Curcumin Prevents Cerebellar Hypoplasia and Restores the Behavior in Hyperbilirubinemic Gunn Rat by a Pleiotropic Effect on the Molecular Effectors of Brain Damage

 ,
,  ,
,  and
and
Abstract
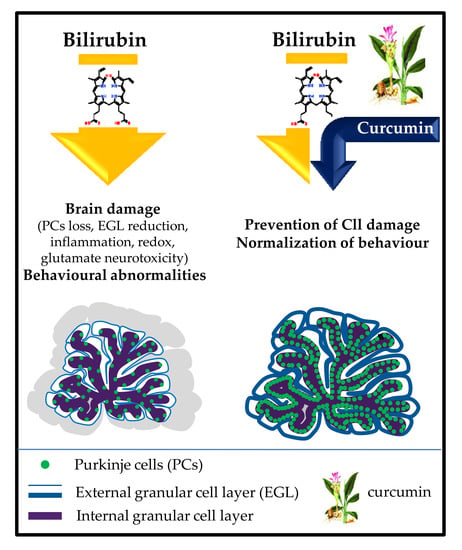
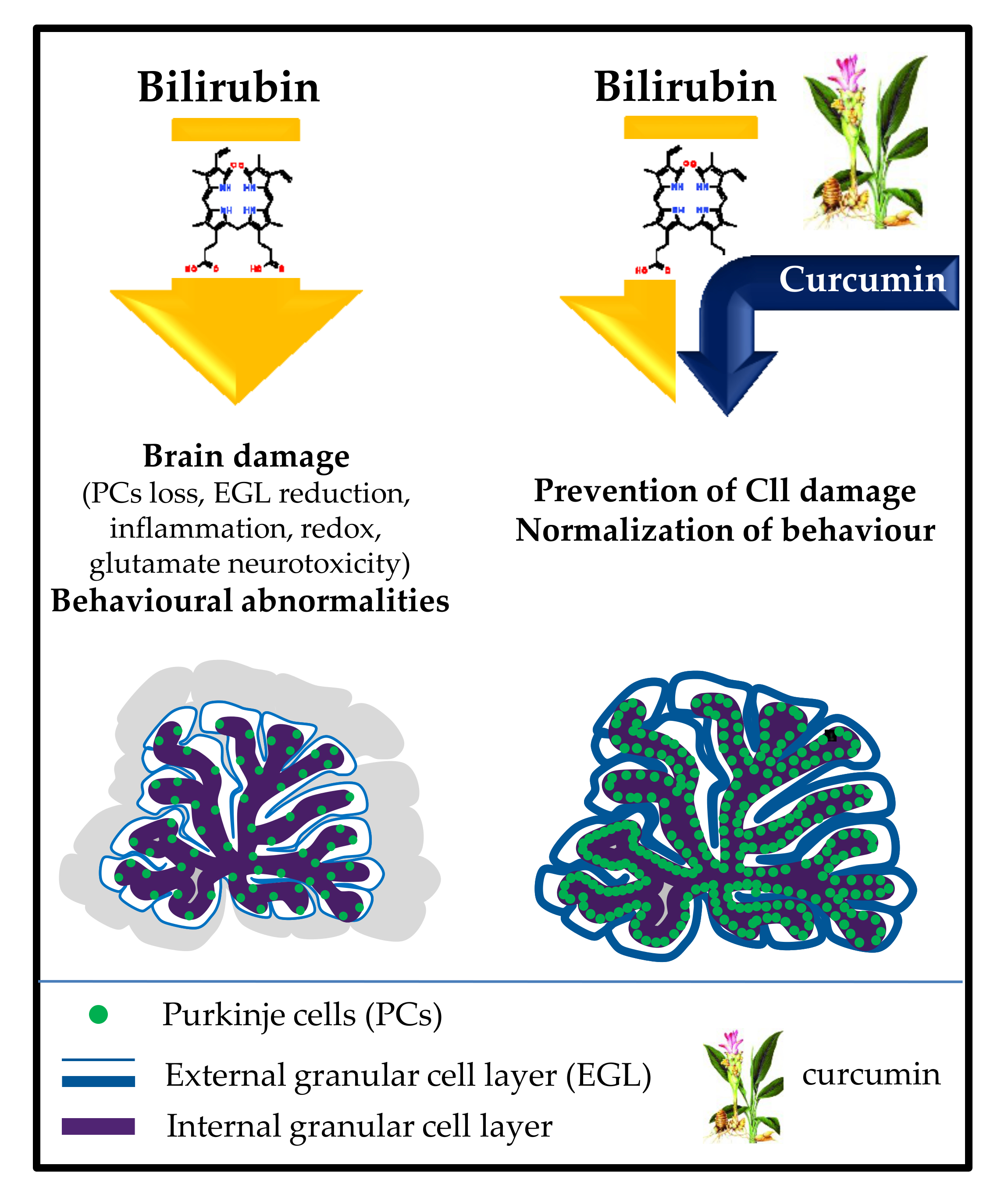
1. Introduction
2. Results
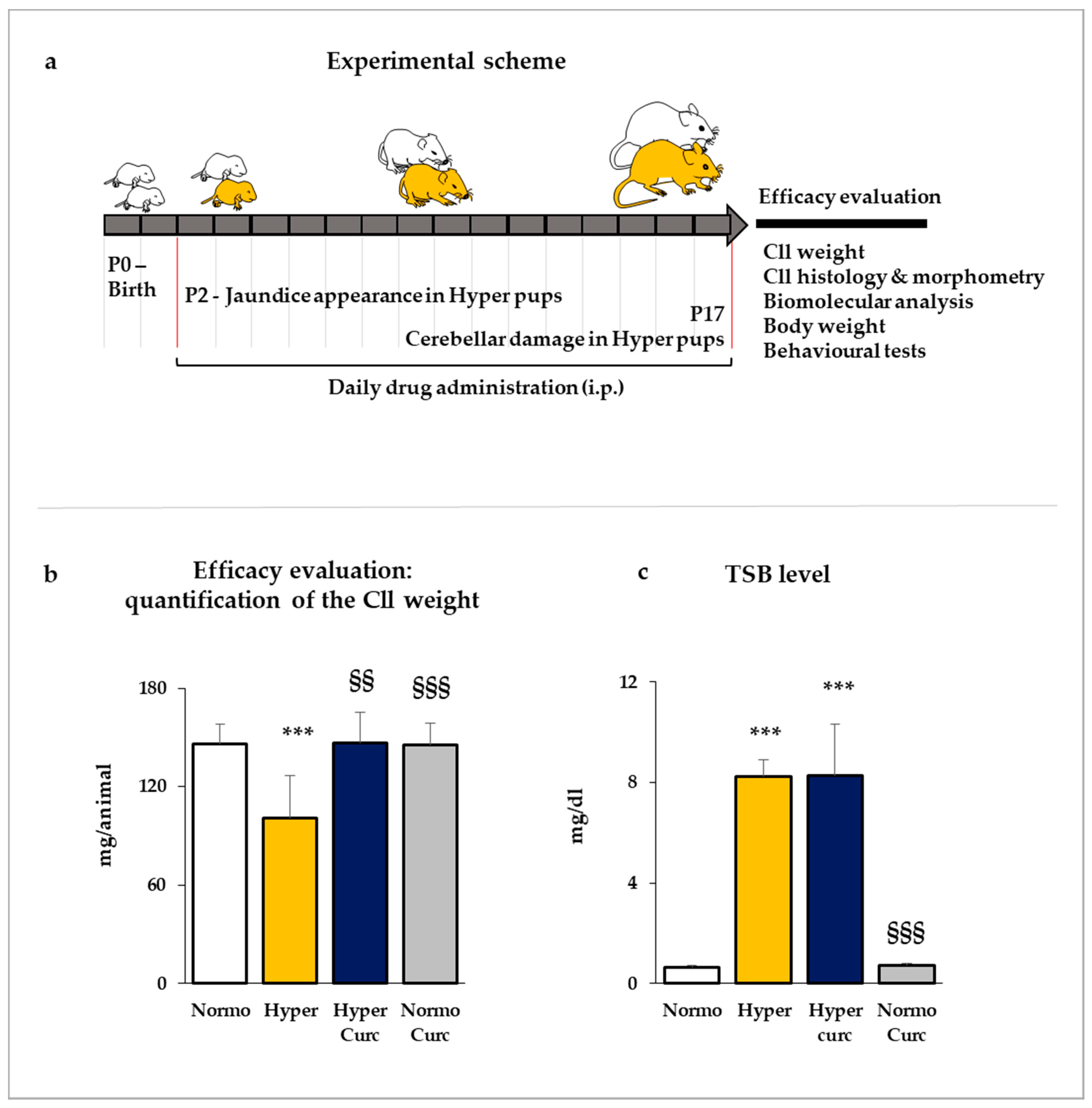
2.1. Evaluation of the Efficacy of Curcumin in Preventing the Cerebellar Damage in the Gunn Rat
2.1.1. Evaluation of the Cll Weight
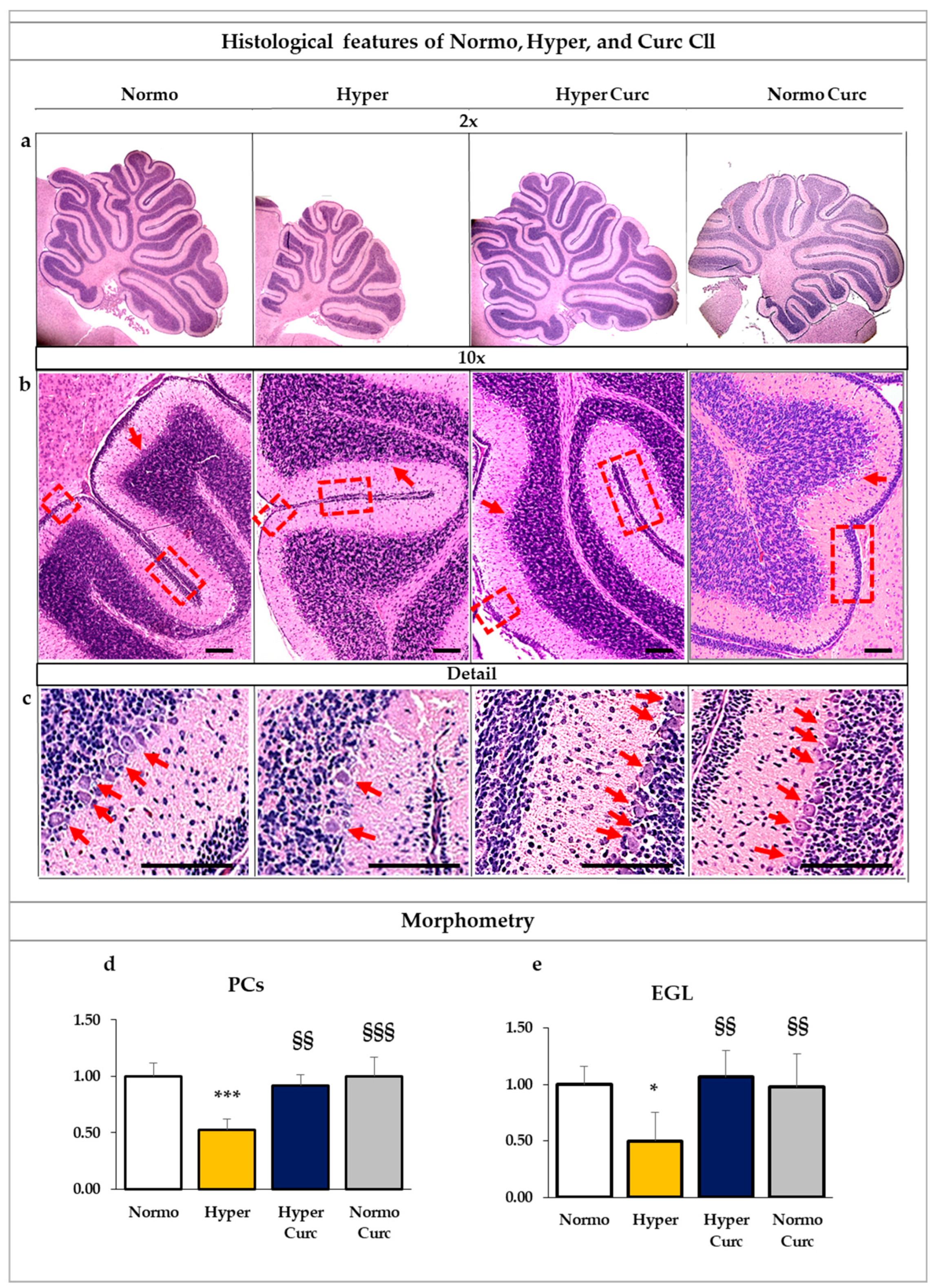
2.1.2. Assessment of the Histological Findings of the Cll under the Curc Treatment
2.1.3. Total Serum Bilirubin Quantification
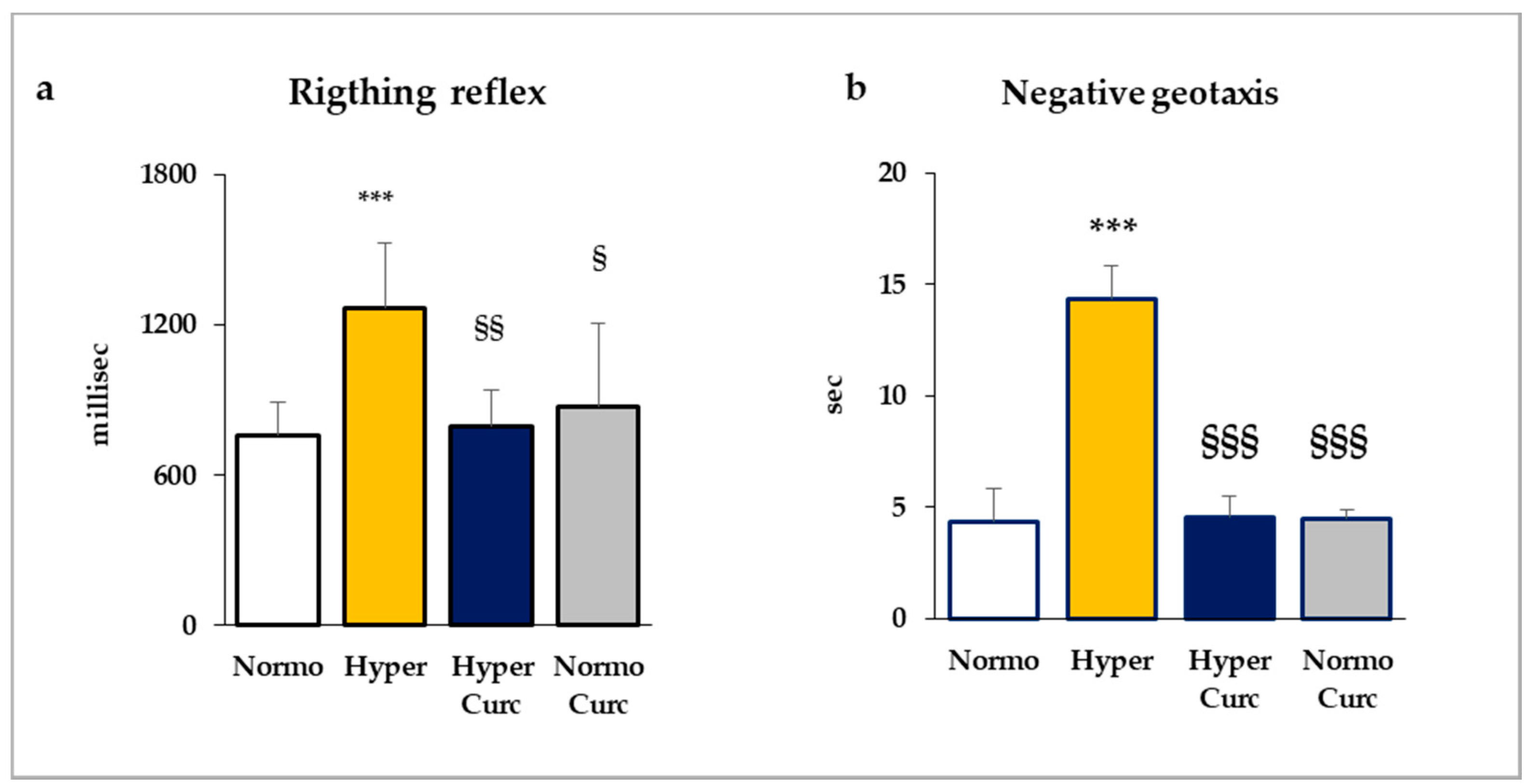
2.2. Behavioral Tests
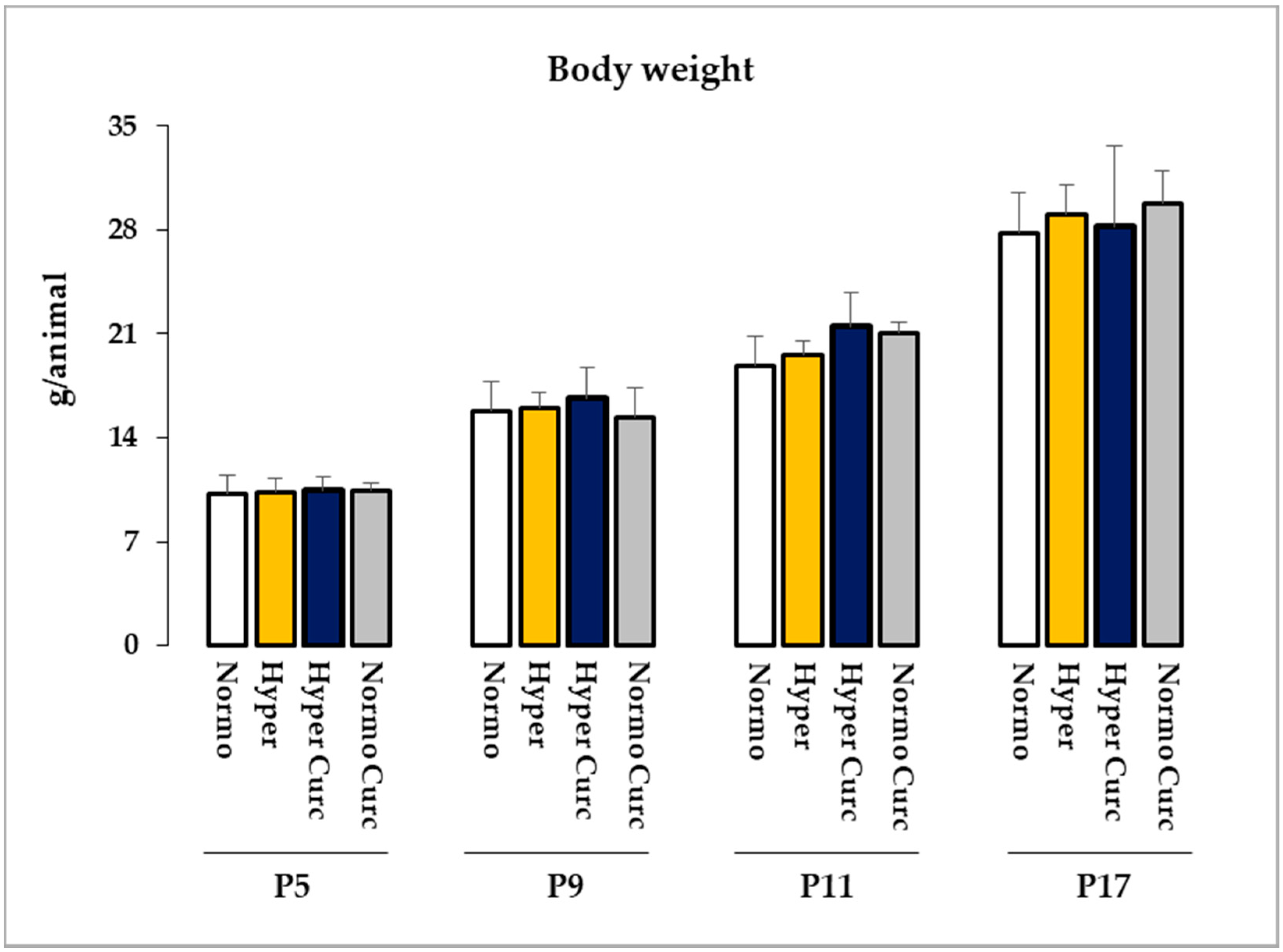
2.3. Monitoring of the Side Effects of the Treatment
2.4. Effects of Curc on the Main Molecular Effectors Involved in Bilirubin Brain Damage
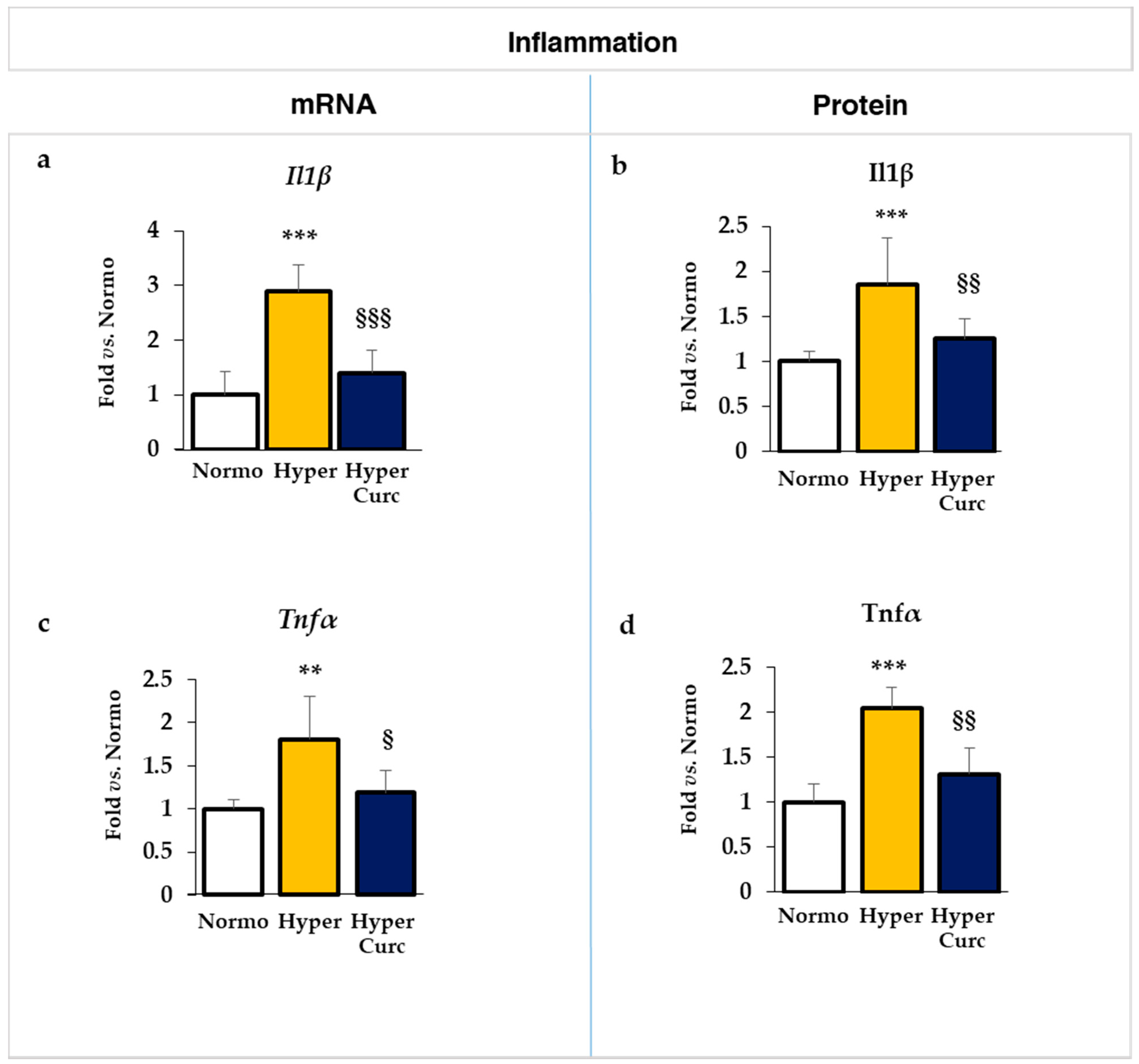
2.4.1. Evaluation of the Effect of Curc on the Inflammatory Markers
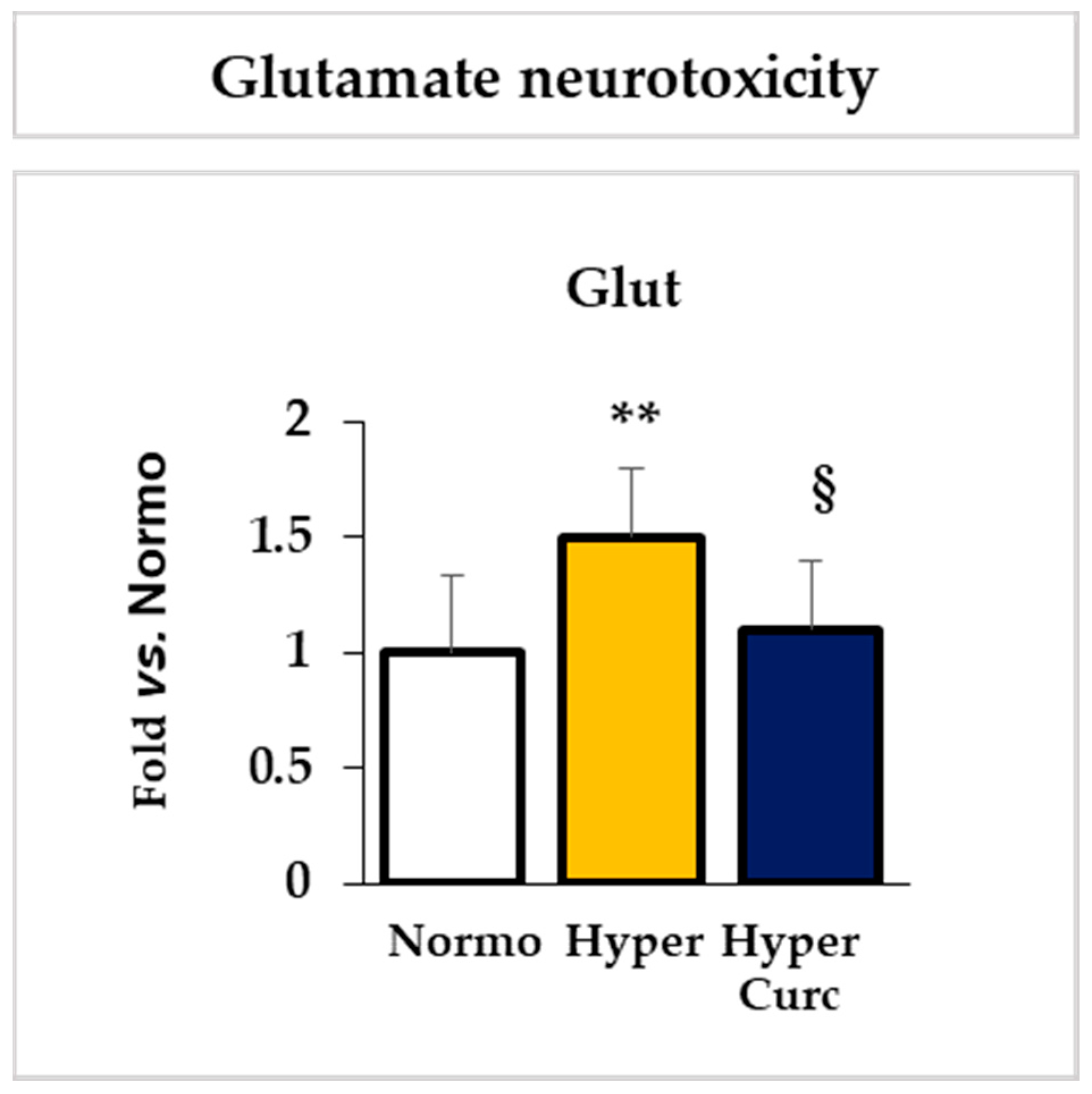
2.4.2. Evaluation of the Effect of Curc on the Glutamate Cll Content
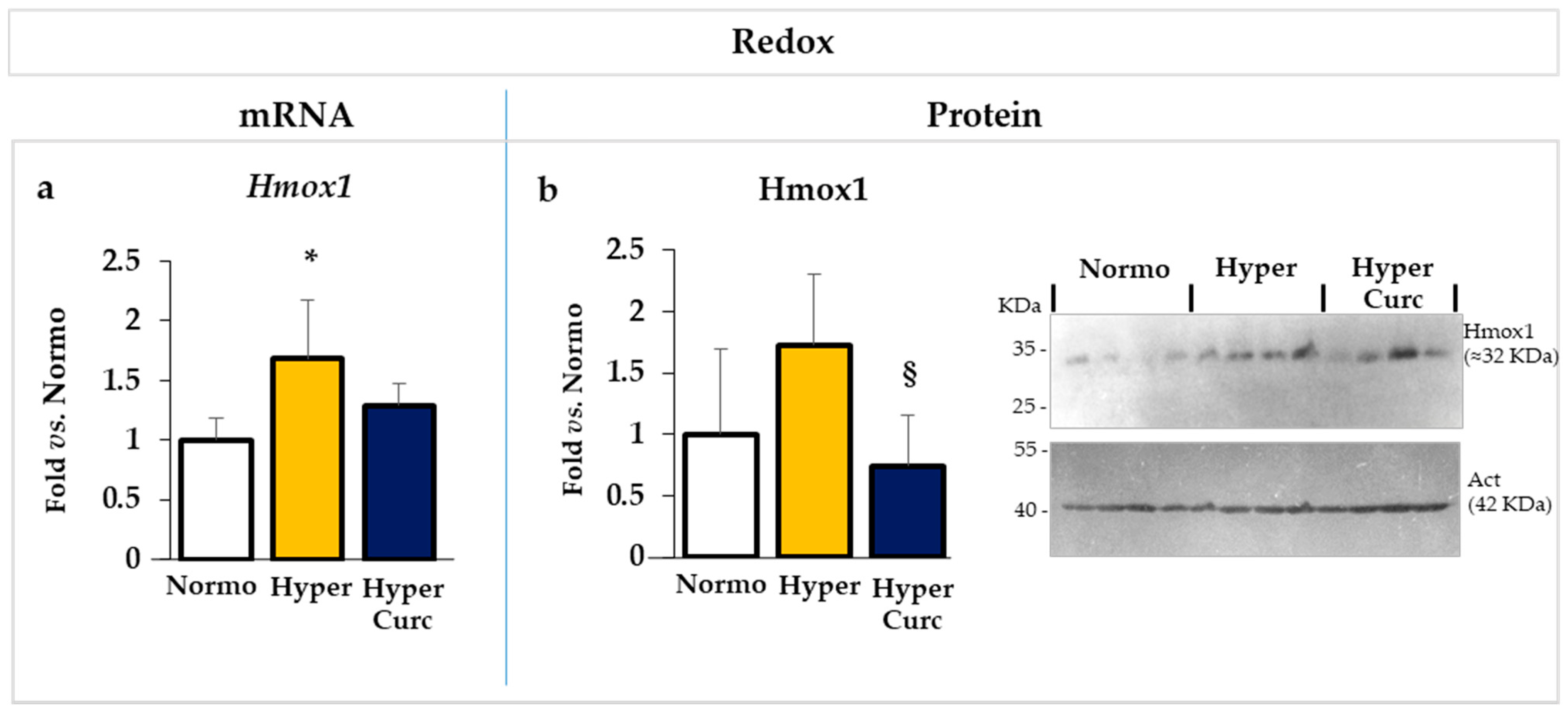
2.4.3. Evaluation of the Effect of Curc on the Redox Markers
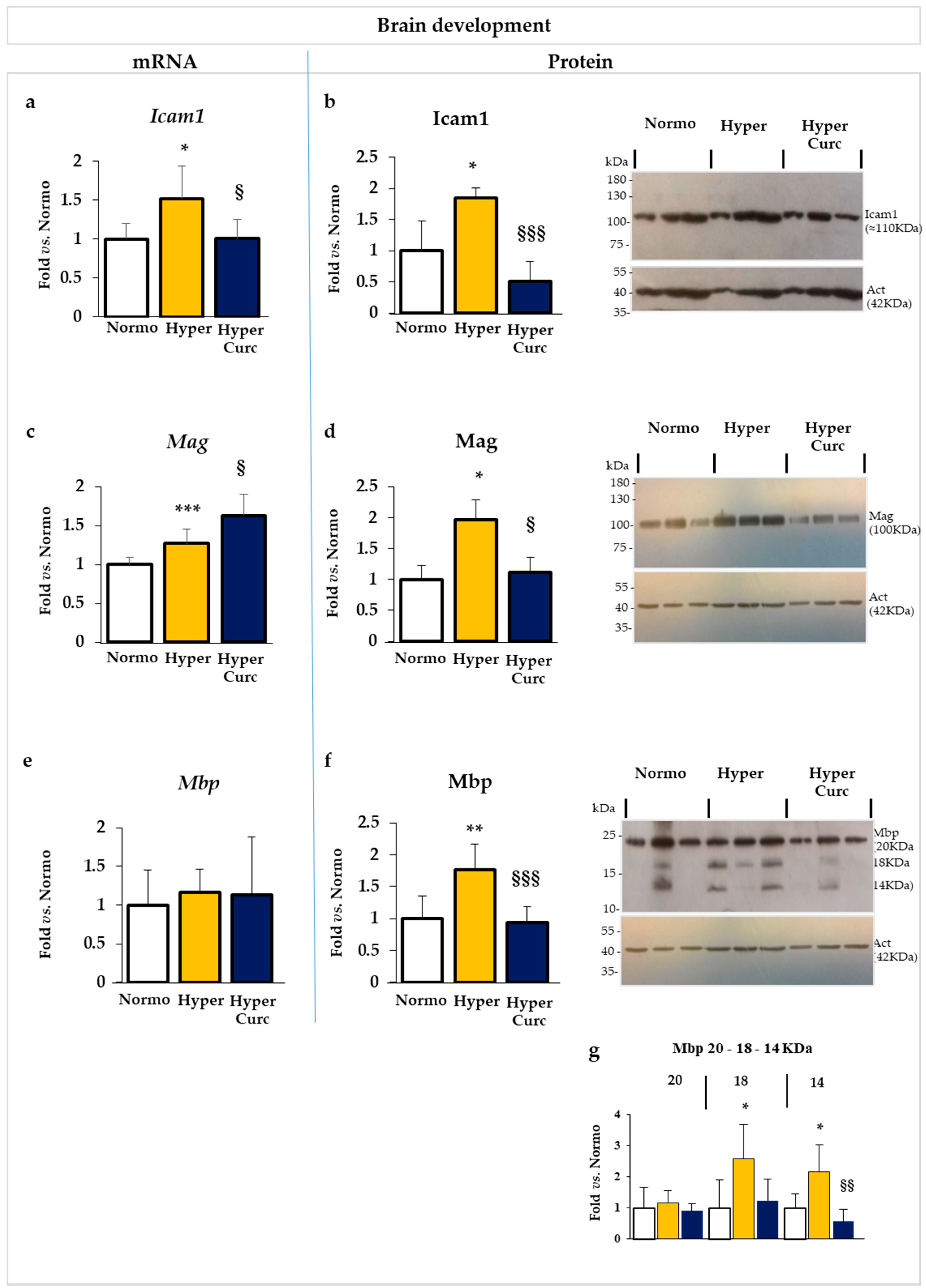
2.5. Additional Evaluation of Curc Protection: Selected Markers of Brain Development
3. Discussion
4. Materials and Methods
4.1. Litter Composition and Treatment Scheme
4.2. Cll Weight
4.3. Cll histology and Morphometry
4.4. Total Serum Bilirubin (TSB)
4.5. Behavioral Tests
4.6. Real-Time PCR of Selected Markers for Inflammation, Redox Imbalance, and Brain Development
4.7. Glutamate Quantification
4.8. Protein Evaluation of Selected Markers for Inflammation, Redox Imbalance, and Brain Development
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Bf | free bilirubin |
| bp | base pairs |
| Cll | cerebellum |
| Curc | curcumin |
| EGL | external granular layer |
| ELISA | enzyme-linked immunosorbent assay |
| Glut | glutamate |
| Hmox1 | heme oxygenase 1 |
| Hyper | Hyperbilirubinemic Gunn pups |
| Hyper Curc | Hyperbilirubinemic Gunn pups treated with curcumin |
| Icam1 | intracellular adhesion molecule |
| Il1β | interleukin 1 beta |
| i.p. | intraperitoneal injection |
| Mag | Myelin associated glycoprotein |
| Mbp | Myelin basic protein |
| Normo | Normobilirubinemic Gunn pups |
| Normo Curc | Normobilirubinemic Gunn pups treated with curcumin |
| P | post-natal age in days |
| PT | phototherapy |
| PCs | Purkinje cells |
| RTqPCR | Real-Time PCR |
| Tnfα | tumor necrosis factor alpha |
| TSB | Total serum bilirubin |
| KSD | kernicterus spectrum disorder |
References
- Pichon, J.-B.L.; Riordan, S.M. The Neurological Sequelae of Neonatal Hyperbilirubinemia: Definitions, Diagnosis and Treatment of the Kernicterus Spectrum Disorders (KSDs). Curr. Pediatric Rev. 2017, 13. [Google Scholar] [CrossRef]
- Hegyi, T.; Kleinfeld, A.; Huber, A.; Weinberger, B.; Memon, N.; Shih, W.J.; Carayannopoulos, M.; Oh, W. Unbound bilirubin measurements by a novel probe in preterm infants. J. Matern. Neonatal Med. 2019, 32, 2721–2726. [Google Scholar] [CrossRef] [PubMed]
- Riordan, S.M.; Bittel, D.C.; Le Pichon, J.-B.; Gazzin, S.; Tiribelli, C.; Watchko, J.F.; Wennberg, R.P.; Shapiro, S. A Hypothesis for Using Pathway Genetic Load Analysis for Understanding Complex Outcomes in Bilirubin Encephalopathy. Front. Neurosci. 2016, 10, 376. [Google Scholar] [CrossRef]
- Riordan, S.M.; Gazzin, S. Where do we stand in the field of neonatal jaundice? Commentary on the 2017 J. Donald Ostrow Trieste Yellow Retreat. Pediatr. Res. 2018, 83, 1090–1092. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.; Riordan, S.M. Review of bilirubin neurotoxicity II: Preventing and treating acute bilirubin encephalopathy and kernicterus spectrum disorders. Pediatr. Res. 2019, 87, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Olusanya, B.O.; Teeple, S.; Kassebaum, N.J. The Contribution of Neonatal Jaundice to Global Child Mortality: Findings from the GBD 2016 Study. Pediatrics 2018, 141. [Google Scholar] [CrossRef] [PubMed]
- Hyperbilirubinemia, S.O. Improvement on Practice Parameter: Management of Hyperbilirubinemia in the Healthy Term Newborn. Pediatrics 1994, 94, 558–565. [Google Scholar]
- Bhutani, V.K.; Johnson, L. Kernicterus in the 21st century: Frequently asked questions. J. Perinatol. 2009, 29, S20–S24. [Google Scholar] [CrossRef]
- Watchko, J.F. Refractory Causes of Kernicterus in Developed Countries: Can We Eradicate G6PD Deficiency Triggered and Low-Bilirubin Kernicterus? Curr. Pediatric Rev. 2017, 13. [Google Scholar] [CrossRef]
- Watchko, J.F.; Maisels, M.J. The enigma of low bilirubin kernicterus in premature infants: Why does it still occur, and is it preventable? Semin. Perinatol. 2014, 38, 397–406. [Google Scholar] [CrossRef]
- Watchko, J.F.; Spitzer, A.R.; Clark, R.H. Prevalence of Hypoalbuminemia and Elevated Bilirubin/Albumin Ratios in a Large Cohort of Infants in the Neonatal Intensive Care Unit. J. Pediatr. 2017, 188, 280–286.e4. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, T.; Kleinfeld, A.; Huber, A.; Weinberger, B.; Memon, N.; Shih, W.J.; Carayannopoulos, M.; Oh, W. Effects of Soybean Lipid Infusion on Unbound Free Fatty Acids and Unbound Bilirubin in Preterm Infants. J. Pediatr. 2017, 184, 45–50.e1. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hegyi, T.; Kathiravan, S.; Stahl, G.E.; Huber, A.H.; Kleinfeld, A. Unbound Free Fatty Acids from Preterm Infants Treated with Intralipid Decouples Unbound from Total Bilirubin Potentially Making Phototherapy Ineffective. Neonatology 2013, 104, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, T.; Kleinfeld, A.; Huber, A.; Weinberger, B.; Memon, N.; Carayannopoulos, M.; Oh, W. Unbound Bilirubin Levels in Phototherapy-treated Preterm Infants Receiving Soy-based Lipid Emulsion. Pediatr. Int. 2020. [Google Scholar] [CrossRef]
- Riordan, S.M.; Shapiro, S.M. Review of bilirubin neurotoxicity I: Molecular biology and neuropathology of disease. Pediatr. Res. 2019, 87, 327–331. [Google Scholar] [CrossRef]
- Sampurna, M.T.A.; Ratnasari, K.A.; Etika, R.; Hulzebos, C.V.; Dijk, P.H.; Bos, A.F.; Sauer, P.J.J. Adherence to hyperbilirubinemia guidelines by midwives, general practitioners, and pediatricians in Indonesia. PLoS ONE 2018, 13, e0196076. [Google Scholar] [CrossRef]
- Wennberg, R.P.; Watchko, J.F.; Shapiro, S. Maternal Empowerment—An underutilized strategy to prevent kernicterus? Curr. Pediatr. Rev. 2017, 13, 210–219. [Google Scholar] [CrossRef]
- Jašprová, J.J.; Dal Ben, M. Neuro-inflammatory effects of photodegradative products of bilirubin. Sci. Rep. 2018, 8, 7444. [Google Scholar] [CrossRef]
- Jasprova, J.; Ben, M.D.; Vianello, E.; Goncharova, I.; Urbanova, M.; Vyroubalova, K.; Gazzin, S.; Tiribelli, C.; Sticha, M.; Cerna, M.; et al. The Biological Effects of Bilirubin Photoisomers. PLoS ONE 2016, 11, e0148126. [Google Scholar] [CrossRef]
- Grosse, S.D.; Prosser, L.A. Screening for Neonatal Hyperbilirubinemia-First Do No Harm? JAMA Pediatr. 2019, 173, 617–618. [Google Scholar] [CrossRef]
- Hegyi, T.; Chefitz, D.; Weller, A.; Huber, A.; Carayannopoulos, M.; Kleinfeld, A. Unbound bilirubin measurements in term and late-preterm infants. J. Matern. Neonatal Med. 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dal Ben, M.; Bottin, C. Evaluation of region selective bilirubin-induced brain damage as a basis for a pharmacological treatment. Sci. Rep. 2017, 7, 41032. [Google Scholar] [CrossRef] [PubMed]
- Daood, M.J.; Hoyson, M. Lipid peroxidation is not the primary mechanism of bilirubin-induced neurologic dysfunction in jaun-diced Gunn rat pups. Pediatr. Res. 2012, 72, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Barateiro, A.; Vaz, A.R.; Silva, S.D.; Fernandes, A.; Brites, D. ER Stress, Mitochondrial Dysfunction and Calpain/JNK Activation are Involved in Oligodendrocyte Precursor Cell Death by Unconjugated Bilirubin. Neuromol. Med. 2012, 14, 285–302. [Google Scholar] [CrossRef]
- Barateiro, A.; Domingues, H.S. Rat cerebellar slice cultures exposed to bilirubin evidence reactive gliosis, excitotoxicity and im-paired myelinogenesis that is prevented by AMPA and TNF-α inhibitors. Mol. Neurobiol. 2014, 49, 424–439. [Google Scholar] [CrossRef]
- Brites, D.; Fernandes, A. Bilirubin-induced neural impairment: A special focus on myelination, age-related windows of suscepti-bility and associated co-morbidities. Semin. Fetal Neonatal Med. 2015, 20, 14–19. [Google Scholar] [CrossRef]
- Brito, M.A.; Zurolo, E.; Pereira, P.; Barroso, C.; Aronica, E.; Brites, D. Cerebellar Axon/Myelin Loss, Angiogenic Sprouting, and Neuronal Increase of Vascular Endothelial Growth Factor in a Preterm Infant with Kernicterus. J. Child Neurol. 2011, 27, 615–624. [Google Scholar] [CrossRef]
- Fernandes, A.; Silva, R.F.M. Cytokine production, glutamate release and cell death in rat cultured astrocytes treated with un-conjugated bilirubin and LPS. J. Neuroimmunol. 2004, 153, 64–75. [Google Scholar] [CrossRef]
- Fernandes, A.; Falcão, A.S.; Silva, R.F.M.; Gordo, A.C.; Gama, M.J.; Brito, M.A.; Brites, D. Inflammatory signalling pathways involved in astroglial activation by unconjugated bilirubin. J. Neurochem. 2006, 96, 1667–1679. [Google Scholar] [CrossRef]
- Fernandes, A.; Falcão, A.S.; Abranches, E.; Bekman, E.; Henrique, D.; Lanier, L.M.; Brites, D. Bilirubin as a determinant for altered neurogenesis, neuritogenesis, and synaptogenesis. Dev. Neurobiol. 2009, 69, 568–582. [Google Scholar] [CrossRef]
- Gambaro, S.E.; Robert, M.C.; Tiribelli, C.; Gazzin, S. Role of brain cytochrome P450 mono-oxygenases in bilirubin oxidation-specific induction and activity. Arch. Toxicol. 2014, 90, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Gazzin, S.; Zelenka, J.; Zdrahalova, L.; Koníčková, R.; Zabetta, C.C.; Giraudi, P.J.; Berengeno, A.L.; Raseni, A.; Robert, M.C.; Vítek, L.; et al. Bilirubin accumulation and Cyp mRNA expression in selected brain regions of jaundiced Gunn rat pups. Pediatr. Res. 2012, 71, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Grojean, S.; Koziel, V.; Vert, P.; Daval, J.-L. Bilirubin Induces Apoptosis via Activation of NMDA Receptors in Developing Rat Brain Neurons. Exp. Neurol. 2000, 166, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.W.; Bratlid, D. Bilirubin decreases phosphorylation of synapsin I, a synaptic vesicle-associated neuronal phospho-protein, in intact synaptosomes from rat cerebral cortex. Pediatr. Res. 1988, 23, 219–223. [Google Scholar] [CrossRef]
- A Johnson, J.; Hayward, J.J.; E Kornguth, S.; Siegel, F.L. Effects of hyperbilirubinaemia on glutathione S-transferase isoenzymes in cerebellar cortex of the Gunn rat. Biochem. J. 1993, 291, 453–461. [Google Scholar] [CrossRef][Green Version]
- McDonald, J.W.; Shapiro, S.M.; Silverstein, F.S.; Johnston, M.V. Role of Glutamate Receptor-Mediated Excitotoxicity in Bilirubin-Induced Brain Injury in the Gunn Rat Model. Exp. Neurol. 1998, 150, 21–29. [Google Scholar] [CrossRef]
- Qaisiya, M.; Zabetta, C.D.C.; Bellarosa, C.; Tiribelli, C. Bilirubin mediated oxidative stress involves antioxidant response activation via Nrf2 pathway. Cell. Signal. 2014, 26, 512–520. [Google Scholar] [CrossRef]
- Qaisiya, M.; Brischetto, C.; Jasprova, J.; Vítek, L.; Tiribelli, C.; Bellarosa, C. Bilirubin-induced ER stress contributes to the inflammatory response and apoptosis in neuronal cells. Arch. Toxicol. 2017, 91, 1847–1858. [Google Scholar] [CrossRef]
- Robert, M.C.; Furlan, G.; Rosso, N.; Gambaro, S.E.; Apitsionak, F.; Vianello, E.; Tiribelli, C.; Gazzin, S. Alterations in the Cell Cycle in the Cerebellum of Hyperbilirubinemic Gunn Rat: A Possible Link with Apoptosis? PLoS ONE 2013, 8, e79073. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; Solá, S.; Brites, D. Bilirubin induces apoptosis via the mitochondrial pathway in developing rat brain neurons. Hepatology 2002, 35, 1186–1195. [Google Scholar] [CrossRef]
- Silva, S.D.; Vaz, A.R.; Diógenes, M.J.; Van Rooijen, N.; Sebastião, A.M.; Fernandes, A.; Silva, R.F.M.; Brites, D. Neuritic growth impairment and cell death by unconjugated bilirubin is mediated by NO and glutamate, modulated by microglia, and prevented by glycoursodeoxycholic acid and interleukin. Neuropharmacology 2012, 62, 2398–2408. [Google Scholar] [CrossRef] [PubMed]
- Gunn, C.H. Hereditary acholuric jaundice in a New Mutant Strain of Rats. J. Hered. 1938, 29, 137–139. [Google Scholar] [CrossRef]
- Schutta, H.S.; Johnson, L. Bilirubin encephalopathy in the Gunn rat: A fine structure study of the cerebellar cortex. J. Neuropathol. Exp. Neurol. 1967, 26, 377–396. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, J.R.; Kondapalli, R. Gunn rat: A model for inherited deficiency of bilirubin glucuronidation. Adv. Vet. Sci. Comp. Med. 1993, 37, 149–173. [Google Scholar]
- Geiger, A.S.; Rice, A.C.; Shapiro, S.M. Minocycline Blocks Acute Bilirubin-Induced Neurological Dysfunction in Jaundiced Gunn Rats. Neonatology 2007, 92, 219–226. [Google Scholar] [CrossRef]
- Lin, S.; Wei, X.; Bales, K.R.; Paul, A.B.C.; Ma, Z.; Yan, G.; Paul, S.M.; Du, Y. Minocycline blocks bilirubin neurotoxicity and prevents hyperbilirubinemia-induced cerebellar hypoplasia in the Gunn rat. Eur. J. Neurosci. 2005, 22, 21–27. [Google Scholar] [CrossRef]
- Smith, K.; Leyden, J.J. Safety of doxycycline and minocycline: A systematic review. Clin. Ther. 2005, 27, 1329–1342. [Google Scholar] [CrossRef]
- Chen, K.; An, Y.A.; Tie, L.; Pan, Y.; Li, X. Curcumin Protects Neurons from Glutamate-Induced Excitotoxicity by Membrane Anchored AKAP79-PKA Interaction Network. Evid. Based Complement. Altern. Med. 2015, 2015, 1–9. [Google Scholar] [CrossRef]
- Khalil, R.M.; Khedr, N.F. Curcumin Protects against Monosodium Glutamate Neurotoxicity and Decreasing NMDA2B and mGluR5 Expression in Rat Hippocampus. Neurosignals 2016, 24, 81–87. [Google Scholar] [CrossRef]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic Roles of Curcumin: Lessons Learned from Clinical Trials. AAPS J. 2012, 15, 195–218. [Google Scholar] [CrossRef]
- Fadus, M.C.; Lau, C.; Bikhchandani, J.; Lynch, H.T. Curcumin: An age-old anti-inflammatory and anti-neoplastic agent. J. Tradit. Complement. Med. 2017, 7, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Heger, M.; van Golen, R.F. The molecular basis for the pharmacokinetics and pharmacodynamics of curcumin and its metabolites in relation to cancer. Pharmacol. Rev. 2014, 66, 222–307. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its’ Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Larasati, Y.A.; Yoneda-Kato, N.; Nakamae, I.; Yokoyama, T.; Meiyanto, E.; Kato, J.-Y. Curcumin targets multiple enzymes involved in the ROS metabolic pathway to suppress tumor cell growth. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Marchiani, A.; Rozzo, C. Curcumin and Curcumin-like Molecules: From Spice to Drugs. Curr. Med. Chem. 2013, 21. [Google Scholar] [CrossRef]
- Ramsewak, R.; DeWitt, D.; Nair, M. Cytotoxicity, antioxidant and anti-inflammatory activities of Curcumins I–III from Curcuma longa. Phytomedicine 2000, 7, 303–308. [Google Scholar] [CrossRef]
- Salehi, B.; Stojanović-Radić, Z.; Matejic, J.S.; Sharifi-Rad, M.; Kumar, N.V.A.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of curcumin: A review of clinical trials. Eur. J. Med. Chem. 2019, 163, 527–545. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lu, C.W.; Wang, C.-C.; Wang, Y.-C.; Wang, S.-J. Curcumin inhibits glutamate release in nerve terminals from rat prefrontal cortex: Possible relevance to its antidepressant mechanism. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2011, 35, 1785–1793. [Google Scholar] [CrossRef]
- Suskind, D.L.; Wahbeh, G. Tolerability of curcumin in pediatric inflammatory bowel disease: A forced-dose titration study. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 277–279. [Google Scholar] [CrossRef]
- European Medicine Agency. Assessment Report on Curcuma longa L. (C. Domestica Valeton), Rhizoma. 2017. Available online: https://www.ema.europa.eu/documents/herbal-report/draft-assessment-report-curcuma-longa-l-c-domestica-valeton-rhizome-revision-1_en.pdf (accessed on 25 September 2018).
- Begum, A.N.; Jones, M.R.; Lim, G.P.; Morihara, T.; Kim, P.; Heath, D.D.; Rock, C.L.; Pruitt, M.A.; Yang, F.; Hudspeth, B.; et al. Curcumin Structure-Function, Bioavailability, and Efficacy in Models of Neuroinflammation and Alzheimer’s Disease. J. Pharmacol. Exp. Ther. 2008, 326, 196–208. [Google Scholar] [CrossRef]
- Perkins, S.; Verschoyle, R.D.; Hill, K.; Parveen, I.; Threadgill, M.D.; Sharma, R.A.; Williams, M.L.; Steward, W.P.; Gescher, A.J. Chemopreventive efficacy and pharmacokinetics of curcumin in the min/+ mouse, a model of familial adenomatous polyposis. Cancer Epidemiol. Biomark. Prev. 2002, 11, 535–540. [Google Scholar]
- Ryu, E.K.; Choe, Y.S.; Lee, K.-H.; Choi, A.Y.; Kim, B.-T. Curcumin and Dehydrozingerone Derivatives: Synthesis, Radiolabeling, and Evaluation for β-Amyloid Plaque Imaging. J. Med. Chem. 2006, 49, 6111–6119. [Google Scholar] [CrossRef] [PubMed]
- Conlee, J.W.; Shapiro, S.M. Development of cerebellar hypoplasia in jaundiced Gunn rats: A quantitative light microscopic analysis. Acta Neuropathol. 1997, 93, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Daood, M.J.; Watchko, J.F. Calculated In Vivo Free Bilirubin Levels in the Central Nervous System of Gunn Rat Pups. Pediatr. Res. 2006, 60, 44–49. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sawasaki, Y.; Yamada, N.; Nakajima, H. Developmental features of cerebellar hypoplasia and brain bilirubin levels in a mutant (gunn) rat with hereditary hyperbilirubinaemia. J. Neurochem. 1976, 27, 577–583. [Google Scholar] [CrossRef]
- Vianello, E.; Zampieri, S.; Marcuzzo, T.; Tordini, F.; Bottin, C.; Dardis, A.; Zanconati, F.; Tiribelli, C.; Gazzin, S. Histone acetylation as a new mechanism for bilirubin-induced encephalopathy in the Gunn rat. Sci. Rep. 2018, 8, 13690. [Google Scholar] [CrossRef]
- Takagishi, Y.; Yamamura, H. Purkinje cell abnormalities and synaptogenesis in genetically jaundiced rats (Gunn rats). Brain Res. 1989, 492, 116–128. [Google Scholar] [CrossRef]
- Wolfensohn, S.E.; Lloyd, M. Practical use of distress scoring systems in the application of humane end points. In Humane Endpoints in Animal Experiments for Biomedical Research; Hendriksen, C., Morton, D., Eds.; Royal Society of Medicine Press: London, UK, 1999. [Google Scholar]
- Harauz, G.; Boggs, J.M. Myelin management by the 18.5-kDa and 21.5-kDa classic myelin basic protein isoforms. J. Neurochem. 2013, 125, 334–361. [Google Scholar] [CrossRef]
- Akiyama, K.; Ichinose, S.; Omori, A.; Sakurai, Y.; Asou, H. Study of expression of myelin basic proteins (MBPs) in developing rat brain using a novel antibody reacting with four major isoforms of MBP. J. Neurosci. Res. 2002, 68, 19–28. [Google Scholar] [CrossRef]
- Nguyen, T.; Mehta, N.R.; Conant, K.; Kim, K.-J.; Jones, M.; Calabresi, P.A.; Melli, G.; Hoke, A.; Schnaar, R.L.; Ming, G.-L.; et al. Axonal Protective Effects of the Myelin-Associated Glycoprotein. J. Neurosci. 2009, 29, 630–637. [Google Scholar] [CrossRef]
- Vodret, S.; Bortolussi, G.; Iaconcig, A.; Martinelli, E.; Tiribelli, C.; Muro, A. Attenuation of neuro-inflammation improves survival and neurodegeneration in a mouse model of severe neonatal hyperbilirubinemia. Brain Behav. Immun. 2018, 70, 166–178. [Google Scholar] [CrossRef]
- Kliem, C.; Merling, A.; Giaisi, M.; Köhler, R.; Krammer, P.H.; Li-Weber, M. Curcumin Suppresses T Cell Activation by Blocking Ca 2+ Mobilization and Nuclear Factor of Activated T Cells (NFAT) Activation. J. Biol. Chem. 2012, 287, 10200–10209. [Google Scholar] [CrossRef] [PubMed]
- Yong, V.W.; Wells, J.; Giuliani, F.; Casha, S.; Power, C.; Metz, L.M. The promise of minocycline in neurology. Lancet Neurol. 2004, 3, 744–751. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L. The Essential Medicinal Chemistry of Curcumin: Miniperspective. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Hassell, L.A.; Roanh, L.D. Potential response to curcumin in infantile hemangioendothelioma of the liver. Pediatr. Blood Cancer 2010, 55, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.E.; E Brown, R.; Buryanek, J.; Pfister, S.M.; Vats, T.S.; Rytting, M.E. Preliminary experience with personalized and targeted therapy for pediatric brain tumors. Pediatr. Blood Cancer 2011, 59, 27–33. [Google Scholar] [CrossRef]
- Ferrucci, V.; Boffa, I.; De Masi, G.; Zollo, M. Natural compounds for pediatric cancer treatment. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 389, 131–149. [Google Scholar] [CrossRef]
- Dani, C.; Pratesi, S. Task Force for Hyperbilirubinemia of the Italian Society of Neonatology. Italian guidelines for the man-agement and treatment of neonatal cholestasis. Ital. J. Pediatr. 2015, 41, 69. [Google Scholar] [CrossRef]
- Amos, R.C.; Jacob, H.; Leith, W. Jaundice in newborn babies under 28 days: NICE guideline 2016 (CG98). Arch. Dis. Child. Educ. Pr. Ed. 2017, 102, 207–209. [Google Scholar] [CrossRef]
- Fan, C.; Song, Q.; Wang, P.; Li, Y.; Yang, M.; Yu, S.Y. Neuroprotective Effects of Curcumin on IL-1β-Induced Neuronal Apoptosis and Depression-Like Behaviors Caused by Chronic Stress in Rats. Front. Cell. Neurosci. 2019, 12, 516. [Google Scholar] [CrossRef]
- Ikram, M.; Saeed, K. Natural Dietary Supplementation of Curcumin Protects Mice Brains against Ethanol-Induced Oxidative Stress-Mediated Neurodegeneration and Memory Impairment via Nrf2/TLR4/RAGE Signaling. Nutrients 2019, 11, 1082. [Google Scholar] [CrossRef] [PubMed]
- Sarker, M.R.; Franks, S.F. Efficacy of curcumin for age-associated cognitive decline: A narrative review of preclinical and clinical studies. GeroScience 2018, 40, 73–95. [Google Scholar] [CrossRef] [PubMed]
- Roca, L.; Calligaris, S.; Wennberg, R.P.; E Ahlfors, C.; Malik, S.G.; Ostrow, J.D.; Tiribelli, C. Factors Affecting the Binding of Bilirubin to Serum Albumins: Validation and Application of the Peroxidase Method. Pediatr. Res. 2006, 60, 724–728. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Waddell, J.; He, M.; Tang, N.; Rizzuto, C.; Bearer, C.F. A Gunn rat model of preterm hyperbilirubinemia. Pediatr. Res. 2020, 87, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Ruhela, R.K.; Soni, S.; Sarma, P.; Prakash, A.; Medhi, B. Negative geotaxis: An early age behavioral hallmark to VPA rat model of autism. Ann. Neurosci. 2019, 26, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Feather-Schussler, D.N.; Ferguson, T.S. A Battery of Motor Tests in a Neonatal Mouse Model of Cerebral Palsy. J. Vis. Exp. 2016, 117, e53569. [Google Scholar] [CrossRef]
- O’Neill, A.; Abranches, E.; Robertson, M.J.; Schaffer, D.V.; Cabral, J. Development of quantitative PCR methods to analyse neural progenitor cell culture state. Biotechnol. Appl. Biochem. 2006, 44, 1–8. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward Primer | Reverse Primer | Amplicon Length (bp) |
|---|---|---|---|---|
| Hprt | NM_012583.2 | AGACTGAAGAGCTACTGTAATGAC | GGCTGTACTGCTTGACCAAG | 163 |
| Gapdh | NM_017008.2 | CTCTCTGCTCCTCCCTGTTC | CACCGACCTTCACCATCTTG | 87 |
| Hmox1 | NM_012580.2 | GGTGATGGCCTCCTTGTA | ATAGACTGGGTTCTGCTTGT | 76 |
| Srxn1 | NM_001047858.3 | AAGGCGGTGACTACTACT | TTGGCAGGAATGGTCTCT | 85 |
| Tnfα | NM_012675.2 | CAACTACGATGCTCAGAAACAC | AGACAGCCTGATCCACTCC | 172 |
| IL1β | NM_031512.2 | AACAAGATAGAAGTCAAGA | ATGGTGAAGTCAACTATG | 137 |
| IL6 | NM_012589.1 | GCCCACCAGGAACGAAAGTC | ATCCTCTGTGAAGTCTCCTCTCC | 161 |
| Cox2 | NM_017232.3 | CTTTCAATGTGCAAGACC | TACTGTAGGGTTAATGTCATC | 92 |
| Icam1 | NM_012967 | ACCTACATACATTCCTACC | ATGAGACTCCATTGTTGA | 91 |
| Mag | NM_017190 | ACCATCCAACCTTCTGTATC | CTGATTCCGCTCCAAGTG | 90 |
| Mbp | NM_001025291 | CACAGAAGAGACCCTCACAGCGACA | TCCATCGGGCGCTTCTTTAGCGG | 150 |
| Target | Technique | Ab I Code and Dilution | Ab II Code and Dilution |
|---|---|---|---|
| Il1β | ELISA | ER2IL1B (Thermo Scientific, MA, USA) | |
| Tnfα | ELISA | Ab100785 (Abcam, ProdottiGianni, Milano, Italy) | |
| Actin | Western blot | A2066 (1:3000) (Sigma-Aldrich, Darmstadt, Germany) | Anti-Rabbit HRP (P0448; 1:3000) (Dako, CA, USA) |
| Hmox1 | Western blot | Sc136961 (1:100) (SantaCruz, Aachen, Germany) | Anti-Mouse HRP (P0260; 1:3500) (Dako, CA, USA) |
| Icam1 | Western blot | Sc1511 (1:50) (SantaCruz, Aachen, Germany) | Anti-Goat HRP (P0449; 1:4000) (Dako, CA, USA) |
| Mag | Western blot | Sc 166849 (1:1000) (MyBioSource, San Diego, CA, USA) | Anti-Rabbit HRP (1:3000) (Dako, CA, USA) |
| Mbp | Western blot | MBS175140 (1:100) (MyBioSource, San Diego, CA, USA) | Anti-Rabbit HRP (1:3000) (Dako, CA, USA) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gazzin, S.; Dal Ben, M.; Montrone, M.; Jayanti, S.; Lorenzon, A.; Bramante, A.; Bottin, C.; Moretti, R.; Tiribelli, C. Curcumin Prevents Cerebellar Hypoplasia and Restores the Behavior in Hyperbilirubinemic Gunn Rat by a Pleiotropic Effect on the Molecular Effectors of Brain Damage. Int. J. Mol. Sci. 2021, 22, 299. https://doi.org/10.3390/ijms22010299
Gazzin S, Dal Ben M, Montrone M, Jayanti S, Lorenzon A, Bramante A, Bottin C, Moretti R, Tiribelli C. Curcumin Prevents Cerebellar Hypoplasia and Restores the Behavior in Hyperbilirubinemic Gunn Rat by a Pleiotropic Effect on the Molecular Effectors of Brain Damage. International Journal of Molecular Sciences. 2021; 22(1):299. https://doi.org/10.3390/ijms22010299
Chicago/Turabian StyleGazzin, Silvia, Matteo Dal Ben, Michele Montrone, Sri Jayanti, Andrea Lorenzon, Alessandra Bramante, Cristina Bottin, Rita Moretti, and Claudio Tiribelli. 2021. "Curcumin Prevents Cerebellar Hypoplasia and Restores the Behavior in Hyperbilirubinemic Gunn Rat by a Pleiotropic Effect on the Molecular Effectors of Brain Damage" International Journal of Molecular Sciences 22, no. 1: 299. https://doi.org/10.3390/ijms22010299
APA StyleGazzin, S., Dal Ben, M., Montrone, M., Jayanti, S., Lorenzon, A., Bramante, A., Bottin, C., Moretti, R., & Tiribelli, C. (2021). Curcumin Prevents Cerebellar Hypoplasia and Restores the Behavior in Hyperbilirubinemic Gunn Rat by a Pleiotropic Effect on the Molecular Effectors of Brain Damage. International Journal of Molecular Sciences, 22(1), 299. https://doi.org/10.3390/ijms22010299