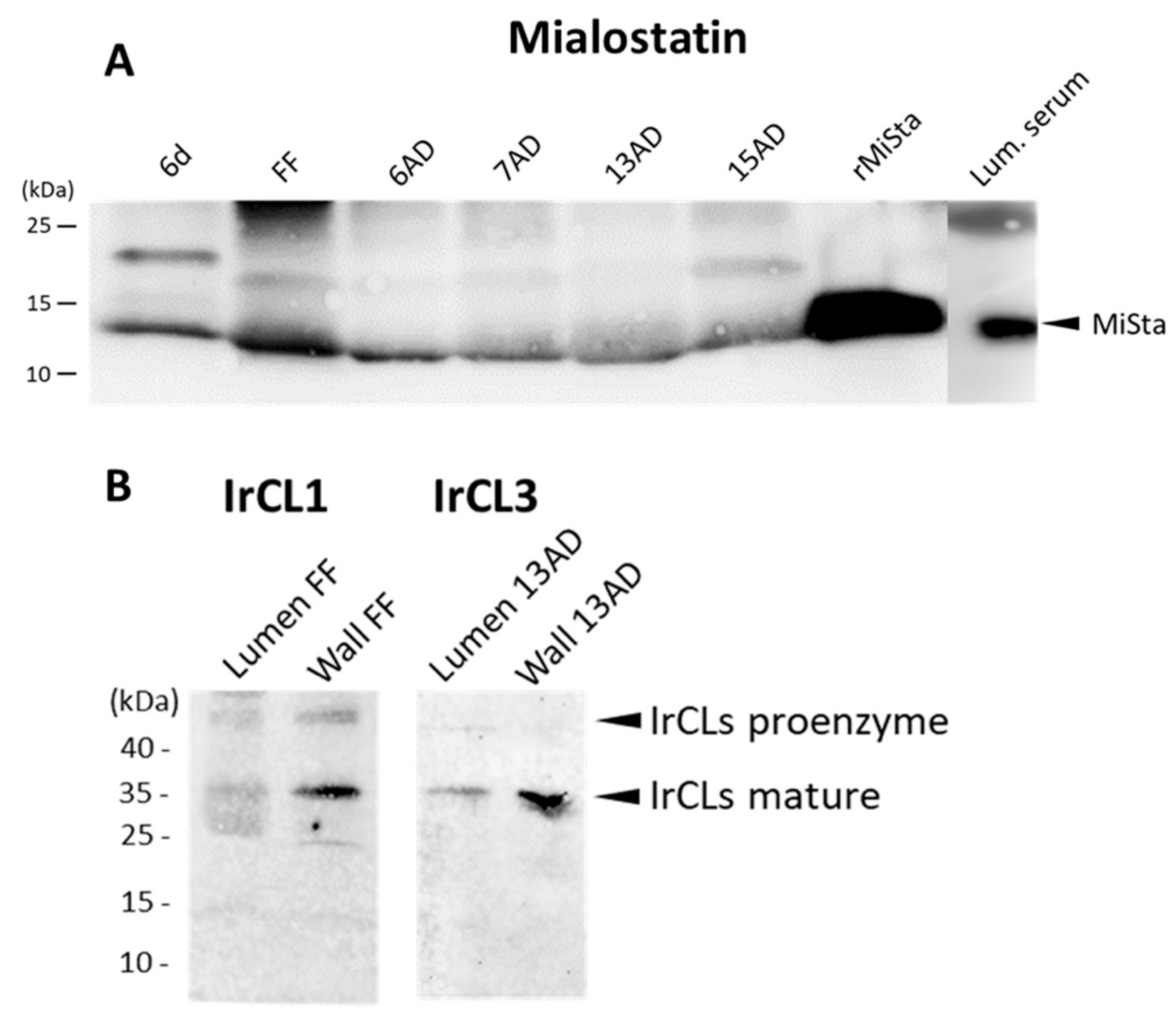
2.3. Mialostatin Is Present in the Tick Gut Wall and Lumen
2.4. Mialostatin Inhibits Blood-Protein Digestion Catalyzed by Tick Gut Cysteine Cathepsins
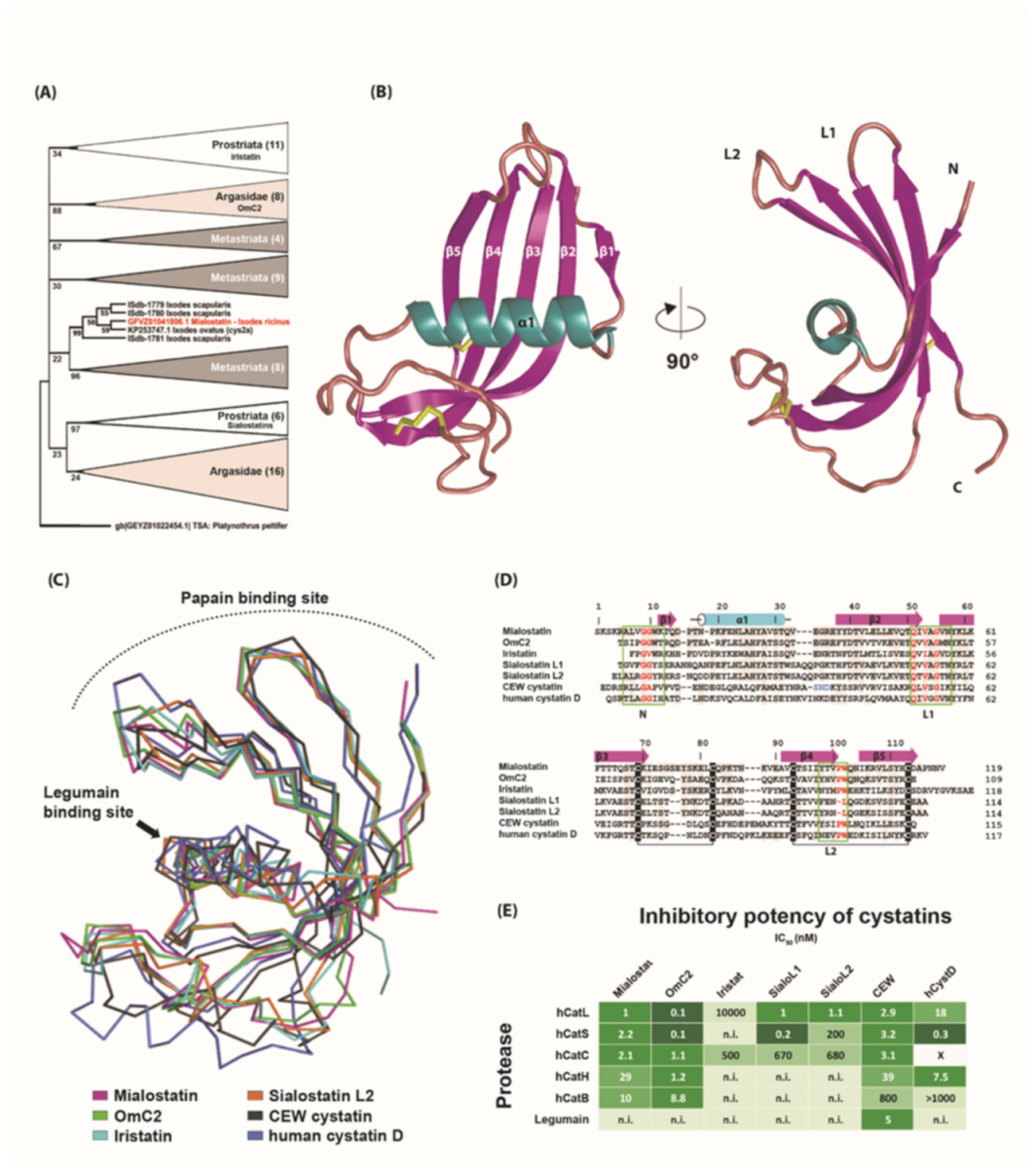
2.5. Phylogenetic Analysis and Three-Dimensional Structure of Mialostatin and Its Reactive Site
3. Discussion
4. Materials and Methods
4.1. Ticks and Laboratory Animals
4.2. Quantitative Real-Time PCR
4.3. Mialostatin Cloning, Expression, Refolding, and Purification and Antibody Production
4.4. Preparation of Tick Gut Samples
4.5. Protease Inhibition Assays
4.5.1. Inhibition of Proteases in Tick Gut Homogenates
4.5.2. Inhibition Assays of Recombinant Tick Proteases
4.5.3. Inhibition Assays of Human Proteases
4.6. Protein Digestion Assay
4.7. Reducing SDS-PAGE and Western Blotting
4.8. Immunohistochemistry
4.9. Evolutionary Analysis by the Maximum Likelihood Method
4.10. Crystallization and Data Collection
4.11. Structure Determination
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindgren, E.; Talleklint, L.; Polfeldt, T. Impact of climatic change on the northern latitude limit and population density of the disease-transmitting European tick Ixodes ricinus. Environ. Health Perspect. 2000, 108, 119–123. [Google Scholar] [CrossRef]
- Yang, Y.; Christie, J.; Köster, L.; Du, A.; Yao, C. Emerging Human Babesiosis with “Ground Zero” in North America. Microorganisms 2021, 9, 440. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Roe, R.M. Biology of Ticks, 2nd ed.; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Lara, F.A.; Lins, U.; Bechara, G.H.; Oliveira, P.L. Tracing heme in a living cell: Hemoglobin degradation and heme traffic in digest cells of the cattle tick Boophilus microplus. J. Exp. Biol. 2005, 208, 3093–3101. [Google Scholar] [CrossRef] [PubMed]
- Sojka, D.; Pytelková, J.; Perner, J.; Horn, M.; Konvičková, J.; Schrenková, J.; Mareš, M.; Kopáček, P. Multienzyme degradation of host serum albumin in ticks. Ticks Tick-Borne Dis. 2016, 7, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.; Nussbaumerová, M.; Šanda, M.; Kovářová, Z.; Srba, J.; Franta, Z.; Sojka, D.; Bogyo, M.; Caffrey, C.R.; Kopáček, P.; et al. Hemoglobin Digestion in Blood-Feeding Ticks: Mapping a Multipeptidase Pathway by Functional Proteomics. Chem. Biol. 2009, 16, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Sojka, D.; Hajdušek, O.; Dvořák, J.; Sajid, M.; Franta, Z.; Schneider, E.L.; Craik, C.S.; Vancová, M.; Burešová, V.; Bogyo, M.; et al. IrAE—An asparaginyl endopeptidase (legumain) in the gut of the hard tick Ixodes ricinus. Int. J. Parasitol. 2007, 37, 713–724. [Google Scholar] [CrossRef]
- Sojka, D.; Franta, Z.; Frantová, H.; Bartošová, P.; Horn, M.; Váchová, J.; O’Donoghue, A.J.; Eroy-Reveles, A.A.; Craik, C.S.; Knudsen, G.M.; et al. Characterization of Gut-associated Cathepsin D Hemoglobinase from Tick Ixodes ricinus (IrCD1). J. Biol. Chem. 2012, 287, 21152–21163. [Google Scholar] [CrossRef] [PubMed]
- Franta, Z.; Sojka, D.; Frantova, H.; Dvorak, J.; Horn, M.; Srba, J.; Talacko, P.; Mares, M.; Schneider, E.; Craik, C.S.; et al. IrCL1—The haemoglobinolytic cathepsin L of the hard tick, Ixodes ricinus. Int. J. Parasitol. 2011, 41, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Sojka, D.; Franta, Z.; Horn, M.; Hajdušek, O.; Caffrey, C.R.; Mares, M.; Kopáček, P. Profiling of proteolytic enzymes in the gut of the tick Ixodes ricinus reveals an evolutionarily conserved network of aspartic and cysteine peptidases. Parasites Vectors 2008, 1, 7. [Google Scholar] [CrossRef]
- Franta, Z.; Frantová, H.; Konvičková, J.; Horn, M.; Sojka, D.; Mareš, M.; Kopáček, P. Dynamics of digestive proteolytic system during blood feeding of the hard tick Ixodes ricinus. Parasites Vectors 2010, 3, 119. [Google Scholar] [CrossRef]
- Sojka, D.; Francischetti, I.M.B.; Calvo, E.; Kotsyfakis, M. Cysteine Proteases from Bloodfeeding Arthropod Ectoparasites. Adv. Exp. Med. Biol. 2011, 712, 177–191. [Google Scholar] [CrossRef]
- Sojka, D.; Franta, Z.; Horn, M.; Caffrey, C.R.; Mareš, M.; Kopáček, P. New insights into the machinery of blood digestion by ticks. Trends Parasitol. 2013, 29, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, C.R.; Goupil, L.; Rebello, K.M.; Dalton, J.P.; Smith, D. Cysteine proteases as digestive enzymes in parasitic helminths. PLoS Negl. Trop. Dis. 2018, 12, e0005840. [Google Scholar] [CrossRef] [PubMed]
- Novinec, M.; Lenarčič, B.; Turk, B. Cysteine Cathepsin Activity Regulation by Glycosaminoglycans. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Turk, V.; Bode, W. The cystatins: Protein inhibitors of cysteine proteinases. FEBS Lett. 1991, 285, 213–219. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Waller, M.; Barrett, A.J.; Bateman, A. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2014, 42, D503–D509. [Google Scholar] [CrossRef]
- Schwarz, A.; Valdés, J.J.; Kotsyfakis, M. The role of cystatins in tick physiology and blood feeding. Ticks Tick-Borne Dis. 2012, 3, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Chmelař, J.; Kotál, J.; Langhansová, H.; Kotsyfakis, M. Protease Inhibitors in Tick Saliva: The Role of Serpins and Cystatins in Tick-host-Pathogen Interaction. Front. Cell. Infect. Microbiol. 2017, 7, 216. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.A.; Kotál, J.; Bensaoud, C.; Chmelař, J.; Kotsyfakis, M. Small protease inhibitors in tick saliva and salivary glands and their role in tick-host-pathogen interactions. Biochim. Biophys. Acta BBA Proteins Proteom. 2020, 1868, 140336. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.A.; Sasaki, S.D.; Tanaka, A.S. Bmcystatin, a cysteine proteinase inhibitor characterized from the tick Boophilus microplus. Biochem. Biophys. Res. Commun. 2006, 347, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Parizi, L.F.; Githaka, N.W.; Acevedo, C.; Benavides, U.; Seixas, A.; Logullo, C.; Konnai, S.; Ohashi, K.; Masuda, A.; Vaz, I.D.S. Sequence characterization and immunogenicity of cystatins from the cattle tick Rhipicephalus (Boophilus) microplus. Ticks Tick-Borne Dis. 2013, 4, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Grunclová, L.; Horn, M.; Vancová, M.; Sojka, D.; Franta, Z.; Mares, M.; Kopáček, P. Two secreted cystatins of the soft tick Ornithodoros moubata: Differential expression pattern and inhibitory specificity. Biol. Chem. 2006, 387, 1635–1644. [Google Scholar] [CrossRef]
- Salát, J.; Paesen, G.C.; Řezáčová, P.; Kotsyfakis, M.; Kovářová, Z.; Šanda, M.; Majtán, J.; Grunclová, L.; Horká, H.; Andersen, J.F.; et al. Crystal structure and functional characterization of an immunomodulatory salivary cystatin from the soft tick Ornithodoros moubata. Biochem. J. 2010, 429, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Zavašnik-Bergant, T.; Vidmar, R.; Sekirnik, A.; Fonović, M.; Salát, J.; Grunclová, L.; Kopáček, P.; Turk, B. Salivary Tick Cystatin OmC2 Targets Lysosomal Cathepsins S and C in Human Dendritic Cells. Front. Cell. Infect. Microbiol. 2017, 7, 288. [Google Scholar] [CrossRef] [PubMed]
- Parizi, L.F.; Sabadin, G.A.; Alzugaray, M.F.; Seixas, A.; Logullo, C.; Konnai, S.; Ohashi, K.; Masuda, A.; da Sliva Vaz, I., Jr. Rhipicephalus microplus and Ixodes ovatus cystatins in tick blood digestion and evasion of host immune response. Parasites Vectors 2015, 8, 122. [Google Scholar] [CrossRef]
- Rangel, C.K.; Parizi, L.F.; Sabadin, G.A.; Costa, E.P.; Romeiro, N.C.; Isezaki, M.; Githaka, N.W.; Seixas, A.; Logullo, C.; Konnai, S.; et al. Molecular and structural characterization of novel cystatins from the taiga tick Ixodes persulcatus. Ticks Tick-Borne Dis. 2017, 8, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Charrier, N.P.; Couton, M.; Voordouw, M.J.; Rais, O.; Durand-Hermouet, A.; Hervet, C.; Plantard, O.; Rispe, C. Whole body transcriptomes and new insights into the biology of the tick Ixodes ricinus. Parasites Vectors 2018, 11, 1–15. [Google Scholar] [CrossRef]
- Kotsyfakis, M.; Schwarz, A.; Erhart, J.; Ribeiro, J.M.C. Tissue- and time-dependent transcription in Ixodes ricinus salivary glands and midguts when blood feeding on the vertebrate host. Sci. Rep. 2015, 5, srep09103. [Google Scholar] [CrossRef]
- Cramaro, W.J.; Revets, D.; Hunewald, O.E.; Sinner, R.; Reye, A.L.; Muller, C.P. Integration of Ixodes ricinus genome sequencing with transcriptome and proteome annotation of the naïve midgut. BMC Genom. 2015, 16, 871. [Google Scholar] [CrossRef]
- Perner, J.; Provazník, J.; Schrenková, J.; Urbanová, V.; Ribeiro, J.M.C.; Kopáček, P. RNA-seq analyses of the midgut from blood- and serum-fed Ixodes ricinus ticks. Sci. Rep. 2016, 6, 36695. [Google Scholar] [CrossRef]
- Kotál, J.; Stergiou, N.; Buša, M.; Chlastáková, A.; Beránková, Z.; Řezáčová, P.; Langhansová, H.; Schwarz, A.; Calvo, E.; Kopecký, J.; et al. The structure and function of Iristatin, a novel immunosuppressive tick salivary cystatin. Cell. Mol. Life Sci. 2019, 76, 2003–2013. [Google Scholar] [CrossRef] [PubMed]
- Kotsyfakis, M.; Horka, H.; Salat, J.; Andersen, J.F. The crystal structures of two salivary cystatins from the tick Ixodes scapularis and the effect of these inhibitors on the establishment of Borrelia burgdorferi infection in a murine model. Mol. Microbiol. 2010, 77, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Kotsyfakis, M.; Sá-Nunes, A.; Francischetti, I.M.B.; Mather, T.N.; Andersen, J.F.; Ribeiro, J.M.C. Antiinflammatory and Immunosuppressive Activity of Sialostatin L, a Salivary Cystatin from the Tick Ixodes scapularis. J. Biol. Chem. 2006, 281, 26298–26307. [Google Scholar] [CrossRef]
- Balbin, M.; Hall, A.; Grubb, A.; Mason, R.W.; Lopez-Otin, C.; Abrahamson, M. Structural and functional characterization of two allelic variants of human cystatin D sharing a characteristic inhibition spectrum against mammalian cysteine proteinases. J. Biol. Chem. 1994, 269, 23156–23162. [Google Scholar] [CrossRef]
- Anastasi, A.; Brown, M.A.; Kembhavi, A.A.; Nicklin, M.J.H.; Sayers, C.A.; Sunter, D.C.; Barrett, A.J. Cystatin, a protein inhibitor of cysteine proteinases. Improved purification from egg white, characterization, and detection in chicken serum. Biochem. J. 1983, 211, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Vasiljeva, O.; Dolinar, M.; Turk, V.; Turk, B. Recombinant Human Cathepsin H Lacking the Mini Chain Is an Endopeptidase. Biochemistry 2003, 42, 13522–13528. [Google Scholar] [CrossRef] [PubMed]
- Pidugu, L.S.; Maity, K.; Ramaswamy, K.; Surolia, N.; Suguna, K. Analysis of proteins with the ’Hot dog’ fold: Prediction of function and identification of catalytic residues of hypothetical proteins. BMC Struct. Biol. 2009, 9, 37. [Google Scholar] [CrossRef]
- Alvarez-Fernandez, M.; Liang, Y.-H.; Abrahamson, M.; Su, X.-D. Crystal Structure of Human Cystatin D, a Cysteine Peptidase Inhibitor with Restricted Inhibition Profile. J. Biol. Chem. 2005, 280, 18221–18228. [Google Scholar] [CrossRef]
- Bode, W.; Engh, R.; Musil, D.; Thiele, U.; Huber, R.; Karshikov, A.; Brzin, J.; Kos, J.; Turk, V. The 2.0 A X-ray crystal structure of chicken egg white cystatin and its possible mode of interaction with cysteine proteinases. EMBO J. 1988, 7, 2593–2599. [Google Scholar] [CrossRef]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochim. Biophys. Acta BBA Proteins Proteom. 2012, 1824, 68–88. [Google Scholar] [CrossRef]
- Nandy, S.K.; Seal, A. Structural Dynamics Investigation of Human Family 1 & 2 Cystatin-Cathepsin L1 Interaction: A Comparison of Binding Modes. PLoS ONE 2016, 11, e0164970. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, X.; Cao, J.; Zhou, Y.; Gong, H.; Zhang, H.; Li, X.; Zhou, J. Characterization of a secreted cystatin from the tick Rhipicephalus haemaphysaloides. Exp. Appl. Acarol. 2015, 67, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; da Rocha, L.A.; Torquato, R.J.; Junior, I.D.S.V.; Florin-Christensen, M.; Tanaka, A.S. A novel type 1 cystatin involved in the regulation of Rhipicephalus microplus midgut cysteine proteases. Ticks Tick-Borne Dis. 2020, 11, 101374. [Google Scholar] [CrossRef] [PubMed]
- Kordiš, D.; Turk, V. Phylogenomic analysis of the cystatin superfamily in eukaryotes and prokaryotes. BMC Evol. Biol. 2009, 9, 266. [Google Scholar] [CrossRef]
- Kopáček, P.; Perner, J.; Sojka, D.; Šíma, R.; Hajdušek, O. Molecular Targets to Impair Blood Meal Processing in Ticks. In Ectoparasites; Wiley: New York, NY, USA, 2018; Volume 8, pp. 139–165. [Google Scholar]
- Reyes, J.; Ayala-Chavez, C.; Sharma, A.; Pham, M.; Nuss, A.B.; Gulia-Nuss, M. Blood Digestion by Trypsin-Like Serine Proteases in the Replete Lyme Disease Vector Tick, Ixodes scapularis. Insects 2020, 11, 201. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.E.; Fogaça, A.C.; Nakayasu, E.S.; Angeli, C.B.; Belmonte, R.; Almeida, I.C.; Miranda, A.; Miranda, M.T.M.; Tanaka, A.S.; Braz, G.R.; et al. Characterization of proteinases from the midgut of Rhipicephalus (Boophilus) microplus involved in the generation of antimicrobial peptides. Parasites Vectors 2010, 3, 63. [Google Scholar] [CrossRef]
- Xavier, M.A.; Tirloni, L.; Torquato, R.; Tanaka, A.; Pinto, A.F.M.; Diedrich, J.K.; Yates, J.R., 3rd; da Silva Vaz, I., Jr.; Seixas, A.; Termignoni, C. Blood anticlotting activity of a Rhipicephalus microplus cathepsin L-like enzyme. Biochimie 2019, 163, 12–20. [Google Scholar] [CrossRef]
- Rodríguez-Mallon, A. Developing Anti-tick Vaccines. Methods Mol. Biol. 2016, 1404, 243–259. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Balk, J.A.; Postigo, M.; Jongejan, F. Selection of reference genes for quantitative RT-PCR studies in Rhipicephalus (Boophilus) microplus and Rhipicephalus appendiculatus ticks and determination of the expression profile of Bm86. BMC Mol. Biol. 2009, 10, 112. [Google Scholar] [CrossRef]
- Urbanová, V.; Hartmann, D.; Grunclová, L.; Šíma, R.; Flemming, T.; Hajdušek, O.; Kopáček, P. IrFC—An Ixodes ricinus injury-responsive molecule related to Limulus Factor C. Dev. Comp. Immunol. 2014, 46, 439–447. [Google Scholar] [CrossRef]
- Vechtova, P.; Fussy, Z.; Cegan, R.; Sterba, J.; Erhart, J.; Benes, V.; Grubhoffer, L. Catalogue of stage-specific transcripts in Ixodes ricinus and their potential functions during the tick life-cycle. Parasites Vectors 2020, 13, 1–19. [Google Scholar] [CrossRef]
- Kopácek, P.; Zdychová, J.; Yoshiga, T.; Weise, C.; Rudenko, N.; Law, J.H. Molecular cloning, expression and isolation of ferritins from two tick species--Ornithodoros moubata and Ixodes ricinus. Insect Biochem. Mol. Biol. 2003, 33, 103–113. [Google Scholar] [CrossRef]
- Hurrell, J.G.R. Monoclonal Hybridoma Antibodies: Techniques and Applications; CRC Press: Boca Raton, FL, USA, 2017; p. 239. [Google Scholar]
- Russo, C.; Callegaro, L.; Lanza, E.; Ferrone, S. Purification of IgG monoclonal antibody by caprylic acid precipitation. J. Immunol. Methods 1983, 65, 269–271. [Google Scholar] [CrossRef]
- Hartmann, D.; Šíma, R.; Konvičková, J.; Perner, J.; Kopáček, P.; Sojka, D. Multiple legumain isoenzymes in ticks. Int. J. Parasitol. 2018, 48, 167–178. [Google Scholar] [CrossRef]
- Hánová, I.; Brynda, J.; Houštecká, R.; Alam, N.; Sojka, D.; Kopáček, P.; Marešová, L.; Vondrášek, J.; Horn, M.; Schueler-Furman, O.; et al. Novel Structural Mechanism of Allosteric Regulation of Aspartic Peptidases via an Evolutionarily Conserved Exosite. Cell Chem. Biol. 2018, 25, 318–329.e4. [Google Scholar] [CrossRef]
- Schagger, H. Tricine-SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evol. Int. J. Org. Evol. 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Mueller, U.; Darowski, N.; Fuchs, M.R.; Förster, R.; Hellmig, M.; Paithankar, K.S.; Pühringer, S.; Steffien, M.; Zocher, G.; Weiss, M.S. Facilities for macromolecular crystallography at the Helmholtz-Zentrum Berlin. J. Synchrotron Radiat. 2012, 19, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. Integration, scaling, space-group assignment and post-refinement. Acta Cryst. Sect. D Biol. Cryst. 2010, 66, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Int. Tables Crystallogr. 2012, 66, 364–366. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of theCCP4 suite and current developments. Acta Cryst. Sect. D Biol. Cryst. 2011, 67, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development ofCoot. Acta Cryst. Sect. D Biol. Cryst. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Cryst. D Biol. Cryst. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibition of I. ricinus Midgut Homogenate | Inhibition of Recombinant Digestive I. ricinus Proteases | Inhibition of Human Cysteine Cathepsins | |||
|---|---|---|---|---|---|
| Targeted Activity | IC50 (nM) | Proteases | IC50 (nM) | Protease | IC50 (nM) |
| Cathepsin L | 0.18 ± 0.02 | Ir-cathepsin L1 (IrCL1) | 0.071 ± 0.01 | Hs-cathepsin L | 0.38 ± 0.03 |
| Cathepsins L and B | 3.1 ± 0.4 | Ir-cathepsin L3 (IrCL3) | 0.39 ± 0.18 | Hs-cathepsin C | 2.1 ± 0.8 |
| Cathepsin B | 12.1 ± 1.5 | Ir-legumain (IrAE) | n.i. | Hs-cathepsin S | 2.2 ± 0.4 |
| Cathepsin C | 91.7 ± 5.5 | Ir-cathepsin D1 (IrCD1) | n.i. | Hs-cathepsin B | 9.0 ± 0.3 |
| Hs-cathepsin K | 9.7 ± 1.3 | ||||
| Hs-cathepsin H | 24.0 ± 3.5 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotál, J.; Buša, M.; Urbanová, V.; Řezáčová, P.; Chmelař, J.; Langhansová, H.; Sojka, D.; Mareš, M.; Kotsyfakis, M. Mialostatin, a Novel Midgut Cystatin from Ixodes ricinus Ticks: Crystal Structure and Regulation of Host Blood Digestion. Int. J. Mol. Sci. 2021, 22, 5371. https://doi.org/10.3390/ijms22105371
Kotál J, Buša M, Urbanová V, Řezáčová P, Chmelař J, Langhansová H, Sojka D, Mareš M, Kotsyfakis M. Mialostatin, a Novel Midgut Cystatin from Ixodes ricinus Ticks: Crystal Structure and Regulation of Host Blood Digestion. International Journal of Molecular Sciences. 2021; 22(10):5371. https://doi.org/10.3390/ijms22105371
Chicago/Turabian StyleKotál, Jan, Michal Buša, Veronika Urbanová, Pavlína Řezáčová, Jindřich Chmelař, Helena Langhansová, Daniel Sojka, Michael Mareš, and Michail Kotsyfakis. 2021. "Mialostatin, a Novel Midgut Cystatin from Ixodes ricinus Ticks: Crystal Structure and Regulation of Host Blood Digestion" International Journal of Molecular Sciences 22, no. 10: 5371. https://doi.org/10.3390/ijms22105371
APA StyleKotál, J., Buša, M., Urbanová, V., Řezáčová, P., Chmelař, J., Langhansová, H., Sojka, D., Mareš, M., & Kotsyfakis, M. (2021). Mialostatin, a Novel Midgut Cystatin from Ixodes ricinus Ticks: Crystal Structure and Regulation of Host Blood Digestion. International Journal of Molecular Sciences, 22(10), 5371. https://doi.org/10.3390/ijms22105371