Possible Adverse Effects of Food Additive E171 (Titanium Dioxide) Related to Particle Specific Human Toxicity, Including the Immune System

 ,
,  , , ,
, , , 
Abstract
1. Background of TiO2 as a Food Additive
- Does oral exposure to E171 or nano-sized TiO2 reveal a relevant toxicological hazard?
- How reliable are these in-vitro and in-vivo studies?
- Are the animal models, the exposure conditions, and the effects observed in these studies relevant to humans?
- Can the data from in-vitro and in-vivo studies with TiO2 be extrapolated to humans?
- Are there epidemiological studies on the effects of E171 in humans after oral exposure?
2. Physicochemical Properties and Characterization of E171
3. Exposure to E171
4. Toxicokinetics of Ingested E171
5. Health Effect of Ingested E171
5.1. In Vivo Toxicity of E171
5.2. In Vitro and Ex Vivo Toxicity of E171
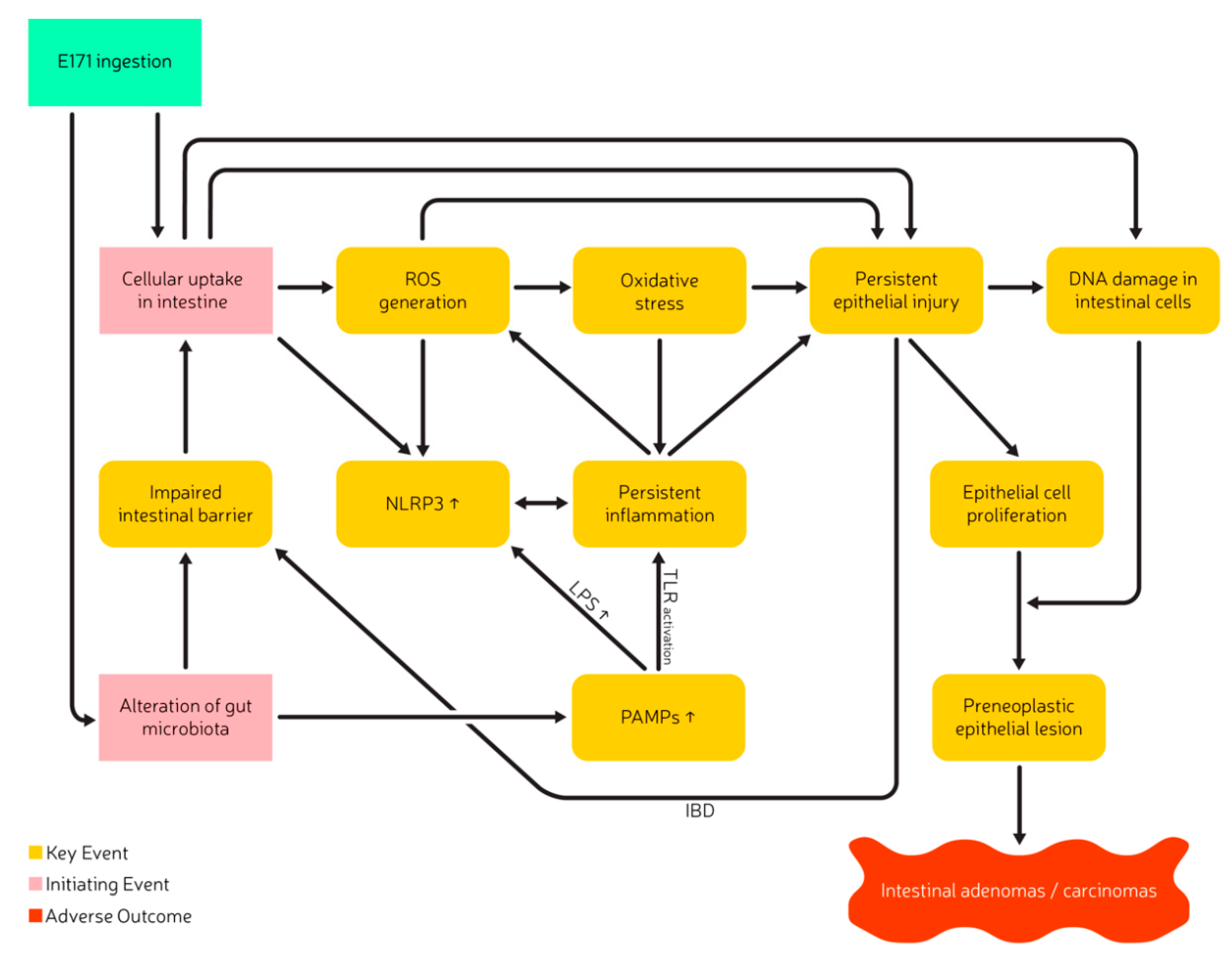
6. Mode of Action
7. Recommendation and Outlook
7.1. Particle Characterization
7.2. In Vitro Models
7.3. Rodent Studies
7.4. Non-Rodent Studies and Human Intervention Study
8. Summarizing Conclusions of the Workshop
- Does oral exposure to E171 reveal a relevant toxicological hazard in in-vitro and in-vivo studies and how reliable are these studies?
- 2.
- Are the animal models, the exposure conditions, and the effects observed in these studies relevant to humans?
- 3.
- Can the data from in vitro and in vivo studies with TiO2 be extrapolated to humans?
- 4.
- Are there epidemiological studies on the effects of E171 in humans after oral exposure?
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TiO2 | Titanium dioxide |
| BuRO | Dutch Office for Risk Assessment and Research |
| AOP | Adverse Outcome Pathway |
| EU | European Union |
| IARC | International Agency for Research and Cancer |
| RAC | Risk Assessment Committee |
| ECHA | European Chemical Agency |
| CLP | Classification, Labelling and Packaging |
| ROS | Reactive Oxygen Species |
| EFSA | European Food Safety Authority |
| ANSES | French Agency for Food, Environmental and Occupational Health |
| NVWA | Netherlands Food and Consumer Products Safety Authority |
| TEM | Transmission Electron Microscopy |
| Sp-ICP-MS | Single Particle Inductively Couple Plasma Mass Spectrometry |
| LPS | Lipopolysaccharides |
| NF-kB | Nuclear Factor Kappa Light-Chain-Enhancer of activated B-cell |
| IRF-3 | Interferon Regulatory-Factor 3 |
| Bw | Body weight |
| TJ | Tight junction |
| PBPK | Physiologically Based Pharmacokinetics |
| TEM-EDX | Transmission Electron Microscopy -Energy Dispersive X-Ray Analysis |
| IFN-γ | Interferon-gamma |
| TNF-⍺ | Tumor Necrosis Factor alpha |
| ALP | Alkaline Phosphatase |
| ALT | Alanine Aminotransferase |
| ASP | Aspartate Aminotransferase |
| GSH | Glutathione |
| GSSG | Glutathione Disulfide |
| GPx | Glutathione Peroxidase |
| SOD | Superoxid Dismutase |
| CAT | Catalase |
| IL | Interleukin |
| CYP450 | Cytochrome P450 |
| COX-2 | Cyclooxygenase-2 |
| Ki67 | Antigen Ki67 |
| Th-1 | T-helper-1 |
| P65 | Transcription factor p65 |
| MAPK | Mitogen-Activated Protein Kinase |
| GPCR | G-Protein Coupled Receptor |
| ER | Endoplasmic Reticulum |
| I.V. | Intravenous |
| I.P. | Intraperitoneal |
| IBD | Inflammatory Bowel Disease |
| CRC | Colorectal Cancer |
| BBB | Brain-Blood-Barrier |
| NO | Nitric Oxide |
| GT | Glutamyltransferase |
| StAR | Steroidogenic Regulatory Protein |
| BCL-2 | B-cell Lymphoma-2 |
| MAD | Malondialdehyde |
| PSA | Prostate-Specific Antigen |
| OECD | Organization for Economic Cooperation and Development |
| GR | Glutathione Reductase |
| LC-3 | 1A/1B-Light-Chain-3 |
| ERK | Extracellular Signaling Regulated Kinase |
| BAX | Bcl-2-Associated X protein |
| Apaf-1 | Apoptotic Peptidase Activating Factor 1 |
| Nrf2 | Nuclear Factor Erythroid 2-related Factor 2 |
| IE | Initiating Event |
| KE | Key Event |
| PAMP | Pathogen-Associated-Molecular-Pattern |
| ATP | Adenosine Triphosphate |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| EGFR | Epidermal Growth Factor Receptor |
| Cdk2 | Cyclin Dependent Kinase 2 |
| TfEB | Transcriptional Factor EB |
| GLP | Good Laboratory Practice |
| T/SEM | Transmission/Scanning Electron Microscopy |
| DLS | Dynamic Light Scattering |
| NTA | Nanoparticle Tracking Analysis |
| FCS | Fluorescence Correlation Spectroscopy |
References
- Heringa, M.B.; Geraets, L.; van Eijkeren, J.C.; Vandebriel, R.J.; de Jong, W.H.; Oomen, A.G. Risk assessment of titanium dioxide nanoparticles via oral exposure, including toxicokinetic considerations. Nanotoxicology 2016, 10, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Winkler, H.C.; Notter, T.; Meyer, U.; Naegeli, H. Critical review of the safety assessment of titanium dioxide additives in food. J. Nanobiotechnol. 2018, 16, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Braakhuis, H.M.; Gosens, I.; Heringa, M.B.; Oomen, A.G.; Vandebriel, R.J.; Groenewold, M.; Cassee, F.R. Mechanism of Action of TiO2: Recommendations to reduce uncertainties related to carcinogenic potential. Annu. Rev. Pharmacol. Toxicol. 2020, 61. [Google Scholar] [CrossRef]
- Warheit, D.B. How to measure hazards/risks following exposures to nanoscale or pigment-grade titanium dioxide particles. Toxicol. Lett. 2013, 220, 193–204. [Google Scholar] [CrossRef]
- Shi, H.; Magaye, R.; Castranova, V.; Zhao, J. Titanium dioxide nanoparticles: A review of current toxicological data. Part. Fibre Toxicol. 2013, 10, 15. [Google Scholar] [CrossRef]
- Warheit, D.B.; Donner, E.M. Risk assessment strategies for nanoscale and fine-sized titanium dioxide particles: Recognizing hazard and exposure issues. Food. Chem. Toxicol. 2015, 85, 138–147. [Google Scholar] [CrossRef]
- EFSA. EFSA Statement on the Review of the Risk Related to the Exposure to the Food Additive Titanium Dioxide (E171) Performed by the French Agency for Food, Environment and Occupational Health and Safety (ANSES); EFSA: Parma, Italy, 2019. [Google Scholar]
- EFSA; Additives PoF; Food NSat; Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; et al. EFSA statement on the review of the risks related to the exposure to the food additive titanium dioxide (E 171) performed by the French Agency for Food, Environmental and Occupational Health and Safety (ANSES). EFSA J. 2019, 5714. [Google Scholar] [CrossRef]
- Lee, K.P.; Trochimowicz, H.J.; Reinhardt, C.F. Pulmonary response of rats exposed to titanium dioxide (TiO2) by inhalation for two years. Toxicol. Appl. Pharmacol. 1985, 79, 179–192. [Google Scholar] [CrossRef]
- IARC. IARC monographs on the evaluation of carcinogenic risks to humans. In Carbon Black, Titanium Dioxide and Talc; IARC: Lyon, France, 2010; Volume 93. [Google Scholar]
- ECHA. Opinion Proposing Harmonised Classification and Labelling a EU Level of Titanium Dioxide; ECHA: Helsinki, Finland, 2017. [Google Scholar]
- European Parliament. Regulation (EC) No 1272/2008 of the European Parliament and of the Council of 16 December 2008 on Classification, Labelling and Packaging of Substances and Mixtures, Amending and Repealing Directives 67/548/EEC and 1999/45/EC, and Amending Regulation (EC) No 1907/2006; European Parliament: Brussels, Belgium, 2020. [Google Scholar]
- Brand, W.; Peters, R.J.B.; Braakhuis, H.M.; Maslankiewicz, L.; Oomen, A.G. Possible effects of titanium dioxide particles on human liver, intestinal tissue, spleen and kidney after oral exposure. Nanotoxicology 2020, 14, 985–1007. [Google Scholar] [CrossRef]
- Bettini, S.; Boutet-Robinet, E.; Cartier, C.; Comera, C.; Gaultier, E.; Dupuy, J.; Naud, N.; Tache, S.; Grysan, P.; Reguer, S.; et al. Food-grade TiO2 impairs intestinal and systemic immune homeostasis, initiates preneoplastic lesions and promotes aberrant crypt development in the rat colon. Sci. Rep. 2017, 7, 40373. [Google Scholar] [CrossRef]
- Urrutia-Ortega, I.M.; Garduno-Balderas, L.G.; Delgado-Buenrostro, N.L.; Freyre-Fonseca, V.; Flores-Flores, J.O.; Gonzalez-Robles, A.; Pedraza-Chaverri, J.; Hernandez-Pando, R.; Rodriguez-Sosa, M.; Leon-Cabrera, S.; et al. Food-grade titanium dioxide exposure exacerbates tumor formation in colitis associated cancer model. Food. Chem. Toxicol. 2016, 93, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Proquin, H.; Rodriguez-Ibarra, C.; Moonen, C.G.; Urrutia Ortega, I.M.; Briede, J.J.; de Kok, T.M.; van Loveren, H.; Chirino, Y.I. Titanium dioxide food additive (E171) induces ROS formation and genotoxicity: Contribution of micro and nano-sized fractions. Mutagenesis 2017, 32, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Blevins, L.K.; Crawford, R.B.; Bach, A.; Rizzo, M.D.; Zhou, J.; Henriquez, J.E.; Khan, D.; Sermet, S.; Arnold, L.L.; Pennington, K.L.; et al. Evaluation of immunologic and intestinal effects in rats administered an E 171-containing diet, a food grade titanium dioxide (TiO2). Food Chem. Toxicol. 2019, 133, 110793. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.D.C., Jr.; Azevedo, L.F.; de Souza Rocha, C.C.; Carneiro, M.F.H.; Venancio, V.P.; de Almeida, M.R.; Antunes, L.M.G.; de Carvalho Hott, R.; Rodrigues, J.L.; Ogunjimi, A.T.; et al. Evaluation of distribution, redox parameters, and genotoxicity in Wistar rats co-exposed to silver and titanium dioxide nanoparticles. J. Toxicol. Environ. Health A 2017, 80, 1156–1165. [Google Scholar] [CrossRef] [PubMed]
- ANSES. OPINION of the French Agency for Food, Environmental and Occupational Health and Safety on the Risk Associated with Ingestion of the Food Additive E171; ANSES: Maisons-Alfort, France, 2019. [Google Scholar]
- Bundesinstitut für Berufsbildung. Titanium Dioxide: Further Research Is Still Needed; Bundesinstitut für Berufsbildung: Bonn, Germany, 2020. [Google Scholar]
- EFSA. Re-Evaluation of titanium dioxide (E 171) as a food additive. EFSA J. 2016, 14. [Google Scholar] [CrossRef]
- EFSA. Scientific Panel on food additives and flavourings. In Proceedings of the Minutes of the 16th Pleenary Meeting, Parma, Italy, 21–23 September 2020. [Google Scholar]
- BuRO. Opinion on Possible Health Effects of the Food Additive Titanium Dioxide (E171); Netherlands Food and Consumer Product Safety Authority: The Hague, The Netherlands, 2019. [Google Scholar]
- Sayes, C.M.; Wahi, R.; Kurian, P.A.; Liu, Y.; West, J.L.; Ausman, K.D.; Warheit, D.B.; Colvin, V.L. Correlating nanoscale titania structure with toxicity: A cytotoxicity and inflammatory response study with human dermal fibroblasts and human lung epithelial cells. Toxicol. Sci. 2006, 92, 174–185. [Google Scholar] [CrossRef]
- Peters, R.J.; van Bemmel, G.; Herrera-Rivera, Z.; Helsper, H.P.; Marvin, H.J.; Weigel, S.; Tromp, P.C.; Oomen, A.G.; Rietveld, A.G.; Bouwmeester, H. Characterization of titanium dioxide nanoparticles in food products: Analytical methods to define nanoparticles. J. Agric. Food Chem 2014, 62, 6285–6293. [Google Scholar] [CrossRef]
- Dudefoi, W.; Terrisse, H.; Popa, A.F.; Gautron, E.; Humbert, B.; Ropers, M.H. Evaluation of the content of TiO2 nanoparticles in the coatings of chewing gums. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018, 35, 211–221. [Google Scholar] [CrossRef]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; von Goetz, N. Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef]
- Geiss, O.; Ponti, J.; Senaldi, C.; Bianchi, I.; Mehn, D.; Barrero, J.; Gilliland, D.; Matissek, R.; Anklam, E. Characterisation of food grade titania with respect to nanoparticle content in pristine additives and in their related food products. Food Addit. Contam. Part. A Chem. Anal. Control Expo. Risk Assess. 2020, 37, 239–253. [Google Scholar] [CrossRef]
- Commission, T.E. Definition of nanomaterial. Off. J. Eur. Union 2011, 38–40. [Google Scholar] [CrossRef]
- Verleysen, E.; Waegeneers, N.; Brassinne, F.; De Vos, S.; Jimenez, I.O.; Mathioudaki, S.; Mast, J. Physicochemical Characterization of the Pristine E171 Food Additive by Standardized and Validated Methods. Nanomaterials 2020, 10, 592. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Bae, D.; Fong, A. Titanium Dioxide in Food Products: Quantitative Analysis Using ICP-MS and Raman Spectroscopy. J. Agric. Food Chem. 2018, 66, 13533–13540. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; George, S.; Ji, Z.; Lin, S.; Yu, H.; Damoiseaux, R.; France, B.; Ng, K.W.; Loo, S.C.J. Size of TiO(2) nanoparticles influences their phototoxicity: An in vitro investigation. Arch. Toxicol. 2013, 87, 99–109. [Google Scholar] [CrossRef]
- Musial, J.; Krakowiak, R.; Mlynarczyk, D.T.; Goslinski, T.; Stanisz, B.J. Titanium dioxide nanoparticles in food and personal care products-what do we know about their safety? Nanomaterials 2020, 10, 1110. [Google Scholar] [CrossRef]
- Warheit, D.B.; Brown, S.C. What is the impact of surface modifications and particle size on commercial titanium dioxide particle samples? A review of in vivo pulmonary and oral toxicity studies—Revised 11-6-2018. Toxicol. Lett. 2019, 302, 42–59. [Google Scholar] [CrossRef]
- Ali, S.A.; Rizk, M.Z.; Hamed, M.A.; Aboul-Ela, E.I.; El-Rigal, N.S.; Aly, H.F.; Abdel-Hamid, A.Z. Assessment of titanium dioxide nanoparticles toxicity via oral exposure in mice: Effect of dose and particle size. Biomarkers 2019, 24, 492–498. [Google Scholar] [CrossRef]
- Posgai, R.; Cipolla-McCulloch, C.B.; Murphy, K.R.; Hussain, S.M.; Rowe, J.J.; Nielsen, M.G. Differential toxicity of silver and titanium dioxide nanoparticles on Drosophila melanogaster development, reproductive effort, and viability: Size, coatings and antioxidants matter. Chemosphere 2011, 85, 34–42. [Google Scholar] [CrossRef]
- Moore, T.L.; Rodriguez-Lorenzo, L.; Hirsch, V.; Balog, S.; Urban, D.; Jud, C.; Rothen-Rutishauser, B.; Lattuada, M.; Petri-Fink, A. Nanoparticle colloidal stability in cell culture media and impact on cellular interactions. Chem. Soc. Rev. 2015, 44, 6287–6305. [Google Scholar] [CrossRef]
- Janer, G.; Mas del Molino, E.; Fernandez-Rosas, E.; Fernandez, A.; Vazquez-Campos, S. Cell uptake and oral absorption of titanium dioxide nanoparticles. Toxicol. Lett. 2014, 228, 103–110. [Google Scholar] [CrossRef]
- Murugadoss, S.; Brassinne, F.; Sebaihi, N.; Petry, J.; Cokic, S.M.; Van Landuyt, K.L.; Godderis, L.; Mast, J.; Lison, D.; Hoet, P.H.; et al. Agglomeration of titanium dioxide nanoparticles increases toxicological responses in vitro and in vivo. Part. Fibre Toxicol. 2020, 17, 10. [Google Scholar] [CrossRef] [PubMed]
- Foroozandeh, P.; Aziz, A.A. Merging worlds of nanomaterials and biological environment: Factors governing protein corona formation on nanoparticles and its biological consequences. Nanoscale Res. Lett. 2015, 10, 221. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.O.; Di Maio, A.; Guggenheim, E.J.; Chetwynd, A.J.; Pencross, D.; Tang, S.; Belinga-Desaunay, M.A.; Thomas, S.G.; Rappoport, J.Z.; Lynch, I. Surface chemistry-dependent evolution of the nanomaterial corona on TiO2 nanomaterials following uptake and sub-cellular localization. Nanomaterials 2020, 10, 401. [Google Scholar] [CrossRef] [PubMed]
- Mazzolini, J.; Weber, R.J.; Chen, H.S.; Khan, A.; Guggenheim, E.; Shaw, R.K.; Chipman, J.K.; Viant, M.R.; Rappoport, J.Z. Protein corona modulates uptake and toxicity of nanoceria via clathrin-mediated endocytosis. Biol. Bull. 2016, 231, 40–60. [Google Scholar] [CrossRef] [PubMed]
- Saptarshi, S.R.; Duschl, A.; Lopata, A.L. Interaction of nanoparticles with proteins: Relation to bio-reactivity of the nanoparticle. J. Nanobiotechnol. 2013, 11, 26. [Google Scholar] [CrossRef]
- Bianchi, M.G.; Allegri, M.; Chiu, M.; Costa, A.L.; Blosi, M.; Ortelli, S.; Bussolati, O.; Bergamaschi, E. Lipopolysaccharide adsorbed to the bio-corona of TIO2 nanoparticles powerfully activates selected pro-inflammatory transduction pathways. Front. Immunol. 2017, 8, 866. [Google Scholar] [CrossRef]
- Chen, X.X.; Cheng, B.; Yang, Y.X.; Cao, A.; Liu, J.H.; Du, L.J.; Liu, Y.; Zhao, Y.; Wang, H. Characterization and preliminary toxicity assay of nano-titanium dioxide additive in sugar-coated chewing gum. Small 2013, 9, 1765–1774. [Google Scholar] [CrossRef]
- Dudefoi, W.; Moniz, K.; Allen-Vercoe, E.; Ropers, M.H.; Walker, V.K. Impact of food grade and nano-TiO2 particles on a human intestinal community. Food Chem. Toxicol. 2017, 106, 242–249. [Google Scholar] [CrossRef]
- Dudefoi, W.; Terrisse, H.; Richard-Plouet, M.; Gautron, E.; Popa, F.; Humbert, B.; Ropers, M.H. Criteria to define a more relevant reference sample of titanium dioxide in the context of food: A multiscale approach. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2017, 34, 653–665. [Google Scholar] [CrossRef]
- Yang, Y.; Doudrick, K.; Bi, X.; Hristovski, K.; Herckes, P.; Westerhoff, P.; Kaegi, R. Characterization of food-grade titanium dioxide: The presence of nanosized particles. Environ. Sci. Technol. 2014, 48, 6391–6400. [Google Scholar] [CrossRef]
- Rompelberg, C.; Heringa, M.B.; van Donkersgoed, G.; Drijvers, J.; Roos, A.; Westenbrink, S.; Peters, R.; van Bemmel, G.; Brand, W.; Oomen, A.G. Oral intake of added titanium dioxide and its nanofraction from food products, food supplements and toothpaste by the Dutch population. Nanotoxicology 2016, 10, 1404–1414. [Google Scholar] [CrossRef]
- Wu, F.; Hicks, A.L. Estimating human exposure to titanium dioxide from personal care products through a social survey approach. Integr. Environ. Assess. Manag. 2020, 16, 10–16. [Google Scholar] [CrossRef]
- Bachler, G.; von Goetz, N.; Hungerbuhler, K. Using physiologically based pharmacokinetic (PBPK) modeling for dietary risk assessment of titanium dioxide (TiO2) nanoparticles. Nanotoxicology 2015, 9, 373–380. [Google Scholar] [CrossRef]
- Sprong, C.; Bakker, M.; Niekerk, M.; Vennemann, M. Exposure Assessment of the Food Additive Titanium Dioxide (E 171) Based on Use Levels Provided by the Industry; RIVM Letter Report 2015-0195; National Institute for Public Health and the Environment: Bilthoven, The Netherlands, 2016.
- Christensen, F.M.; Christensen, T.B.; Vogel, U.; Jensen, K.A.; Lassen, C. Better Control of Nanomaterials—Summary of the 4-Year Danish Initiative on Nanomaterials; The Danish Environmental Protection Agency: København, Denmark, 2015. [Google Scholar]
- Powell, J.J.; Faria, N.; Thomas-McKay, E.; Pele, L.C. Origin and fate of dietary nanoparticles and microparticles in the gastrointestinal tract. J. Autoimmun. 2010, 34, J226–J233. [Google Scholar] [CrossRef]
- Golja, V.; Drazic, G.; Lorenzetti, M.; Vidmar, J.; Scancar, J.; Zalaznik, M.; Kalin, M.; Novak, S. Characterisation of food contact non-stick coatings containing TiO2 nanoparticles and study of their possible release into food. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2017, 34, 421–433. [Google Scholar] [CrossRef]
- Teubl, B.J.; Leitinger, G.; Schneider, M.; Lehr, C.M.; Frohlich, E.; Zimmer, A.; Roblegg, E. The buccal mucosa as a route for TiO2 nanoparticle uptake. Nanotoxicology 2015, 9, 253–261. [Google Scholar] [CrossRef]
- Teubl, B.J.; Schimpel, C.; Leitinger, G.; Bauer, B.; Frohlich, E.; Zimmer, A.; Roblegg, E. Interactions between nano-TiO2 and the oral cavity: Impact of nanomaterial surface hydrophilicity/hydrophobicity. J. Hazard. Mater. 2015, 286, 298–305. [Google Scholar] [CrossRef]
- Pedata, P.; Ricci, G.; Malorni, L.; Venezia, A.; Cammarota, M.; Volpe, M.G.; Iannaccone, N.; Guida, V.; Schiraldi, C.; Romano, M.; et al. In vitro intestinal epithelium responses to titanium dioxide nanoparticles. Food Res. Int. 2019, 119, 634–642. [Google Scholar] [CrossRef]
- Deciga-Alcaraz, A.; Delgado-Buenrostro, N.L.; Ispanixtlahuatl-Meraz, O.; Freyre-Fonseca, V.; Flores-Flores, J.O.; Ganem-Rondero, A.; Vaca-Paniagua, F.; Pilar Ramos-Godinez, M.D.; Morales-Barcenas, R.; Sanchez-Perez, Y.; et al. Irreversible disruption of the cytoskeleton as induced by non-cytotoxic exposure to titanium dioxide nanoparticles in lung epithelial cells. Chem. Biol. Interact. 2020, 323, 109063. [Google Scholar] [CrossRef]
- Brun, E.; Barreau, F.; Veronesi, G.; Fayard, B.; Sorieul, S.; Chaneac, C.; Carapito, C.; Rabilloud, T.; Mabondzo, A.; Herlin-Boime, N.; et al. Titanium dioxide nanoparticle impact and translocation through ex vivo, in vivo and in vitro gut epithelia. Part. Fibre Toxicol. 2014, 11, 13. [Google Scholar] [CrossRef]
- Comera, C.; Cartier, C.; Gaultier, E.; Catrice, O.; Panouille, Q.; El Hamdi, S.; Tirez, K.; Nelissen, I.; Theodorou, V.; Houdeau, E. Jejunal villus absorption and paracellular tight junction permeability are major routes for early intestinal uptake of food-grade TiO2 particles: An in vivo and ex vivo study in mice. Part. Fibre Toxicol. 2020, 17, 26. [Google Scholar] [CrossRef]
- Kreyling, W.G.; Holzwarth, U.; Haberl, N.; Kozempel, J.; Hirn, S.; Wenk, A.; Schleh, C.; Schaffler, M.; Lipka, J.; Semmler-Behnke, M.; et al. Quantitative biokinetics of titanium dioxide nanoparticles after intravenous injection in rats: Part 1. Nanotoxicology 2017, 11, 434–442. [Google Scholar] [CrossRef]
- Demir, E. An in vivo study of nanorod, nanosphere, and nanowire forms of titanium dioxide using Drosophila melanogaster: Toxicity, cellular uptake, oxidative stress, and DNA damage. J Toxicol. Environ. Health A 2020, 83, 456–469. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, G.; Chen, C.; Yu, H.; Wang, T.; Ma, Y.; Jia, G.; Gao, Y.; Li, B.; Sun, J.; et al. Acute toxicity and biodistribution of different sized titanium dioxide particles in mice after oral administration. Toxicol. Lett. 2007, 168, 176–185. [Google Scholar] [CrossRef]
- Shinohara, N.; Danno, N.; Ichinose, T.; Sasaki, T.; Fukui, H.; Honda, K.; Gamo, M. Tissue distribution and clearance of intravenously administered titanium dioxide (TiO2) nanoparticles. Nanotoxicology 2014, 8, 132–141. [Google Scholar] [CrossRef]
- Tassinari, R.; Cubadda, F.; Moracci, G.; Aureli, F.; D’Amato, M.; Valeri, M.; De Berardis, B.; Raggi, A.; Mantovani, A.; Passeri, D.; et al. Oral, short-term exposure to titanium dioxide nanoparticles in Sprague-Dawley rat: Focus on reproductive and endocrine systems and spleen. Nanotoxicology 2014, 8, 654–662. [Google Scholar] [CrossRef]
- Donner, E.M.; Myhre, A.; Brown, S.C.; Boatman, R.; Warheit, D.B. In vivo micronucleus studies with 6 titanium dioxide materials (3 pigment-grade & 3 nanoscale) in orally-exposed rats. Regul. Toxicol. Pharmacol. 2016, 74, 64–74. [Google Scholar] [CrossRef]
- Bockmann, J.; Lahl, H.; Eckert, T.; Unterhalt, B. Blood titanium levels before and after oral administration titanium dioxide. Pharmazie 2000, 55, 140–143. [Google Scholar]
- Jones, K.; Morton, J.; Smith, I.; Jurkschat, K.; Harding, A.H.; Evans, G. Human in vivo and in vitro studies on gastrointestinal absorption of titanium dioxide nanoparticles. Toxicol. Lett. 2015, 233, 95–101. [Google Scholar] [CrossRef]
- Pele, L.C.; Thoree, V.; Bruggraber, S.F.; Koller, D.; Thompson, R.P.; Lomer, M.C.; Powell, J.J. Pharmaceutical/food grade titanium dioxide particles are absorbed into the bloodstream of human volunteers. Part. Fibre Toxicol. 2015, 12, 26. [Google Scholar] [CrossRef]
- Geraets, L.; Oomen, A.G.; Krystek, P.; Jacobsen, N.R.; Wallin, H.; Laurentie, M.; Verharen, H.W.; Brandon, E.F.; de Jong, W.H. Tissue distribution and elimination after oral and intravenous administration of different titanium dioxide nanoparticles in rats. Part. Fibre Toxicol. 2014, 11, 30. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, Y.; Dong, L.; Huang, J.; Zhang, Y.; Su, J.; Zang, J. A hybrid of titanium nitride and nitrogen-doped amorphous carbon supported on SiC as a noble metal-free electrocatalyst for oxygen reduction reaction. Chem. Commun. 2015, 51, 2625–2628. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Z.; Ba, T.; Pu, J.; Chen, T.; Song, Y.; Gu, Y.; Qian, Q.; Xu, Y.; Xiang, K.; et al. Susceptibility of young and adult rats to the oral toxicity of titanium dioxide nanoparticles. Small 2013, 9, 1742–1752. [Google Scholar] [CrossRef]
- Hummel, T.Z.; Kindermann, A.; Stokkers, P.C.; Benninga, M.A.; ten Kate, F.J. Exogenous pigment in Peyer patches of children suspected of having IBD. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 477–480. [Google Scholar] [CrossRef]
- Powell, J.J.; Ainley, C.C.; Harvey, R.S.; Mason, I.M.; Kendall, M.D.; Sankey, E.A.; Dhillon, A.P.; Thompson, R.P. Characterisation of inorganic microparticles in pigment cells of human gut associated lymphoid tissue. Gut 1996, 38, 390–395. [Google Scholar] [CrossRef]
- Heringa, M.B.; Peters, R.J.B.; Bleys, R.; van der Lee, M.K.; Tromp, P.C.; van Kesteren, P.C.E.; van Eijkeren, J.C.H.; Undas, A.K.; Oomen, A.G.; Bouwmeester, H. Detection of titanium particles in human liver and spleen and possible health implications. Part. Fibre Toxicol. 2018, 15, 15. [Google Scholar] [CrossRef]
- Guillard, A.; Gaultier, E.; Cartier, C.; Devoille, L.; Noireaux, J.; Chevalier, L.; Morin, M.; Grandin, F.; Lacroix, M.Z.; Comera, C.; et al. Basal Ti level in the human placenta and meconium and evidence of a materno-foetal transfer of food-grade TiO2 nanoparticles in an ex vivo placental perfusion model. Part. Fibre Toxicol. 2020, 17, 51. [Google Scholar] [CrossRef]
- Cho, W.S.; Kang, B.C.; Lee, J.K.; Jeong, J.; Che, J.H.; Seok, S.H. Comparative absorption, distribution, and excretion of titanium dioxide and zinc oxide nanoparticles after repeated oral administration. Part. Fibre Toxicol. 2013, 10, 9. [Google Scholar] [CrossRef]
- Da Silva, A.B.; Miniter, M.; Thom, W.; Hewitt, R.E.; Wills, J.; Jugdaohsingh, R.; Powell, J.J. Gastrointestinal Absorption and Toxicity of Nanoparticles and Microparticles: Myth, Reality and Pitfalls explored through Titanium Dioxide. Curr. Opin. Toxicol. 2020, 19, 112–120. [Google Scholar] [CrossRef]
- Lamas, B.; Martins Breyner, N.; Houdeau, E. Impacts of foodborne inorganic nanoparticles on the gut microbiota-immune axis: Potential consequences for host health. Part. Fibre Toxicol. 2020, 17, 19. [Google Scholar] [CrossRef]
- NTP. Bioassay of Titanium Dioxide for Possible Carcinogenicity; National Cancer Institute: Bathesda, MD, USA, 1979.
- Bartsch, R.; Brinkmann, B.; Jahnke, G.; Laube, B.; Lohmann, R.; Michaelsen, S.; Neumann, I.; Greim, H. Human relevance of follicular thyroid tumors in rodents caused by non-genotoxic substances. Regul. Toxicol. Pharmacol. 2018, 98, 199–208. [Google Scholar] [CrossRef]
- Warheit, D.B.; Brown, S.C.; Donner, E.M. Acute and subchronic oral toxicity studies in rats with nanoscale and pigment grade titanium dioxide particles. Food Chem. Toxicol. 2015, 84, 208–224. [Google Scholar] [CrossRef]
- Warheit, D.B.; Boatman, R.; Brown, S.C. Developmental toxicity studies with 6 forms of titanium dioxide test materials (3 pigment-different grade & 3 nanoscale) demonstrate an absence of effects in orally-exposed rats. Regul. Toxicol. Pharmacol. 2015, 73, 887–896. [Google Scholar]
- Chen, Z.; Zhou, D.; Han, S.; Zhou, S.; Jia, G. Hepatotoxicity and the role of the gut-liver axis in rats after oral administration of titanium dioxide nanoparticles. Part. Fibre Toxicol. 2019, 16, 48. [Google Scholar] [CrossRef]
- Moradi, A.; Ziamajidi, N.; Ghafourikhosroshahi, A.; Abbasalipourkabir, R. Effects of vitamin A and vitamin E on attenuation of titanium dioxide nanoparticles-induced toxicity in the liver of male Wistar rats. Mol. Biol. Rep. 2019, 46, 2919–2932. [Google Scholar] [CrossRef]
- Orazizadeh, M.; Fakhredini, F.; Mansouri, E.; Khorsandi, L. Effect of glycyrrhizic acid on titanium dioxide nanoparticles-induced hepatotoxicity in rats. Chem. Biol. Interact. 2014, 220, 214–221. [Google Scholar] [CrossRef]
- Hu, H.; Li, L.; Guo, Q.; Zong, H.; Yan, Y.; Yin, Y.; Wang, Y.; Oh, Y.; Feng, Y.; Wu, Q.; et al. RNA sequencing analysis shows that titanium dioxide nanoparticles induce endoplasmic reticulum stress, which has a central role in mediating plasma glucose in mice. Nanotoxicology 2018, 12, 341–356. [Google Scholar] [CrossRef]
- Ruiz, P.A.; Moron, B.; Becker, H.M.; Lang, S.; Atrott, K.; Spalinger, M.R.; Scharl, M.; Wojtal, K.A.; Fischbeck-Terhalle, A.; Frey-Wagner, I.; et al. Titanium dioxide nanoparticles exacerbate DSS-induced colitis: Role of the NLRP3 inflammasome. Gut 2017, 66, 1216–1224. [Google Scholar] [CrossRef]
- Trouiller, B.; Reliene, R.; Westbrook, A.; Solaimani, P.; Schiestl, R.H. Titanium dioxide nanoparticles induce DNA damage and genetic instability in vivo in mice. Cancer Res. 2009, 69, 8784–8789. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Goyary, D.; Karmakar, S.; Chattopadhyay, P. Exploration of cytotoxic and genotoxic endpoints following sub-chronic oral exposure to titanium dioxide nanoparticles. Toxicol. Ind. Health 2019, 35, 577–592. [Google Scholar] [CrossRef]
- Proquin, H.; Jetten, M.J.; Jonkhout, M.C.M.; Garduno-Balderas, L.G.; Briede, J.J.; de Kok, T.M.; Chirino, Y.I.; van Loveren, H. Gene expression profiling in colon of mice exposed to food additive titanium dioxide (E171). Food Chem. Toxicol. 2018, 111, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Sycheva, L.P.; Zhurkov, V.S.; Iurchenko, V.V.; Daugel-Dauge, N.O.; Kovalenko, M.A.; Krivtsova, E.K.; Durnev, A.D. Investigation of genotoxic and cytotoxic effects of micro- and nanosized titanium dioxide in six organs of mice in vivo. Mutat. Res. 2011, 726, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Jensen, D.M.; Lohr, M.; Sheykhzade, M.; Lykkesfeldt, J.; Wils, R.S.; Loft, S.; Moller, P. Telomere length and genotoxicity in the lung of rats following intragastric exposure to food-grade titanium dioxide and vegetable carbon particles. Mutagenesis 2019, 34, 203–214. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, B.; Li, L.; Guo, Q.; Yang, D.; Wei, X.; Fan, X.; Liu, J.; Wu, Q.; Oh, Y.; et al. The toxic effects of titanium dioxide nanoparticles on plasma glucose metabolism are more severe in developing mice than in adult mice. Environ. Toxicol. 2020, 35, 443–456. [Google Scholar] [CrossRef]
- Fadoju, O.; Ogunsuyi, O.; Akanni, O.; Alabi, O.; Alimba, C.; Adaramoye, O.; Cambier, S.; Eswara, S.; Gutleb, A.C.; Bakare, A. Evaluation of cytogenotoxicity and oxidative stress parameters in male Swiss mice co-exposed to titanium dioxide and zinc oxide nanoparticles. Environ. Toxicol. Pharmacol. 2019, 70, 103204. [Google Scholar] [CrossRef]
- Louro, H.; Tavares, A.; Vital, N.; Costa, P.M.; Alverca, E.; Zwart, E.; de Jong, W.H.; Fessard, V.; Lavinha, J.; Silva, M.J. Integrated approach to the in vivo genotoxic effects of a titanium dioxide nanomaterial using LacZ plasmid-based transgenic mice. Environ. Mol. Mutagen. 2014, 55, 500–509. [Google Scholar] [CrossRef]
- Riedle, S.; Pele, L.C.; Otter, D.E.; Hewitt, R.E.; Singh, H.; Roy, N.C.; Powell, J.J. Pro-inflammatory adjuvant properties of pigment-grade titanium dioxide particles are augmented by a genotype that potentiates interleukin 1beta processing. Part. Fibre Toxicol. 2017, 14, 51. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, D.; Zhou, S.; Jia, G. Gender difference in hepatic toxicity of titanium dioxide nanoparticles after subchronic oral exposure in Sprague-Dawley rats. J. Appl. Toxicol. 2019, 39, 807–819. [Google Scholar] [CrossRef]
- Bu, Q.; Yan, G.; Deng, P.; Peng, F.; Lin, H.; Xu, Y.; Cao, Z.; Zhou, T.; Xue, A.; Wang, Y.; et al. NMR-based metabonomic study of the sub-acute toxicity of titanium dioxide nanoparticles in rats after oral administration. Nanotechnology 2010, 21, 125105. [Google Scholar] [CrossRef]
- Waller, T.; Chen, C.; Walkerm, L. Food and industrial grade titanium dioxide impacts gut microbiota. Environ. Eng. Sci. 2017, 34, 537–550. [Google Scholar] [CrossRef]
- Mao, Z.; Li, Y.; Dong, T.; Zhang, L.; Zhang, Y.; Li, S.; Hu, H.; Sun, C.; Xia, Y. Exposure to Titanium Dioxide Nanoparticles During Pregnancy Changed Maternal Gut Microbiota and Increased Blood Glucose of Rat. Nanoscale Res. Lett 2019, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.; Wang, Y.; Huang, C.; Fu, Y.; Li, J.; Wang, H.; Jia, X.; Ba, Q. Effect of Long-Term Intake of Dietary Titanium Dioxide Nanoparticles on Intestine Inflammation in Mice. J. Agric. Food Chem. 2019, 67, 9382–9389. [Google Scholar] [CrossRef] [PubMed]
- Pinget, G.; Tan, J.; Janac, B.; Kaakoush, N.O.; Angelatos, A.S.; O’Sullivan, J.; Koay, Y.C.; Sierro, F.; Davis, J.; Divakarla, S.K.; et al. Impact of the Food Additive Titanium Dioxide (E171) on Gut Microbiota-Host Interaction. Front. Nutr. 2019, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- Flemer, B.; Lynch, D.B.; Brown, J.M.; Jeffery, I.B.; Ryan, F.J.; Claesson, M.J.; O’Riordain, M.; Shanahan, F.; O’Toole, P.W. Tumour-associated and non-tumour-associated microbiota in colorectal cancer. Gut 2017, 66, 633–643. [Google Scholar] [CrossRef]
- Saus, E.; Iraola-Guzman, S.; Willis, J.R.; Brunet-Vega, A.; Gabaldon, T. Microbiome and colorectal cancer: Roles in carcinogenesis and clinical potential. Mol. Asp. Med. 2019, 69, 93–106. [Google Scholar] [CrossRef]
- Sobhani, I.; Bergsten, E.; Couffin, S.; Amiot, A.; Nebbad, B.; Barau, C.; de’Angelis, N.; Rabot, S.; Canoui-Poitrine, F.; Mestivier, D.; et al. Colorectal cancer-associated microbiota contributes to oncogenic epigenetic signatures. Proc. Natl. Acad. Sci. USA 2019, 116, 24285–24295. [Google Scholar] [CrossRef]
- Grissa, I.; ElGhoul, J.; Mrimi, R.; Mir, L.E.; Cheikh, H.B.; Horcajada, P. In deep evaluation of the neurotoxicity of orally administered TiO2 nanoparticles. Brain Res. Bull. 2020, 155, 119–128. [Google Scholar] [CrossRef]
- Grissa, I.; Guezguez, S.; Ezzi, L.; Chakroun, S.; Sallem, A.; Kerkeni, E.; Elghoul, J.; El Mir, L.; Mehdi, M.; Cheikh, H.B.; et al. The effect of titanium dioxide nanoparticles on neuroinflammation response in rat brain. Environ. Sci. Pollut. Res. Int. 2016, 23, 20205–20213. [Google Scholar] [CrossRef]
- Heidari, Z.; Mohammadipour, A.; Haeri, P.; Ebrahimzadeh-Bideskan, A. The effect of titanium dioxide nanoparticles on mice midbrain substantia nigra. Iran. J. Basic Med. Sci. 2019, 22, 745–751. [Google Scholar]
- Jensen, D.M.; Christophersen, D.V.; Sheykhzade, M.; Skovsted, G.F.; Lykkesfeldt, J.; Munter, R.; Roursgaard, M.; Loft, S.; Moller, P. Vasomotor function in rat arteries after ex vivo and intragastric exposure to food-grade titanium dioxide and vegetable carbon particles. Part. Fibre Toxicol. 2018, 15, 12. [Google Scholar] [CrossRef]
- Jensen, D.M.; Skovsted, G.F.; Lykkesfeldt, J.; Dreier, R.; Berg, J.O.; Jeppesen, J.L.; Sheykhzade, M.; Loft, S.; Moller, P. Vasomotor dysfunction in human subcutaneous arteries exposed ex vivo to food-grade titanium dioxide. Food Chem. Toxicol. 2018, 120, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Khorsandi, L.; Nejaddehbashi, F. Protective effects of Curcumin on testicular toxicity induced by titanium dioxide nanoparticles in mice. JBRA Assist. Reprod. 2019, 23, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Tainaka, H.; Oba, T.; Mizuo, K.; Umezawa, M.; Takeda, K. Maternal exposure to nanoparticulate titanium dioxide during the prenatal period alters gene expression related to brain development in the mouse. Part. Fibre Toxicol. 2009, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Shahin, N.N.; Mohamed, M.M. Nano-Sized titanium dioxide toxicity in rat prostate and testis: Possible ameliorative effect of morin. Toxicol. Appl. Pharmacol. 2017, 334, 129–141. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, X.; Zhang, X.; Zhao, Z.; Liu, H.; George, R.; Wilson-Rawls, J.; Chang, Y.; Chen, Y. Disruption of zebrafish (Danio rerio) reproduction upon chronic exposure to TiO(2) nanoparticles. Chemosphere 2011, 83, 461–467. [Google Scholar] [CrossRef]
- Ma, H.; Lenz, K.A.; Gao, X.; Li, S.; Wallis, L.K. Comparative toxicity of a food additive TiO2, a bulk TiO2, and a nano-sized P25 to a model organism the nematode C. elegans. Environ. Sci. Pollut. Res. Int. 2019, 26, 3556–3568. [Google Scholar] [CrossRef]
- Jovanovic, B.; Cvetkovic, V.J.; Mitrovic, T. Effects of human food grade titanium dioxide nanoparticle dietary exposure on Drosophila melanogaster survival, fecundity, pupation and expression of antioxidant genes. Chemosphere 2016, 144, 43–49. [Google Scholar] [CrossRef]
- Jovanovic, B.; Jovanovic, N.; Cvetkovic, V.J.; Matic, S.; Stanic, S.; Whitley, E.M.; Mitrovic, T.L. The effects of a human food additive, titanium dioxide nanoparticles E171, on Drosophila melanogaster—A 20 generation dietary exposure experiment. Sci. Rep. 2018, 8, 17922. [Google Scholar] [CrossRef]
- Kim, D.H.; Kundu, J.; Chae, I.G.; Lee, J.K.; Heo, J.S.; Chun, K.S. Titanium dioxide nanoparticles induce COX-2 expression through ROS generation in human periodontal ligament cells. J. Toxicol. Sci. 2019, 44, 335–345. [Google Scholar] [CrossRef]
- Gerloff, K.; Fenoglio, I.; Carella, E.; Kolling, J.; Albrecht, C.; Boots, A.W.; Forster, I.; Schins, R.P. Distinctive toxicity of TiO2 rutile/anatase mixed phase nanoparticles on Caco-2 cells. Chem Res. Toxicol. 2012, 25, 646–655. [Google Scholar] [CrossRef]
- Dorier, M.; Beal, D.; Marie-Desvergne, C.; Dubosson, M.; Barreau, F.; Houdeau, E.; Herlin-Boime, N.; Carriere, M. Continuous in vitro exposure of intestinal epithelial cells to E171 food additive causes oxidative stress, inducing oxidation of DNA bases but no endoplasmic reticulum stress. Nanotoxicology 2017, 11, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Dorier, M.; Brun, E.; Veronesi, G.; Barreau, F.; Pernet-Gallay, K.; Desvergne, C.; Rabilloud, T.; Carapito, C.; Herlin-Boime, N.; Carriere, M. Impact of anatase and rutile titanium dioxide nanoparticles on uptake carriers and efflux pumps in Caco-2 gut epithelial cells. Nanoscale 2015, 7, 7352–7360. [Google Scholar] [CrossRef] [PubMed]
- Dorier, M.; Tisseyre, C.; Dussert, F.; Beal, D.; Arnal, M.E.; Douki, T.; Valdiglesias, V.; Laffon, B.; Fraga, S.; Brandao, F.; et al. Toxicological impact of acute exposure to E171 food additive and TiO2 nanoparticles on a co-culture of Caco-2 and HT29-MTX intestinal cells. Mutat. Res. 2019, 845, 402980. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; DeLoid, G.M.; Bitounis, D.; De La Torre-Roche, R.; White, J.C.; Zhang, Z.; Ho, C.G.; Ng, K.W.; Eitzer, B.D.; Demokritou, P. Co-exposure to the food additives SiO2 (E551) or TiO2 (E171) and the pesticide boscalid increases cytotoxicity and bioavailability of the pesticide in a tri-culture small intestinal epithelium model: Potential health implications. Environ. Sci. Nano 2019, 6, 2786–2800. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jia, M.X.; Deng, J.; Wang, J.H.; Zuberi, Z.; Yang, S.; Ba, J.; Chen, Z. MicroRNA Response and toxicity of potential pathways in human colon cancer cells exposed to titanium dioxide nanoparticles. Cancers 2020, 12, 1236. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Chakraborty, A.; Mukherjee, A. Cytotoxic, genotoxic and the hemolytic effect of titanium dioxide (TiO2) nanoparticles on human erythrocyte and lymphocyte cells in vitro. J. Appl. Toxicol. 2013, 33, 1097–1110. [Google Scholar] [CrossRef]
- Dai, X.; Liu, R.; Li, N.; Yi, J. Titanium dioxide nanoparticles induce in vitro autophagy. Hum. Exp. Toxicol. 2019, 38, 56–64. [Google Scholar] [CrossRef]
- Shukla, R.K.; Kumar, A.; Gurbani, D.; Pandey, A.K.; Singh, S.; Dhawan, A. TiO(2) nanoparticles induce oxidative DNA damage and apoptosis in human liver cells. Nanotoxicology 2013, 7, 48–60. [Google Scholar] [CrossRef]
- Shi, Z.; Niu, Y.; Wang, Q.; Shi, L.; Guo, H.; Liu, Y.; Zhu, Y.; Liu, S.; Liu, C.; Chen, X.; et al. Reduction of DNA damage induced by titanium dioxide nanoparticles through Nrf2 in vitro and in vivo. J. Hazard. Mater. 2015, 298, 310–319. [Google Scholar] [CrossRef]
- Brzicova, T.; Sikorova, J.; Milcova, A.; Vrbova, K.; Klema, J.; Pikal, P.; Lubovska, Z.; Philimonenko, V.; Franco, F.; Topinka, J.; et al. Nano-TiO2 stability in medium and size as important factors of toxicity in macrophage-like cells. Toxicol. In Vitro 2019, 54, 178–188. [Google Scholar] [CrossRef]
- Popp, L.; Tran, V.; Patel, R.; Segatori, L. Autophagic response to cellular exposure to titanium dioxide nanoparticles. Acta Biomater. 2018, 79, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Costa, C.; Silva, S.; Costa, S.; Dhawan, A.; Oliveira, P.A.; Teixeira, J.P. Effects of titanium dioxide nanoparticles in human gastric epithelial cells in vitro. Biomed. Pharmacother. 2014, 68, 59–64. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Ling, C.; Xu, M.; Hu, M.; Wang, H.; Liu, J.; Song, G.; Liu, J. Oxidative damage induced by nano-titanium dioxide in rats and mice: A systematic review and meta-analysis. Biol. Trace Elem. Res. 2020, 194, 184–202. [Google Scholar] [CrossRef] [PubMed]
- Canli, E.G.; Atli, G.; Canli, M. Response of the antioxidant enzymes of the erythrocyte and alterations in the serum biomarkers in rats following oral administration of nanoparticles. Environ. Toxicol. Pharmacol. 2017, 50, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, K.M.; El-Amir, Y.O. Protective effects of thymoquinone and avenanthramides on titanium dioxide nanoparticles induced toxicity in Sprague-Dawley rats. Pathol. Res. Pract. 2017, 213, 13–22. [Google Scholar] [CrossRef]
- Hassanein, K.M.A.; El-Amir, Y.O. Ameliorative effects of thymoquinone on titanium dioxide nanoparticles induced acute toxicity in rats. Int. J. Vet. Sci. Med. 2018, 6, 16–21. [Google Scholar] [CrossRef]
- Pelclova, D.; Zdimal, V.; Kacer, P.; Komarc, M.; Fenclova, Z.; Vlckova, S.; Zikova, N.; Schwarz, J.; Makes, O.; Navratil, T.; et al. Markers of lipid oxidative damage among office workers exposed intermittently to air pollutants including nanoTiO2 particles. Rev. Environ. Health 2017, 32, 193–200. [Google Scholar] [CrossRef]
- Charles, S.; Jomini, S.; Fessard, V.; Bigorgne-Vizade, E.; Rousselle, C.; Michel, C. Assessment of the in vitro genotoxicity of TiO2 nanoparticles in a regulatory context. Nanotoxicology 2018, 12, 357–374. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019, 10, 128. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Nunez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Winter, M.; Beer, H.D.; Hornung, V.; Kramer, U.; Schins, R.P.; Forster, I. Activation of the inflammasome by amorphous silica and TiO2 nanoparticles in murine dendritic cells. Nanotoxicology 2011, 5, 326–340. [Google Scholar] [CrossRef] [PubMed]
- Baron, L.; Gombault, A.; Fanny, M.; Villeret, B.; Savigny, F.; Guillou, N.; Panek, C.; Le Bert, M.; Lagente, V.; Rassendren, F.; et al. The NLRP3 inflammasome is activated by nanoparticles through ATP, ADP and adenosine. Cell Death Dis. 2015, 6, e1629. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.C. Microbiota dysbiosis and barrier dysfunction in inflammatory bowel disease and colorectal cancers: Exploring a common ground hypothesis. J. Biomed. Sci. 2018, 25, 79. [Google Scholar] [CrossRef] [PubMed]
- Karimipour, M.; Zirak Javanmard, M.; Ahmadi, A.; Jafari, A. Oral administration of titanium dioxide nanoparticle through ovarian tissue alterations impairs mice embryonic development. Int. J. Reprod. Biomed. 2018, 16, 397–404. [Google Scholar] [CrossRef]
- Jin, C.; Tang, Y.; Fan, X.Y.; Ye, X.T.; Li, X.L.; Tang, K.; Zhang, Y.F.; Li, A.G.; Yang, Y.J. In vivo evaluation of the interaction between titanium dioxide nanoparticle and rat liver DNA. Toxicol. Ind. Health 2013, 29, 235–244. [Google Scholar] [CrossRef]
- Latif, M.A.; Jabeen, F.; Ali, M.; Rasul, A.; Naz, S.; Akram, M. Neurotoxic effects of titanium dioxide nanoparticles on the brain of male sprague dawley rats. Pak. J. Pharm. Sci. 2019, 32, 2311–2316. [Google Scholar]
- Kazimirova, A.; Baranokova, M.; Staruchova, M.; Drlickova, M.; Volkovova, K.; Dusinska, M. Titanium dioxide nanoparticles tested for genotoxicity with the comet and micronucleus assays in vitro, ex vivo and in vivo. Mutat. Res. 2019, 843, 57–65. [Google Scholar] [CrossRef]
- Kim, H.; Jeon, D.; Oh, S.; Nam, K.; Son, S.; Gye, M.C.; Shin, I. Titanium dioxide nanoparticles induce apoptosis by interfering with EGFR signaling in human breast cancer cells. Environ. Res. 2019, 175, 117–123. [Google Scholar] [CrossRef]
- Montalvo-Quiros, S.; Luque-Garcia, J.L. Combination of bioanalytical approaches and quantitative proteomics for the elucidation of the toxicity mechanisms associated to TiO2 nanoparticles exposure in human keratinocytes. Food Chem. Toxicol. 2019, 127, 197–205. [Google Scholar] [CrossRef]
- Du, X.; Gao, S.; Hong, L.; Zheng, X.; Zhou, Q.; Wu, J. Genotoxicity evaluation of titanium dioxide nanoparticles using the mouse lymphoma assay and the Ames test. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 838, 22–27. [Google Scholar] [CrossRef]
- Freyre-Fonseca, V.; Tellez-Medina, D.I.; Medina-Reyes, E.I.; Cornejo-Mazon, M.; Lopez-Villegas, E.O.; Alamilla-Beltran, L.; Ocotlan-Flores, J.; Chirino, Y.I.; Gutierrez-Lopez, G.F. Morphological and physicochemical characterization of agglomerates of titanium dioxide nanoparticles in cell culture media. J. Nanomater. 2016, 2016, 5937932. [Google Scholar] [CrossRef]
- Kobayashi, K.; Kubota, H.; Hojo, R.; Miyagawa, M. Effective dispersal of titanium dioxide nanoparticles for toxicity testing. J. Toxicol. Sci. 2019, 44, 515–521. [Google Scholar] [CrossRef] [PubMed]
- JRC. Titanium dioxide, NM-100, NM-101, NM-102, NM-103, NM-104, NM-105: Characterisation and physico-chemical properties. JRC 2014. [Google Scholar] [CrossRef]
- Action, N.J. Facilitating the Safety Evaluation of Manufactured Nanomaterials by Characterising Their Potential Genotoxic Hazard; NANOGENOTOX Final Report; French Agency for Food, Environmental and Occupational Health & Safety (ANSES): Maisons-Alfort, France, 2013. [Google Scholar]
- Jensen, K.; Kembouche, Y.; Christiansen, E.; Jacobsen, N.; Wallin, H.; Guiot, C.; Spalla, O.; Witschger, O. The Generic NANOGENOTOX Dispersion Protocol—Standard Operation Procedure (SOP); NANOGENOTOX Deliverable Report; French Agency for Food, Environmental and Occupational Health & Safety (ANSES): Maisons-Alfort, France, 2011; Volume 3. [Google Scholar]
- Donovan, A.R.; Adams, C.D.; Ma, Y.; Stephan, C.; Eichholz, T.; Shi, H. Single particle ICP-MS characterization of titanium dioxide, silver, and gold nanoparticles during drinking water treatment. Chemosphere 2016, 144, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assuncao, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carriere, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Abdelkhaliq, A.; van der Zande, M.; Undas, A.K.; Peters, R.J.B.; Bouwmeester, H. Impact of in vitro digestion on gastrointestinal fate and uptake of silver nanoparticles with different surface modifications. Nanotoxicology 2020, 14, 111–126. [Google Scholar] [CrossRef]
- Sohal, I.S.; Cho, Y.K.; O’Fallon, K.S.; Gaines, P.; Demokritou, P.; Bello, D. Dissolution Behavior and Biodurability of Ingested Engineered Nanomaterials in the Gastrointestinal Environment. ACS Nano 2018, 12, 8115–8128. [Google Scholar] [CrossRef]
- Sohal, I.S.; O’Fallon, K.S.; Gaines, P.; Demokritou, P.; Bello, D. Ingested engineered nanomaterials: State of science in nanotoxicity testing and future research needs. Part. Fibre Toxicol. 2018, 15, 29. [Google Scholar] [CrossRef]
- Bettencourt, A.; Goncalves, L.M.; Gramacho, A.C.; Vieira, A.; Rolo, D.; Martins, C.; Assuncao, R.; Alvito, P.; Silva, M.J.; Louro, H. Analysis of the characteristics and cytotoxicity of titanium dioxide nanomaterials following simulated In Vitro digestion. Nanomaterials 2020, 10, 1516. [Google Scholar] [CrossRef]
- Terrisse, H.M.-B.M. Titanium Dioxide as Food Additive; INTECH: London, UK, 2017. [Google Scholar]
- Hwang, J.S.; Yu, J.; Kim, H.M.; Oh, J.M.; Choi, S.J. Food additive titanium dioxide and its fate in commercial foods. Nanomaterials 2019, 9, 1175. [Google Scholar] [CrossRef]
- Noben, M.; Vanhove, W.; Arnauts, K.; Santo Ramalho, A.; Van Assche, G.; Vermeire, S.; Verfaillie, C.; Ferrante, M. Human intestinal epithelium in a dish: Current models for research into gastrointestinal pathophysiology. United Eur. Gastroenterol. J. 2017, 5, 1073–1081. [Google Scholar] [CrossRef]
- Augustyniak, J.; Bertero, A.; Coccini, T.; Baderna, D.; Buzanska, L.; Caloni, F. Organoids are promising tools for species-specific in vitro toxicological studies. J. Appl. Toxicol. 2019, 39, 1610–1622. [Google Scholar] [CrossRef] [PubMed]
- Kijanska, M.; Kelm, J. In vitro 3D spheroids and microtissues: ATP-based cell viability and toxicity assays. In Assay Guidance Manual; Sittampalam, G.S., Grossman, A., Brimacombe, K., Arkin, M., Auld, D., Austin, C.P., Baell, J., Bejcek, B., Caaveiro, J.M.M., Chung, T.D.Y., et al., Eds.; Eli Lilly & Company and the National Center for Advancing: Bethesda, MD, USA, 2004. [Google Scholar]
- Michels, B.E.; Mosa, M.H.; Grebbin, B.M.; Yepes, D.; Darvishi, T.; Hausmann, J.; Urlaub, H.; Zeuzem, S.; Kvasnicka, H.M.; Oellerich, T.; et al. Human colon organoids reveal distinct physiologic and oncogenic Wnt responses. J. Exp. Med. 2019, 216, 704–720. [Google Scholar] [CrossRef] [PubMed]
- Onozato, D.; Yamashita, M.; Nakanishi, A.; Akagawa, T.; Kida, Y.; Ogawa, I.; Hashita, T.; Iwao, T.; Matsunaga, T. Generation of intestinal organoids suitable for pharmacokinetic studies from human induced pluripotent stem cells. Drug Metab. Dispos. 2018, 46, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Dinse, G.E.; Peddada, S.D.; Harris, S.F.; Elmore, S.A. Comparison of NTP historical control tumor incidence rates in female Harlan Sprague Dawley and Fischer 344/N Rats. Toxicol. Pathol. 2010, 38, 765–775. [Google Scholar] [CrossRef]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef]
- Bode, G.; Clausing, P.; Gervais, F.; Loegsted, J.; Luft, J.; Nogues, V.; Sims, J.; Steering Group of the RETHINK Project. The utility of the minipig as an animal model in regulatory toxicology. J. Pharmacol. Toxicol. Methods 2010, 62, 196–220. [Google Scholar] [CrossRef]
- Henze, L.J.; Koehl, N.J.; O’Shea, J.P.; Kostewicz, E.S.; Holm, R.; Griffin, B.T. The pig as a preclinical model for predicting oral bioavailability and in vivo performance of pharmaceutical oral dosage forms: A PEARRL review. J. Pharm. Pharmacol. 2019, 71, 581–602. [Google Scholar] [CrossRef]



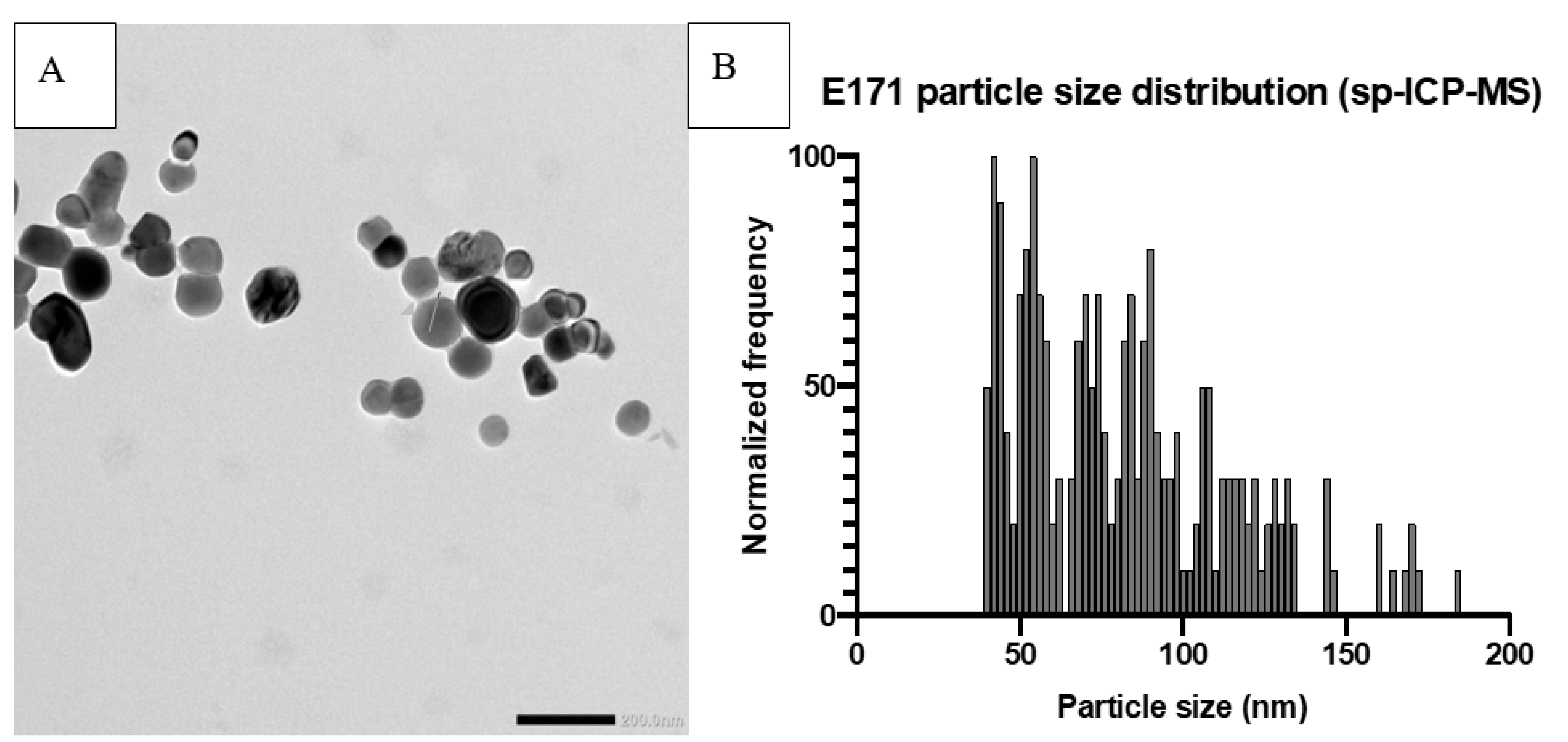{kind=link}
{kind=link}
| Author | Year | Country | Mean (mg/kg bw/Day) | 95th Percentile (mg/kg bw/Day) |
|---|---|---|---|---|
| Wu [50] | 2020 | USA | 0.15–3.9 * (PCP survey and usage patterns, no food included) | n/a |
| EFSA [21] | 2016 | Europe | <11 months: 0.2–0.8 1–3 years: 0.6–4.6 3–9 years: 0.9–5.5 10–17 years: 0.4–4.1 18–64 years: 0.3–4.0 >65 years: 0.2–2.8 | <11 months: 0.7–3.9 1–3 years: 2.0–6.8 3–9 years: 2.4–14.8 10–17 years: 1.3–10.8 18–64 years: 1.1–9.7 >65 years: 0.5–7.0 |
| Rompelberg [49] | 2016 | NL | 2–6 years: 0.66–0.70 7–69 years: 0.16–0.18 >69 years: 0.05–0.07 | 2–6 years: 1.19–1.40 7–69 years: 0.47–0.54 >69 years: 0.20–0.28 |
| Bachler [51] | 2015 | DE | “Other Children”: ~2 Toddlers, adolescents, adults, elderly: 0.5–1 | “Other Children”: ~0.7–7.2 Toddlers, adolescents, adults, elderly: ~0.1–4.2 |
| Sprong [52] | 2015 | NL | 2–6 years: 1.3–1.5 7–69 years: 0.6–0.7 >70 years: 0.5–0.6 | 2–6 years: 4.5–5.6 7–69 years: 2.6–3.0 >70 years: 1.7–2.2 |
| Christensen [53] | 2015 | DK | Children: 2 Adults: 1 | n/a |
| Weir [27] | 2012 | UK | <10 years 2–3 >10 years: 1 | n/a |
| Weir [27] | 2012 | US | <10 years: 1–2 >10 years: 0.2–0.7 | n/a |
| Powell [54] | 2010 | UK | 5 * | n/a |
| Reference | Testing | Material | Ø Primary Size (nm) | Hydrodynamic Diameter (nm) | Zeta Potential (mV) | Species/Sex ♀♂ | Duration (days) | Dose (mg/kg bw/day) | Administration | Observation |
|---|---|---|---|---|---|---|---|---|---|---|
| Chen 2019 [99] | 90-days repeated dose | TiO2 NP Spherical anatase (purity 99.90%) | 29 ± 9 | In water ~30 In gastric juices ~105 In intestinal juices ~110 | In water ~+10 In gastric juices ~+5 In intestinal juices ~−15 | Rats Sprague-Dawley | 90 | 0, 2, 10, 50 | Oral gavage in ultra-pure water |
|
| Moradi 2019 [86] | 14-days repeated dose | TiO2 NP 80% anatase, 20% rutile | 20 | Not assessed | Not assessed | Rats ♂ Wistar | 14 | 300 | Oral gavage in bi-distilled water |
|
| Jensen 2019 [94] | 70-day repeated exposure | E171 99.8% anatase, 0.2% rutile | 135 ± 6 305 ± 61 900 ± 247 | In filtered water, 2% FBS 270 ± 25 | Not assessed | Rats ♀ Zucker | 70 | 50 *, 500 * | Oral gavage in sterile water 2% FBS |
|
| Blevins 2019 [17] | 100-days repeated dose | E171 85% anatase 25% rutile | 110, 115 | In food, no assessment of E171 characteristics in food | In food, no assessment of E171 characteristics in food | Rats ♂ Wistar (immune response and DMH colon carcinogenesis model) | 7100 | 40, 400, 5000 | Ingestion via food in Purina 5002R33 diet |
|
| Bettini 2017 [14] | 100-days repeated dose (E171), 7-days repeater dose (E171 and NM-105) | E171 (45% nanosized by particles number), NM-105 85% anatase 25% rutile | E171: 118 ± 53 NM-105: 22 ± 1 | In purified water E171: 373 ± 20 NM-105: 192 ± 2 | In purified water E171: −23.9 ± 2.4 NM-105: +5.03 ± 0.02 | Rats ♂ Wistar (immune response) | 7100 | 10 | Oral gavage in purified water |
|
| Bettini 2017 [14] | 100-days repeated dose | E171, | E171: 118 ± 53 | In purified water E171: 373 ± 20 | In purified water E171: −23.9 ± 2.4 | Rats ♂ Wistar (normal wild type and DMH colon carcinogenesis model) Wistar | 7100 | 0.2, 10 | Oral gavage in purified water |
|
| Martins 2017 [18] | 45-days repeated dose | TiO2 NP | 41.99 ± 1.63 | Sodium citrate buffer 0.1M, pH 4.5 447.67 ± 6.43 | Not assessed | Rats ♂ Wistar | 45 | 0.5 | Oral gavage in sodium citrate buffer |
|
| Donner 2016 [67] | OECD 474 | TiO2 **** | **** | **** | **** | Rats ♂♀ Sprague-Dawley | 2, 3 | 500, 1000, 2000 | Single dose oral gavage in water |
|
| Warheit 2015 [83] | OECD 407 | TiO2 2 types of rutile | 173 | In 0.01% tetrasodium hexametaphosphate + MilliQ 253 | **** | Rats ♂ Sprague-Dawley | 28 | 24000 | Oral gavage in sterile water |
|
| Warheit 2015 [6] | OECD 414 | TiO2 **** | 20–206 | **** | **** | Rats ♀ Sprague-Dawley, Wistar | 14 | 100, 300, 1000 | Oral gavage in sterile water |
|
| Orazizadeh 2014 [87] | 14-days repeated dose | TiO2 NP | 50–100 | Not assessed | Not assessed | Rats ♂ Wistar | 14 | 300 | Oral gavage in milli-Q water |
|
| Hu 2020 [95] | 56-, 182-days repeated dose | TiO2 NP | 25.37 ± 4.17 | In PBS 34.34 ± 6.33 | In PBS “negative value” | Mice ♂ ICR (Y/A) | 56, 182 | 50 | Oral gavage in PBS |
|
| Ali 2019 [35] | 5-days repeated dose | TiO2 NP | 21, 80 | Not assessed | Not assessed | Mice ♂ Swiss-Albino | 5 | 50, 250, 500 | Oral gavage in 0.9% physiological saline solution |
|
| Chakrabarti2019 [91] | OECD 408 | TiO2 NP | 58.25± 8.11 | Not assessed | Not assessed | Mice ♂♀ Swiss-Albino | 90 | 200, 500 | “orally” in water |
|
| Proquin 2018 [92] | 21-days repeated dose | E171 | 535 | In water at 1 mg/mL 316.8 ± 282.4 | In water at 1 mg/mL −12.78 ± 0.52 | Mice ♂♀ BALB/c | 2, 7, 14, 21 | 5 | Oral gavage in water |
|
| Hu 2018 [88] | 182-days repeated exposure | TiO2 NP | 26.42± 7.73 | In PBS 42.15 ± 6.71 | In PBS “negative values” | Mice ICR | 182 | 10, 20, 50, 100, 200 | Oral gavage in PBS |
|
| Ruiz 2017 [89] | 8-days repeated dose | TiO2 rutile | 30–50 | Not assessed | Not assessed | Mice ♀ C57BL/6 NLRP3− (DSS colitis model) | 8 | 50, 500 | Oral gavage in water |
|
| Urrutia-Ortega 2016 [15] | 77-days repeated dose | E171 (purity > 99%) | 382 502 626 | In water pH7 300 | In water pH7 -30 | Mice ♂♀ BALB/c+ (CAC model) | 45 *** | 5 | Oral gavage in water |
|
| Sycheva 2011 [93] | 7-days repeated dose | TiO2 MP TiO2 NP | MP: 160 ± 59.4 NP: 33 ± 16.7 | Not assessed | Not assessed | Mice ♂♀ CBAB6F1 | 7 | 40, 200, 1000 | Oral gavage in distilled water |
|
| Trouiller 2009 [90] | 5-days repeated exposure | TiO2 NP 75% anatase, 25% rutile (purity 99.5%) | 21 | In water 160 ± 5 | Not assessed | Mice C57B1/6Jpun/pun | 5 | 60, 120, 300, 600 ** | Orally in drinking water |
|
| Reference | Material | Ø Primary Size (nm) | Hydrodynamic Diameter (nm) | Zeta-Potential (mV) | Cell Type | Duration (h) | Conc. (µg/mL) | Observation |
|---|---|---|---|---|---|---|---|---|
| Teubl 2015 [57] | NM-103 rutile dimethicone coated NM-104 rutile glycerol coated | NM-103: 20 NM-104: 20 | In PBS NM-103: 1819 ± 61.56 NM-104: 1539 ± 489 In artificial saliva NM-103: 3061 ± 134.4 NM-104: 2597 ± 426 | In PBS NM-103: −25.2 ± 8.2 NM-104: −21 ± 4.3 Inartificial saliva NM-103: −9.5 ± 3.7 NM-104: −9.1 ± 3.3 | Human buccal epithelial TR146 | 4 | 1–200 |
|
| Teuble 2015 [56] | NM-100 anatase NM-101 anatase NM-105 80% anatase, 20% rutile | NM-100: 148 ± 45/34 ± 15 NM-101: 28 ± 8 NM-105: 36 ± 10 | In PBS (d0.5) NM-100: 1346 ± 76 NM-101: 627 ± 5 NM-105: 868 ± 16 In artificial saliva (d0.5) NM-100: 2850 ± 482 NM-101: 1095 ± 28 NM-105: 1183 ± 74 | In PBS (d0.5) NM-100: −10.3 ± 8.2 NM-101: −28.7 ± 4.3 NM-105: −24.0 ± 3.4 Inartificial saliva (d0.5) NM-100: −12.0 ± 5.7 NM-101: −8.46 ± 4.8 NM-105: −11.7 ± 5.6 | Human buccal epithelial TR146 | 4, 24 | 1–200 |
|
| Kim 2019 [120] | TiO2 NPs anatase 99%anatase | 15 | n/a | n/a | Periodontal Ligament cells | 0.5, 1, 3, 8, 24, 48 | 2.5–50 |
|
| Gerloff 2012 [121] | TiO2 NPs * (anatase/rutile) | 4–215 * | Was assessed * | Was assessed * | Caco-2 | 4, 24 | 20, 80 ** |
|
| Botelho 2014 [133] | E171 80% anatase, 20% rutile | 21 | In milli-Q water or RPMI supplemented with 10% FBS, or 2% BSA in PBS 420.7 | In milli-Q water or RPMI supplemented with 10% FBS, or 2% BSA in PBS −27.8 | AGS human gastric epithelial | 3, 6, 24 | 20–150 |
|
| Dorier 2015 [123] | TiO2 NPs A12 R20 | A12: 12 R20: 20 | In water A12: 132 ± 1 R20: >1000 In exposure medium A12: 320 R20: >1000 | In water A12: −20,0 ± 0.6 R20: −19.5 ± 0.9 In exposure medium A12: −10.8 ± 0.6 R20: −11.7 ± 0.8 | Caco-2 | 6, 24, 48 | 50 |
|
| Dorier 2017 [122] | A12 E171 * P25 (anatase/rutile) | A12: 12 ± 3 P25: 24 ± 6 E171: 118 ± 53 * | In water A12: 85 ± 2.9 E171 *: 415.6 ± 69.5 P25: 157.6 ± 1.0 In cell culture medium A12: 447.9 ± 0.3 E171 *: 739.3 ± 355.3 P25: 439.9 ± 6.7 | In cell culture medium A12: −10.8 ± 0.6 E171 *: −19 ± 0.7 P25: −11.2 ± 0.8 | Caco-2/HT29-MTX (co-culture) | 6, 48, 3 weeks | 0–200 |
|
| Proquin 2017 [16] | E171 TiO2 MPs TiO2 NPs 99.5% anatase | E171: 50–250 TiO2 MPs: 535 TiO2 NPs: 10–25 | In DMEM 0.05% BSA E171: 669.62 ± 30.13 TiO2 MPs: 1385.83 ± 38.85 TiO2 NPs: > 1000 HBSS E171: > 1000 TiO2 MPs: > 1000 TiO2 NPs: > 1000 McCoy + 10% FBS at 1 mg/mL E171: 316 ± 282.4 | In DMEM 0.05% BSA E171: −12.97 ± 0.29 TiO2 MPs: −14.10 ±0.56 TiO2 NPs: −13.12 ± 0.44 HBSS E171: −4.39 ± 0.12 TiO2 MPs: −5.62 ± 0.9 TiO2 NPs: −6.08 ± 0.09 McCoy +10% FBS at 1 mg/mL E171: −12.56 ± 8.3 | Caco-2, HCT116 | 24 | 0.001–1000 |
|
| Popp 2018 [132] | TiO2 NPs NanoAmor, mkNano MKN-TiO2-A100 (purity > 98%) | NanoAmor:15 mkNano: 50 MKN-TiO2-A100: 100 | In DMEM at 25 µg/mL NanoAmor: ~300 mkNano: ~800 MKN-TiO2-A100: ~500 | In DMEM at 25 µg/mL NanoAmor: ~−15 mkNano: ~−12 MKN-TiO2-A100: ~−12 | HeLa | 24, 48, 72 | 0–500 |
|
| Pedata 2019 [58] | TiO2 NPs P25 | P25: 21 | Not assessed | Not assessed | Caco-2 (differentiated) | 4, 24, 48 | 42, 84 |
|
| Dorier 2019 [124] | E171 A12 NM-105 | E171: 118 ± 53 A12: 12 ± 3 NM-105: 24 ± 6 | In ultrapure sterile water at 10 mg/mL E171:415 ± 69 A12: 85 ± 3 NM-105: 158 ± 1 In DMEM + 10% FBS E171:739 ± 355 A12: 448 ± 1 NM-105: 440 ± 7 | In ultrapure sterile water at 10 mg/mL E171: −19 ± 1 A12: −11 ± 1 NM-105: −11 ± 1 | Caco-2/HT29-MTX (co-culture) | 6, 24, 48 | 0–200 |
|
| Cao 2019 [125] | E171 | E171: 113.4 ± 37.2 | Not assessed | Not assessed | Caco-2/HT29-MTX (co-culture), + Raji B | 6, 24 | 10, 150 *** |
|
| Li 2020 [126] | TiO2 NPs | 25 | Not assessed | Not assessed | HCT116, NCM4660 | 24 | 2, 30 |
|
| Ghosh 2013 [127] | TiO2 NPs | 100 | In PBS 6180.73 | Not assessed | Human lymphocyte, human erythrocytes | 3 | 25, 50, 100 |
|
| Dai 2019 [128] | TiO2 NPs rutile | 30–50 50–100 | Not assessed | Not assessed | RAW 264.7 | 24 | 50, 100, 200, 300, 400 |
|
| Brzicova 2019 [131] | TiO2 NPs | 20–60 * | Was assessed* | Was assessed * | THP-1 | 24 | 32, 128, 256 |
|
| Shukla 2013 [129] | TiO2 NPs Anatase (purity 99.7%) | 30–70 * | In Milli-Q water 192.5 ± 2.00 In CMEM 124.9 ± 3.20 | In Milli-Q water −11.4 ± 0.25 In CMEM −17.6 ± 0.48 | HepG2 | 6, 24, 48 | 1, 10, 20, 40, 80 |
|
| Shi 2015 [130] | TiO2 NPs Anatase (purity 99.7%) | 10–25 | DMEM + 1% FCS 366.5 ± 30 | DMEM + 1% FCS −26.93 ± 0.43 | HepG2 | 24 | 0.1, 1, 10 |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bischoff, N.S.; de Kok, T.M.; Sijm, D.T.H.M.; van Breda, S.G.; Briedé, J.J.; Castenmiller, J.J.M.; Opperhuizen, A.; Chirino, Y.I.; Dirven, H.; Gott, D.; et al. Possible Adverse Effects of Food Additive E171 (Titanium Dioxide) Related to Particle Specific Human Toxicity, Including the Immune System. Int. J. Mol. Sci. 2021, 22, 207. https://doi.org/10.3390/ijms22010207
Bischoff NS, de Kok TM, Sijm DTHM, van Breda SG, Briedé JJ, Castenmiller JJM, Opperhuizen A, Chirino YI, Dirven H, Gott D, et al. Possible Adverse Effects of Food Additive E171 (Titanium Dioxide) Related to Particle Specific Human Toxicity, Including the Immune System. International Journal of Molecular Sciences. 2021; 22(1):207. https://doi.org/10.3390/ijms22010207
Chicago/Turabian StyleBischoff, Nicolaj S., Theo M. de Kok, Dick T.H.M. Sijm, Simone G. van Breda, Jacco J. Briedé, Jacqueline J.M. Castenmiller, Antoon Opperhuizen, Yolanda I. Chirino, Hubert Dirven, David Gott, and et al. 2021. "Possible Adverse Effects of Food Additive E171 (Titanium Dioxide) Related to Particle Specific Human Toxicity, Including the Immune System" International Journal of Molecular Sciences 22, no. 1: 207. https://doi.org/10.3390/ijms22010207
APA StyleBischoff, N. S., de Kok, T. M., Sijm, D. T. H. M., van Breda, S. G., Briedé, J. J., Castenmiller, J. J. M., Opperhuizen, A., Chirino, Y. I., Dirven, H., Gott, D., Houdeau, E., Oomen, A. G., Poulsen, M., Rogler, G., & van Loveren, H. (2021). Possible Adverse Effects of Food Additive E171 (Titanium Dioxide) Related to Particle Specific Human Toxicity, Including the Immune System. International Journal of Molecular Sciences, 22(1), 207. https://doi.org/10.3390/ijms22010207