
Plant Copper Amine Oxidases: Key Players in Hormone Signaling Leading to Stress-Induced Phenotypic Plasticity

 , and
, and 
Abstract
1. Introduction: The Plant Amine Oxidase Network
2. The Amine Oxidase CuAO Gene Family: Subcellular Localization and Biochemical Properties of CuAOs from Different Plant Species
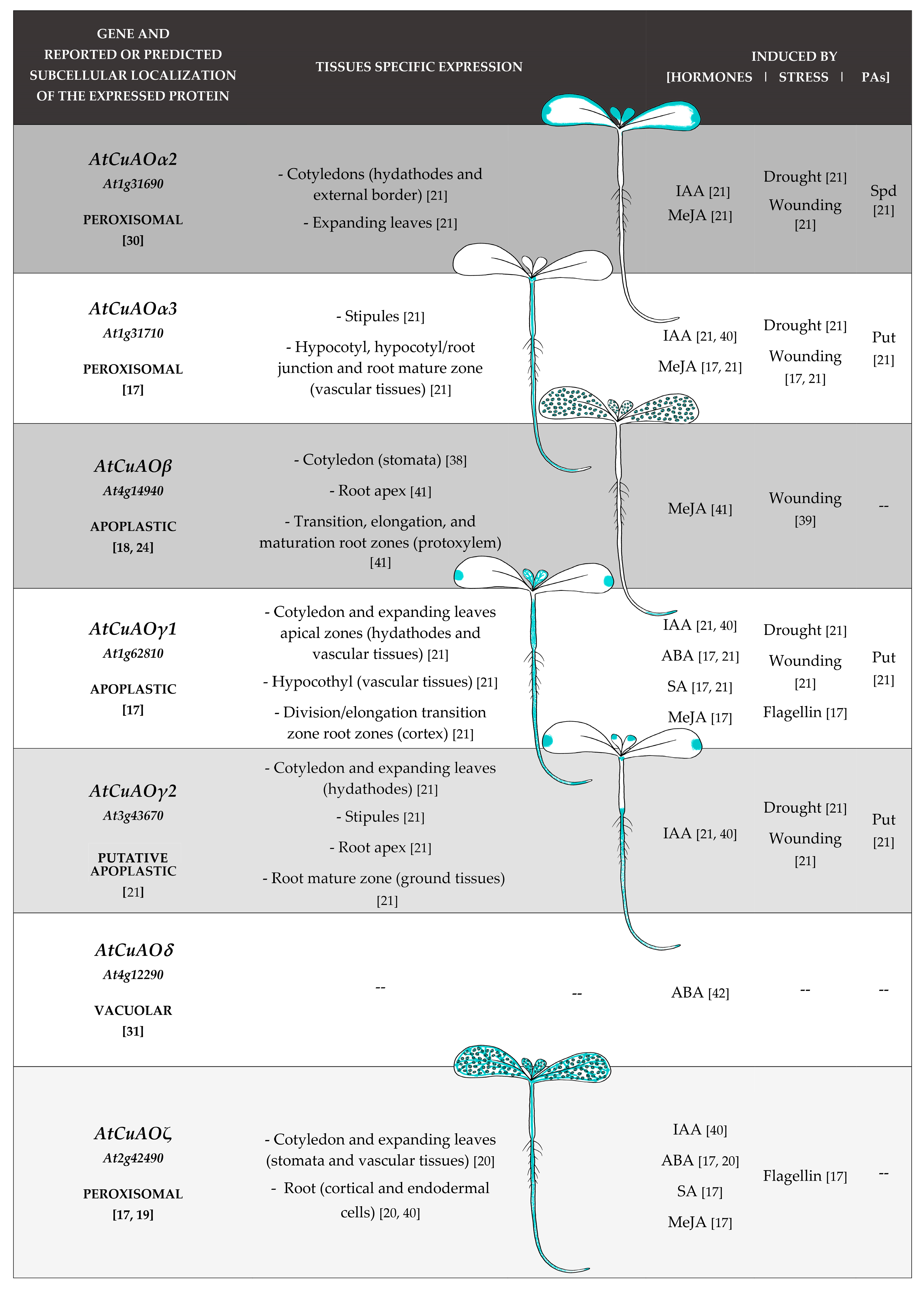
3. CuAOs in Development and Hormonal Signaling Pathways: Analysis of Tissues Specific- and Hormone/Stress-Modulated-CuAO Expression Profiles
4. CuAOs as PA Level Modulators and Sources of Reactive Compounds in the Cytoplasm and Cell Wall
5. AOs Play a Role in Xylem Phenotypic Plasticity
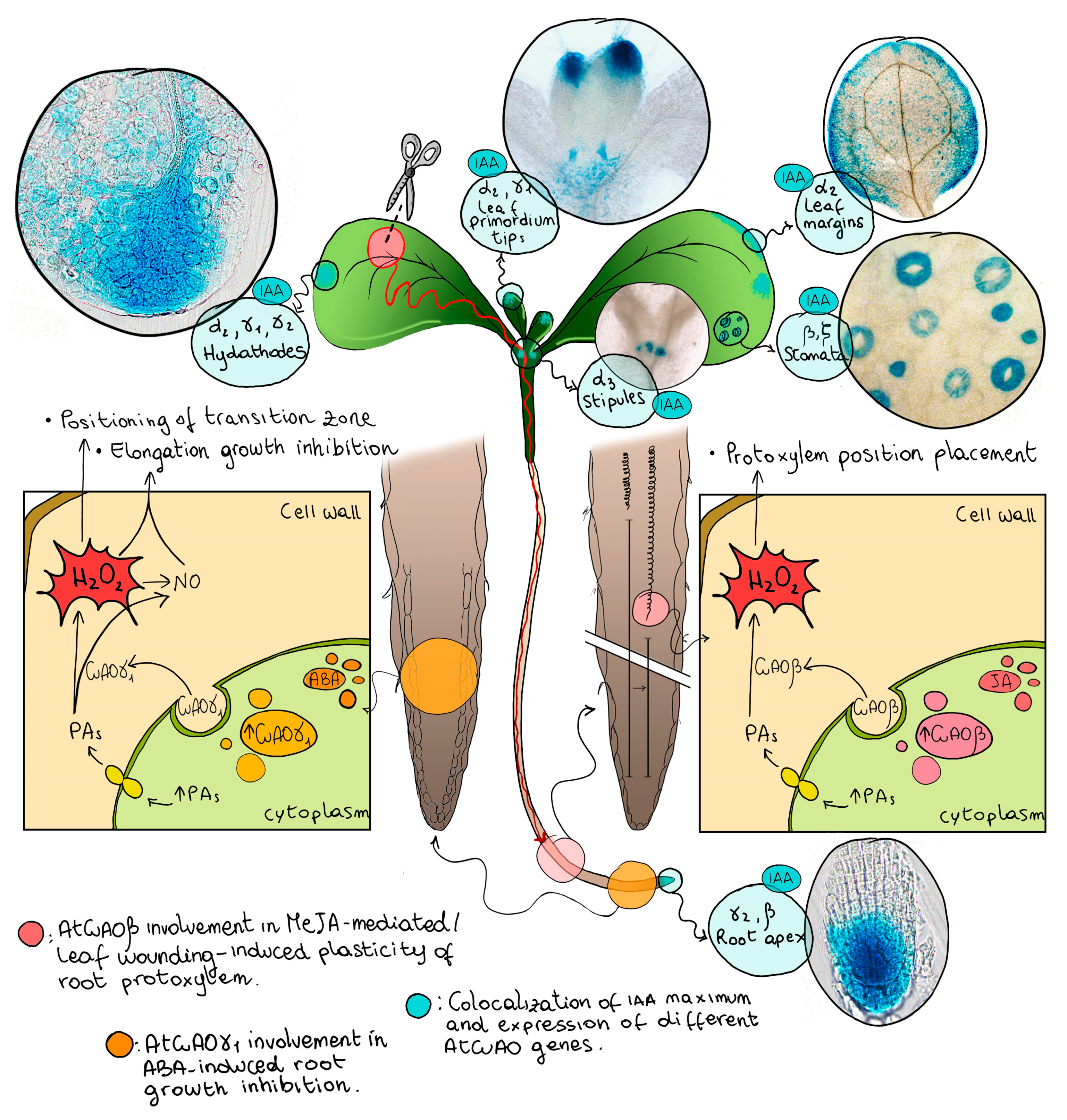
6. Expression of IAA- and Stress-Inducible AtCuAOs in Free-Auxin Maximum Zones Suggest an Role in Both Developmentally Regulated and Stress-Induced Xylem Differentiation in the Leaf and in the Root
7. The AtCuAOβ-Driven H2O2 Production Plays a Role in Stress-Induced/MeJA-Signaled Early Protoxylem Differentiation in the Root
8. AtCuAOβ Is Involved in Early Root Protoxylem Differentiation Induced by Leaf Wounding through Systemic Signaling
9. AtCuAOγ1 Is Involved in ABA-Associated Growth Responses
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wuddineh, W.; Minocha, R.; Minocha, S.C. Polyamines in the Context of Metabolic Networks. Methods Mol. Biol. 2018, 1694, 1–23. [Google Scholar] [PubMed]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Podlešáková, K.; Ugena, L.; Spíchal, L.; Doležal, K.; De Diego, N. Phytohormones and polyamines regulate plant stress responses by altering GABA pathway. New Biotechnol. 2019, 48, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, A.; Sharma, N.; Bhardwaj, R.; Ohri, P. Emerging Role of Polyamines in Plant Stress Tolerance. Curr. Protein Pept. Sci. 2018, 19, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front. Chem. 2018, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Fortes, A.M.; Agudelo-Romero, P. Polyamine Metabolism in Climacteric and Non-Climacteric Fruit Ripening. Methods Mol. Biol. 2018, 1694, 433–447. [Google Scholar]
- Wang, W.; Paschalidis, K.; Feng, J.C.; Song, J.; Liu, J.H. Polyamine Catabolism in Plants: A Universal Process with Diverse Functions. Front. Plant Sci. 2019, 10, 561. [Google Scholar] [CrossRef]
- Tavladoraki, P.; Cona, A.; Federico, R.; Tempera, G.; Viceconte, N.; Saccoccio, S.; Battaglia, V.; Toninello, A.; Agostinelli, E. Polyamine catabolism: Target for antiproliferative therapies in animals and stress tolerance strategies in plants. Amino Acids 2012, 42, 411–426. [Google Scholar] [CrossRef]
- Tavladoraki, P.; Cona, A.; Angelini, R. Copper-containing amine oxidases and FAD-dependent polyamine oxidases are key players in plant tissue differentiation and organ development. Front. Plant Sci. 2016, 7, 824. [Google Scholar] [CrossRef]
- Moschou, P.N.; Wu, J.; Cona, A.; Tavladoraki, P.; Angelini, R.; Roubelakis-Angelakis, K.A. The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J. Exp. Bot. 2012, 63, 5003–5015. [Google Scholar] [CrossRef]
- Angelini, R.; Cona, A.; Federico, R.; Fincato, P.; Tavladoraki, P.; Tisi, A. Plant amine oxidases “on the move”: An update. Plant Physiol. Biochem. 2010, 48, 560–564. [Google Scholar] [CrossRef]
- Cona, A.; Rea, G.; Angelini, R.; Federico, R.; Tavladoraki, P. Functions of amine oxidases in plant development and defence. Trends Plant Sci. 2006, 11, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Federico, R.; Angelini, R. Polyamine catabolism in plants. In Biochemistry and Physiology of Polyamines in Plants, 1st ed.; Slocum, R.D., Flores, H.E., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 41–56. [Google Scholar]
- Šebela, M.; Radová, A.; Angelini, R.; Tavladoraki, P.; Frébort, I.; Peč, P. FAD-containing polyamine oxidases: A timely challenge for researchers in biochemistry and physiology of plants. Plant Sci. 2001, 160, 197–207. [Google Scholar] [CrossRef]
- Groß, F.; Rudolf, E.E.; Thiele, B.; Durner, J.; Astier, J. Copper amine oxidase 8 regulates arginine-dependent nitric oxide production in Arabidopsis thaliana. J. Exp. Bot. 2017, 68, 2149–2162. [Google Scholar] [CrossRef] [PubMed]
- Planas-Portell, J.; Gallart, M.; Tiburcio, A.F.; Altabella, T. Copper containing amine oxidases contribute to terminal polyamine oxidation in peroxisomes and apoplast of Arabidopsis thaliana. BMC Plant Biol. 2013, 13, 109. [Google Scholar] [CrossRef]
- Møller, S.G.; McPherson, M.J. Developmental expression and biochemical analysis of the Arabidopsis atao1 gene encoding an H2O2-generating diamine oxidase. Plant J. 1998, 13, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Naconsie, M.; Kato, K.; Shoji, T.; Hashimoto, T. Molecular evolution of N-methylputrescine oxidase in tobacco. Plant Cell Physiol. 2014, 55, 436–444. [Google Scholar] [CrossRef]
- Qu, Y.; An, Z.; Zhuang, B.; Jing, W.; Zhang, Q.; Zhang, W. Copper amine oxidase and phospholipase D act independently in abscisic acid (ABA)-induced stomatal closure in Vicia faba and Arabidopsis. J. Plant Res. 2014, 127, 533–544. [Google Scholar] [CrossRef]
- Fraudentali, I.; Ghuge, S.A.; Carucci, A.; Tavladoraki, P.; Angelini, R.; Rodrigues-Pousada, R.A.; Cona, A. Developmental, hormone- and stress-modulated expression profiles of four members of the Arabidopsis copper-amine oxidase gene family. Plant Physiol. Biochem. 2020, 147, 141–160. [Google Scholar] [CrossRef]
- Wang, W.; Wu, H.; Liu, J.H. Genome-wide identification and expression profiling of copper-containing amine oxidase genes in sweet orange (Citrus sinensis). Tree Genet. Genomes 2017, 13, 31. [Google Scholar] [CrossRef]
- Bharalee, R.; Gupta, S.; Bandyopadhyay, T.; Gohain, B.; Agarwala, N.; Kalita, C.M.; Das, S. Molecular cloning, expression and computational analysis of a water stress inducible copper-containing amine oxidase gene (CuAO) from tea plant [Camellia sinensis (L.) O. Kuntze]. Afr. J. Biotechnol. 2012, 11, 15547–15555. [Google Scholar]
- Boudart, G.; Jamet, E.; Rossignol, M.; Lafitte, C.; Borderies, G.; Jauneau, A.; Esquerré-Tugayé, M.T.; Pont-Lezica, R. Cell wall proteins in apoplastic fluids of Arabidopsis thaliana rosettes: Identification by mass spectrometry and bioinformatics. Proteomics 2005, 5, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Zarei, A.; Trobacher, C.P.; Cooke, A.R.; Meyers, A.J.; Hall, J.C.; Shelp, B.J. Apple fruit copper amine oxidase isoforms: Peroxisomal MdAO1 prefers diamines as substrates, whereas extracellular MdAO2 exclusively utilizes monoamines. Plant Cell Physiol. 2015, 56, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Laurenzi, M.; Tipping, A.J.; Marcus, S.E.; Knox, J.P.; Federico, R.; Angelini, R.; McPherson, M.J. Analysis of the distribution of copper amine oxidase in cell walls of legume seedlings. Planta 2001, 214, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Padiglia, A.; Medda, R.; Scanu, T.; Longu, S.; Rossi, A.; Floris, G. Structure and nucleotide sequence of Euphorbia characias copper/TPQ-containing amine oxidase gene. J. Protein Chem. 2002, 21, 435–441. [Google Scholar] [CrossRef]
- Rossi, A.; Petruzzelli, R.; Agrò, A.F. cDNA-derived amino-acid sequence of lentil seedlings’ amine oxidase. FEBS Lett. 1992, 301, 253–257. [Google Scholar] [CrossRef]
- An, Z.; Jing, W.; Liu, Y.; Zhang, W. Hydrogen peroxide generated by copper amine oxidase is involved in abscisic acid-induced stomatal closure in Vicia faba. J. Exp. Bot. 2008, 59, 815–825. [Google Scholar] [CrossRef]
- Fukao, Y.; Hayashi, M.; Hara-Nishimura, I.; Nishimura, M. Novel glyoxysomal protein kinase, GPK1, identified by proteomic analysis of glyoxysomes in etiolated cotyledons of Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 1002–1012. [Google Scholar] [CrossRef]
- Carter, C.; Pan, S.; Zouhar, J.; Avila, E.L.; Girke, T.; Raikhel, N.V. The vegetative vacuole proteome of Arabidopsis thaliana reveals predicted and unexpected proteins. Plant Cell 2004, 16, 3285–3303. [Google Scholar] [CrossRef]
- Pietrangeli, P.; Federico, R.; Mondovì, B.; Morpurgo, L. Substrate specificity of copper-containing plant amine oxidases. J. Inorg. Biochem. 2007, 101, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Padiglia, A.; Floris, G.; Longu, S.; Schininà, M.E.; Pedersen, J.Z.; Finazzi Agrò, A.; De Angelis, F.; Medda, R. Inhibition of lentil copper/TPQ amine oxidase by the mechanism-based inhibitor derived from tyramine. Biol. Chem. 2004, 385, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Katoh, A.; Shoji, T.; Hashimoto, T. Molecular cloning of N-Methyl putrescine oxidase from tobacco. Plant Cell Physiol. 2007, 48, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Marina, M.; Maiale, S.J.; Rossi, F.R.; Romero, M.F.; Rivas, E.I.; Gárriz, A.; Ruiz, O.A.; Pieckenstain, F.L. Apoplastic polyamine oxidation plays different roles in local responses of tobacco to infection by the necrotrophic fungus Sclerotinia sclerotiorum and the biotrophic bacterium Pseudomonas viridiflava. Plant Physiol. 2008, 147, 2164–2178. [Google Scholar] [CrossRef] [PubMed]
- Šebela, M.; Brauner, F.; Radová, A.; Jacobsen, S.; Havlis, J.; Galuszka, P.; Pec, P. Characterisation of a homogeneous plant aminoaldehyde dehydrogenase. Biochim. Biophys. Acta 2000, 1480, 329–341. [Google Scholar] [CrossRef]
- Petřivalský, M.; Brauner, F.; Luhová, L.; Gagneul, D.; Šebela, M. Aminoaldehyde dehydrogenase activity during wound healing of mechanically injured pea seedlings. J. Plant Physiol. 2007, 164, 1410–1418. [Google Scholar] [CrossRef]
- Ghuge, S.A.; Carucci, A.; Rodrigues-Pousada, R.A.; Tisi, A.; Franchi, S.; Tavladoraki, P.; Angelini, R.; Cona, A. The MeJA-inducible copper amine oxidase AtAO1 is expressed in xylem tissue and guard cells. Plant Signal. Behav. 2015, 10, e1073872. [Google Scholar] [CrossRef]
- Fraudentali, I.; Rodrigues-Pousada, R.A.; Tavladoraki, P.; Angelini, R.; Cona, A. Leaf-Wounding Long-Distance Signaling Targets AtCuAOβ Leading to Root Phenotypic Plasticity. Plants 2020, 9, 249. [Google Scholar] [CrossRef]
- Qu, Y.; Wang, Q.; Guo, J.; Wang, P.; Song, P.; Jia, Q.; Zhang, X.; Kudla, J.; Zhang, W.; Zhang, Q. Peroxisomal CuAOζ and its product H2O2 regulate the distribution of auxin and IBA-dependent lateral root development in Arabidopsis. J. Exp. Bot. 2017, 68, 4851–4867. [Google Scholar] [CrossRef]
- Ghuge, S.A.; Carucci, A.; Rodrigues-Pousada, R.A.; Tisi, A.; Franchi, S.; Tavladoraki, P.; Angelini, R.; Cona, A. The apoplastic copper AMINE OXIDASE1 mediates jasmonic acid-induced protoxylem differentiation in Arabidopsis roots. Plant Physiol. 2015, 168, 690–707. [Google Scholar] [CrossRef]
- Fraudentali, I.; Ghuge, S.A.; Carucci, A.; Tavladoraki, P.; Angelini, R.; Cona, A.; Rodrigues-Pousada, R.A. The Copper Amine Oxidase Atcuaoδ participates in Abscisic Acid-Induced Stomatal Closure in Arabidopsis. Plants 2019, 8, 183. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.; Cowley, T.; Mitchell, A. Methyl jasmonate alters polyamine metabolism and induces systemic protection against powdery mildew infection in barley seedlings. J. Exp. Bot. 2002, 53, 747–756. [Google Scholar] [CrossRef]
- Biondi, S.; Scaramagli, S.; Capitani, F.; Altamura, M.M.; Torrigiani, P. Methyl jasmonate upregulates biosynthetic gene expression, oxidation and conjugation of polyamines, and inhibits shoot formation in tobacco thin layers. J. Exp. Bot. 2001, 52, 231–242. [Google Scholar] [CrossRef]
- Rea, G.; Metoui, O.; Infantino, A.; Federico, R.; Angelini, R. Copper amine oxidase expression in defense responses to wounding and Ascochyta rabiei invasion. Plant Physiol. 2002, 128, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Moschou, P.N.; Paschalidis, K.A.; Delis, I.D.; Andriopoulou, A.H.; Lagiotis, G.D.; Yakoumakis, D.I.; Roubelakis-Angelakis, K.A. Spermidine exodus and oxidation in the apoplast induced by abiotic stress is responsible for H2O2 signatures that direct tolerance responses in tobacco. Plant Cell. 2008, 20, 1708–1724. [Google Scholar] [CrossRef]
- Xing, S.G.; Jun, Y.B.; Hau, Z.W.; Liang, L.Y. Higher accumulation of gamma-aminobutyric acid induced by salt stress through stimulating the activity of diamine oxidases in Glycine max (L.) Merr. roots. Plant Physiol. Biochem. 2007, 45, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Wimalasekera, R.; Villar, C.; Begum, T.; Scherer, G.F.E. COPPER AMINE OXIDASE1 (CuAO1) of Arabidopsis thaliana contributes to abscisic acid- and polyamine-induced nitric oxide biosynthesis and abscisic acid signal transduction. Mol. Plant. 2011, 4, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Flores, T.; Todd, C.D.; Tovar-Mendez, A.; Dhanoa, P.K.; Correa-Aragunde, N.; Hoyos, M.E.; Brownfield, D.M.; Mullen, R.T.; Lamattina, L.; Polacco, J.C. Arginase-negative mutants of Arabidopsis exhibit increased nitric oxide signaling in root development. Plant Physiol. 2008, 147, 1936–1946. [Google Scholar] [CrossRef] [PubMed]
- Torrigiani, P.; Serafini-Fracassini, D.; Fara, A. Diamine Oxidase Activity in Different Physiological Stages of Helianthus tuberosus Tuber. Plant Physiol. 1989, 89, 69–73. [Google Scholar] [CrossRef]
- Gemperlová, L.; Eder, J.; Cvikrová, M. Polyamine metabolism during the growth cycle of tobacco BY-2 cells. Plant Physiol. Biochem. 2005, 43, 375–381. [Google Scholar] [CrossRef]
- Paschalidis, K.A.; Roubelakis-Angelakis, K.A. Sites and regulation of polyamine catabolism in the tobacco plant. Correlations with cell division/expansion, cell cycle progression, and vascular development. Plant Physiol. 2005, 138, 2174–2184. [Google Scholar] [CrossRef]
- Alharbi, B.; Hunt, J.D.; Dimitrova, S.; Spadafora, N.D.; Cort, A.P.; Colombo, D.; Müller, C.T.; Ghuge, S.A.; Davoli, D.; Cona, A.; et al. Mutation of Arabidopsis copper-containing amine oxidase gene AtCuAOδ alters polyamines, reduces gibberellin content and affects development. Int. J. Mol. Sci. 2020, 21, 7789. [Google Scholar] [CrossRef]
- Hatmi, S.; Villaume, S.; Trotel-Aziz, P.; Barka, E.A.; Clément, C.; Aziz, A. Osmotic Stress and ABA Affect Immune Response and Susceptibility of Grapevine Berries to Gray Mold by Priming Polyamine Accumulation. Front. Plant Sci. 2018, 9, 1010. [Google Scholar] [CrossRef]
- Rodríguez, A.A.; Maiale, S.J.; Menéndez, A.B.; Ruiz, O.A. Polyamine oxidase activity contributes to sustain maize leaf elongation under saline stress. J. Exp. Bot. 2009, 60, 4249–4262. [Google Scholar] [CrossRef] [PubMed]
- Rea, G.; de Pinto, M.C.; Tavazza, R.; Biondi, S.; Gobbi, V.; Ferrante, P.; de Gara, L.; Federico, R.; Angelini, R.; Tavladoraki, P. Ectopic expression of maize polyamine oxidase and pea copper amine oxidase in the cell wall of tobacco plants. Plant Physiol. 2004, 134, 1414–1426. [Google Scholar] [CrossRef] [PubMed]
- Cona, A.; Tisi, A.; Ghuge, S.A.; Franchi, S.; De Lorenzo, G.; Angelini, R. Wound healing response and xylem differentiation in tobacco plants over-expressing a fungal endopolygalacturonase is mediated by copper amine oxidase activity. Plant Physiol. Biochem. 2014, 82, 54–65. [Google Scholar] [CrossRef]
- Huang, A.X.; Wang, Y.S.; She, X.P.; Mu, J.; Zhao, J.L. Copper amine oxidase-catalysed hydrogen peroxide involves production of nitric oxide in darkness-induced stomatal closure in broad bean. Funct. Plant Biol. 2015, 42, 1057–1067. [Google Scholar] [CrossRef]
- Liu, C.; Atanasov, K.E.; Arafaty, N.; Murillo, E.; Tiburcio, A.F.; Zeier, J.; Alcázar, R. Putrescine elicits ROS-dependent activation of the salicylic acid pathway in Arabidopsis thaliana. Plant Cell Environ. 2020, 43, 2755–2768. [Google Scholar] [CrossRef] [PubMed]
- Tisi, A.; Federico, R.; Moreno, S.; Lucretti, S.; Moschou, P.N.; Roubelakis-Angelakis, K.A.; Angelini, R.; Cona, A. Perturbation of polyamine catabolism can strongly affect root development and xylem differentiation. Plant Physiol. 2011, 157, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Ghuge, S.A.; Tisi, A.; Carucci, A.; Rodrigues-Pousada, R.A.; Franchi, S.; Tavladoraki, P.; Angelini, R.; Cona, A. Cell wall amine oxidases: New players in root xylem differentiation under stress conditions. Plants 2015, 4, 489–504. [Google Scholar] [CrossRef]
- Cui, X.; Ge, C.; Wang, R.; Wang, H.; Chen, W.; Fu, Z.; Jiang, X.; Li, J.; Wang, Y. The BUD2 mutation affects plant architecture through altering cytokinin and auxin responses in Arabidopsis. Cell Res. 2010, 20, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Baima, S.; Forte, V.; Possenti, M.; Peñalosa, A.; Leoni, G.; Salvi, S.; Felici, B.; Ruberti, I.; Morell, G. Negative feedback regulation of auxin signaling by ATHB8/ACL5 BUD2 transcription module. Mol. Plant. 2014, 7, 1006–1025. [Google Scholar] [CrossRef] [PubMed]
- Salvi, E.; Rutten, J.P.; Di Mambro, R.; Polverari, L.; Licursi, V.; Negri, R.; Dello Ioio, R.; Sabatini, S.; Ten Tusscher, K. A Self-Organized PLT/Auxin/ARR-B Network Controls the Dynamics of Root Zonation Development in Arabidopsis thaliana. Dev. Cell 2020, 53, 431–443.e23. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T. Root Development: A Go-Faster Stripe and Spoilers. Dev. Cell 2020, 53, 372–374. [Google Scholar] [CrossRef] [PubMed]
- Rost, T.L.; Baum, S. On the correlation of primary root length, meristem size and protoxylem tracheary element position in pea seedlings. Am. J. Bot. 1988, 75, 414–424. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef]
- Zhou, X.; Xiang, Y.; Li, C.; Yu, G. Modulatory Role of Reactive Oxygen Species in Root Development in Model Plant of Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 485932. [Google Scholar] [CrossRef]
- Fraudentali, I.; Rodrigues-Pousada, R.A.; Volpini, A.; Tavladoraki, P.; Angelini, R.; Cona, A. Stress-Triggered Long-Distance Communication Leads to Phenotypic Plasticity: The Case of the Early Root Protoxylem Maturation Induced by Leaf Wounding in Arabidopsis. Plants 2018, 7, 107. [Google Scholar] [CrossRef]
- Aloni, R. The induction of vascular tissues by auxin and cytokinin. In Plant Hormones: Physiology, Biochemistry and Molecular Biology, 2nd ed.; Davies, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 531–546. [Google Scholar]
- Aloni, R. Foliar and axial aspects of vascular differentiation: Hypotheses and evidence. J. Plant Growth Regul. 2001, 20, 22–34. [Google Scholar] [CrossRef]
- Aloni, R.; Schwalm, K.; Langhans, M.; Ullrich, C.I. Gradual shifts in sites of free-auxin production during leaf-primordium development and their role in vascular differentiation and leaf morphogenesis in Arabidopsis. Planta 2003, 216, 841–853. [Google Scholar] [CrossRef]
- Jacobs, J.; Roe, J.L. SKS6, a multicopper oxidase-like gene, participates in cotyledon vascular patterning during Arabidopsis thaliana development. Planta 2005, 222, 652–666. [Google Scholar] [CrossRef] [PubMed]
- Møller, S.G.; Urwin, P.E.; Atkinson, H.J.; McPherson, M.J. Nematode induced expression of atao1, a gene encoding an extracellular diamine oxidase associated with developing vascular tissue. Physiol. Mol. Plant Pathol. 1998, 53, 73–79. [Google Scholar] [CrossRef]
- Hilleary, R.; Gilroy, S. Systemic signaling in response to wounding and pathogens. Curr. Opin. Plant Biol. 2018, 43, 57–62. [Google Scholar] [CrossRef]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef] [PubMed]
- Heyman, J.; Canher, B.; Bisht, A.; Christiaens, F.; De Veylder, L. Emerging role of the plant ERF transcription factors in coordinating wound defense responses and repair. J Cell Sci. 2018, 29, 131–132. [Google Scholar] [CrossRef]
- Lee, H.J.; Park, J.S.; Shin, S.Y.; Kim, S.G.; Lee, G.; Kim, H.S.; Jeon, J.H.; Cho, H.S. Submergence deactivates wound-induced plant defence against herbivores. Commun. Biol. 2020, 3, 651. [Google Scholar] [CrossRef]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, I.S.; Scherer, G.F.E. Polyamines induce rapid biosynthesis of nitric oxide (NO) in Arabidopsis thaliana seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Tebartz, F.; Scherer, G.F.E. Polyamines, polyamines oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 2011, 181, 593–603. [Google Scholar] [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Courtois, C.; Besson, A.; Dahan, J.; Bourque, S.; Dobrowolska, G.; Pugin, A.; Wendehenne, D. Nitric oxide signaling in plants: Interplays with Ca2+ and protein kinases. J. Exp. Bot. 2008, 59, 155–163. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CuAOs/Plant Sources | Subcellular Localization | Reported Physiological Functions |
|---|---|---|
| AtCuAOβ Arabidopsis | Apoplast [18,24] | |
| AtCuAOα2 Arabidopsis | Peroxisome [30] |
|
| AtCuAOγ1 Arabidopsis | Apoplast [17] | |
| AtCuAOδ Arabidopsis | Vacuole [31] | |
| AtCuAOζ Arabidopsis | Peroxisome [17,19] | |
| MdAO1 Malus domestica | Peroxisome [25] |
|
| MdAO2 Malus domestica | Apoplast [25] |
|
| PSAO Pisum sativum | Apoplast [14,26] | |
| ELAO Euphorbia characias | Apoplast [27] | - |
| LSAO Lens culinaris | Apoplast [14,26,28] |
|
| CaCuAO Cicer arietinum | Apoplast [14,26] | |
| GmCuAO Glycine max | - |
|
| HvCuAO Hordeum vulgare | - |
|
| HtCuAO Helianthus tuberosus | - |
|
| VfCuAO Vicia faba | Apoplast [29] |
|
| NtCuAO Nicotiana tabacum | - | |
| NtDAO1 Nicotiana tabacum | Peroxisome [19] |
|
| NtMPO1 Nicotiana tabacum | Peroxisome [19] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraudentali, I.; Rodrigues-Pousada, R.A.; Angelini, R.; Ghuge, S.A.; Cona, A. Plant Copper Amine Oxidases: Key Players in Hormone Signaling Leading to Stress-Induced Phenotypic Plasticity. Int. J. Mol. Sci. 2021, 22, 5136. https://doi.org/10.3390/ijms22105136
Fraudentali I, Rodrigues-Pousada RA, Angelini R, Ghuge SA, Cona A. Plant Copper Amine Oxidases: Key Players in Hormone Signaling Leading to Stress-Induced Phenotypic Plasticity. International Journal of Molecular Sciences. 2021; 22(10):5136. https://doi.org/10.3390/ijms22105136
Chicago/Turabian StyleFraudentali, Ilaria, Renato A. Rodrigues-Pousada, Riccardo Angelini, Sandip A. Ghuge, and Alessandra Cona. 2021. "Plant Copper Amine Oxidases: Key Players in Hormone Signaling Leading to Stress-Induced Phenotypic Plasticity" International Journal of Molecular Sciences 22, no. 10: 5136. https://doi.org/10.3390/ijms22105136
APA StyleFraudentali, I., Rodrigues-Pousada, R. A., Angelini, R., Ghuge, S. A., & Cona, A. (2021). Plant Copper Amine Oxidases: Key Players in Hormone Signaling Leading to Stress-Induced Phenotypic Plasticity. International Journal of Molecular Sciences, 22(10), 5136. https://doi.org/10.3390/ijms22105136