Genetic Factors of Nitric Oxide’s System in Psychoneurologic Disorders



Abstract
1. Introduction
2. Results
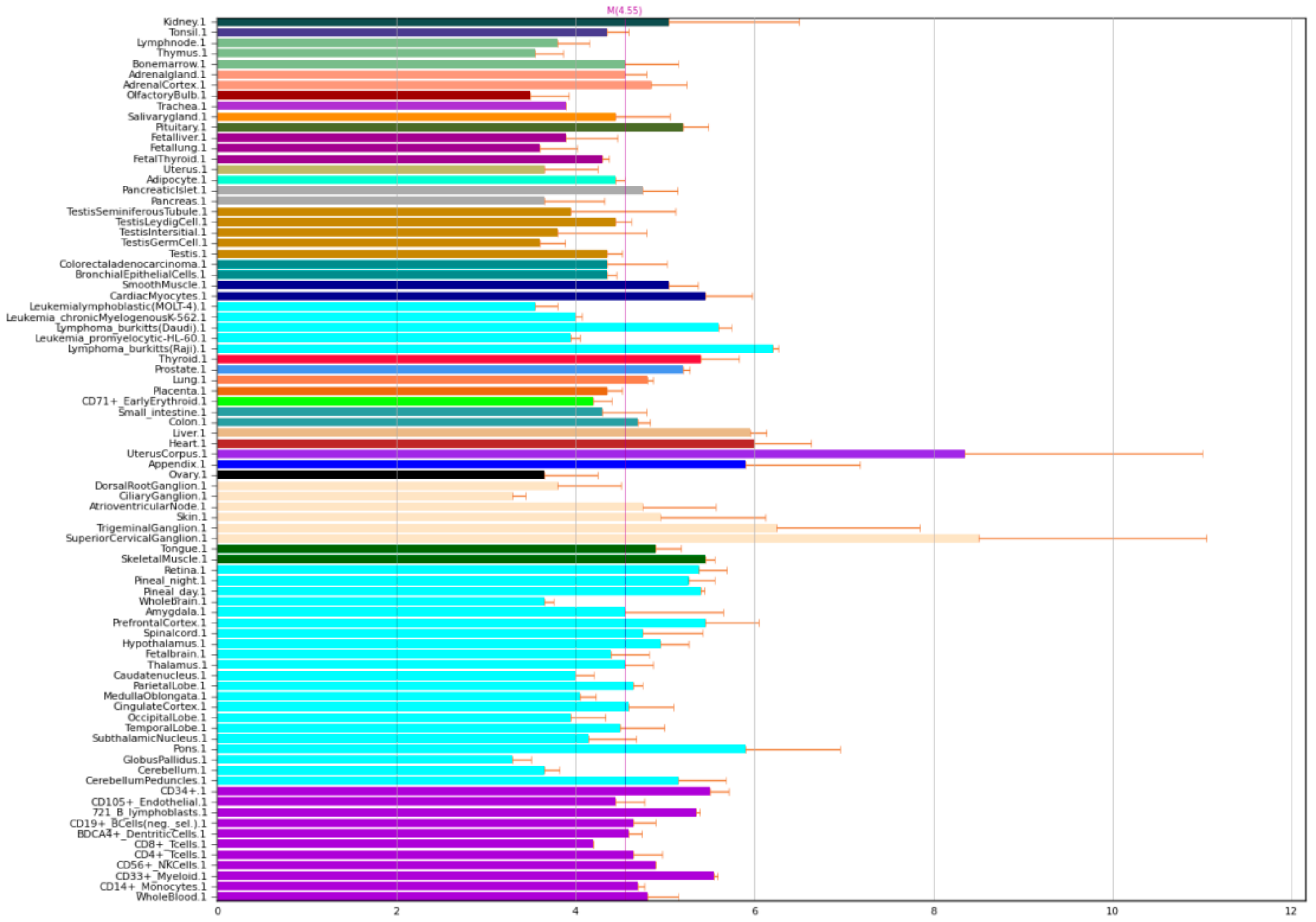
2.1. NOS1 Gene
2.1.1. Schizophrenia
2.1.2. Depression and Bipolar Affective Disorder
2.1.3. Autism Spectrum Disorders
2.1.4. Parkinson’s Disease
2.1.5. Brain Tumors
2.1.6. Cognitive Disorders
2.1.7. Ischemic Stroke
2.1.8. Restless Legs Syndrome
2.1.9. Multiple Sclerosis
2.2. NOS1AP Gene
2.2.1. Schizophrenia
2.2.2. Post-Traumatic Stress Disorder
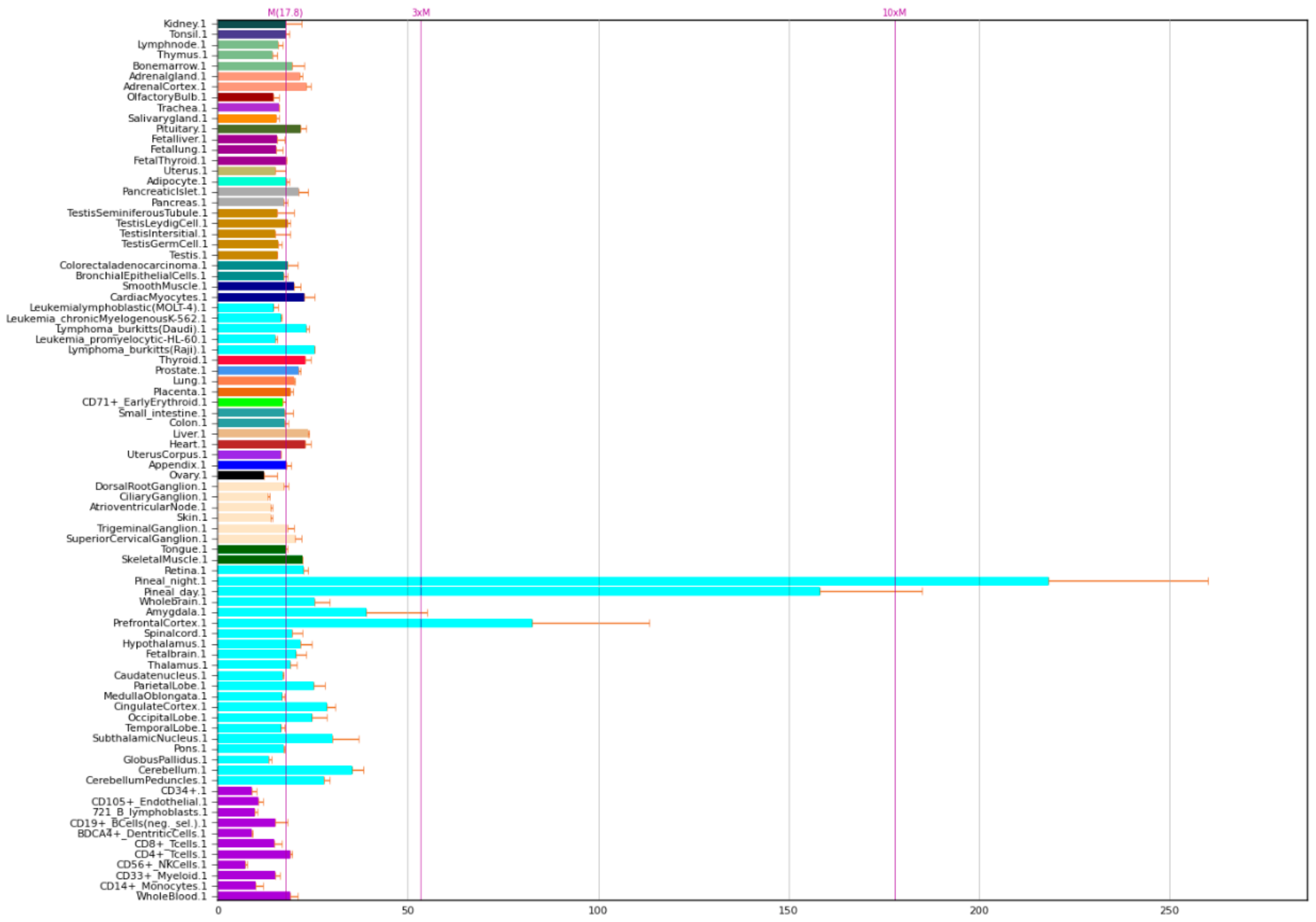
2.3. NOS2 Gene
2.3.1. Depression
2.3.2. Autism Spectrum Disorders
2.3.3. Parkinson’s Disease
2.3.4. Migraines
2.3.5. Brain Tumors
2.3.6. Ischemic Stroke
2.3.7. Multiple Sclerosis
2.4. NOS3 Gene
2.4.1. Methamphetamine-Induced Psychosis
2.4.2. Depression
2.4.3. Suicide
2.4.4. Parkinson’s Disease
2.4.5. Ischemic Stroke
2.4.6. Dementia
2.4.7. Migraines
2.4.8. Brain Tumors
2.4.9. Infantile Cerebral Palsy
2.4.10. Multiple Sclerosis
2.4.11. Gentamicin-Induced Vestibular Dysfunction
2.4.12. Hypoxic-Ischemic Encephalopathy (HIE)
2.4.13. Cerebral Ischemia Following Subarachnoid Hemorrhage
3. Conclusions
- nNOS are expressed mainly in central and peripheral neurons, as well as some other cell types, providing a synaptic plasticity in the central nervous system, central regulation of blood pressure, relaxation of smooth muscles, and vasodilation through peripheral nitrergic nerves.
- iNOS are expressed in many cells in response to lipopolysaccharides, cytokines, or other agents. iNOS generates a large amount of NO, which has a cytostatic effect on target cells of microorganisms, thereby underlining the pathophysiology of inflammatory diseases and septic shock, and are also involved in autoimmune reactions.
- eNOS are predominantly expressed in endothelial cells. They ensure the expansion of blood vessels, control blood pressure, and are responsible for other vasoprotective and anti-atherosclerotic effects. Many cardiovascular risk factors lead to oxidative stress and endothelial dysfunction in the vasculature.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| iNOS | Inducible nitric oxide synthase |
| nNOS | Neuronal nitric oxide synthase |
| eNOS | Endothelial nitric oxide synthase |
| SNV | Single nucleotide variant |
| BAD | Bipolar affective disorder |
| GWAS | Genome-wide association study |
| OR | Odds ratio |
| GDS | Geriatric Depression Scale |
| ASD | Autism spectrum disorders |
| PD | Parkinson’s disease |
| MMSE | Mini mental state examination |
| IQ | Intelligence quotient |
| EEG | Electroencephalogram |
| IS | Ischemic stroke |
| RLS | Restless legs syndrome |
| MS | Multiple sclerosis |
| NMDAR | N-methyl-d-aspartate-receptors |
| DNA | Deoxyribonucleic acid |
| NMDA | N-methyl-d-aspartate |
| PTSD | Post-traumatic stress disorder |
| BDI | Beck Depression Inventory |
| ROS | Reactive oxygen species |
| RNS | Reactive nitrogen species |
| 8-oxoG | 8-hydroxyguanine |
| 8-iso-PGF2a | 8-iso prostaglandin F2a |
| MDA | Malondialdehyde |
| CNS | Central nervous system |
| METH | Methamphetamine |
| METH-i.ps. | METH-induced psychosis |
| CAD | Coronary artery disease |
| VNTR | Variable number of tandem repeats |
| ICP | Infantile cerebral palsy |
| GM | Gentamicin |
| GM-i.v.d. | GM-induced vestibular dysfunction |
| HIE | Hypoxic-ischemic encephalopathy |
| aSAH | Aneurysmal subarachnoid hemorrhage |
References
- NCBI. Available online: https://www.ncbi.nlm.nih.gov (accessed on 1 February 2020).
- Geller, D.A.; Lowenstein, C.J.; Shapiro, R.A.; Nussler, A.K.; Di Silvio, M.; Wang, S.C.; Nakayama, D.K.; Simmons, R.L.; Snyder, S.H.; Billiar, T.R. Molecular cloning and expression of inducible nitric oxide synthase from human hepatocytes. Proc. Natl. Acad. Sci. USA 1993, 90, 3491–3495. [Google Scholar] [CrossRef] [PubMed]
- Hancock, D.B.; Martin, E.R.; Vance, J.M.; Scott, W.K. Nitric oxide synthase genes and their interactions with environmental factors in Parkinson’s disease. Neurogenetics 2008, 9, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Su, A.I.; Wiltshire, T.; Batalov, S.; Lapp, H.; Ching, K.A.; Block, D.; Zhang, J.; Soden, R.; Hayakawa, M.; Kreiman, G.; et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc. Natl. Acad. Sci. USA 2004, 101, 6062–6067. [Google Scholar] [CrossRef] [PubMed]
- BioGPS. Available online: https://www.BioGPS.org (accessed on 1 February 2020).
- Kishimoto, J.; Spurr, N.; Liao, M.; Lizhi, L.; Emson, P.; Xu, W. Localization of brain nitric oxide synthase (NOS) to human chromosome 12. Genomics 1992, 14, 802–804. [Google Scholar] [CrossRef]
- GeneCards. Available online: https://www.genecards.org (accessed on 1 February 2020).
- Shinkai, T.; Ohmori, O.; Hori, H.; Nakamura, J. Allelic association of the neuronal nitric oxide synthase (NOS1) gene with schizophrenia. Mol. Psychiatry 2002, 7, 560–563. [Google Scholar] [CrossRef][Green Version]
- Fallin, M.D.; Lasseter, V.K.; Avramopoulos, D.; Nicodemus, K.K.; Wolyniec, P.S.; McGrath, J.A.; Steel, G.; Nestadt, G.; Liang, K.Y.; Huganir, R.L.; et al. Bipolar I disorder and schizophrenia: A 440-single-nucleotide polymorphism screen of 64 candidate genes among Ashkenazi Jewish case-parent trios. Am. J. Hum. Genet. 2005, 77, 918–936. [Google Scholar] [CrossRef]
- Tang, W.; Huang, K.; Tang, R.; Zhou, G.; Fang, C.; Zhang, J.; Du, L.; Feng, G.; He, L.; Shi, Y. Evidence for association between the 5′ flank of the NOS1 gene and schizophrenia in the Chinese population. Int. J. Neuropsychopharmacol. 2008, 1, 1063–1071. [Google Scholar] [CrossRef]
- Reif, A.; Herterich, S.; Strobel, A.; Ehlis, A.C.; Saur, D.; Jacob, C.P.; Wienker, T.; Topner, T.; Fritzen, S.; Walter, U.; et al. A neuronal nitric oxide synthase (NOS-I) haplotype associated with schizophrenia modifies prefrontal cortex function. Mol. Psychiatry 2006, 11, 286–300. [Google Scholar] [CrossRef]
- Moskvina, V.; Craddock, N.; Holmans, P.; Nikolov, I.; Pahwa, J.S.; Green, E.; Wellcome Trust Case Control Consortium; Owen, M.J.; O’Donovan, M.C. Gene-wide analyses of genome-wide association data sets: Evidence for multiple common risk alleles for schizophrenia and bipolar disorder and for overlap in genetic risk. Mol. Psychiatry 2009, 14, 252–260. [Google Scholar] [CrossRef]
- Okumura, T.; Okochi, T.; Kishi, T.; Ikeda, M.; Kitajima, T.; Yamanouchi, Y.; Kinoshita, Y.; Kawashima, K.; Tsunoka, T.; Ujike, H.; et al. No association between polymorphisms of neuronal oxide synthase 1 gene (NOS1) and schizophrenia in a Japanese population. NeuromolecularMed. 2009, 11, 123–127. [Google Scholar] [CrossRef]
- Cui, H.; Nishiguchi, N.; Yanagi, M.; Fukutake, M.; Mouri, K.; Kitamura, N.; Hashimoto, T.; Shirakawa, O.; Hishimoto, A. A putative cis-acting polymorphism in the NOS1 gene is associated with schizophrenia and NOS1 immunoreactivity in the postmortem brain. Schizophr. Res. 2010, 121, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, X.H.; Xiang, B.; Wu, J.Y.; Wang, Y.C.; Deng, W.; Li, M.L.; Wang, Q.; He, Z.L.; Li, T. Association study of NOS1 gene polymorphisms and schizophrenia. Zhonghua Yi Xue Yi ChuanXueZaZhi 2012, 29, 459–463. (In Chinese) [Google Scholar] [CrossRef]
- Weber, H.; Klamer, D.; Freudenberg, F.; Kittel-Schneider, S.; Rivero, O.; Scholz, C.J.; Volkert, J.; Kopf, J.; Heupel, J.; Herterich, S.; et al. The genetic contribution of the NO system at the glutamatergic post-synapse to schizophrenia: Further evidence and meta-analysis. Eur. Neuropsychopharmacol. 2014, 24, 65–85. [Google Scholar] [CrossRef] [PubMed]
- Riley, B.; Thiselton, D.; Maher, B.S.; Bigdeli, T.; Wormley, B.; McMichael, G.O.; Fanous, A.H.; Vladimirov, V.; O’Neill, F.A.; Walsh, D.; et al. Replication of association between schizophrenia and ZNF804A in the Irish Case-Control Study of Schizophrenia sample. Mol. Psychiatry 2010, 15, 29–37. [Google Scholar] [CrossRef]
- Joca, S.R.; Guimarães, F.S.; Del-Bel, E. Inhibition of nitric oxide synthase increases synaptophysin mRNA expression in the hippocampal formation of rats. Neurosci. Lett. 2007, 421, 72–76. [Google Scholar] [CrossRef]
- Zhou, Q.G.; Zhu, L.J.; Chen, C.; Wu, H.Y.; Luo, C.X.; Chang, L.; Zhu, D.Y. Hippocampal neuronal nitric oxide synthase mediates the stress-related depressive behaviors of glucocorticoids by downregulating glucocorticoid receptor. J.Neurosci. 2011, 31, 7579–7590. [Google Scholar] [CrossRef]
- Sarginson, J.E.; Deakin, J.F.; Anderson, I.M.; Downey, D.; Thomas, E.; Elliott, R.; Juhasz, G. Neuronal nitric oxide synthase (NOS1) polymorphisms interact with financial hardship to affect depression risk. Neuropsychopharmacology 2014, 39, 2857–2866. [Google Scholar] [CrossRef]
- Montesanto, A.; Crocco, P.; Tallaro, F.; Pisani, F.; Mazzei, B.; Mari, V.; Corsonello, A.; Lattanzio, F.; Passarino, G.; Rose, G. Common polymorphisms in nitric oxide synthase (NOS) genes influence quality of aging and longevity in humans. Biogerontology 2013, 14, 177–186. [Google Scholar] [CrossRef]
- Wigner, P.; Czarny, P.; Synowiec, E.; Bijak, M.; Białek, K.; Talarowska, M.; Galecki, P.; Szemraj, J.; Sliwinski, T. Variation of genes involved in oxidative and nitrosative stresses in depression. Eur. Psychiatry 2018, 48, 38–48. [Google Scholar] [CrossRef]
- Bernstein, H.G.; Heinemann, A.; Krell, D.; Mawrin, C.; Bielau, H.; Danos, P.; Diekmann, S.; Keilhoff, G.; Bogerts, B.; Baumann, B. Further immunohistochemical evidence for impaired NO signaling in the hypothalamus of depressed patients. Ann. N. Y. Acad. Sci. 2002, 973, 91–93. [Google Scholar] [CrossRef]
- Savas, H.A.; Herken, H.; Yürekli, M.; Uz, E.; Tutkun, H.; Zoroğlu, S.S.; Ozen, M.E.; Cengiz, B.; Akyol, O. Possible role of nitric oxide and adrenomedullin in bipolar affective disorder. Neuropsychobiology 2002, 45, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Okumura, T.; Kishi, T.; Okochi, T.; Ikeda, M.; Kitajima, T.; Yamanouchi, Y.; Kinoshita, Y.; Kawashima, K.; Tsunoka, T.; Inada, T.; et al. Genetic association analysis of functional polymorphisms in neuronal nitric oxide synthase 1 gene (NOS1) and mood disorders and fluvoxamine response in major depressive disorder in the Japanese population. Neuropsychobiology 2010, 61, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Buttenschon, H.; Mors, O.; Ewald, H.; McQuillin, A.; Kalsi, G.; Lawrence, J.; Gurling, H.; Kruse, T. No association between a neuronal nitric oxide synthase (NOS1) gene polymorphism on chromosome 12q24 and bipolar disorder. Am. J. Med. Genet. 2004, 124B, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Cho, S.C.; Kim, J.W.; Cho, I.H.; Kim, S.A.; Park, M.; Cho, E.J.; Yoo, H.J. Family-based association study between NOS-I and -IIA polymorphisms and autism spectrum disorders in Korean trios. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2009, 150B, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, M.; Siddiqui, M.K. Low level lead exposure and oxidative stress: Current opinions. Clin.Chim.Acta 2007, 383, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Silbergeld, E.K. Facilitative mechanisms of lead as a carcinogen. MutatRes. 2003, 533, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, P.; Stewart, P.A.; Hutchinson, A.; Rothman, N.; Linet, M.S.; Inskip, P.D.; Rajaraman, P. Lead exposure, polymorphisms in genes related to oxidative stress, and risk of adult brain tumors. Cancer Epidemiol.Biomark. Prev. 2009, 18, 1841–1848. [Google Scholar] [CrossRef]
- Donohoe, G.; Walters, J.; Morris, D.W.; Quinn, E.M.; Judge, R.; Norton, N.; Giegling, I.; Hartmann, A.M.; Möller, H.J.; Muglia, P.; et al. Influence of NOS1 on verbal intelligence and working memory in both patients with schizophrenia and healthy control subjects. Arch. Gen. Psychiatry 2009, 66, 1045–1054. [Google Scholar] [CrossRef]
- O’Donoghue, T.; Morris, D.W.; Fahey, C.; Da Costa, A.; Foxe, J.J.; Hoerold, D.; Tropea, D.; Gill, M.; Corvin, A.; Donohoe, G. A NOS1 variant implicated in cognitive performance influences evoked neural responses during a high density EEG study of early visual perception. Hum. Brain Mapp. 2012, 33, 1202–1211. [Google Scholar] [CrossRef]
- Rose, E.J.; Greene, C.; Kelly, S.; Morris, D.W.; Robertson, I.H.; Fahey, C.; Jacobson, S.; O’Doherty, J.; Newell, F.N.; McGrath, J.; et al. The NOS1 variant rs6490121 is associated with variation in prefrontal function and grey matter density in healthy individuals. Neuroimage 2012, 60, 614–622. [Google Scholar] [CrossRef]
- Reif, A.; Schecklmann, M.; Eirich, E.; Jacob, C.P.; Jarczok, T.A.; Kittel-Schneider, S.; Lesch, K.P.; Fallgatter, A.J.; Ehlis, A.C. A functional promoter polymorphism of neuronal nitric oxide synthase moderates prefrontal functioning in schizophrenia. Int. J.Neuropsychopharmacol. 2011, 14, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, X.; Yu, P.; Zhang, Q.; Sun, X.; Gu, H.; Zhang, H.; Zhai, J.; Chen, M.; Du, B.; et al. Evidence for the contribution of NOS1 gene polymorphism (rs3782206) to prefrontal function in schizophrenia patients and healthy controls. Neuropsychopharmacology 2015, 40, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, R.; Sulman, E.P.; Scheurer, M.E.; Boehling, N.; Armstrong, G.N.; Tsavachidis, S.; Liang, F.W.; Etzel, C.J.; Conrad, C.A.; et al. Genetic Modulation of Neurocognitive Function in Glioma Patients. Clin. Cancer Res. 2015, 21, 3340–3346. [Google Scholar] [CrossRef] [PubMed]
- Toda, N.; Ayajiki, K.; Okamura, T. Cerebral blood flow regulation by nitric oxide: Recent advances. Pharmacol. Rev. 2009, 61, 62–97. [Google Scholar] [CrossRef]
- Manso, H.; Krug, T.; Sobral, J.; Albergaria, I.; Gaspar, G.; Ferro, J.M.; Oliveira, S.A.; Vicente, A.M. Variants within the nitric oxide synthase 1 gene are associated with stroke susceptibility. Atherosclerosis. 2012, 220, 443–448. [Google Scholar] [CrossRef]
- Dai, Y.; He, Z.; Sui, R.; Jiang, Z.; Ma, S. Association of nNOS gene polymorphism with ischemic stroke in Han Chinese of North China. Sci. World J. 2013, 3, 891581. [Google Scholar] [CrossRef]
- Winkelmann, J.; Lichtner, P.; Schormair, B.; Uhr, M.; Hauk, S.; Stiasny-Kolster, K.; Trenkwalder, C.; Paulus, W.; Peglau, I.; Eisensehr, I.; et al. Variants in the neuronal nitric oxide synthase (nNOS, NOS1) gene are associated with restless leg ssyndrome. Mov. Disord. 2008, 23, 350–358. [Google Scholar] [CrossRef]
- Al Fadhli, S.; Mohammed, E.M.; Al Shubaili, A. Association analysis of nitric oxide synthases: NOS1, NOS2A and NOS3 genes, with multiple sclerosis. Ann. Hum.Biol. 2013, 40, 368–375. [Google Scholar] [CrossRef]
- Brzustowicz, L.M.; Simone, J.; Mohseni, P.; Hayter, J.E.; Hodgkinson, K.A.; Chow, E.W.; Bassett, A.S. Linkage disequilibrium mapping of schizophrenia susceptibility to the CAPON region of chromosome 1q22. Am. J. Hum. Genet. 2004, 74, 1057–1063. [Google Scholar] [CrossRef][Green Version]
- Wratten, N.S.; Memoli, H.; Huang, Y.; Dulencin, A.M.; Matteson, P.G.; Cornacchia, M.A.; Azaro, M.A.; Messenger, J.; Hayter, J.E.; Bassett, A.S.; et al. Identification of a schizophrenia-associated functional noncoding variant in NOS1AP. Am. J. Psychiatry 2009, 166, 434–441. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, H.; Qin, W.; Chen, W.; Duan, Y.; Xiao, Y.; Li, C.; Zhang, J.; Li, X.; Feng, G.; et al. Association of the carboxyl-terminal PDZ ligand of neuronal nitric oxide synthase gene with schizophrenia in the Chinese Han population. Biochem. Biophys. Res.Commun. 2005, 328, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Lawford, B.R.; Morris, C.P.; Swagell, C.D.; Hughes, I.P.; Young, R.M.; Voisey, J. NOS1AP is associated with increased severity of PTSD and depression in untreated combat veterans. J. Affect. Disord. 2013, 147, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative andnitrosative stress (O&NS) pathways in major depression and their possiblecontribution to the (neuro)degenerative processes in that illness. Prog.Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar] [CrossRef] [PubMed]
- Stefanescu, C.; Ciobica, A. The relevance of oxidative stress status in first episode and recurrent depression. J. Affect.Disord. 2012, 143, 34–38. [Google Scholar] [CrossRef]
- Yager, S.; Forlenza, M.J.; Miller, G.E. Depression and oxidative damage to lipids. Psychoneuroendocrinology 2010, 35, 1356–1362. [Google Scholar] [CrossRef]
- Moylan, S.; Berk, M.; Dean, O.M.; Samuni, Y.; Williams, L.J.; O’Neil, A.; Hayley, A.C.; Pasco, J.A.; Anderson, G.; Jacka, F.N.; et al. Oxidative &nitrosative stress in depression: Why so much stress? Neurosci. Biobehav. Rev. 2014, 45, 46–62. [Google Scholar] [CrossRef]
- Leonard, B.; Maes, M. Mechanistic explanations how cell-mediated immuneactivation, inflammation and oxidative and nitrosative stress pathways and their sequels and concomitants play a role in the pathophysiology of unipolar depression. Neurosci. Biobehav. Rev. 2012, 36, 764–785. [Google Scholar] [CrossRef]
- Black, M.D.; Selk, D.E.; Hitchcock, J.M.; Wettstein, J.G.; Sorensen, S.M. On the effect of neonatal nitric oxide synthase inhibition in rats: A potential neurodevelopmental model of schizophrenia. Neuropharmacology 1999, 38, 1299–1306. [Google Scholar] [CrossRef]
- Herken, H.; Uz, E.; Ozyurt, H.; Akyol, O. Red blood cell nitric oxide levels in patients with schizophrenia. Schizophr. Res. 2001, 52, 289–290. [Google Scholar] [CrossRef]
- Pardo, C.A.; Vargas, D.L.; Zimmerman, A.W. Immunity, neuroglia and neuroinflammation in autism. Int. Rev. Psychiatry 2005, 17, 485–495. [Google Scholar] [CrossRef]
- De OS Mansur, T.; Gonçalves, F.M.; Martins-Oliveira, A.; Speciali, J.G.; Dach, F.; Lacchini, R.; Tanus-Santos, J.E. Inducible nitric oxide synthase haplotype associated with migraine and aura. Mol. Cell.Biochem. 2012, 364, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.M.; Luizon, M.R.; Speciali, J.G.; Martins-Oliveira, A.; Dach, F.; Tanus-Santos, J.E. Interaction among nitric oxide (NO)-related genes in migraine susceptibility. Mol. Cell.Biochem. 2012, 370, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.T.; Zhang, L.; Xu, Y.J.; Wang, X.J.; Wang, C.Y.; Wang, D.W. Polymorphisms of genes in nitric oxide-forming pathway associated with ischemic stroke in Chinese Han population. ActaPharmacol. Sin. 2011, 32, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cheng, S.; Brophy, V.H.; Erlich, H.A.; Mannhalter, C.; Berger, K.; Lalouschek, W.; Browner, W.S.; Shi, Y.; Ringelstein, E.B.; et al. RMS Stroke SNV Consortium. A meta-analysis of candidate gene polymorphisms and ischemic stroke in 6 study populations: Association of lymphotoxin-alpha in nonhypertensive patients. Stroke 2009, 40, 683–695. [Google Scholar] [CrossRef]
- Okochi, T.; Kishi, T.; Ikeda, M.; Kitajima, T.; Kinoshita, Y.; Kawashima, K.; Okumura, T.; Tsunoka, T.; Fukuo, Y.; Inada, T.; et al. Genetic Association Analysis of NOS3 and Methamphetamine-Induced Psychosis Among Japanese. Curr. Neuropharmacol. 2011, 9, 151–154. [Google Scholar] [CrossRef]
- Ikenouchi-Sugita, A.; Yoshimura, R.; Kishi, T.; Umene-Nakano, W.; Hori, H.; Hayashi, K.; Katsuki, A.; Ueda, N.; Iwata, N.; Nakamura, J. Three polymorphisms of thee NOS gene and plasma levels of metabolites of nitric oxide in depressed Japanese patients: A preliminary report. Hum.Psychopharmacol. 2011, 26, 531–534. [Google Scholar] [CrossRef]
- Sáiz, P.A.; García-Portilla, M.P.; Paredes, B.; Arango, C.; Morales, B.; Alvarez, V.; Coto, E.; Bascarán, T.; Bousoño, M.; Bobes, J. Lack of association between endothelial nitric oxide synthase (NOS3) gene polymorphisms and suicide attempts. Behav. Brain Funct. 2007, 3, 32. [Google Scholar] [CrossRef]
- Du, D.; Gao, P.; Hu, L.; Yang, Y.; Wang, F.; Ye, L.; Zhang, X.; Chang, M.; Zhao, J.; Wu, J.; et al. A genetic study of the NOS3 gene for ischemic stroke in a Chinese population. Int. J. Gen. Med. 2008, 1, 65–68. [Google Scholar] [CrossRef]
- Morris, C.M.; Ballard, C.G.; Allan, L.; Rowan, E.; Stephens, S.; Firbank, M.; Ford, G.A.; Kenny, R.A.; O’Brien, J.T.; Kalaria, R.N. NOS3 gene rs1799983 polymorphism and incident dementia in elderly stroke survivors. Neurobiol. Aging 2011, 32, 554.e1-6. [Google Scholar] [CrossRef]
- Schürks, M.; Kurth, T.; Buring, J.E.; Zee, R.Y. A candidate gene association study of 77 polymorphisms in migraine. J. Pain 2009, 10, 759–766. [Google Scholar] [CrossRef]
- Toriello, M.; Oterino, A.; Pascual, J.; Castillo, J.; Colás, R.; Alonso-Arranz, A.; Ruiz-Alegría, C.; Quintela, E.; Montón, F.; Ruiz-Lavilla, N. Lack of association of endothelial nitric oxide synthase polymorphisms and migraine. Headache 2008, 48, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.M.; Martins-Oliveira, A.; Speciali, J.G.; Luizon, M.R.; Izidoro-Toledo, T.C.; Silva, P.S.; Dach, F.; Tanus-Santos, J.E. Endothelial nitric oxide synthase haplotypes associated with aura in patients with migraine. DNA Cell Biol. 2011, 30, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zou, Y.F.; Xu, X.Y.; Feng, X.L.; Yang, L.; Zhang, G.C.; Bu, X.S.; Tang, J.L. The association of genetic polymorphisms with cerebral palsy: A meta-analysis. Dev. Med. Child Neurol. 2011, 53, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.M.; Williams, S.M.; Jiang, L.; Menon, K.S.; Jeka, J.J. Susceptibility genes forgentamicin-induced vestibular dysfunction. J.Vestib. Res. 2008, 18, 59–68. [Google Scholar] [PubMed]
- Wu, Y.; Zhu, Z.; Fang, X.; Yin, L.; Liu, Y.; Xu, S.; Li, A. The Association between NOS3 Gene Polymorphisms and Hypoxic-Ischemic Encephalopathy Susceptibility and Symptoms in Chinese Han Population. Biomed. Res. Int. 2016, 2016, 1957374. [Google Scholar] [CrossRef]
- KuzmanićSamija, R.; Primorac, D.; Resić, B.; Lozić, B.; Krzelj, V.; Tomasović, M.; Stoini, E.; Samanović, L.; Benzon, B.; Pehlić, M.; et al. Association of NOS3 tag polymorphisms with hypoxic-ischemic encephalopathy. Croat. Med. J. 2011, 52, 396–402. [Google Scholar] [CrossRef]
- Hendrix, P.; Foreman, P.M.; Harrigan, M.R.; Fisher, W.S., 3rd; Vyas, N.A.; Lipsky, R.H.; Lin, M.; Walters, B.C.; Tubbs, R.S.; Shoja, M.M.; et al. Endothelial Nitric Oxide Synthase Polymorphism Is Associated with Delayed Cerebral IschemiaH Following Aneurysmal Subarachnoid Hemorrhage. World Neurosurg. 2017, 101, 514–519. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNV | Psychiatric Disorders | Neurologic Disorders | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Schizophrenia | Depression | BAD | ASD | PD | Brain Tumors | Stroke | Cognitive Disorders | MS | RLS | |
| rs1004356 | + [20] | |||||||||
| rs1047735 | – [9,11] | – [9] | – [27] | + [3] | ||||||
| rs10507279 | + [20] | – [40] | ||||||||
| rs10774907 | – [40] | |||||||||
| rs10774910 | – [3] | |||||||||
| rs10850803 | – [30] | |||||||||
| rs10850807 | – [40] | |||||||||
| rs1093329 | – [40] | |||||||||
| rs1093330 | – [38] | |||||||||
| rs11068428 | – [38] | |||||||||
| rs11068438 | – [38] | |||||||||
| rs11068445 | – [38] | |||||||||
| rs11068446 | – [40] | |||||||||
| rs11068447 | + [3] | |||||||||
| rs1123425 | – [3] | – [30] | – [38] | – [40] | ||||||
| rs11611788 | – [3] | – [38] | + [36] | |||||||
| rs12578547 | – [3] | – [40] | ||||||||
| rs12829185 | + [3] | – [38] | ||||||||
| rs1353939 | – [30] | |||||||||
| rs1483757 | – [3] | + [38] – [39] | ||||||||
| rs1520810 | – [20] | – [38] | ||||||||
| rs1520811 | + [15] | – [3] | ||||||||
| rs1552227 | – [3] | – [38] | – [40] | |||||||
| rs1552228 | – [3] | |||||||||
| rs1607817 | – [38] | |||||||||
| rs17509231 | – [38] | |||||||||
| rs1875140 | – [20] | |||||||||
| rs1879417 | + [16] – [10] | – [21] – [22] | – [38] | – [21] | – [40] | |||||
| rs2133681 | – [11] | |||||||||
| rs2139733 | + [38] – [39] | |||||||||
| rs2291908 | – [30] | |||||||||
| rs2243044 | – [27] | |||||||||
| rs2293048 | – [40] | |||||||||
| rs2293050 | + [20] | + [38] – [39] | ||||||||
| rs2293051 | – [9] | – [9] | ||||||||
| rs2293052 | + [3] | |||||||||
| rs2293054 | – [11] | – [27] | – [3] | – [40] | ||||||
| rs2293055 | – [3] | – [40] | ||||||||
| rs2650163 | – [40] | |||||||||
| rs2682826 | + [8] - [9,10,13,14] | + [21] | - [9,26] | - [27] | + [3] | - [21] | - [41] | - [40] | ||
| rs3741473 | - [10] | |||||||||
| rs3741475 | - [27] | + [3] | - [38] | |||||||
| rs3741476 | -[27] | |||||||||
| rs3741480 | - [27] | |||||||||
| rs3782206 | + [10] – [9,13,14,16] | – [9] | + [35] | |||||||
| rs3782218 | + [20] | + [3] | – [38] | |||||||
| rs3782219 | + [9] – [10,13,14,16] | – [9] | ||||||||
| rs3782221 | + [9] – [10,13,14,16] | – [9] | – [3] | – [38] | ||||||
| rs3837437 | +* [10] | |||||||||
| rs41279104 | +[11,14,16] – [10,13] | – [20,25] | – [25] | + [34] | [41] | |||||
| rs473640 | – [40] | |||||||||
| rs4766836 | – [40] | |||||||||
| rs4767523 | – [3] | |||||||||
| rs4767533 | − [20] | |||||||||
| rs4767535 | − [30] | − [40] | ||||||||
| rs4767540 | − [10,16] | |||||||||
| rs478597 | − [3] | |||||||||
| rs483589 | − [30] | |||||||||
| rs499776 | +* [10] + [16] | |||||||||
| rs499813 | – [40] | |||||||||
| rs522910 | + [20] | |||||||||
| rs527590 | – [20] | – [38] | – [40] | |||||||
| rs530393 | – [40] | |||||||||
| rs532967 | – [9] | – [9] | ||||||||
| rs545654 | – [3] | – [30] | – [40] | |||||||
| rs547954 | – [3] | – [38] | ||||||||
| rs561712 | +* [10] – [9,13,14] | + [20] | – [9] | – [3] | ||||||
| rs576881 | – [38] | |||||||||
| rs579604 | – [20] | |||||||||
| rs6490121 | + [12] – [13,14,17] | – [30] | + [31,32,33] | – [40] | ||||||
| rs693534 | + [20] | – [3] | + [40] | |||||||
| rs7133438 | – [40] | |||||||||
| rs7139256 | – [3] | |||||||||
| rs7298903 | – [30] | – [38] | ||||||||
| rs7295972 | + [3] | |||||||||
| rs7308402 | + [38] – [39] | |||||||||
| rs7309163 | – [38] | |||||||||
| rs7314935 | – [38] | |||||||||
| rs7959232 | – [20] | |||||||||
| rs7977109 | – [38] | + [40] | ||||||||
| rs816292 | – [40] | |||||||||
| rs816293 | – [30] | – [38] | ||||||||
| rs816296 | – [20] | |||||||||
| rs816346 | – [38] | |||||||||
| rs816347 | – [40] | |||||||||
| rs816351 | – [30] | |||||||||
| rs816353 | – [38] | |||||||||
| rs816354 | – [3] | – [38] | ||||||||
| rs816357 | - [20] | - [38] | ||||||||
| rs816361 | - [38] | |||||||||
| rs877995 | - [40] | |||||||||
| rs904658 | - [38] | |||||||||
| rs9658247 | – [27] | |||||||||
| rs9658255 | – [27] | |||||||||
| rs9658266 | – [38] | |||||||||
| rs9658267 | – [38] | |||||||||
| rs9658281 | + [20] | – [38] | ||||||||
| rs9658536 | – [38] | |||||||||
| rs9658570 | – [40] | |||||||||
| SNV | Psychiatric Disorders | Neurologic Disorders | |||||
|---|---|---|---|---|---|---|---|
| Depression | ASD | PD | Migraines | IS | Brain Tumors | MS | |
| rs944725 | – [3] | + [30] | |||||
| rs10459953 | –* [22] | – [27] | |||||
| rs1060826 | + [27] | – [3] | |||||
| rs1137933 | – [27] | – [3] | – [57] | ||||
| rs2072324 | + [3] | ||||||
| rs2248814 | – [3] | ||||||
| rs2255929 | – [27] | + [3] | |||||
| rs2297515 | +* [22] | – [3] | |||||
| rs2297516 | + [3] | + [30] | |||||
| rs2297518 | – [22], +* [22] | – [27] | – [3] | + [54,55] | – [56] | ||
| rs2314810 | – [3] | ||||||
| rs2779248 | – [27] | ||||||
| rs2779249 | + [54], – [55] | ||||||
| rs2779252 | + [30] | ||||||
| rs3201742 | – [27] | ||||||
| rs3730014 | – [3] | ||||||
| rs3794764 | + [3] | ||||||
| rs3794766 | – [3] | ||||||
| rs4795067 | – [3] | + [30] | |||||
| rs7406657 | – [27] | ||||||
| rs8068149 | + [27] | ||||||
| rs8072199 | – [3] | + [30] | |||||
| rs10459953 | – [22], +* [22] | – [27] | |||||
| rs12944039 | + [3] | ||||||
| rs16966563 | – [3] | ||||||
| rs17722851 | – [3] | ||||||
| (CCTTT)n/(TAAA)n | + [41] | ||||||
| SNV | Psychiatric Disorders | Neurologic Disorders | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| METH-i.ps | Suicide | Depression | PD | Migraines | Brain tumors | Dementia | IS | ICP | MS | HIE | GM-i.v.d. | aSAH | |
| rs7830 | – [58] | ||||||||||||
| rs743506 | – [65] | ||||||||||||
| rs743507 | – [58] | ||||||||||||
| rs1549758 | – [3] | ||||||||||||
| rs1799983 | – [58] | – [59] | – [3] | – [63,64,65] | + [30] | + [62] | – [56,57] | – [66] | +* [69] | + [67] | |||
| rs1808593 | + [69] | ||||||||||||
| rs1800779 | – [58] | – [63,64] | – [57] | – [66] | + [41] | – [68] | |||||||
| rs1800783 | – [3] | – [38] | – [41] | – [68,69] +* [69] | |||||||||
| rs1808593 | – [3] | + [69] | |||||||||||
| rs2070744 | – [58] | – [60] | – [59] | – [65] | – [56] | – [41] | + [68] | – [70] | |||||
| rs2373929 | – [38] | ||||||||||||
| rs3918181 | + [61] | ||||||||||||
| rs3918186 | – [69] | ||||||||||||
| rs3918188 | – [58] | – [69] | |||||||||||
| rs3918226 | + [63] – [65] | – [57] | – [66] | ||||||||||
| rs3918227 | – [3] | – [69] | |||||||||||
| rs4496877 | + [30] | ||||||||||||
| rs10952298 | – [67] | ||||||||||||
| rs12703107 | + [30] | ||||||||||||
| 27bpVNTR | – [60] | – [59] | – [65] | + [41] | – [67] | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasyrova, R.F.; Moskaleva, P.V.; Vaiman, E.E.; Shnayder, N.A.; Blatt, N.L.; Rizvanov, A.A. Genetic Factors of Nitric Oxide’s System in Psychoneurologic Disorders. Int. J. Mol. Sci. 2020, 21, 1604. https://doi.org/10.3390/ijms21051604
Nasyrova RF, Moskaleva PV, Vaiman EE, Shnayder NA, Blatt NL, Rizvanov AA. Genetic Factors of Nitric Oxide’s System in Psychoneurologic Disorders. International Journal of Molecular Sciences. 2020; 21(5):1604. https://doi.org/10.3390/ijms21051604
Chicago/Turabian StyleNasyrova, Regina F., Polina V. Moskaleva, Elena E. Vaiman, Natalya A. Shnayder, Nataliya L. Blatt, and Albert A. Rizvanov. 2020. "Genetic Factors of Nitric Oxide’s System in Psychoneurologic Disorders" International Journal of Molecular Sciences 21, no. 5: 1604. https://doi.org/10.3390/ijms21051604
APA StyleNasyrova, R. F., Moskaleva, P. V., Vaiman, E. E., Shnayder, N. A., Blatt, N. L., & Rizvanov, A. A. (2020). Genetic Factors of Nitric Oxide’s System in Psychoneurologic Disorders. International Journal of Molecular Sciences, 21(5), 1604. https://doi.org/10.3390/ijms21051604