Flavodoxins as Novel Therapeutic Targets against Helicobacter pylori and Other Gastric Pathogens



Abstract
1. Introduction
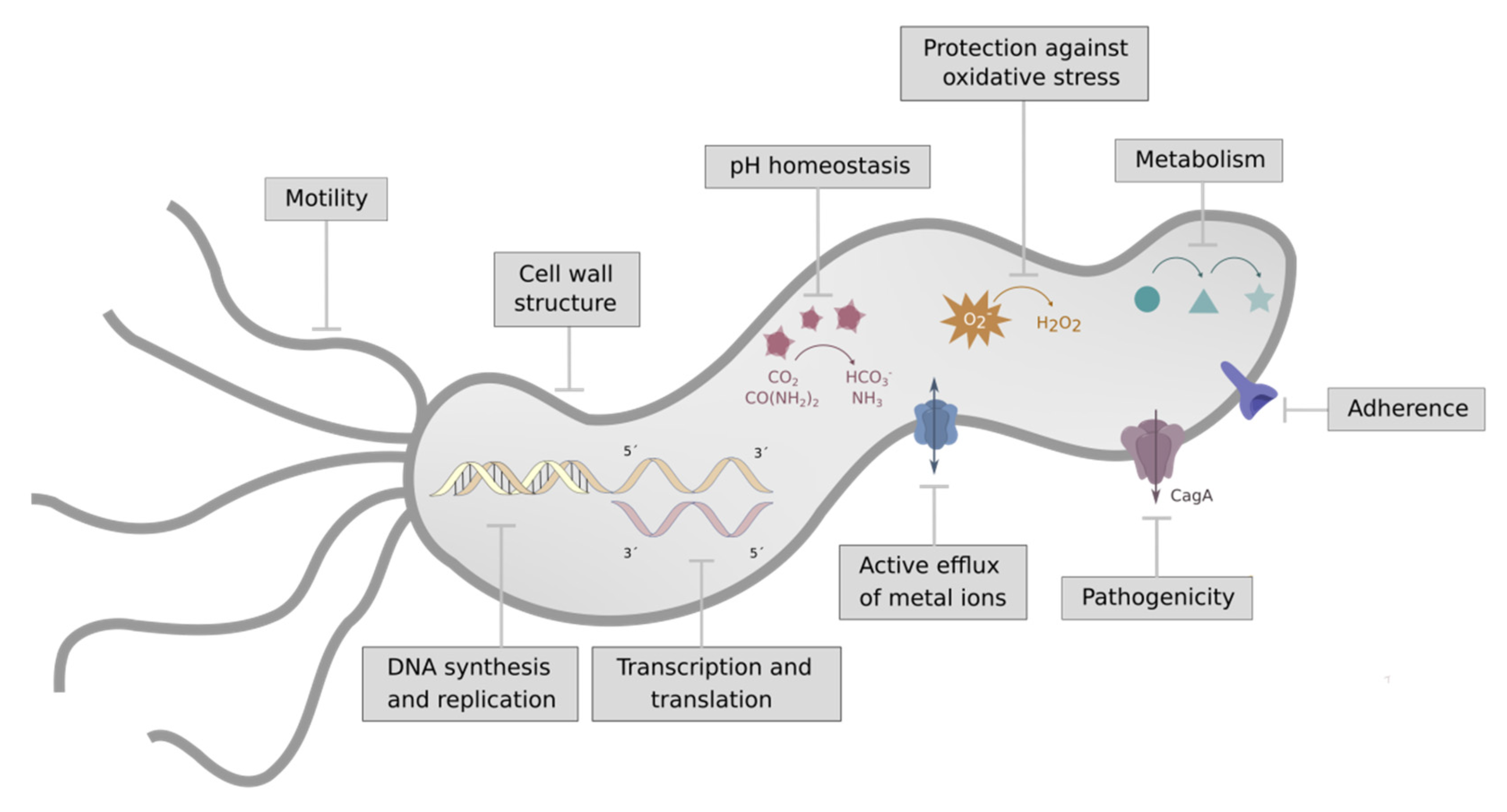
2. Targets for Hp Infection
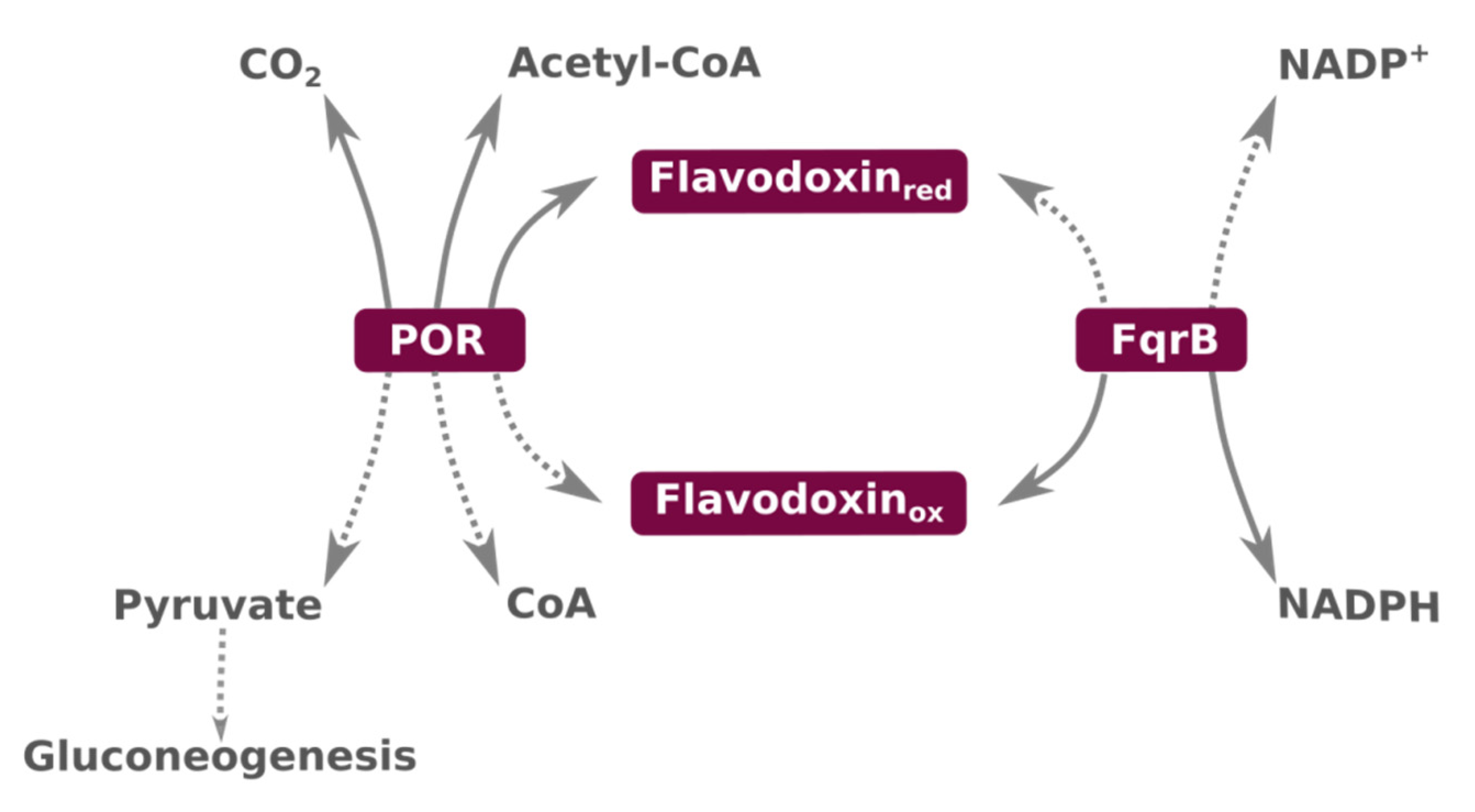
2.1. Metabolism
2.2. Cell Wall Structure
2.3. pH Homeostasis
2.4. Virulence (Adherence, Motility, and Pathogenicity)
2.5. Active Efflux of Metal Ions
2.6. Protection against Oxidative Stress
3. An Overview of Flavodoxins and of the Flavodoxin from Hp
4. Flavodoxins in other Pathogenic Bacteria and in the Gut Microbiota
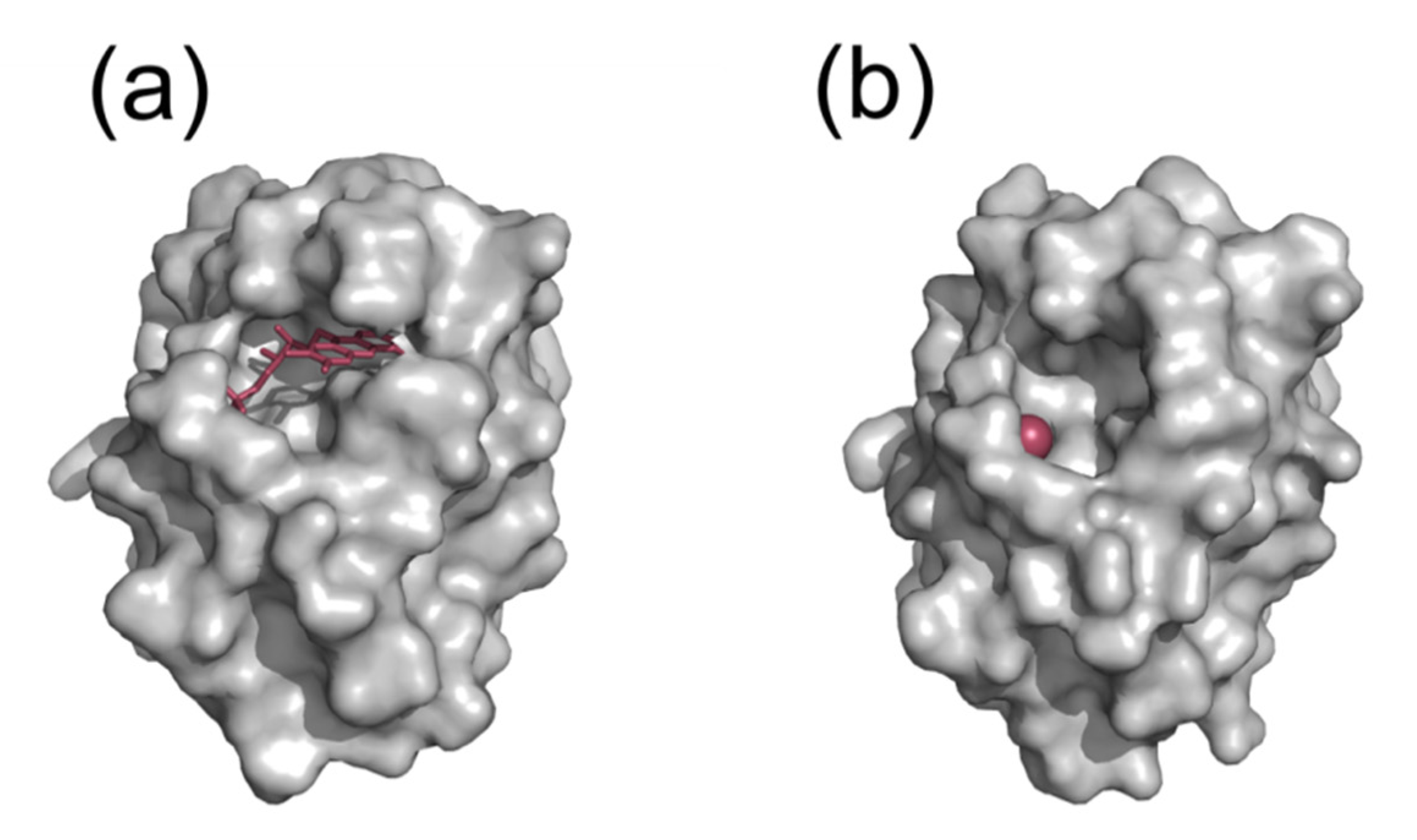
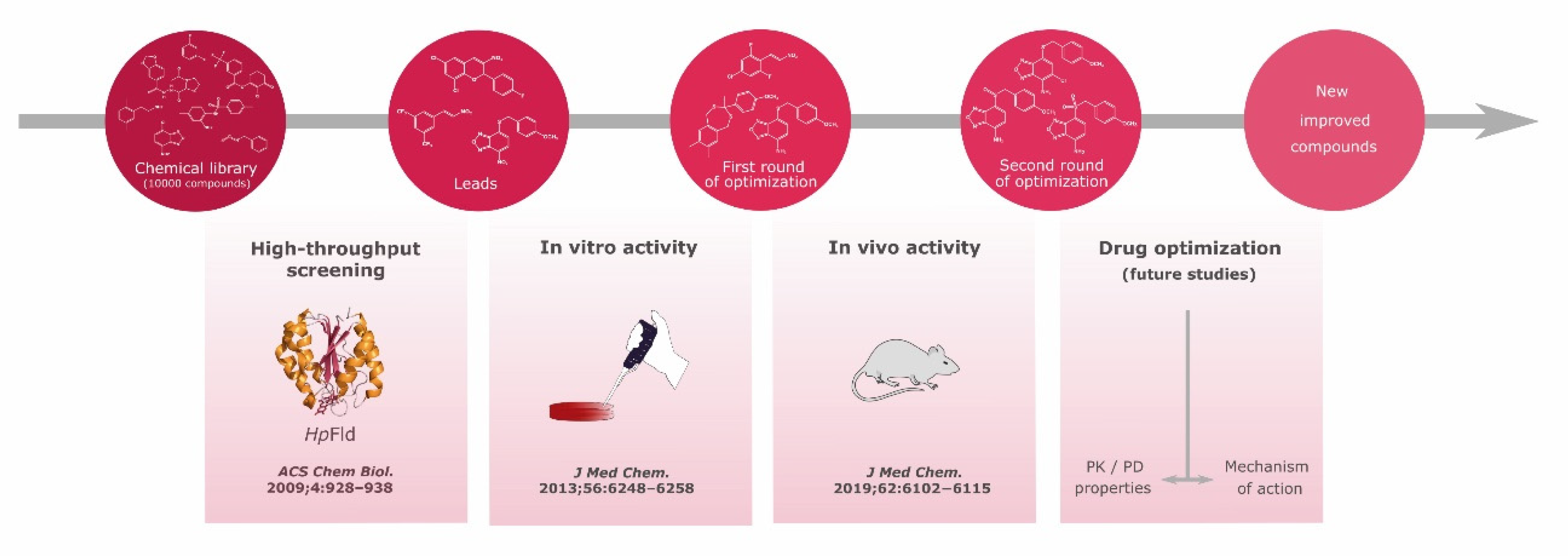
5. Discovery of Specific Inhibitors of Hp Flavodoxin Using an Approach that Can Be Transferred to Other Pathogens
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Ala (A) | Alanine |
| ATCC | American Type Culture Collection |
| DNA | Deoxyribonucleic acid |
| EUCAST | European Committee on Antimicrobial Susceptibility Testing |
| Fld | Flavodoxin |
| FMN | Flavinmononucleotide |
| FqrB | Flavodoxin:quinone reductase |
| Hp | Helicobacter pylori |
| MALT | Gastric mucosa-associated lymphoid tissue lymphoma |
| MCC | Minimal cytotoxic concentration |
| MIC | Minimal inhibitory concentration |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NCBI | National Center for Biotechnology Information |
| NMR | Nuclear Magnetic Resonance |
| PD | Pharmacodynamics |
| PDB | Protein Data Bank |
| POR | Pyruvate oxidoreductase complex |
| PPI | Proton-pump inhibitor |
| PK | Pharmacokinetics |
| TI | Therapeutic index |
| Trp (W) | Tryptophan |
| Tyr (Y) | Tyrosine |
Appendix A


References
- Malfertheiner, P.; Venerito, M.; Schulz, C. Helicobacter pylori Infection: New facts in clinical management. Curr. Treat. Options Gastroenterol. 2018, 16, 605–615. [Google Scholar] [CrossRef]
- Sjomina, O.; Pavlova, J.; Niv, Y.; Leja, M. Epidemiology of Helicobacter pylori infection. Helicobacter 2018, 23, e12514. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Williams, D.W. Chapter 7—Helicobacter pylori. In Microbiology of Waterborne Diseases; Academic Press: Cambridge, MA, USA, 2014; pp. 119–154. ISBN 978-0-12-415846-7. [Google Scholar]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global prevalence of Helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Azuma, T.; Konishi, J.; Tanaka, Y.; Hirai, M.; Ito, S.; Kato, T.; Kohli, Y. Contribution of HLA-DQA gene to host’s response against Helicobacter pylori. Lancet 1994, 343, 542–543. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Zhang, P.-Y.; Aboul-Soud, M.A.M. From inflammation to gastric cancer: Role of Helicobacter pylori (review). Oncol. Lett. 2017, 13, 543–548. [Google Scholar] [CrossRef] [PubMed]
- WHO. IARC Agents Classified by the IARC Monographs, Volumes 1–124. Available online: https://monographs.iarc.fr/list-of-classifications (accessed on 9 March 2020).
- Plummer, M.; Franceschi, S.; Vignat, J.; Forman, D.; De Martel, C. Global burden of gastric cancer attributable to Helicobacter pylori. Int. J. Cancer 2015, 136, 487–490. [Google Scholar] [CrossRef]
- Polk, D.B.; Peek, R.M. Helicobacter pylori: Gastric cancer and beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef]
- Gravina, A.G.; Zagari, R.M.; De Musis, C.; Romano, L.; Loguercio, C.; Romano, M. Helicobacter pylori and extragastric diseases: A review. World J. Gastroenterol. 2018, 24, 3204–3221. [Google Scholar] [CrossRef]
- Tsay, F.-W.; Hsu, P.-I. H pylori infection and extra-gastroduodenal diseases. J. Biomed. Sci. 2018, 26, 65. [Google Scholar] [CrossRef]
- Hu, Y.; Zhu, Y.; Lu, N.-H. Novel and effective therapeutic regimens for Helicobacter pylori in an era of increasing antibiotic resistance. Front. Cell. Infect. Microbiol. 2017, 7, 168. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori infection-the Maastricht V/Florence Consensus Report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [PubMed]
- Flores-Treviño, S.; Mendoza-Olazarán, S.; Bocanegra-Ibarias, P.; Maldonado-Garza, H.J.; Garza-González, E. Helicobacter pylori drug resistance: Therapy changes and challenges. Expert Rev. Gastroenterol. Hepatol. 2018, 12, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y.; Dore, M.P. Helicobacter pylori therapy: A paradigm shift. Expert Rev. Anti-Infect. Ther. 2016, 14, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lv, Z.; Zhong, Y.; Liu, D.; Chen, S.; Xie, Y. The internalization of Helicobacter pylori plays a role in the failure of H pylori eradication. Helicobacter 2017, 22, e12324. [Google Scholar] [CrossRef]
- Debraekeleer, A.; Remaut, H. Future perspective for potential Helicobacter pylori eradication therapies. Future Microbiol. 2018, 13, 671–687. [Google Scholar] [CrossRef]
- Abadi, A.T.B. Strategies used by Helicobacter pylori to establish persistent infection. World J. Gastroenterol. 2017, 23, 2870–2882. [Google Scholar] [CrossRef]
- Kusters, J.G.; Van Vliet, A.H.M.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Goderska, K.; Agudo-Pena, S.; Alarcon, T. Helicobacter pylori treatment: Antibiotics or probiotics. Appl. Microbiol. Biotechnol. 2018, 102, 1–7. [Google Scholar] [CrossRef]
- O’Morain, N.R.; Dore, M.P.; O’Connor, A.J.P.; Gisbert, J.P.; O’Morain, C.A. Treatment of Helicobacter pylori infection in 2018. Helicobacter 2018, 23, e12519. [Google Scholar] [CrossRef]
- Secka, O.; Berg, D.E.; Antonio, M.; Corrah, T.; Tapgun, M.; Walton, R.; Thomas, V.; Galano, J.J.; Sancho, J.; Adegbola, R.A.; et al. Antimicrobial susceptibility and resistance patterns among Helicobacter pylori strains from the gambia, West Africa. Antimicrob. Agents Chemother. 2013, 57, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Ayala, G.; Escobedo-Hinojosa, W.I.; De la Cruz-Herrera, C.F.; Romero, I. Exploring alternative treatments for Helicobacter pylori infection. World J. Gastroenterol. 2014, 20, 1450–1469. [Google Scholar] [CrossRef] [PubMed]
- Zawahir, S.; Czinn, S.J.; Nedrud, J.G.; Blanchard, T.G. Vaccinating against Helicobacter pylori in the developing world. Gut Microbes 2013, 4, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Chalker, A.F.; Minehart, H.W.; Hughes, N.J.; Koretke, K.K.; Lonetto, M.A.; Brinkman, K.K.; Warren, P.V.; Lupas, A.; Stanhope, M.J.; Brown, J.R.; et al. Systematic identification of selective essential genes in Helicobacter pylori by genome prioritization and allelic replacement mutagenesis. J. Bacteriol. 2001, 183, 1259–1268. [Google Scholar] [CrossRef]
- Sancho, J. Flavodoxins: Sequence, folding, binding, function and Beyond. Cell. Mol. Life Sci. 2006, 63, 855–864. [Google Scholar] [CrossRef]
- Cremades, N.; Bueno, M.; Toja, M.; Sancho, J. Towards a new therapeutic target: Helicobacter pylori flavodoxin. Biophys. Chem. 2005, 115, 267–276. [Google Scholar] [CrossRef][Green Version]
- Puan, K.J.; Wang, H.; Dairi, T.; Kuzuyama, T.; Morita, C.T. fldA is an essential gene required in the 2-C-methyl-D-erythritol 4-phosphate pathway for isoprenoid biosynthesis. FEBS Lett. 2005, 579, 3802–3806. [Google Scholar] [CrossRef]
- González-Bello, C. Inhibition of shikimate kinase and type II dehydroquinase for antibiotic discovery: Structure-based design and simulation studies. Curr. Top. Med. Chem. 2016, 16, 960–977. [Google Scholar] [CrossRef]
- Sánchez-Sixto, C.; Prazeres, V.F.V.; Castedo, L.; Suh, S.W.; Lamb, H.; Hawkins, A.R.; Cañada, F.J.; Jiménez-Barbero, J.; González-Bello, C. Competitive Inhibitors of Helicobacter pylori type II dehydroquinase: Synthesis, biological evaluation, and NMR studies. ChemMedChem 2008, 3, 756–770. [Google Scholar] [CrossRef]
- Prazeres, V.F.V.; Tizón, L.; Otero, J.M.; Guardado-Calvo, P.; Llamas-Saiz, A.L.; Van Raaij, M.J.; Castedo, L.; Lamb, H.; Hawkins, A.R.; González-Bello, C. Synthesis and biological evaluation of new nanomolar competitive inhibitors of Helicobacter pylori type II dehydroquinase. Structural details of the role of the aromatic moieties with essential residues. J. Med. Chem. 2010, 53, 191–200. [Google Scholar] [CrossRef]
- Duckworth, M.J.; Okoli, A.S.; Mendz, G.L. Novel Helicobacter pylori therapeutic targets: The unusual aspects. Expert Rev. Anti-Infect. Ther. 2009, 7, 835–867. [Google Scholar] [CrossRef] [PubMed]
- Pernas, M.; Blanco, B.; Lence, E.; Thompson, P.; Hawkins, A.R.; González-Bello, C. Synthesis of rigidified shikimic acid derivatives by ring-closing metathesis to imprint inhibitor efficacy against shikimate kinase enzyme. Org. Chem. Front. 2019, 6, 2514–2528. [Google Scholar] [CrossRef]
- Cheng, C.-S.; Chen, C.-H.; Luo, Y.-C.; Chen, W.-T.; Chang, S.-Y.; Lyu, P.-C.; Kao, M.-C.; Yin, H.-S. Crystal structure and biophysical characterisation of Helicobacter pylori phosphopantetheine adenylyltransferase. Biochem. Biophys. Res. Commun. 2011, 408, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-S.; Jia, K.-F.; Chen, T.; Chang, S.-Y.; Lin, M.-S.; Yin, H.-S. Experimentally validated novel inhibitors of Helicobacter pylori phosphopantetheine adenylyltransferase discovered by virtual high-throughput screening. PLoS ONE 2013, 8, e74271. [Google Scholar] [CrossRef] [PubMed]
- Neelapu, N.R.R.; Mutha, N.V.R.; Akula, S. Identification of potential drug targets in Helicobacter pylori strain HPAG1 by in silico genome analysis. Infect. Disord. Drug Targets 2015, 15, 106–117. [Google Scholar] [CrossRef]
- Ge, Z. Potential of fumarate reductase as a novel therapeutic target in Helicobacter pylori infection. Expert Opin. Targets 2002, 6, 135–146. [Google Scholar] [CrossRef]
- Amundsen, S.K.; Spicer, T.; Karabulut, A.C.; Londoño, L.M.; Eberhardt, C.; Fernandez Vega, V.; Bannister, T.D.; Hodder, P.; Smith, G.R. Small-molecule inhibitors of bacterial AddAB and RecBCD Helicase-nuclease DNA repair enzymes. ACS Chem. Biol. 2012, 7, 879–891. [Google Scholar] [CrossRef]
- Shadrick, W.R.; Ndjomou, J.; Kolli, R.; Mukherjee, S.; Hanson, A.M.; Frick, D.N. Discovering new medicines targeting helicases: Challenges and recent progress. J. Biomol. Screen. 2013, 18, 761–781. [Google Scholar] [CrossRef]
- Freigang, J.; Diederichs, K.; Schäfer, K.P.; Welte, W.; Paul, R. Crystal structure of oxidized flavodoxin, an essential protein in Helicobacter pylori. Protein Sci. 2002, 11, 253–261. [Google Scholar] [CrossRef]
- Cremades, N.; Velazquez-Campoy, A.; Martinez-Julvez, M.; Neira, J.L.; Perez-Dorado, I.; Hermoso, J.; Jimenez, P.; Lanas, A.; Hoffman, P.S.; Sancho, J. Discovery of specific flavodoxin inhibitors as potential therapeutic agents against Helicobacter pylori infection. ACS Chem. Biol. 2009, 4, 928–938. [Google Scholar] [CrossRef]
- Galano, J.J.; Alías, M.; Pérez, R.; Velázquez-Campoy, A.; Hoffman, P.S.; Sancho, J. Improved flavodoxin inhibitors with potential therapeutic effects against Helicobacter pylori infection. J. Med. Chem. 2013, 56, 6248–6258. [Google Scholar] [CrossRef] [PubMed]
- Hughes, N.J.; Chalk, P.A.; Clayton, C.L.; Kelly, D.J. Identification of carboxylation enzymes and characterization of a novel four-subunit pyruvate: Flavodoxin oxidoreductase from Helicobacter pylori. J. Bacteriol. 1995, 177, 3953–3959. [Google Scholar] [CrossRef] [PubMed]
- Hughes, N.J.; Clayton, C.L.; Chalk, P.A.; Kelly, D.J. Helicobacter pylori porCDAB and oorDABC genes encode distinct pyruvate: Flavodoxin and 2-Oxoglutarate: Acceptor oxidoreductases which mediate electron transport to NADP. J. Bacteriol. 1998, 180, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Maurice, M.; Cremades, N.; Croxen, M.A.; Sisson, G.; Sancho, J.; Hoffman, P.S. Flavodoxin: Quinone reductase (FqrB): A redox partner of pyruvate: Ferredoxin oxidoreductase that reversibly couples pyruvate oxidation to NADPH production in Helicobacter pylori and Campylobacter jejuni. J. Bacteriol. 2007, 189, 4764–4773. [Google Scholar] [CrossRef]
- Mandal, R.S.; Das, S. In silico approach towards identification of potential inhibitors of Helicobacter pylori DapE. J. Biomol. Struct. Dyn. 2014, 33, 1460–1473. [Google Scholar] [CrossRef]
- Karita, M.; Etterbeek, M.L.; Forsyth, M.H.; Tummuru, M.K.R.; Blaser, M.J. Characterization of Helicobacter pylori dapE and Construction of a Conditionally Lethal dapE Mutant. Infect. Immun. 1997, 65, 4158–4164. [Google Scholar] [CrossRef]
- Basarab, G.S.; Hill, P.J.; Rastagar, A.; Webborn, P.J.H. Design of Helicobacter pylori glutamate racemase inhibitors as selective antibacterial agents: A novel pro-drug approach to increase exposure. Bioorg. Med. Chem. Lett. 2008, 18, 4716–4722. [Google Scholar] [CrossRef]
- Keating, T.A. Resistance mechanism to an uncompetitive inhibitor of a single-substrate, single-product enzyme: A study of Helicobacter pylori glutamate racemase. Future Med. Chem. 2013, 5, 1203–1214. [Google Scholar] [CrossRef]
- Chiu, H.-C.; Lin, T.-L.; Yang, J.-C.; Wang, J.-T. Synergistic effect of imp/ostA and msbA in hydrophobic drug resistance of Helicobacter pylori. BMC Microbiol. 2009, 9, 136. [Google Scholar] [CrossRef]
- Kwon, D.-H.; Woo, J.-S.; Perng, C.-L.; Go, M.F.; Graham, D.Y.; El-Zaatari, F.A.K. The effect of galE gene inactivation on lipopolysaccharide profile of Helicobacter pylori. Curr. Microbiol. 1998, 37, 144–148. [Google Scholar] [CrossRef]
- Loughlin, M.F. Novel therapeutic targets in Helicobacter pylori. Expert Opin. Targets 2003, 7, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, A.H.M.; Ernst, F.D.; Kusters, J.G. NikR-mediated regulation of Helicobacter pylori acid adaptation. Trends Microbiol. 2004, 12, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Chellia, R.; Hu, X.; Kathiresan, K.; Oh, D.-H.; Wang, M.-H. Eradication of Helicobacter pylori through the inhibition of urease and peptide deformylase: Computational and biological studies. Microb. Pathog. 2019, 128, 236–244. [Google Scholar] [CrossRef]
- Carlini, C.R.; Ligabue-Braun, R. Ureases as multifunctional toxic proteins: A review. Toxicon 2016, 110, 90–109. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-S.; Su, M.-M.; Zhang, X.-P.; Liu, Q.-X.; He, Z.-X.; Xu, C.; Zhu, H.-L. Developing potential Helicobacter pylori urease inhibitors from novel oxoindoline derivatives: Synthesis, biological evaluation and in silico study. Bioorg. Med. Chem. Lett. 2018, 28, 3182–3186. [Google Scholar] [CrossRef] [PubMed]
- Kafarski, P.; Talma, M. Recent advances in design of new urease inhibitors: A review. J. Adv. Res. 2018, 13, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Rowinska-Zyrek, M.; Witkowska, D.; Valensin, D.; Kamyszc, W.; Kozlowski, H. The C terminus of HspA—A potential target for native Ni(II) and Bi(III) anti-ulcer drugs. Dalton Trans. 2010, 39, 5814–5826. [Google Scholar] [CrossRef]
- Nammi, D.; Srimath-Tirumala-Peddinti, R.C.P.K.; Neelapu, N.R.R. Identification of Drug Targets in Helicobacter pylori by in silico Analysis: Possible Therapeutic Implications for Gastric cancer. Curr. Cancer Drug Targets 2016, 16, 79–98. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Cheng, T.; Xia, W.; Lai, Y.-T.; Sun, H. Nickel translocation between metallochaperones HypA and UreE in Helicobacter pylori. Metallomics 2014, 6, 1731–1736. [Google Scholar] [CrossRef]
- Tarsia, C.; Danielli, A.; Florini, F.; Cinelli, P.; Ciurli, S.; Zambelli, B. Targeting Helicobacter pylori urease activity and maturation: In-cell highthroughput approach for drug discovery. BBA Gen. Subj. 2018, 1862, 2245–2253. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Hu, H.Q.; Merrell, D.S.; Maroney, M.J. Dynamic HypA zinc site is essential for acid viability and proper urease maturation in Helicobacter pylori. Metallomics 2015, 7, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Rowinska-Zyrek, M.; Witkowska, D.; Bielinska, S.; Kamyszb, W.; Kozlowski, H. The -Cys–Cys- motif in Helicobacter pylori’s Hpn and HspA proteins is an essential anchoring site for metal ions. Dalton Trans. 2011, 40, 5604–5610. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.; Sun, X.; Gu, Q.; Watt, R.M.; Tanner, J.A.; Wong, B.C.Y.; Xia, H.H.; Huang, J.-D.; He, Q.-Y.; Sun, H. A proteomic approach for the identification of bismuth-binding proteins in Helicobacter pylori. J. Biol. Inorg. Chem. 2007, 12, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y.; Miftahussurur, M. Helicobacter pylori urease for diagnosis of Helicobacter pylori infection: A mini review. J. Adv. Res. 2018, 13, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, I.; Onishi, S.; Takeuchi, H.; Supuran, C.T. The α and β classes carbonic anhydrases from Helicobacter pylori as novel drug targets. Curr. Pharm. Des. 2008, 14, 622–630. [Google Scholar] [PubMed]
- Modakh, J.K.; Liu, Y.C.; Machuca, M.A.; Supuran, C.T.; Roujeinikova, A. Structural basis for the inhibition of Helicobacter pylori α-carbonic anhydrase by sulfonamides. PLoS ONE 2015, 10, e0127149. [Google Scholar] [CrossRef]
- Modak, J.K.; Liu, Y.C.; Supuran, C.T.; Roujeinikova, A. Structure-activity relationship for sulfonamide inhibition of Helicobacter pylori α-carbonic anhydrase. J. Med. Chem. 2016, 59, 11098–11109. [Google Scholar] [CrossRef]
- Marcus, E.A.; Moshfegh, A.P.; Sachs, G.; Scott, D.R. The Periplasmic α-carbonic anhydrase activity of Helicobacter pylori is essential for acid acclimation. J. Bacteriol. 2005, 187, 729–738. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. Bacterial, fungal and protozoan carbonic anhydrases as drug targets. Expert Opin. Targets 2015, 19, 1689–1704. [Google Scholar] [CrossRef]
- Nishimori, I.; Minakuch, T.; Kohsaki, T.; Onishi, S.; Takeuchi, H.; Vullo, D.; Scozzafavac, A.; Supuran, C.T. Carbonic anhydrase inhibitors: The β-carbonic anhydrase from Helicobacter pylori is a new target for sulfonamide and sulfamate inhibitors. Bioorg. Med. Chem. Lett. 2007, 17, 3585–3594. [Google Scholar] [CrossRef]
- Ménard, R.; Schoenhofen, I.C.; Tao, L.; Aubry, A.; Bouchard, P.; Reid, C.W.; Lachance, P.; Twine, S.M.; Fulton, K.M.; Cui, Q.; et al. Small-Molecule Inhibitors of the Pseudaminic Acid Biosynthetic Pathway: Targeting Motility as a Key Bacterial Virulence Factor. Antimicrob. Agents Chemother. 2014, 58, 7430–7440. [Google Scholar] [CrossRef] [PubMed]
- Schoenhofen, I.C.; Lunin, V.V.; Julien, J.-P.; Li, Y.; Ajamian, E.; Matte, A.; Cygler, M.; Brisson, J.-R.; Aubry, A.; Logan, S.M.; et al. Structural and functional characterization of PseC, an aminotransferase involved in the biosynthesis of pseudaminic acid, an essential flagellar modification in Helicobacter pylori. J. Biol. Chem. 2006, 281, 8907–8916. [Google Scholar] [CrossRef] [PubMed]
- Legrain, P.; Strosberg, D. Protein interaction domain mapping for the selection of validated targets and lead compounds in the anti-infectious area. Curr. Pharm. Des. 2002, 8, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Sheu, B.-S.; Yang, H.-B.; Yeh, Y.-C.; Wu, J.-J. Helicobacter pylori colonization of the human gastric epithelium: A bug’s first step is a novel target for us. J. Gastroenterol. Hepatol. 2010, 25, 26–32. [Google Scholar] [CrossRef]
- Pulic, I.; Loconte, V.; Zanotti, G. Structural Characterization at the atomic level of a molecular nano-machine: The state of the art of Helicobacter pylori flagellum organization. Am. J. Biochem. Biotechnol. 2014, 10, 143–161. [Google Scholar]
- Wu, J.-J.; Sheu, B.-S.; Huang, A.-H.; Lin, S.-T.; Yang, H.-B. Characterization of flgK gene and FlgK protein required for H pylori Colonization-from cloning to clinical relevance. World J. Gastroenterol. 2006, 12, 3989–3993. [Google Scholar] [CrossRef]
- Van Amsterdam, K.; Van Der Ende, A. Helicobacter pylori HP1034 (ylxH) is required for motility. Helicobacter 2004, 9, 387–395. [Google Scholar] [CrossRef]
- Ishijima, N.; Suzuki, M.; Ashida, H.; Ichikawa, Y.; Kanegae, Y.; Saito, I.; Borén, T.; Haas, R.; Sasakawa, C.; Mimuro, H. BabA-mediated adherence is a potentiator of the Helicobacter pylori Type IV secretion system activity. J. Biol. Chem. 2011, 286, 25256–25264. [Google Scholar] [CrossRef]
- Moonens, K.; Gideonsson, P.; Subedi, S.; Bugaytsova, J.; Romao, E.; Mendez, M.; Nordén, J.; Fallah, M.; Rakhimova, L.; Shevtsova, A.; et al. Structural insights into polymorphic ABO Glycan binding by Helicobacter pylori. Cell Host Microbe 2016, 19, 55–66. [Google Scholar] [CrossRef]
- Prinz, C.; Schoniger, M.; Rad, R.; Becker, I.; Keiditsch, E.; Wagenpfeil, S.; Classen, M.; Rosch, T.; Schepp, W.; Gerhard, M. Key importance of the Helicobacter pylori adherence factor blood group antigen binding adhesin during chronic gastric inflammation. Cancer Res. 2001, 61, 1903–1909. [Google Scholar]
- Tohidpour, A. CagA-mediated pathogenesis of Helicobacter pylori. Microb. Pathog. 2016, 93, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Javaheri, A.; Kruse, T.; Moonens, K.; Mejías-Luque, R.; Debraekeleer, A.; Asche, C.I.; Tegtmeyer, N.; Kalali, B.; Bach, N.C.; Sieber, S.A.; et al. Helicobacter pylori adhesin HopQ engages in a virulence-enhancing interaction with human CEACAMs. Nat. Microbiol. 2016, 2, 16189. [Google Scholar] [CrossRef] [PubMed]
- Tegtmeyer, N.; Moodley, Y.; Yamaoka, Y.; Pernitzsch, S.R.; Schmidt, V.; Rivas Traverso, F.; Schmidt, T.P.; Rad, R.; Yeoh, K.G.; Bow, H.; et al. Characterisation of worldwide Helicobacter pylori strains reveals genetic conservation and essentiality of serine protease HtrA. Mol. Microbiol. 2016, 99, 925–944. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Moss, S.F. Targeting Helicobacter pylori in gastric carcinogenesis. Expert Opin. Targets 2007, 11, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Hoy, B.; Lower, M.; Weydig, C.; Carra, G.; Tegtmeyer, N.; Geppert, T.; Schroder, P.; Sewald, N.; Backert, S.; Schneider, G.; et al. Helicobacter pylori HtrA is a new secreted virulence factor that cleaves E-cadherin to disrupt intercellular adhesion. EMBO Rep. 2010, 11, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, J.; Sondén, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Ångström, J.; Larsson, T.; Teneberg, S.; Karlsson, K.-A.; et al. Helicobacter pylori SabA Adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Stähler, F.N.; Odenbreit, S.; Haas, R.; Wilrich, J.; Van Vliet, A.H.M.; Kusters, J.G.; Kist, M.; Bereswill, S. The novel Helicobacter pylori CznABC metal efflux pump is required for cadmium, zinc, and nickel resistance, urease modulation, and gastric colonization. Infect. Immun. 2006, 74, 3845–3852. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, S.; Li, W.; Shan, Y.; Li, X.; Lu, X.; Li, Y.; Guo, Q.; Zhou, Y.; Jia, J. Proteomic analysis of the function of sigma factor σ54 in Helicobacter pylori survival with nutrition deficiency stress in vitro. PLoS ONE 2013, 8, e72920. [Google Scholar] [CrossRef]
- Dorer, M.S.; Sessler, T.H.; Salama, N.R. Recombination and DNA Repair in Helicobacter pylori. Annu. Rev. Microbiol. 2011, 65, 329–348. [Google Scholar] [CrossRef]
- Olekhnovich, I.N.; Vitko, S.; Valliere, M.; Hoffmana, P.S. Response to metronidazole and oxidative stress is mediated through homeostatic regulator HsrA (HP1043) in Helicobacter pylori. J. Bacteriol. 2014, 196, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Pelliciari, S.; Pinatel, E.; Vannini, A.; Peano, C.; Puccio, S.; De Bellis, G.; Danielli, A.; Scarlato, V.; Roncarati, D. Insight into the essential role of the Helicobacter pylori HP1043 orphan response regulator: Genome-wide identification and characterization of the DNA-binding sites. Sci. Rep. 2017, 7, 41063. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Salillas, S.; Velázquez-Campoy, A.; Espinosa Angarica, V.; Fillat, M.F.; Sancho, J.; Lanas, Á. Identifying potential novel drugs against Helicobacter pylori by targeting the essential response regulator HsrA. Sci. Rep. 2019, 9, 11294. [Google Scholar] [CrossRef]
- González, A.; Casado, J.; Chueca, E.; Salillas, S.; Velázquez-Campoy, A.; Angarica, V.E.; Bénejat, L.; Guignard, J.; Giese, A.; Sancho, J.; et al. Repurposing dihydropyridines for treatment of Helicobacter pylori infection. Pharmaceutics 2019, 11, 681. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, G.; Sanguinetti, M.; Gallo, A.; Posteraro, B. Review article: Biofilm formation by Helicobacter pylori as a target for eradication of resistant infection. Aliment. Pharm. 2012, 36, 222–230. [Google Scholar] [CrossRef]
- Huynh, H.Q.; Couper, R.T.L.; Tran, C.D.; Moore, L.; Kelso, R.; Butler, R.N. N-Acetylcysteine, a novel treatment for Helicobacter pylori infection. Dig. Dis. Sci. 2004, 49, 1853–1861. [Google Scholar] [CrossRef]
- Abut, E.; Yasar, B.; Güveli, H.; Bölükbas, C.; Bölükbas, F.F.; Dalay, A.R.; Kurdas, O.Ö. Effect of the mucolytic erdosteine on the success rate of PPI-based first-line triple therapy for Helicobacter pylori eradication: A prospective, double-blind, randomized, placebo-controlled study. Scand. J. Gastroenterol. 2010, 45, 677–683. [Google Scholar] [CrossRef]
- Backert, S.; Bernegger, S.; Skórko-Glonek, J.; Wessler, S. Extracellular HtrA serine proteases: An emerging new strategy in bacterial pathogenesis. Cell. Microbiol. 2018, 20, e12845. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.M.; Leite, M.; Seruca, R.; Figueiredo, C. Adherens junctions as targets of microorganisms: A focus on Helicobacter pylori. FEBS Lett. 2013, 587, 259–265. [Google Scholar] [CrossRef]
- Orengo, C.; Michie, A.; Jones, S.; Jones, D.; Swindells, M.; Thornton, J. CATH—A hierarchic classification of protein domain structures. Structure 1997, 5, 1093–1108. [Google Scholar] [CrossRef]
- Fillat, M.F.; Sandmann, G.; Gomez-Moreno, C. Flavodoxin from the nitrogen-fixing cyanobacterium Anabaena PCC 7119. Arch. Microbiol. 1988, 150, 160–164. [Google Scholar] [CrossRef]
- Sandmann, G.; Peleato, M.L.; Fillat, M.F.; Lázaro, M.C.; Gómez-Moreno, C. Consequences of the iron-dependent formation of ferredoxin and flavodoxin on photosynthesis and nitrogen fixation on Anabaena strains. Photosynth. Res. 1990, 26, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Ifuku, O.; Koga, N.; Haze, S.; Kishimoto, J.; Wachi, Y. Flavodoxin is required for conversion of dethiobiotin to biotin in Escherichia coli. Eur. J. Biochem. 1994, 224, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Vetter, H.; Knappe, J. Flavodoxin and ferredoxin of Escherichia coli. Hoppe Seylers Z. Physiol. Chem. 1971, 352, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Van Lin, B.; Bothe, H. Flavodoxin from Azotobacter vinelandii. Arch. Microbiol. 1972, 82, 155–172. [Google Scholar]
- Knauf, M.A.; Lohr, F.; Blumel, M.; Mayhew, S.G.; Ruterjans, H. NMR investigation of the solution conformation of oxidized flavodoxin from Desulfovibrio vulgaris. Determination of the tertiary structure and detection of protein-bound water molecules. Eur. J. Biochem. 1996, 238, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Rascio, N.; La Rocca, N. Biological Nitrogen Fixation. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar] [CrossRef]
- Simondsen, R.P.; Tollin, G. Structure-function relations in flavodoxins. Mol. Cell. Biochem. 1980, 33, 13–24. [Google Scholar] [CrossRef]
- Osborne, C.; Chen, L.-M.; Matthews, R.G. Isolation, cloning, mapping, and nucleotide sequencing of the gene encoding flavodoxin in Escherichia coli. J. Bacteriol. 1991, 173, 1729–1737. [Google Scholar] [CrossRef]
- Hoover, D.M.; Ludwig, M.L. A flavodoxin that is required for enzyme activation: The structure of oxidized flavodoxin from Escherichia coli at 1.8 A resolution. Protein Sci. 1997, 6, 2525–2537. [Google Scholar] [CrossRef]
- Sanyal, I.; Gibson, K.J.; Flint, D.H. Escherichia coli biotin synthase: An investigation into the factors required for its activity and its sulfur donor. Arch. Biochem. Biophys. 1996, 326, 48–56. [Google Scholar] [CrossRef]
- Bianchi, V.; Eliasson, R.; Fontecave, M.; Mulliez, E.; Hoover, D.M.; Matthews, R.G.; Reichard, P. Flavodoxin is required for the activation of the anaerobic ribonucleotide reductase. Biochem. Biophys. Res. Commun. 1993, 197, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Lawson, R.J.; Von Wachenfeldt, C.; Haq, I.; Perkins, J.; Munro, A.W. Expression and characterization of the two flavodoxin proteins of Bacillus subtilis, YkuN and YkuP: Biophysical properties and interactions with cytochrome P450 bioI. Biochemistry 2004, 43, 12390–12409. [Google Scholar] [CrossRef] [PubMed]
- Gangeswaran, R.; Eady, R.R. Flavodoxin 1 of Azotobacter vinelandii : Characterization and role in electron donation to purified assimilatory nitrate reductase. Biochem. J. 1996, 317, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Sawers, G.; Watson, G. A glycyl radical solution: Oxygen-dependent interconversion of pyruvate formate-lyase. Mol. Microbiol. 1998, 29, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.A.; Sancho, J. Native-Specific Stabilization of Flavodoxin by the FMN Cofactor: Structural and Thermodynamical Explanation. Proteins Struct. Funct. Bioinform. 2006, 63, 581–594. [Google Scholar] [CrossRef]
- Martıínez-Júlvez, M.; Cremades, N.; Bueno, M.; Pérez-Dorado, I.; Maya, C.; Cuesta-López, S.; Prada, D.; Falo, F.; Hermoso, J.A.; Sancho, J. Common conformational changes in flavodoxins induced by FMN and anion binding: The structure of Helicobacter pylori apoflavodoxin. Proteins 2007, 69, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cárdenas, Á.; Rojas, A.L.; Conde-Giménez, M.; Velázquez-Campoy, A.; Hurtado-Guerrero, R.; Sancho, J. Streptococcus pneumoniae TIGR4 flavodoxin: Structural and biophysical characterization of a novel drug target. PLoS ONE 2016, 11, e0161020. [Google Scholar] [CrossRef] [PubMed]
- López-Llano, J.; Maldonado, S.; Bueno, M.; Lostao, A.; Jiménez, M.Á.; Lillo, M.P.; Sancho, J. The long and short flavodoxins: I. The role of the differentiating loop in apoflavodoxin structure and FMN binding. J. Biol. Chem. 2004, 279, 47177–47183. [Google Scholar] [CrossRef]
- López-Llano, J.; Maldonado, S.; Jain, S.; Lostao, A.; Godoy-Ruiz, R.; Sanchez-Ruiz, J.M.; Cortijo, M.; Fernández-Recio, J.; Sancho, J. The long and short flavodoxins: II. The role of the differentiating loop in apoflavodoxin stability and folding mechanism. J. Biol. Chem. 2004, 279, 47184–47191. [Google Scholar] [CrossRef]
- Hall, D.A.; Kooi, C.W.V.; Stasik, C.N.; Stevens, S.Y.; Zuiderweg, E.R.P.; Matthews, R.G. Mapping the interactions between flavodoxin and its physiological partners flavodoxin reductase and cobalamin-dependent methionine synthase. Proc. Natl. Acad. Sci. USA 2001, 98, 9521–9526. [Google Scholar] [CrossRef]
- Alagaratnam, S.; Van Pouderoyen, G.; Pijning, T.; Dijkstra, B.W.; Cavazzini, D.; Rossi, G.L.; Van Dongen, W.M.A.M.; Van Mierlo, C.P.M.; Van Berkel, W.J.H.; Canters, G.W. A crystallographic study of Cys69Ala flavodoxin II from Azotobacter vinelandii: Structural determinants of redox potential. Protein Sci. 2005, 14, 2284–2295. [Google Scholar] [CrossRef]
- Kaihovaaraa, P.; Höök-Nikannea, J.; Uusi-Oukarib, M.; Kosunenc, T.U.; Salaspuro, M. Flavodoxin-dependent pyruvate oxidation, acetate production and metronidazole reduction by Helicobacter pylori. J. Antimicrob. Chemother. 1998, 41, 171–177. [Google Scholar] [CrossRef]
- Paul, R.; Bosch, F.U.; Schafer, K.P. Overexpression and Purification of Helicobacter pylori Flavodoxin and Induction of a Specific Antiserum in Rabbits. Protein Expr. Purif. 2001, 22, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Chen, L.; Yang, J.; Lin, J.; Chang, K.; Wang, J. Isolation of a Helicobacter pylori Protein, FldA, Associated With Mucosa-Associated Lymphoid Tissue Lymphoma of the Stomach. Gastroenterology 1999, 117, 82–88. [Google Scholar] [CrossRef]
- Gerrits, M.M.; Van Der Wouden, E.J.; Bax, D.A.; Van Zwet, A.A.; Van Vliet, A.H.M.; De Jong, A.; Kusters, J.G.; Thijs, J.C.; Kuipers, E.J. Role of the rdxA and frxA genes in oxygen-dependent metronidazole resistance of Helicobacter pylori. J. Med. Microbiol. 2004, 53, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Müller, M. Mode of action of metronidazole on anaerobic bacteria and protozoa. Surgery 1983, 93, 165–171. [Google Scholar]
- Kwon, D.H.; El-Zaatari, F.A.K.; Kato, M.; Osato, M.S.; Reddy, R.; Yamaoka, Y.; Graham, D.Y. Analysis of rdxA and involvement of additional genes encoding NAD(P)H flavin oxidoreductase (FrxA) and ferredoxin-like protein (FdxB) in metronidazole resistance of Helicobacter pylori. Antimicrob. Agents Chemother. 2000, 44, 2133–2142. [Google Scholar] [CrossRef]
- Kim, S.Y.; Joo, Y.M.; Lee, H.S.; Chung, I.S.; Yoo, Y.J.; Merrell, D.S.; Cha, J.H. Genetic analysis of Helicobacter pylori clinical isolates suggests resistance to metronidazole can occur without the loss of functional rdxA. J. Antibiot. (Tokyo) 2009, 62, 43–50. [Google Scholar] [CrossRef]
- Tomb, J.F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Dougherty, B.A.; et al. The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 1997, 389, 412. [Google Scholar] [CrossRef]
- Cremades, N.; Velazquez-Campoy, A.; Freire, E.; Sancho, J. The flavodoxin from Helicobacter pylori: Structural Determinants of Thermostability and FMN Cofactor Binding. Biochemistry 2008, 47, 627–639. [Google Scholar] [CrossRef][Green Version]
- Gray, L.R.; Tompkins, S.C.; Taylor, E.B. Regulation of pyruvate metabolism and human disease. Cell. Mol. Life Sci. 2014, 71, 2577–2604. [Google Scholar] [CrossRef]
- Hutcherson, J.A.; Sinclair, K.M.; Belvin, B.R.; Gui, Q.; Hoffman, P.S.; Lewis, J.P. Amixicile, a novel strategy for targeting oral anaerobic pathogens. Sci. Rep. 2017, 7, 10474. [Google Scholar] [CrossRef]
- Kletzin, A.; Adams, M.W.W. Molecular and phylogenetic characterization of pyruvate and 2-ketoisovalerate ferredoxin oxidoreductases from Pyrococcus furiosus and pyruvate ferredoxin oxidoreductase from Thermotoga maritima. J. Bacteriol. 1996, 178, 248–257. [Google Scholar] [CrossRef] [PubMed]
- UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef] [PubMed]
- Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018, 46, D8–D13. [CrossRef] [PubMed]
- Chazarreta-Cifre, L.; Martiarena, L.; de Mendoza, D.; Altabe, S.G. Role of ferredoxin and flavodoxins in Bacillus subtilis fatty acid desaturation. J. Bacteriol. 2011, 193, 4043–4048. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Lawson, R.J.; Buddha, M.R.; Wei, C.C.; Crane, B.R.; Munro, A.W.; Stuehr, D.J. Bacterial flavodoxins support nitric oxide production by Bacillus subtilis nitric-oxide synthase. J. Biol. Chem. 2007, 282, 2196–2202. [Google Scholar] [CrossRef]
- Yeom, J.; Jeon, C.O.; Madsen, E.L.; Park, W. In vitro and in vivo interactions of ferredoxin-NADP+ reductases in Pseudomonas putida. J. Biochem. 2009, 145, 481–491. [Google Scholar] [CrossRef]
- Yeom, J.-K.; Park, W.-J. Biochemical characterization of ferredoxin-NADP+ reductase interaction with flavodoxin in Pseudomonas putida. BMB Rep. 2012, 45, 476–481. [Google Scholar] [CrossRef]
- Yeom, J.; Park, W. Pleiotropic effects of the mioC mutation on the physiology of Pseudomonas aeruginosa PAO1. FEMS Microbiol. Lett. 2012, 335, 47–57. [Google Scholar] [CrossRef][Green Version]
- Moyano, A.J.; Tobares, R.A.; Rizzi, Y.S.; Krapp, A.R.; Mondotte, J.A.; Bocco, J.L.; Saleh, M.C.; Carrillo, N.; Smania, A.M. A Long-Chain Flavodoxin Protects Pseudomonas aeruginosa from Oxidative Stress and Host Bacterial Clearance. PLoS Genet. 2014, 10, e1004163. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Kapil, A.; Sharma, A.; Ragupathi, N.K.D.; Inbanathan, F.Y.; Veeraraghavan, B.; Kang, G. Whole-Genome Shotgun Sequencing of Cephalosporin-Resistant Salmonella enterica Serovar Typhi. Genome Announc. 2017, 5, e01639-16. [Google Scholar] [CrossRef] [PubMed]
- Kasai, S. Freshwater bioluminescence in Vibrio albensis (Vibrio cholerae biovar albensis) NCIMB 41 is caused by a two-nucleotide deletion in luxO. J. Biochem. 2006, 139, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.J.; Kelly, D.J. The Function, Biogenesis and Regulation of the Electron Transport Chains in Campylobacter jejuni: New Insights into the Bioenergetics of a Major Food-Borne Pathogen; Elsevier: Amsterdam, The Netherlands, 2019; Volume 74, ISBN 9780128177129. [Google Scholar]
- Weerakoon, D.R.; Olson, J.W. The Campylobacter jejuni NADH: Ubiquinone oxidoreductase (complex I) utilizes flavodoxin rather than NADH. J. Bacteriol. 2008, 190, 915–925. [Google Scholar] [CrossRef]
- Kendall, J.J.; Barrero-Tobon, A.M.; Hendrixson, D.R.; Kelly, D.J. Hemerythrins in the microaerophilic bacterium Campylobacter jejuni help protect key iron-sulphur cluster enzymes from oxidative damage. Environ. Microbiol. 2014, 16, 1105–1121. [Google Scholar] [CrossRef]
- Kolker, E.; Picone, A.F.; Galperin, M.Y.; Romine, M.F.; Higdon, R.; Makarova, K.S.; Kolker, N.; Anderson, G.A.; Qiu, X.; Auberry, K.J.; et al. Global profiling of Shewanella oneidensis MR-1: Expression of hypothetical genes and improved functional annotations. Proc. Natl. Acad. Sci. USA 2005, 102, 2099–2104. [Google Scholar] [CrossRef]
- Biel, S.; Klimmek, O.; Groß, R.; Kröger, A. Flavodoxin from Wolinella succinogenes. Arch. Microbiol. 1996, 166, 122–127. [Google Scholar] [CrossRef]
- Tanaka, M.; Haniu, M.; Yasunobu, K.T. Amind acid sequence of the Peptostreptococcus elsdenii flavodoxin. Biochem. Biophys. Res. Commun. 1971, 44, 886–892. [Google Scholar] [CrossRef]
- Mayhew, S.G.; Massey, V. Studies on the kinetics and mechanism of reduction of flavodoxin from Peptostreptococcus elsdenii by sodium dithionite. Biochim. Biophys. Acta 1973, 315, 181–190. [Google Scholar] [CrossRef][Green Version]
- Mayhew, S.G.; Massey, V. Purification and characterization of flavodoxin from Peptostreptococcus elsdenii. J. Biol. Chem. 1969, 244, 794–802. [Google Scholar] [PubMed]
- Fox, E.M.; Allnutt, T.; Bradbury, M.I.; Fanning, S.; Chandry, P.S. Comparative genomics of the Listeria monocytogenes ST204 subgroup. Front. Microbiol. 2016, 7, 2057. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Shang, J.; Feng, X.; Guo, X.; Zhang, L.; Zhou, Q. PrfA led to reduced biofilm formation and contributed to altered gene expression patterns in biofilm-forming Listeria monocytogenes. Curr. Microbiol. 2013, 67, 372–378. [Google Scholar] [CrossRef]
- Ofer, A.; Kreft, J.; Logan, D.T.; Cohen, G.; Borovok, I.; Aharonowitz, Y. Implications of the inability of Listeria monocytogenes EGD-e to grow anaerobically due to a deletion in the class III NrdD ribonucleotide reductase for its use as a model laboratory strain. J. Bacteriol. 2011, 193, 2931–2940. [Google Scholar] [CrossRef] [PubMed]
- Knight, E., Jr.; Hardy, R.W.F. Flavodoxin of Clostridium pasteurianum. Methods Enzymol. 1971, 18, 592–598. [Google Scholar]
- Plegaria, J.S.; Sutter, M.; Ferlez, B.; Aussignargues, C.; Niklas, J.; Poluektov, O.G.; Fromwiller, C.; Teravest, M.; Utschig, L.M.; Tiede, D.M.; et al. Structural and functional characterization of a short-chain flavodoxin associated with a noncanonical 1,2-propanediol utilization bacterial microcompartment. Biochemistry 2017, 56, 5679–5690. [Google Scholar] [CrossRef]
- Fox, J.L.; Smith, S.S.; Brown, J.R. Amino acid sequences of Clostridium pasteurianum flavodoxin. Z. Naturforsch. B 1972, 27, 1096–1100. [Google Scholar] [CrossRef]
- Luo, H.; Lin, Y.; Gao, F.; Zhang, C.-T.; Zhang, R. DEG 10, an update of the database of essential genes that includes both protein-coding genes and noncoding genomic elements. Nucleic Acids Res. 2014, 42, D574–D580. [Google Scholar] [CrossRef]
- Wilkes-Weiss, D. The Gram-staining properties of Treponema pallidum. Transl. Res. J. Lab. Clin. Med. 1924, 9, 716–718. [Google Scholar]
- Zhou, X.; Li, Y. Chapter 4—Subgingival microbes. In Atlas of Oral Microbiology: From Healthy Microflora to Disease; Elsevier: Amsterdam, The Netherlands, 2015; pp. 67–93. ISBN 978-0-12-802234-4. [Google Scholar]
- Krissinel, E. On the relationship between sequence and structure similarities in proteomics. Bioinformatics 2007, 23, 717–723. [Google Scholar] [CrossRef]
- Nardone, G.; Compare, D. The human gastric microbiota: Is it time to rethink the pathogenesis of stomach diseases? United Eur. Gastroenterol. J. 2015, 3, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Vasile Petra, C.; Rus, A.; Dumitraşcu, D.L. Gastric microbiota: Tracing the culprit. Clujul Med. 2017, 90, 369–376. [Google Scholar]
- Sebastián-Domingo, J.J.; Sánchez-Sánchez, C. From the intestinal flora to the microbiome. Rev. Esp. Enferm. Dig. 2018, 110, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, C.X.; Min, L.; Zhu, L.Y. Good or bad: Gut bacteria in human health and diseases. Biotechnol. Biotechnol. Equip. 2018, 32, 1075–1080. [Google Scholar] [CrossRef]
- Lagier, J.-C.; Million, M.; Hugon, P.; Armougom, F.; Raoult, D. Human gut microbiota: Repertoire and variations. Front. Cell. Infect. Microbiol. 2012, 2, 136. [Google Scholar] [CrossRef]
- Sohn, M.B.; An, L.; Pookhao, N.; Li, Q. Accurate genome relative abundance estimation for closely related species in a metagenomic sample. BMC Bioinform. 2014, 15, 242. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, W.; Schink, M.; Zopf, Y. Microbiota in the Gastrointestinal Tract. Med. Sci. 2018, 6, 116. [Google Scholar] [CrossRef]
- Gorbach, S.L. Chapter 95—Microbiology of the gastrointestinal tract. In Medical Microbiology, 4th ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- World Health Organization. Readings on Diarrhoea: Student Manual; WHO: Geneva, Switzerland, 1992; pp. 1–143. [Google Scholar]
- Ducarmon, Q.R.; Zwittink, R.D.; Hornung, B.V.H.; Van Schaik, W.; Young, V.B.; Kuijper, E.J. Gut microbiota and colonization resistance against bacterial enteric infection. Microbiol. Mol. Biol. Rev. 2019, 83, e00007–e00019. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins (Basel) 2017, 9, 60. [Google Scholar] [CrossRef]
- Chatterjee, R.; Shreenivas, M.M.; Sunil, R.; Chakravortty, D. Enteropathogens: Tuning their gene expression for Hassle-Free survival. Front. Microbiol. 2019, 9, 3303. [Google Scholar] [CrossRef]
- Hone, D.; Hackett, J. Vaccination against Enteric Bacterial Diseases. Rev. Infect. Dis. 1989, 11, 853–877. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M. Review of molecular subtyping methodologies used to investigate outbreaks due to multidrug-resistant enteric bacterial pathogens in sub-Saharan Africa. Afr. J. Lab. Med. 2019, 8, a760. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Mohakud, N.K.; Suar, M.; Sahu, B.R. Vaccine development for enteric bacterial pathogens: Where do we stand? Pathog. Dis. 2018, 76, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sistrunk, J.R.; Nickerson, K.P.; Chanin, R.B.; Rasko, D.A.; Faherty, C.S. Survival of the fittest: How bacterial pathogens utilize bile to enhance infection. Clin. Microbiol. Rev. 2016, 29, 819–836. [Google Scholar] [CrossRef]
- Wallace, N.; Zani, A.; Abrams, E.; Sun, Y. Chapter 4—The Impact of Oxygen on Bacterial Enteric Pathogens; Elsevier: Amsterdam, The Netherlands, 2016; Volume 95, ISBN 9780128048023. [Google Scholar]
- Riddle, M.S.; Chen, W.H.; Kirkwood, C.D.; MacLennan, C.A. Update on vaccines for enteric pathogens. Clin. Microbiol. Infect. 2018, 24, 1039–1045. [Google Scholar] [CrossRef]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar]
- Ishimaru, K.; Sasaki, M.; Narimatsu, H.; Arimizu, Y.; Gotoh, Y.; Nakamura, K.; Hayashi, T.; Ogura, Y. Escherichia coli O8 : H8 carrying a novel variant of the heat-labile enterotoxin LT2 gene caused outbreaks of diarrhea. Open Forum Infect. Dis. 2020, 7, ofaa021. [Google Scholar] [CrossRef]
- Fox, J.G.; Dewhirst, F.E.; Tully, J.G.; Paster, B.J.; Yan, L.; Taylor, N.S.; Collins, M.J.; Gorelick, P.L.; Ward, J.M. Helicobacter hepaticus sp. nov., a microaerophilic bacterium isolated from livers and intestinal mucosal scrapings from mice. J. Clin. Microbiol. 1994, 32, 1238–1245. [Google Scholar] [CrossRef]
- Falsafi, T.; Mahboubi, M. Helicobacter hepaticus, a new pathogenic species of the Helicobacter genus: Similarities and differences with H. Pylori. Iran. J. Microbiol. 2013, 5, 185–194. [Google Scholar]
- Dailidiene, D.; Dailide, G.; Ogura, K.; Zhang, M.; Mukhopadhyay, A.K.; Eaton, K.A.; Cattoli, G.; Kusters, J.G.; Berg, D.E. Helicobacter acinonychis: Genetic and rodent infection studies of a Helicobacter pylori-like gastric pathogen of cheetahs and other big cats. J. Bacteriol. 2004, 186, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Lash, R.H.; Lauwers, G.Y.; Odze, R.D.; Genta, R.M. Chapter 12—Inflammatory disorders of the stomach. In Surgical Pathology of the GI Tract, Liver, Biliary Tract, and Pancreas, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 269–320. [Google Scholar]
- Edwards, D.I. Nitroimidazole drugs—Action and resistance mechanisms. I. Mechanisms of action. J. Antimicrob. Chemother. 1993, 31, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.I. Nitroimidazole drugs—action and resistance mechanisms. II. Mechanisms of resistance. J. Antimicrob. Chemother. 1993, 31, 201–210. [Google Scholar] [CrossRef]
- Ang, C.W.; Jarrad, A.M.; Cooper, M.A.; Blaskovich, M.A.T. Nitroimidazoles: Molecular fireworks that combat a broad spectrum of infectious diseases. J. Med. Chem. 2017, 60, 7636–7657. [Google Scholar] [CrossRef]
- Edwards, D.I. Mechanisms of selective toxicity of metronidazole and other nitroimidazole drugs. Br. J. Vener. Dis. 1980, 56, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Sisson, G.; Goodwin, A.; Raudonikiene, A.; Hughes, N.J.; Mukhopadhyay, A.K.; Berg, D.E.; Hoffman, P.S. Enzymes associated with reductive activation and action of nitazoxanide, nitrofurans, and metronidazole in Helicobacter pylori. Antimicrob. Agents Chemother. 2002, 46, 2116–2123. [Google Scholar] [CrossRef]
- Salillas, S.; Alías, M.; Michel, V.; Mahía, A.; Lucía, A.; Rodrigues, L.; Bueno, J.; Galano-Frutos, J.J.; De Reuse, H.; Velázquez-Campoy, A.; et al. Design, synthesis, and efficacy testing of nitroethylene- and 7-nitrobenzoxadiazol-based flavodoxin inhibitors against Helicobacter pylori drug-resistant clinical strains and in Helicobacter pylori-infected mice. J. Med. Chem. 2019, 62, 6102–6115. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Pathway | Reference |
|---|---|---|
| Metabolism | ||
| Type II 3-dehydroquinase dehydratase (DHQ2) | Shikimate pathway | [30,31,32,33] |
| Shikimate 5-dehydrogenase (SDHase) | Shikimate pathway | [30,33] |
| Shikimate kinase (SK) | Shikimate pathway | [30,33,34] |
| Chorismate synthase | Shikimate pathway | [30,33] |
| Phosphopantetheine adenylyltransferase (PPAT) | Coenzyme A biosynthesis | [35,36] |
| Carbon starvation protein A | Starvation response, utilization of peptides, and host–pathogen interactions | [37] |
| Methylthiotransferase (MiaB) | Protein synthesis | [37] |
| Ribosomal RNA small subunit methyltransferase E | Protein synthesis | [37] |
| Ribosomal protein L11 methyltransferase | Protein synthesis | [37] |
| Tetrapyrrole (Corrin-Porphyrin) methylase family protein | Protein synthesis | [37] |
| Peptide chain release factor 1 | Protein synthesis | [37] |
| Fumarate reductase (FrdA, FrdB, and FrdC) | Krebs cycle and anaerobic respiration | [33,38] |
| Glu-tRNAGln amidotransferase, subunits A (GatA), B (GatB), and C (GatC) | Protein synthesis | [26,33] |
| Helicase-nuclease DNA Repair Enzymes (AddAB) | DNA damage reparation | [39,40] |
| Cytochrome C-type biogenesis protein CcdA | Cytochrome C synthesis | [37] |
| Cytochrome C oxidase, subunits CcoN, CcoO, CcoP and CcoQ | ATP synthesis | [37] |
| Flavodoxin (Fld) | Oxidative decarboxylation of pyruvate | [28,41,42,43] |
| Pyruvate:ferredoxin oxidoreductase (POR), subunit α (porA), β (porB), ϒ (porC or porG) and δ (porD) | Oxidative decarboxylation of pyruvate | [26,33,44,45,46] |
| Flavodoxin:quinone reductase (FqrB) | Oxidative decarboxylation of pyruvate | [33,42,46] |
| 2-oxoglutarate:acceptor oxidoreductase, subunits A (OorA), B (OorB), C (OorC) and D (OorD) | Succinyl-CoA production | [26,33,45] |
| Cell Wall Structure | ||
| N-succinyl-L,L-diaminopimelic acid desuccinylase, SDAP-deacylase (DapE) | Succinylase pathway (lysine biosynthesis) | [17,47,48] |
| Glutamate racemase MurI | Peptidoglycan biosynthesis | [49,50] |
| Multi-drug resistance protein MsbA | Lipopolysaccharide biosynthesis | [51] |
| UDP-galactose 4-epimerase (GalE) | Lipopolysaccharide biosynthesis | [52] |
| pH Homeostasis | ||
| Urease, subunits α (UreA), and β (UreB) | Acclimation to low pH | [33,53,54,55,56,57,58] |
| Nickel-responsive regulator (NikR) | Urease expression and nickel uptake regulator | [53,54] |
| Nickel–cobalt transporter (NixA) | Nickel/cobalt transport | [54,59,60] |
| Urease accessory protein UreE | Urease maturation | [33,54,59,61,62,63] |
| Urease accessory protein UreF | Urease maturation | [54,61,62] |
| Urease accessory protein UreG | Urease maturation | [33,54,62,63] |
| Urease accessory protein UreH | Urease maturation | [54,61] |
| Hydrogenase/urease maturation factor (HypA) | Urease maturation | [61,63] |
| Hydrogenase/urease maturation factor (HypB) | Urease maturation | [61,63] |
| Heat Shock Protein A (HspA) | Nickel homeostasis | [59,64,65] |
| Hpn | Nickel homeostasis and storage | [59,61,64] |
| Acid-activated urea channel (UreI) | Urea permeability | [33,66] |
| α-carbonic anhydrase | Acclimation to low pH | [33,67,68,69,70,71] |
| β-carbonic anhydrase | Acclimation to low pH | [33,67,68,69,71,72] |
| Virulence (Adherence, Motility and Pathogenicity) | ||
| Spore coat polysaccharide biosynthesis protein C (PseC) | Pseudaminic acid biosynthesis pathway (Pse): flagellin glycosylation | [33,73,74] |
| Heat-inducible transcription repressor (HrcA) | Flagella biosynthesis | [37] |
| Transcriptional repressor of DnaK operon (HspR) | Flagella biosynthesis | [37] |
| Major flagellin FlaA | Flagellar filament composition | [33,53,75,76,77] |
| Minor flagellin FlaB | Flagellar filament composition | [53,75,76,77] |
| Flagellar hook-associated protein 2 (FliD) | Flagellum assembly (filament capping) | [33,76,77] |
| Flagellar hook-associated protein 1 (FlgK) | Flagellum assembly (hook-filament junction formation) | [33,76,77,78] |
| ATP-binding protein (YlxH) | Flagella biosynthesis | [33,79] |
| Flagellar basal body L-ring protein (FlgH) | Flagellum assembly (L-ring composition) | [33,77] |
| Flagellar basal body P-ring protein (FlgI) | Flagellum assembly (P-ring composition) | [33,77] |
| Flagellar basal body M-ring protein (FliF) | Flagellum assembly (MS ring composition) | [33,77] |
| Flagellar biosynthetic protein (FliP) | Flagellum assembly (Flagellar export component) | [33,60,77] |
| Flagellar biosynthetic protein (FliQ) | Flagellum assembly (Flagellar export component) | [33,77] |
| Flagellar motor switch protein (FliY) | Flagellum assembly (C-ring composition; Flagellar export component) | [33,77] |
| Flagellum-specific ATP synthase (FliI) | Flagellum assembly (Flagellar export component) | [33,77] |
| Flagella-specific σ factor (FliA) | Flagellum assembly (regulatory protein) | [53,77] |
| FlgM (putative antagonist of FliA) | Flagellum assembly (regulatory protein) | [53,77] |
| Cytotoxin-associated gene A (CagA) | cag pathogenicity island (host cell metabolism modulation, inflammation, metaplasia and precancerous transformations) | [80,81,82,83] |
| cag-Type IV secretion system (T4SS) | cag pathogenicity island (translocation of bacterial factors (e.g., Cag A and peptidoglycan) into host cells) | [80,83] |
| HopQ adhesin (outer membrane protein) | Adhesion to host cells and translocation of CagA into host cells | [84] |
| Vacuolating cytotoxin (VacA) | Cellular vacuolation, apoptosis and inhibition of cell cycle progression and host immune response | [81,82,85,86] |
| Blood group antigen binding adhesin (BabA) | Adhesion to host cells | [80,81,82] |
| High temperature requirement A (HtrA) | Chaperone and proteolytic activities (intercellular adhesion cleavage) | [37,85,87] |
| Sialic acid-binding adhesin (SabA) (outer membrane protein) | Bacterial migration to epithelium surface | [76,88] |
| HopZ adhesin (outer membrane protein) | Adhesion to host cells | [76,80,85] |
| OipA adhesin (outer membrane protein) | Adhesion to host cells | [76,80,85] |
| AlpA/B adhesin (outer membrane protein) | Adhesion to host cells | [76,80,85] |
| Active Efflux of Metal Ions | ||
| Cation efflux system protein CusA | Efflux of cobalt/zinc/cadmium | [37] |
| Cobalt/Zinc/Cadmium efflux system membrane fusion protein | Efflux of cobalt/zinc/cadmium | [37] |
| Cobalt/Zinc/Cadmium resistance protein (CzcA, CzcB and CzcC) | Efflux of cobalt/zinc/cadmium | [37,89] |
| CznABC metal efflux pump | Efflux of cadmium/zinc/nickel | [89] |
| Ferrix siderophore transport system TonB periplasmic binding protein | Iron transport | [37] |
| Ferric siderophore transport system ExbB biopolymer transport protein | Iron transport | [37,60] |
| Haemin uptake system ATP binding protein | Iron transport | [37] |
| Protection Against Stress | ||
| Glutathionyl spermidine synthetase | Intracellular thiol redox balance regulation | [37] |
| Iron-binding ferritin-like antioxidant protein | Prevention of toxic reactive species formation | [37] |
| DNA-binding protein Dps | DNA breaking protection | [37] |
| Superoxide dismutase | Superoxide dismutation | [37] |
| Thioredoxin reductase | Prevention of toxic reactive species formation | [37] |
| RNA polymerase σ54 factor | Survival under stress conditions | [33,90] |
| Multi-drug resistance protein MsbA | Efflux of hydrophobic drugs | [33,51] |
| Exodeoxyribonuclease (LexA) | SOS response activation | [33,91] |
| Homeostatic stress regulator (HsrA) | Regulation of gene expression | [92,93,94,95] |
| Microorganism | Protein Name | Gene Name | Seq. Length | Long/short Chain | Phylum | Gram Stain |
|---|---|---|---|---|---|---|
| Anabaena (Nostoc) sp. | Flavodoxin | isiB | 170 | Long | Cyanobacteria | Negative |
| Aquifex aeolicus | Flavodoxin | fldA | 185 | Long | Aquificae | Negative |
| Azotobacter vinelandii | Flavodoxin 1 | Avin45950 | 174 | Long | Proteobacteria | Negative |
| Azotobacter vinelandii | Flavodoxin 2 | nifF | 180 | Long | Proteobacteria | Negative |
| Azotobacter chroococcum | Flavodoxin B | nifF | 180 | Long | Proteobacteria | Negative |
| Bacillus cereus | Flavodoxin | BC_1376 | 148 | Short | Firmicutes | Positive |
| Bacillus cereus | Flavodoxin | BC_3541 | 154 | Short | Firmicutes | Positive |
| Bacillus subtilis | Probable flavodoxin 2 | ykuP | 151 | Short | Firmicutes | Positive |
| Bacillus subtilis | Probable flavodoxin 1 | ykuN | 158 | Short | Firmicutes | Positive |
| Bacteroides uniformis | Flavodoxin | BACUNI_04544 | 178 | Long | Bacteroidetes | Negative |
| Buchnera aphidicola | Flavodoxin | fldA BUsg_289_ | 154 | Long b | Proteobacteria | Negative |
| Buchnera aphidicola | Flavodoxin | fldA BU299 | 171 | Long | Proteobacteria | Negative |
| Buchnera aphidicola | Flavodoxin | fldA bbp_277 | 174 | Long | Proteobacteria | Negative |
| Campylobacter jejuni | Flavodoxin | fldA | 163 | Long | Proteobacteria | Negative |
| Clostridium beijerinckiic | Flavodoxin | 138 | Short | Firmicutes | Positive | |
| Clostridium pasteurianum | Flavodoxin | CLPA_c13840d | 140 | Short | Firmicutes | Positive |
| Clostridium saccharobutylicum | Flavodoxin | floX | 160 | Long | Firmicutes | Positive |
| Desulfovibrio desulfuricans | Flavodoxin | Ddes_1951 | 148 | Short | Proteobacteria | Negative |
| Desulfovibrio gigasc | Flavodoxin | 146 | Short | Proteobacteria | Negative | |
| Desulfovibrio gigasc | Flavodoxin | 147 | Short | Proteobacteria | Negative | |
| Desulfovibrio salexigens | Flavodoxin | Desal_0805 | 146 | Short | Proteobacteria | Negative |
| Desulfovibrio vulgaris | Flavodoxin | DVU_2680/DvMF_1143 | 148 | Short | Proteobacteria | Negative |
| Escherichia coli | Protein MioC | mioC | 147 | Short | Proteobacteria | Negative |
| Escherichia coli | Uncharacterized protein YqcA | yqcA | 149 | Short | Proteobacteria | Negative |
| Escherichia coli | Flavodoxin 2 | fldB | 173 | Long | Proteobacteria | Negative |
| Escherichia coli | Flavodoxin 1 | fldA | 176 | Long | Proteobacteria | Negative |
| Fusobacterium nucleatum | Flavodoxin | FN0724 | 167 | Long | Fusobacteria | Negative |
| Haemophilus influenzae | Protein MioC homolog | mioC | 146 | Short | Proteobacteria | Negative |
| Haemophilus influenzae | Flavodoxin | fldA | 174 | Long | Proteobacteria | Negative |
| Helicobacter pylori | Flavodoxin | fldA | 164 | Long | Proteobacteria | Negative |
| Klebsiella pneumoniae | Flavodoxin | fldA/nifF | 176 | Long | Proteobacteria | Negative |
| Lactobacillus reuteri | Flavodoxin/nitric oxide synthase | Lreu_1727 | 149 | Short | Firmicutes | Positive |
| Listeria monocytogenes | Lmo2153 protein | lmo2153 | 145 | Short | Firmicutes | Positive |
| Megasphaera elsdenii(Peptostreptococcus elsdenii) c | Flavodoxin | 137 | Short | Firmicutes | Negative | |
| Pantoea agglomerans (Enterobacter agglomerans) | Flavodoxin | nifF | 177 | Long | Proteobacteria | Negative |
| Pasteurella multocida | Protein mioC homolog | mioC | 147 | Short | Proteobacteria | Negative |
| Pectobacterium carotovorumc | Exoenzyme regulation regulon ORF2 | 151 | Short | Proteobacteria | Negative | |
| Pseudomonas aeruginosa | Uncharacterized protein PA3435 | PA3435 | 150 | Short | Proteobacteria | Negative |
| Pseudomonas aeruginosa | Flavodoxin FldP | fldP | 184 | Long | Proteobacteria | Negative |
| Pseudomonas putida | Flavodoxin | mioC | 151 | Short | Proteobacteria | Negative |
| Rhodobacter capsulatus | Flavodoxin | nifF | 182 | Long | Proteobacteria | Negative |
| Salmonella Typhi | Flavodoxin 2 | fldB | 173 | Long | Proteobacteria | Negative |
| Salmonella Typhi | Flavodoxin | fldA | 176 | Long | Proteobacteria | Negative |
| Salmonella Typhimurium | Flavodoxin 2 | fldB | 173 | Long | Proteobacteria | Negative |
| Salmonella Typhimurium | Flavodoxin 1 | fldA | 176 | Long | Proteobacteria | Negative |
| Shewanella oneidensis | Flavodoxin Protein MioC | mioC | 146 | Short | Proteobacteria | Negative |
| Shewanella oneidensis | tRNA pseudouridine synthase C-associated flavoprotein YqcA | yqcA | 154 | Short | Proteobacteria | Negative |
| Shewanella oneidensis | Flavodoxin | fldA | 175 | Long | Proteobacteria | Negative |
| Shigella flexneri | Uncharacterized protein YqcA | yqcA | 149 | Short | Proteobacteria | Negative |
| Shigella flexneri | Flavodoxin 1 | fldA | 176 | Long | Proteobacteria | Negative |
| Streptococcus agalactiae e | Flavodoxin | mioC | 147 | Short | Firmicutes | Positive |
| Streptococcus pneumoniae | Flavodoxin | flaV | 147 | Short | Firmicutes | Positive |
| Synechococcus sp. | Flavodoxin | isiB | 170 | Long | Cyanobacteria | Negative |
| Synechocystis sp. | Flavodoxin | isiB | 170 | Long | Cyanobacteria | Negative |
| Treponema pallidum | Flavodoxin | fldA | 146 | Short | Spirochaetes | Negative f |
| Trichodesmium erythraeum | Flavodoxin | fld | 171 | Long | Cyanobacteria | Negative |
| Vibrio cholerae | Protein MioC homolog | mioC | 144 | Short | Proteobacteria | Negative |
| Vibrio cholerae | Flavodoxin | fld1 | 175 | Long | Proteobacteria | Negative |
| Vibrio cholerae | Flavodoxin | fld2 | 198 | Long | Proteobacteria | Negative |
| Wolinella succinogenes | Flavodoxin | fldA | 171 | Long | Proteobacteria | Negative |
| Microorganism | Sequence Length | Long/Short Chain | Phylum | Gram Stain |
|---|---|---|---|---|
| Campylobacter jejuni | 163 | Long | Proteobacteria | Negative |
| Escherichia coli | 176 | Long | Proteobacteria | Negative |
| Haemophilus influenzae | 174 | Long | Proteobacteria | Negative |
| Helicobacter pylori | 164 | Long | Proteobacteria | Negative |
| Salmonella Typhi | 176 | Long | Proteobacteria | Negative |
| Salmonella Typhimurium | 176 | Long | Proteobacteria | Negative |
| Shewanella oneidensis | 175 | Long | Proteobacteria | Negative |
| Streptococcus agalactiae | 147 | Short | Firmicutes | Positive |
| Vibrio cholerae | 175 | Long | Proteobacteria | Negative |
| Genus | Flavodoxin | Phylum | Gram Stain | Oxygen Requirement |
|---|---|---|---|---|
| Akkermansia | Unreviewed | Verrucomicrobia | Negative | Anaerobe |
| Alistipes | Unreviewed | Bacteroidetes | Negative | Anaerobe |
| Bacteriodes | Yes | Bacteroidetes | Negative | Anaerobe |
| Bifidobacterium | Unreviewed | Actinobacteria | Positive | Anaerobe |
| Clostridium | Yes | Firmicutes | Positive | Anaerobe |
| Eggerthella | Unreviewed | Actinobacteria | Positive | Anaerobe |
| Enterococcus | Unreviewed | Firmicutes | Positive | Facultative anaerobe |
| Escherichia | Yes b | Proteobacteria | Negative | Facultative anaerobe |
| Eubacterium | Unreviewed | Firmicutes | Positive | Anaerobe |
| Fusobacterium | Yes | Fusobacteria | Negative | Anaerobe |
| Haemophilus | Yes b | Proteobacteria | Negative | Facultative anaerobe |
| Lactobacillus | Yes | Firmicutes | Positive | Microaerophile |
| Neisseria | No | Proteobacteria | Negative | Aerobe |
| Odoribacter | Unreviewed | Bacteroidetes | Negative | Anaerobe |
| Parabacteroides | Unreviewed | Bacteroidetes | Negative | Anaerobe |
| Peptococcus | Unreviewed | Firmicutes | Positive | Anaerobe |
| Peptostreptococcus | Yes | Firmicutes | Positive | Anaerobe |
| Porphyromonas | Unreviewed | Bacteroidetes | Negative | Anaerobe |
| Prevotella | Unreviewed | Bacteroidetes | Negative | Anaerobe |
| Propionibacterium | Unreviewed | Actinobacteria | Positive | Anaerobe |
| Pseudomonas | Yes | Proteobacteria | Negative | Aerobe |
| Roseburia | Unreviewed | Firmicutes | Positive | Anaerobe |
| Rothia | Unreviewed | Actinobacteria | Positive | Anaerobe |
| Ruminococcus | Unreviewed | Firmicutes | Positive | Anaerobe |
| Staphylococcus | Unreviewed | Firmicutes | Positive | Facultative anaerobe |
| Streptococcus | Yes b | Firmicutes | Positive | Facultative anaerobe |
| Veillonella | Unreviewed | Firmicutes | Negative | Anaerobe |
| Genus | Flavodoxin | Phylum | Gram Stain | Oxygen Requirement |
|---|---|---|---|---|
| Bacillus | Yes | Firmicutes | Positive | Aerobe |
| Bacteroides | Yes | Bacteroidetes | Negative | Anaerobe |
| Campylobacter | Yes b | Proteobacteria | Negative | Microaerophile |
| Clostridium | Yes | Firmicutes | Positive | Anaerobe |
| Escherichia | Yes b | Proteobacteria | Negative | Facultative anaerobe |
| Helicobacter | Yes b | Proteobacteria | Negative | Microaerophile |
| Listeria | Yes | Firmicutes | Positive | Facultative anaerobe |
| Peptostreptococcus | Yes | Firmicutes | Positive | Anaerobe |
| Salmonella | Yes b | Proteobacteria | Negative | Facultative anaerobe |
| Shigella | Yes | Proteobacteria | Negative | Facultative anaerobe |
| Staphylococcus | Unreviewed | Firmicutes | Positive | Facultative anaerobe |
| Vibrio | Yes b | Proteobacteria | Negative | Facultative anaerobe |
| Yersinia | Unreviewed | Proteobacteria | Negative | Facultative anaerobe |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salillas, S.; Sancho, J. Flavodoxins as Novel Therapeutic Targets against Helicobacter pylori and Other Gastric Pathogens. Int. J. Mol. Sci. 2020, 21, 1881. https://doi.org/10.3390/ijms21051881
Salillas S, Sancho J. Flavodoxins as Novel Therapeutic Targets against Helicobacter pylori and Other Gastric Pathogens. International Journal of Molecular Sciences. 2020; 21(5):1881. https://doi.org/10.3390/ijms21051881
Chicago/Turabian StyleSalillas, Sandra, and Javier Sancho. 2020. "Flavodoxins as Novel Therapeutic Targets against Helicobacter pylori and Other Gastric Pathogens" International Journal of Molecular Sciences 21, no. 5: 1881. https://doi.org/10.3390/ijms21051881
APA StyleSalillas, S., & Sancho, J. (2020). Flavodoxins as Novel Therapeutic Targets against Helicobacter pylori and Other Gastric Pathogens. International Journal of Molecular Sciences, 21(5), 1881. https://doi.org/10.3390/ijms21051881