Abstract
Carotid paragangliomas (CPGLs) are rare neuroendocrine tumors often associated with mutations in SDHx genes. The immunohistochemistry of succinate dehydrogenase (SDH) subunits has been considered a useful instrument for the prediction of SDHx mutations in paragangliomas/pheochromocytomas. We compared the mutation status of SDHx genes with the immunohistochemical (IHC) staining of SDH subunits in CPGLs. To identify pathogenic/likely pathogenic variants in SDHx genes, exome sequencing data analysis among 42 CPGL patients was performed. IHC staining of SDH subunits was carried out for all CPGLs studied. We encountered SDHx variants in 38% (16/42) of the cases in SDHx genes. IHC showed negative (5/15) or weak diffuse (10/15) SDHB staining in most tumors with variants in any of SDHx (94%, 15/16). In SDHA-mutated CPGL, SDHA expression was completely absent and weak diffuse SDHB staining was detected. Positive immunoreactivity for all SDH subunits was found in one case with a variant in SDHD. Notably, CPGL samples without variants in SDHx also demonstrated negative (2/11) or weak diffuse (9/11) SDHB staining (42%, 11/26). Obtained results indicate that SDH immunohistochemistry does not fully reflect the presence of mutations in the genes; diagnostic effectiveness of this method was 71%. However, given the high sensitivity of SDHB immunohistochemistry, it could be used for initial identifications of patients potentially carrying SDHx mutations for recommendation of genetic testing.
1. Introduction
Carotid paraganglioma (CPGL) is a rare neuroendocrine tumor that arises from the carotid body. CPGL represents more than half of all head and neck (HN) paragangliomas (PGLs) [1]. According to the WHO Classification of Head and Neck Tumors 2017, PGLs were reclassified from indeterminate to malignant tumors with variable potential of metastasis [2]. As CPGL associates with the carotid arteries and adjacent nerves, its resection is challenging. Potential surgical complications include stroke, the possibility of cerebrovascular accident, and neural dysfunction, which can manifest clinically as hoarseness, vocal change, aspiration, dysphagia, dysarthria, facial asymmetry, or shoulder weakness [3]. Radiotherapy and chemotherapy can be used for unresectable and metastatic tumors, as well as for bilateral CPGLs; however, surgery remains the primary form of treatment [4,5,6].
CPGLs can develop both in the familial and sporadic forms [7]. Familial CPGLs occur as hereditary paraganglioma/pheochromocytoma (PGL/PCC) syndromes, including PGL1, PGL2, PGL3, PGL4, and PGL5, which are associated with germline mutations in the SDHD, SDHAF2, SDHC, SDHB, and SDHA genes, respectively [8]. SDHx genes encode for subunits of succinate dehydrogenase (SDH, mitochondrial complex II); SDHAF2 protein participates in the assembly of SDH and is required for SDHA flavination [9]. Two novel genes, TMEM127 and MAX, were recently found contributive to hereditary PGL/PCC syndromes. Germline pathogenic variants in these genes are mainly associated with PCCs but have also been seen in CPGLs [10,11]. Apparently sporadic cases of CPGLs have been reported as being associated with germline mutations in SDHA, SDHB, SDHD, and VHL, as well as somatic mutations in VHL, RET, IDH1, and IDH2 [12,13,14,15,16,17,18,19]. Germline pathogenic mutations in the FH and SLC25A11 genes were also determined in CPGLs [20,21].
The immunohistochemistry (IHC) of SDHB and SDHA subunits has been proposed as a useful method to predict underlying SDHx mutations in first-line diagnostics of hereditary PCCs/PGLs. All PGLs carrying SDHA mutations showed negative SDHA staining, while SDHA-positive immunoreactivity was observed in tumors with mutations in other SDHx genes [12]. Loss of cytoplasmic SDHB staining or a weak diffuse cytoplasmic blush instead of a normal granular staining pattern was revealed in PGLs with mutations in SDHB, SDHC, or SDHD [22,23]. Additionally, several cases of PGLs/PCCs with SDHA mutations showed a loss of SDHB protein expression coupled with negative SDHA staining [24]. To validate the reproducibility of SDHA/SDHB immunohistochemistry for the identification of hereditary PGLs/PCCs, seven expert endocrine pathologists performed analyses of 351 tumors with known/unknown genetics using a web-based virtual microscopy approach [24]. Pathologists reached an agreement in 99% (348/351) of the interpretations of SDHA immunohistochemistry and 90% (315/351) of the cases for SDHB. About 90% of PGLs/PCCs with mutations in SDHB, SDHC, SDHD, and SDHAF2 were SDHB immunonegative and SDHA immunopositive. Loss of SDHA/SDHB staining was detected in 75% of tumors with SDHA mutations. Finally, SDHA/SDHB protein expression was positive in 93% of non-SDHx-mutated cases. These results established the usability of SDHA/SDHB immunohistochemistry to identify SDHx mutations in PGLs/PCCs. Nevertheless, in some cases, the IHC results may be misinterpreted or reflect the wrong mutation status. For example, changed SDHB immunostaining can be caused by not only exonic mutations in the SDHx genes but also by the epigenetic alterations (epimutations), mutations in regulatory regions, changes in the expression and functionality of transcription factors, or other components of energy metabolism [23].
Another method for prediction of SDHx mutation status bases on evaluating the succinate-to-fumarate ratio (SFR) using liquid chromatography–mass spectrometry [18]. Recently, a comparative study between predictive capacities of SDHB immunohistochemistry and SFR methods has been reported [25]. In a general set of PGLs/PCCs, metabolite measurements showed higher specificity than SDHB immunohistochemical staining (99.2% versus 92.5%), but their sensitivity was comparable (88.1% versus 85.2%). However, the sensitivity of both methods was lower for HNPGLs than for other PGLs and PCCs. Thus, HNPGLs appear to be more difficult to diagnose using IHC and SFR methods, and need detailed study.
In this work, we first performed a correlation analysis between variants in the SDHx genes and their expression at the protein level in a representative set of CPGLs. The mutation status of the SDHx genes was determined using exome sequencing, and protein levels were estimated with IHC. We also evaluated the possibility of SDHA/SDHB immunostaining use to predict variants in any of the SDHx genes in CPGLs. We have found that SDHB staining does not always correlate with SDHx variants, while several studies proposed the immunohistochemistry of the SDHB subunit as a useful instrument for the prediction of SDHx mutation status. These important results indicate the necessity of genetic testing of SDHx variants along with IHC study in CPGLs.
2. Results
2.1. Pathogenic/Likely Pathogenic Variants of Susceptibility Genes in CPGLs
We used open-sources (ClinVar [26], dbSNP [27], and COSMIC [28]), prediction algorithms (SIFT [29], PolyPhen2 [30], MutationTaster [31], and LRT [32]), as well as position region conservation score (PhastCons [33] and PhyloP [34]) for interpretation of the variants, and analysis was only performed for variants with less than 1% population frequency. Pathogenicity of new variants was assessed using the criteria of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology (ACMG-AMP) [35]. Pathogenic/likely pathogenic variants in the SDHx genes were found in 16 out of 42 (38%) patients with CPGLs (Table 1, Figure 1). Among the 42 studied patients, ten had variants in the SDHD gene (24%), two were characterized with variants in SDHB (5%), three had variants in SDHC (7%), and one carried a variant in SDHA (2%).

Table 1.
Pathogenic/likely pathogenic variants in SDHx, RET, and IDH1 genes found in patients with carotid paragangliomas (CPGLs).
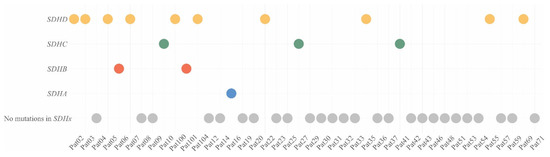
Figure 1.
Distribution of SDHx variants in patients with CPGLs. Yellow dots—patients with SDHD variants, green dots—patients with SDHC variants, red dots—patients with SDHB variants, blue dot—patient with SDHA variant, grey dots—patients with no mutations in SDHx.
In addition, we have analyzed the mutation status of RET, VHL, TMEM127, MAX, IDH1, IDH2, FH, and SLC25A11, as well as of genes belonging to the family of succinate dehydrogenase complex assembly factors (SDHAF1, SDHAF2, SDHAF3, and SDHAF4), which are the main susceptibility genes for CPGLs (Table 1). Pathogenic/likely pathogenic variants were found in RET and IDH1; no variants in VHL, TMEM127, MAX, IDH2, FH, SLC25A11, and SDHAF1-4 genes were identified.
Pathogenic variants in RET were observed in three (7%) patients with CPGLs. A pathogenic variant in RET NM_020975: c.2372A > T, p.(Tyr791Phe) (chr10: 43613908, rs77724903;13936) was revealed in two patients who also carried pathogenic/likely pathogenic variants in SDHA (Pat16) and SDHD (Pat35). In Pat27, we found a pathogenic variant in RET NM_020975: c.2944C > T, p.(Arg982Cys) (chr10: 43620335, rs17158558) that was corepresented with a likely pathogenic frameshift variant in the SDHC gene.
A likely pathogenic variant in IDH1 NM_005896: c.394C > T, p.(Arg132Cys) (chr2: 209113113, rs121913499) has been observed in one (2%) patient (Pat31) together with no mutations in the SDHx genes.
2.2. Correlation of SDHx Mutation Status with Their Immunostaining
Immunohistochemical staining was performed for SDHA, SDHB, SDHC, and SDHD subunits on 42 CPGL samples (Supplementary File S1). SDHB staining was assessed as follows: (+) Positive as granular cytoplasmic staining of tumor cells in parallel with the same intensity staining of internal positive control (endothelial cells); (-) negative as completely absent cytoplasmic staining together with staining of internal positive control; (*) weak diffuse as a cytoplasmic blush lacking definite granularity contrasting the strong granular staining of internal positive control (Figure 2). Immunostaining of SDHA, SDHC, and SDHD was scored as positive or negative in the same manner as SDHB.
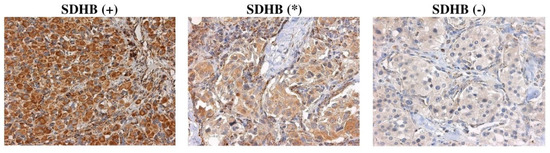
Figure 2.
Representative images of SDHB immunostaining in CPGLs. (+) positive, (*) weak diffuse, (-) negative. Magnification x400.
In the majority of SDHx-mutated tumors (94%, 15/16), we detected negative or weak diffuse staining of SDHB (Supplementary Table S1). The Spearman’s rank correlation coefficient (rs) between the presence of mutations in any SDHx genes and negative or weak diffuse SDHB staining was 0.51, p ≤ 0.05.
Negative or weak diffuse SDHB staining was found in nine out of ten cases with pathogenic/likely pathogenic SDHD variants; one SDHD-mutated tumor showed positive staining of all SDH subunits. In all the samples, SDHD was positively stained.
In two out of three samples with pathogenic/likely pathogenic variants in the SDHC gene, we identified weak diffuse SDHB staining and simultaneous positive SDHC expression. In one case, negative immunohistochemical staining for both SDHB and SDHC was found.
All samples with pathogenic/likely pathogenic variants in the SDHB gene showed negative SDHB staining.
A pathogenic/likely pathogenic variant in the SDHA gene was identified only in one patient. We observed both weak diffuse SDHB staining and negative SDHA expression in this sample.
Among twenty-six CPGLs with no pathogenic/likely pathogenic variants in the SDHx genes, negative or weak diffuse SDHB staining was observed in eleven cases (42%). Fifteen samples were immunopositive for all SDH subunits.
Three tumors with pathogenic variants of the RET gene, which were corepresented with SDHA, SDHC, and SDHD variants, have been characterized by negative or weak diffuse SDHB staining. Positive immunoreactivity was found in one patient with a likely pathogenic variant in the IDH1 gene occurring with no variant in SDHx.
2.3. Calculation of Diagnostic Accuracy
To determine the ability of SDHB immunohistochemistry discriminating SDHx-mutation carriers, we measured the sensitivity, specificity, and diagnostic effectiveness (accuracy) according to the following formulas:
where TP (true positives)—positively diagnosed subjects with the disease, FN (false negatives)—negatively diagnosed subjects with the disease, TN (true negatives)—negatively diagnosed subjects without the disease, and FP (false positives)—positively diagnosed subjects without the disease.
Sensitivity = TP/TP + FN,
Specificity = TN/TN + FP,
Accuracy = TN + TP/TN + TP + FN + FP,
In CPGLs, SDHB immunohistochemistry showed a sensitivity of 94% (15/16) and specificity of 58% (15/26). The diagnostic effectiveness of this method was 71% (30/42).
3. Discussion
Tumor cells are well-known to have alterations in energy metabolism that are exemplified by the Warburg effect [36,37]. A metabolic shift from mitochondrial respiration to glycolysis can be caused by mitochondrial dysfunction or by the reduction in its activity [38,39]. SDH has a critical role in mitochondrial metabolism; disruption of the SDH complex leads to abnormal accumulation of succinate in the cytosol, reprogramming of the energy metabolism, increased ROS production, stabilization of hypoxia-inducible factors (HIFs), and altered gene expression (in particular, for HIF targets) [40]. All of these changes can trigger neoplastic growth [38,41]. SDH abnormalities are associated with a tumorigenesis risk, including the development of PGLs/PCCs, renal and thyroid cancer, as well as composite PGLs/gastrointestinal stromal tumors (GISTs)/pulmonary chondromas (Carney triad) and PGLs/GISTs (Carney–Stratakis syndrome) [42].
SDHx are the most commonly mutated genes in PGLs/PCCs [43]. Variants in SDHD are more frequently observed in HNPGLs, followed by SDHB and SDHC mutations [44,45]. SDHA variants show extremely low penetrance in HNPGLs [10]. We obtained similar results; however, SDHC variants were found more often than SDHB variants. Previously, it has been reported that SDHC mutations are mainly associated with the development of CPGLs, explaining this difference [46].
Mutations in any of the SDHx genes can cause a destabilization of the SDH complex, loss of its enzymatic activity, and a disruption in the electron transport function [47,48,49,50]. Numerous studies have reported a changed expression pattern of SDHB presented as negative or weak diffuse immunostaining in tumors with SDHA-, SDHB-, SDHC-, and SDHD mutations [24,51,52,53]. It was shown that negative SDHB staining is more commonly associated with mutations in SDHB, whereas weak diffuse staining often occurs in SDHD-mutated tumors [52]. We also detected loss of SDHB expression in all patients with SDHB variants and weak diffuse SDHB staining in the majority of SDHD mutation carriers that support this finding. Studied patients carrying variants in SDHC showed both negative and weak diffuse SDHB staining. Notably, a number of authors interpreted SDHB staining only as positive or negative and considered a weak diffuse expression pattern as negative. Generally, both patterns indicate SDH deficiency, which is a surrogate marker for SDHx germline mutations almost always causing the gene biallelic inactivation [23]. Somatic events leading to biallelic inactivation have been rarely reported for the SDHx genes [23]. In the study, we used an archival collection of CPGLs for which paired normal tissues were unavailable; therefore, germline and somatic mutation status could not be estimated. However, based on this conception, we can suppose that in the majority of studied patients with SDH deficiency, the mutations of SDHx genes are germline. In one patient with a novel likely pathogenic frameshift SDHD variant, we found retention of SDHB expression. Possibly, this variant does not have a high impact on the protein structure or it occurs in one allele of the gene.
In a patient with a pathogenic SDHA variant, we have seen completely absent SDHA expression and weak diffuse SDHB immunostaining that is in accordance with the literature. Direct correlation with the presence of the gene mutation and loss of the protein expression is observed only for SDHA. Negative SDHA expression is defined both when mutation leads to the truncated protein and owing to missense mutation [12,54]. SDHB expression at the same time also becomes negative in SDHA-mutated tumors, supported by almost all reported cases (including our results) [12,24,54,55]. Moreover, SDHA mutation is a rare event in PGLs; therefore, the use of SDHB immunohistochemistry seems to be more expedient than SDHA/SDHB immunohistochemistry for prediction of mutations in any SDHx genes.
The loss of SDHC expression was revealed in one out of three patients with variants in the SDHC gene. This variant, NM_003001.3: c.224G > A, p.(Gly75Asp) (chr1: 161310428б rs786205147;189841), was described in the ClinVar database as a germline likely pathogenic variant associated with the hereditary cancer-predisposing syndrome and Carney triad with no experimental evidence of its pathogenicity to date. In this patient, negative SDHB staining was also determined. Therefore, we can suggest that, except for SDHA, no evident correlations have been found between negative SDHC and SDHD immunohistochemistry and the presence of pathogenic variants in the corresponding genes.
Among 42 patients with CPGLs, we revealed pathogenic RET variants in three cases and a likely pathogenic IDH1 variant in one patient. RET variants were presented in SDHx-mutated tumors that showed negative or weak diffuse SDHB staining. SDHx-mutations seem to be the main drivers of SDH efficiency; therefore, we cannot correctly assess the correlation of identified RET variants with the SDHB immunohistochemistry in these samples. The presence of the IDH1 variant was not associated with the changed immunostaining of any SDH subunits. However, more cases are needed to assess the impact of IDHx mutations on the stability of the SDH complex.
Despite the great results showing a high correlation of SDHB immunohistochemistry with the presence of SDHx variants (94%), negative or weak diffuse SDHB staining has also been found in 42% of tumors without pathogenic/likely pathogenic variants in any SDHx genes. In this case, SDH deficiency can be caused by mutations in the DNA regions, which have not been screened, or epimutations.
Given these data, we presumed that SDHB immunohistochemistry could be used for the initial assessment of SDHx variants in CPGLs with genetic testing in parallel. Additional SDHA staining increases the cost of the IHC analysis, but among PGLs/PCCs, SDHA mutation frequency is extremely low, and in the majority of such cases, negative or weak diffuse SDHB staining is also observed.
4. Materials and Methods
4.1. Tumor Samples and Patients
A total of 42 carotid paraganglioma samples (archive material) were used in this study. The formalin-fixed paraffin-embedded (FFPE) tumor tissues were collected in the Vishnevsky Institute of Surgery, Ministry of Health of the Russian Federation. Tumors were obtained from patients who did not receive radiotherapy or chemotherapy before surgery. Samples have no less than 80% of tumor cells. All the patients provided written informed consent. The study was approved by the ethics committee from the Vishnevsky Institute of Surgery with ethics committee approval no. 007/18, 02.10.2018 and performed according to the Declaration of Helsinki (1964). The clinicopathologic characteristics of the patients with CPGLs are presented in Table 2.
Table 2.
Clinicopathologic characteristics of patients with CPGLs.
4.2. DNA Extraction
DNA was extracted from tumor tissues using a High Pure FFPET DNA Isolation Kit (Roche, Basel, Switzerland) according to the manufacturer’s instructions. The quantification of isolated DNA was performed with a Qubit 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA). DNA quality was assessed by quantitative PCR (qPCR) using QuantumDNA Kit (Evrogen, Moscow, Russia).
4.3. Exome Sequencing
Exome libraries were prepared from DNA using a Rapid Capture Exome Kit (Illumina, San Diego, CA, USA) or TruSeq Exome Library Prep Kit (Illumina), according to the guidelines. Capture probes covered the same DNA regions in both kits (predominantly gene-coding regions). Library quantification was carried out using both Qubit 2.0 Fluorometer and qPCR. A quality assay of the libraries was performed on an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). High-throughput sequencing of the libraries was performed on a NextSeq 500 System (Illumina) in a paired-end mode of 76 × 2 bp. The average coverage for each sample was at least 300×. In this study, we used exome data of CPGL samples that were previously sequenced; raw sequence reads for Pat02–Pat51 are available at NCBI Sequence Read Archive (SRA) BioProject PRJNA411769 [56], and sequence data for Pat100–Pat104 are available at SRA BioProject PRJNA476932 [57]. Raw sequence data from an expanded set of CPGL samples (Pat53–Pat71) were added to the NCBI SRA BioProject PRJNA411769.
Bioinformatic analysis of exome sequencing data was carried out in the R environment. Raw reads were qualified using FASTQC (v. 0.11.9, Babraham Bioinformatics, Cambridge, UK). Quality trimming (less than Q20) and adapter removal were done with Trimmomatic (v. 0.39, USADEL LAB, Jülich, Germany) [58]. Then, reads were mapped to the reference human genome GRCh37.75/hg19 using BWA (v. 0.7.17, Wellcome Trust Genome Campus, Cambridge, UK) [59]. SAMtools (v. 1.10, Wellcome Trust Genome Campus, Cambridge, UK) [60] was used for BAM file sorting, and files were then processed with picard-tools (v. 2.23.4, Broad Institute, Cambridge, MA, USA). Base quality score recalibration was done with GATK4 (v. 4.1.2, Broad Institute, Cambridge, MA, USA) [61] and dbSNP (common variants 2015-06-05, Bethesda, MD, USA). Variant detection was carried out with GATK HaplotypeCaller [61]. We used GATK StrandBiasBySample, StrandOddsRatio, and BaseQualityRankSumTest to exclude false positives. Additionally, we excluded mis-sequenced single-nucleotide variants (SNVs) in polyN motifs, such as GGGTG >GGGGG, CCCCG >CCCCC, and others. Variants were annotated with Annovar (v. 20200316, Center for Applied Genomics, Philadelphia, PA, USA) [62]. VCF files included data on allele frequency (1000 Genomes Project [63], ExAC [64], gnomAD [65], Kaviar [66], and ESP-6500 [http://evs.gs.washington.edu/EVS/]), variant annotation (ClinVar, dbSNP, and COSMIC), position region conservation score (PhastCons and PhyloP [both PHAST v. 1.5, Siepel Lab, Cold Spring Harbor, NY, USA]), localization of variants in protein domains (InterPro [v. 81.0, Wellcome Genome Campus, Hinxton, Cambridgeshire, UK] [67]), as well as on pathogenicity prediction (SIFT [v. 6.2.1, Fred Hutchinson Cancer Research Center, Seattle, WA, USA], PolyPhen2 [v. 2.2.2, Harvard Medical School, Boston, MA, USA], MutationTaster [v. 2013-03-20, “Charité–Universitätsmedizin Berlin”, Berlin, Germany], and LRT [v. 0.2, Tel-Aviv University, Tel Aviv, Israel]).
4.4. Immunohistochemistry
IHC staining was used to analyze SDHx gene expression at the protein level. Sections (3–5 µm) from FFPE samples were prepared on glass slides using an HM 355S Automatic Microtome (Thermo Fisher Scientific) and then stained with hematoxylin-eosin (H&E) for histomorphological analysis. Deparaffinization of the sections was performed with xylene, with further rehydration in decreasing alcohol concentrations (absolute, 90%, 70%, and 50%) and washing in distilled water. Immunoreactions were performed in a serial manner using primary antibodies for all four SDH subunits (SDHA, monoclonal, clone 2E3GC12FB2AE2; SDHB, monoclonal, clone 21A11AE7; SDHC, monoclonal, clone EPR11035(B); SDHD, polyclonal) from Abcam (Cambridge, United Kingdom) on a Lab Vision Autostainer 360-2D (Thermo Fisher Scientific), according to the manufacturer’s instructions. Reactions continued in a ready-to-use visualization system Histofine DAB-2V (Nichirei Biosciences, Tokio, Japan) with universal chromogen-labeled (3,3′-diaminobenzidine, DAB) secondary antibodies. Additional Mayer’s hematoxylin staining was performed. Samples incubated without primary antibodies were used as the negative controls (Supplementary Figure S1). Granular cytoplasmic staining of SDH subunits in endothelial cells was used as a positive internal control. The slides were visualized using an Axio Imager 2 (Carl Zeiss Microscopy, Jena, Germany).
4.5. Correlation Analysis
Correlation analysis between SDHB staining and the presence of mutations in any SDHx genes was performed using the Spearman’s rank correlation test with STATISTICA 10 (StatSoft Inc., Tulsa, OK, USA).
5. Conclusions
This is the first study on the correlation between SDHx mutation status and their protein expression, respectively estimated with exome sequencing and IHC in a representative set of CPGLs. It has previously been reported that negative or weak diffuse SDHB staining has high sensitivity and specificity for the prediction of mutations of SDHx in PGLs/PCCs. However, our study showed that altered SDHB immunostaining widely occurs in tumors that do not carry pathogenic/likely pathogenic variants in the genes. These divergent results could be explained by the fact that earlier studies focused on PGLs/PCCs or all HNPGLs, but not only on CPGLs. Nevertheless, the sensitivity of the method remains high. Based on the collected data, we believe that SDHB immunohistochemistry could be used for primary identifications of patients potentially carrying SDHx variants who should be further referred for genetic testing.
Supplementary Materials
Can be found at https://www.mdpi.com/1422-0067/21/18/6950/s1.
Author Contributions
Conceptualization, A.V.K., A.V.S. and D.V.K.; Formal analysis, E.N.L., A.A.D. and G.S.K.; Investigation, V.S.P., M.S.F., E.A.P., M.V.S., O.A.S., T.B.D. and N.V.M.; Resources, A.L.G. and A.A.P.; Writing original draft, A.V.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work and publication costs were financially supported by grant 17-75-20105 from the Russian Science Foundation (collection of CPGL samples, DNA extraction, exome sequencing, IHC analysis) and grant 075-15-2019-1660 from the Ministry of Science and Higher Education of the Russian Federation (the data analysis of pathogenic mutations that will be further used in the development of paraganglioma animal models).
Acknowledgments
Authors thank the Vishnevsky Institute of Surgery for tissue samples, National Medical Research Center of Radiology and A. N. Severtsov Institute of Ecology and Evolution for assistance in the data analysis. This work was performed using the equipment of EIMB RAS “Genome” center (http://www.eimb.ru/ru1/ckp/ccu_genome_c.php).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| CPGL | carotid paraganglioma |
| SDHx | genes encoding for succinate dehydrogenase subunits |
| SDH | succinate dehydrogenase |
| IHC | immunohistochemistry |
| SFR | succinate to fumarate ratio |
References
- Prasad, S.C.; Paties, C.T.; Pantalone, M.R.; Mariani-Costantini, R.; Sanna, M. Carotid Body and Vagal Paragangliomas: Epidemiology, Genetics, Clinicopathological Features, Imaging, and Surgical Management. In Paraganglioma: A Multidisciplinary Approach; Codon Publications: Singapore, 2019; pp. 81–98. [Google Scholar]
- El-Naggar, A.K.; Chan, J.K.C.; Grandis, J.R.; Takata, T.; Slootweg, P.J. WHO Classification of Head and Neck Tumours; International Agency for Research on Cancer: Lyon, France, 2017. [Google Scholar]
- Myssiorek, D.; Persky, M.S.; Information, P.E.K.F.C. Treatment of carotid paraganglioma. Oper. Tech. Otolaryngol. Neck Surg. 2016, 27, 30–35. [Google Scholar] [CrossRef]
- Patel, S.R.; Winchester, D.J.; Benjamin, R.S. A 15-year experience with chemotherapy of patients with paraganglioma. Cancer 1995, 76, 1476–1480. [Google Scholar] [CrossRef]
- Lalya, I.; Mechchat, A.; Kebdani, T.; Hassouni, K.; Kanouni, L.; Andaloussi, K.; Elmarjani, M.; Hadadi, K.; Sifat, H.; Mansouri, H.; et al. Efficacité de la radiothérapie en première intention d’un paragangliome carotidien non résécable. J. des Mal. Vasc. 2011, 36, 185–188. [Google Scholar] [CrossRef]
- Kiziltan, H.S.; Ozucer, B.; Eris, A.H.; Veyseller, B. Bilateral Carotid Paraganglioma: Surgery and Radiotherapy. Clin. Med. Insights Case Rep. 2014, 7, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Boedeker, C.C. Paragangliomas and paraganglioma syndromes. GMS Curr. Top Otorhinolaryngol. Head Neck Surg. 2012, 10. [Google Scholar] [CrossRef]
- Udager, A.M.; Magers, M.J.; Goerke, D.M.; Vinco, M.L.; Siddiqui, J.; Cao, X.; Lucas, D.R.; Myers, J.L.; Chinnaiyan, A.M.; McHugh, J.B.; et al. The utility of SDHB and FH immunohistochemistry in patients evaluated for hereditary paraganglioma-pheochromocytoma syndromes. Hum. Pathol. 2018, 71, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.-X.; Khalimonchuk, O.; Schraders, M.; Dephoure, N.; Bayley, J.-P.; Kunst, H.; Devilee, P.; Cremers, C.W.R.J.; Schiffman, J.D.; Bentz, B.G.; et al. SDH5, a Gene Required for Flavination of Succinate Dehydrogenase, Is Mutated in Paraganglioma. Science 2009, 325, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Bausch, B.; Schiavi, F.; Ni, Y.; Welander, J.; Patocs, A.; Ngeow, J.; Wellner, U.; Malinoc, A.; Taschin, E.; Barbon, G.; et al. Clinical Characterization of the Pheochromocytoma and Paraganglioma Susceptibility Genes SDHA, TMEM127, MAX, and SDHAF2 for Gene-Informed Prevention. JAMA Oncol. 2017, 3, 1204–1212. [Google Scholar] [CrossRef]
- Burnichon, N.; Cascón, A.; Schiavi, F.; Morales, N.P.; Comino-Méndez, I.; Abermil, N.; Inglada-Pérez, L.; De Cubas, A.A.; Amar, L.; Barontini, M.; et al. MAX Mutations Cause Hereditary and Sporadic Pheochromocytoma and Paraganglioma. Clin. Cancer Res. 2012, 18, 2828–2837. [Google Scholar] [CrossRef]
- Korpershoek, E.; Favier, J.; Gaal, J.; Burnichon, N.; Van Gessel, B.; Oudijk, L.; Badoual, C.; Gadessaud, N.; Venisse, A.; Bayley, J.-P.; et al. SDHA Immunohistochemistry Detects Germline SDHA Gene Mutations in Apparently Sporadic Paragangliomas and Pheochromocytomas. J. Clin. Endocrinol. Metab. 2011, 96, 1472–1476. [Google Scholar] [CrossRef] [PubMed]
- Schiavi, F.; Savvoukidis, T.; Trabalzini, F.; Grego, F.; Piazza, M.; Amistà, P.; Demattè, S.; Piano, A.D.; Cecchini, M.E.; Erlic, Z.; et al. Paraganglioma Syndrome: SDHB, SDHC, and SDHD Mutations in Head and Neck Paragangliomas. Ann. N. Y. Acad. Sci. 2006, 1073, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Hermsen, M.; Sevilla, M.A.; Llorente, J.L.; Weiss, M.M.; Grimbergen, A.; Allonca, E.; Garcia-Inclán, C.; Balbín, M.; Suárez, C. Relevance of Germline Mutation Screening in Both Familial and Sporadic Head and Neck Paraganglioma for Early Diagnosis and Clinical Management. Cell. Oncol. 2010, 32, 275–283. [Google Scholar] [PubMed]
- Ding, Y.; Feng, Y.; Wells, M.; Huang, Z.; Chen, X. SDHx gene detection and clinical Phenotypic analysis of multiple paraganglioma in the head and neck. Laryngoscope 2018, 129, E67–E71. [Google Scholar] [CrossRef] [PubMed]
- Gaal, J.; Burnichon, N.; Korpershoek, E.; Roncelin, I.; Bertherat, J.; Plouin, P.-F.; De Krijger, R.R.; Gimenez-Roqueplo, A.-P.; Dinjens, W.N.M. Isocitrate Dehydrogenase Mutations Are Rare in Pheochromocytomas and Paragangliomas. J. Clin. Endocrinol. Metab. 2010, 95, 1274–1278. [Google Scholar] [CrossRef]
- Ercolino, T.; Becherini, L.; Valeri, A.; Maiello, M.; Gaglianò, M.S.; Parenti, G.; Ramazzotti, M.; Piscitelli, E.; Simi, L.; Pinzani, P.; et al. Uncommon clinical presentations of pheochromocytoma and paraganglioma in two different patients affected by two distinct novel VHL germline mutations. Clin. Endocrinol. 2008, 68, 762–768. [Google Scholar] [CrossRef]
- Richter, S.; Gieldon, L.; Pang, Y.; Peitzsch, M.; Huynh, T.; Leton, R.; Viana, B.; Ercolino, T.; Mangelis, A.; Rapizzi, E.; et al. Metabolome-guided genomics to identify pathogenic variants in isocitrate dehydrogenase, fumarate hydratase, and succinate dehydrogenase genes in pheochromocytoma and paraganglioma. Genet. Med. 2018, 21, 705–717. [Google Scholar] [CrossRef]
- Merlo, A.; De Quiros, S.B.; De Santa-María, I.S.; Pitiot, A.S.; Balbin, M.; Astudillo, A.; Scola, B.; Arístegui, M.; Quer, M.; Suárez, C.; et al. Identification of Somatic VHL Gene Mutations in Sporadic Head and Neck Paragangliomas in Association With Activation of the HIF-1?/miR-210 Signaling Pathway. J. Clin. Endocrinol. Metab. 2013, 98, 1661–1666. [Google Scholar] [CrossRef]
- Castro-Vega, L.J.; Buffet, A.; De Cubas, A.A.; Cascón, A.; Menara, M.; Khalifa, E.; Amar, L.; Azriel, S.; Bourdeau, I.; Chabre, O.; et al. Germline mutations in FH confer predisposition to malignant pheochromocytomas and paragangliomas. Hum. Mol. Genet. 2013, 23, 2440–2446. [Google Scholar] [CrossRef]
- Buffet, A.; Morin, A.; Castro-Vega, L.-J.; Habarou, F.; Lussey-Lepoutre, C.; Letouzé, E.; Lefebvre, H.; Guilhem, I.; Haissaguerre, M.; Raingeard, I.; et al. Germline Mutations in the Mitochondrial 2-Oxoglutarate/Malate Carrier SLC25A11 Gene Confer a Predisposition to Metastatic Paragangliomas. Cancer Res. 2018, 78, 1914–1922. [Google Scholar] [CrossRef]
- Van Nederveen, F.H.; Gaal, J.; Favier, J.; Korpershoek, E.; Oldenburg, R.A.; A De Bruyn, E.M.C.; Sleddens, H.F.B.M.; Derkx, P.; Rivière, J.; Dannenberg, H.; et al. An immunohistochemical procedure to detect patients with paraganglioma and phaeochromocytoma with germline SDHB, SDHC, or SDHD gene mutations: A retrospective and prospective analysis. Lancet Oncol. 2009, 10, 764–771. [Google Scholar] [CrossRef]
- Gill, A.J. Succinate dehydrogenase (SDH)-deficient neoplasia. Histopathology 2017, 72, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Papathomas, T.G.; Oudijk, L.; Persu, A.; Gill, A.J.; Van Nederveen, F.; Tischler, A.S.; Tissier, F.; Volante, M.; Matias-Guiu, X.; Smid, M.; et al. SDHB/SDHA immunohistochemistry in pheochromocytomas and paragangliomas: A multicenter interobserver variation analysis using virtual microscopy: A Multinational Study of the European Network for the Study of Adrenal Tumors (ENS@T). Mod. Pathol. 2015, 28, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Wallace, P.W.; Conrad, C.; Brückmann, S.; Pang, Y.; Caleiras, E.; Murakami, M.; Korpershoek, E.; Zhuang, Z.; Rapizzi, E.; Kroiss, M.; et al. Metabolomics, machine learning and immunohistochemistry to predict succinate dehydrogenase mutational status in phaeochromocytomas and paragangliomas. J. Pathol. 2020, 251, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Landrum, M.; Lee, J.M.; Benson, M.; Brown, G.R.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, U.; Jang, W.; et al. ClinVar: Improving access to variant interpretations and supporting evidence. Nucleic Acids Res. 2017, 46, D1062–D1067. [Google Scholar] [CrossRef]
- Sherry, S.T.; Ward, M.-H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. dbSNP: The NCBI database of genetic variation. Nucleic Acids Res. 2001, 29, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res. 2018, 47, D941–D947. [Google Scholar] [CrossRef]
- Vaser, R.; Adusumalli, S.; Leng, S.N.; Sikic, M.; Ng, P.C. SIFT missense predictions for genomes. Nat. Protoc. 2015, 11, 1–9. [Google Scholar] [CrossRef]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat Methods 2010, 7, 248–249. [Google Scholar] [CrossRef]
- Schwarz, J.M.; Cooper, D.N.; Schuelke, M.; Seelow, D. MutationTaster2: Mutation prediction for the deep-sequencing age. Nat. Methods 2014, 11, 361–362. [Google Scholar] [CrossRef]
- Chun, S.; Fay, J.C. Identification of deleterious mutations within three human genomes. Genome Res. 2009, 19, 1553–1561. [Google Scholar] [CrossRef]
- Siepel, A.; Bejerano, G.; Pedersen, J.S.; Hinrichs, A.S.; Hou, M.; Rosenbloom, K.; Clawson, H.; Spieth, J.; Hillier, L.W.; Richards, S.; et al. Evolutionarily conserved elements in vertebrate, insect, worm, and yeast genomes. Genome Res. 2005, 15, 1034–1050. [Google Scholar] [CrossRef] [PubMed]
- Pollard, K.S.; Hubisz, M.J.; Rosenbloom, K.R.; Siepel, A. Detection of nonneutral substitution rates on mammalian phylogenies. Genome Res. 2009, 20, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; on behalf of the ACMG Laboratory Quality Assurance Committee; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–423. [Google Scholar] [CrossRef] [PubMed]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Sci. 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, G.S.; Dmitriev, A.A.; Snezhkina, A.V.; Kudryavtseva, A.V. Deregulation of glycolysis in cancer: Glyceraldehyde-3-phosphate dehydrogenase as a therapeutic target. Expert Opin. Ther. Targets 2013, 17, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Kudryavtseva, A.V.; Krasnov, G.S.; Dmitriev, A.A.; Alekseev, B.Y.; Kardymon, O.; Sadritdinova, A.F.; Fedorova, M.S.; Pokrovsky, A.V.; Melnikova, N.V.; Kaprin, A.D.; et al. Mitochondrial dysfunction and oxidative stress in aging and cancer. Oncotarget 2016, 7, 44879–44905. [Google Scholar] [CrossRef]
- Senyilmaz, D.; Teleman, A.A. Chicken or the egg: Warburg effect and mitochondrial dysfunction. F1000Prime Rep. 2015, 7, 41. [Google Scholar] [CrossRef]
- Zhikrivetskaya, S.O.; Snezhkina, A.V.; Zaretsky, A.R.; Alekseev, B.Y.; Pokrovsky, A.V.; Golovyuk, A.L.; Melnikova, N.V.; Stepanov, O.A.; Kalinin, D.V.; Moskalev, A.; et al. Molecular markers of paragangliomas/pheochromocytomas. Oncotarget 2017, 8, 25756–25782. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxidative Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Aldera, A.P.; Govender, D. Gene of the month: SDH. J. Clin. Pathol. 2017, 71, 95–97. [Google Scholar] [CrossRef]
- Rana, H.; Rainville, I.R.; Vaidya, A. Genetic testing in the clinical care of patients with pheochromocytoma and paraganglioma. Curr. Opin. Endocrinol. Diabetes Obes. 2014, 21, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.; Tischler, A.S. Update from the 4th Edition of the World Health Organization Classification of Head and Neck Tumours: Paragangliomas. Head Neck Pathol. 2017, 11, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Pasini, B.; Stratakis, C.A.; Pasini, B. SDH mutations in tumorigenesis and inherited endocrine tumours: Lesson from the phaeochromocytoma-paraganglioma syndromes. J. Intern. Med. 2009, 266, 19–42. [Google Scholar] [CrossRef] [PubMed]
- Schiavi, F.; Boedeker, C.C.; Bausch, B.; Peczkowska, M.; Gomez, C.F.; Strassburg, T.; Pawlu, C.; Buchta, M.; Salzmann, M.; Hoffmann, M.M.; et al. Predictors and Prevalence of Paraganglioma Syndrome Associated With Mutations of the SDHC Gene. JAMA 2005, 294, 2057. [Google Scholar] [CrossRef]
- Gottlieb, E.; Tomlinson, I.P.M. Mitochondrial tumour suppressors: A genetic and biochemical update. Nat. Rev. Cancer 2005, 5, 857–866. [Google Scholar] [CrossRef]
- Gimenez-Roqueplo, A.-P.; Favier, J.; Rustin, P.; Mourad, J.-J.; Plouin, P.-F.; Corvol, P.; Rötig, A.; Jeunemaître, X. The R22X Mutation of the SDHD Gene in Hereditary Paraganglioma Abolishes the Enzymatic Activity of Complex II in the Mitochondrial Respiratory Chain and Activates the Hypoxia Pathway. Am. J. Hum. Genet. 2001, 69, 1186–1197. [Google Scholar] [CrossRef]
- Dekker, P.D.; Hogendoorn, P.C.; Kuipers-Dijkshoorn, N.; Prins, F.; Van Duinen, S.; Taschner, P.E.; Van Der Mey, A.; Cornelisse, C.J. SDHD mutations in head and neck paragangliomas result in destabilization of complex II in the mitochondrial respiratory chain with loss of enzymatic activity and abnormal mitochondrial morphology. J. Pathol. 2003, 201, 480–486. [Google Scholar] [CrossRef]
- Santi, R.; Rapizzi, E.; Canu, L.; Ercolino, T.; Baroni, G.; Fucci, R.; Costa, G.; Mannelli, M.; Nesi, G. Potential Pitfalls of SDH Immunohistochemical Detection in Paragangliomas and Phaeochromocytomas Harbouring Germline SDHx Gene Mutation. Anticancer Res. 2017, 37, 805–812. [Google Scholar] [CrossRef]
- Castelblanco, E.; Santacana, M.; Valls, J.; De Cubas, A.; Cascón, A.; Robledo, M.; Matias-Guiu, X. Usefulness of Negative and Weak–Diffuse Pattern of SDHB Immunostaining in Assessment of SDH Mutations in Paragangliomas and Pheochromocytomas. Endocr. Pathol. 2013, 24, 199–205. [Google Scholar] [CrossRef]
- Gill, A.J.; Benn, D.E.; Chou, A.; Clarkson, A.; Muljono, A.; Meyer-Rochow, G.Y.; Richardson, A.L.; Sidhu, S.B.; Robinson, B.G.; Clifton-Bligh, R. Immunohistochemistry for SDHB triages genetic testing of SDHB, SDHC, and SDHD in paraganglioma-pheochromocytoma syndromes. Hum. Pathol. 2010, 41, 805–814. [Google Scholar] [CrossRef]
- Pavlov, V.S.; Kalinin, D.V.; Lukyanova, E.N.; Golovyuk, A.L.; Fedorova, M.S.; Pudova, E.A.; Savvateeva, M.V.; Lipatova, A.V.; Guvatova, Z.G.; Kaprin, A.D.; et al. Multiple paragangliomas: A case report. BMC Med Genom. 2020, 13, 125. [Google Scholar] [CrossRef] [PubMed]
- Burnichon, N.; Brière, J.-J.; Libé, R.; Vescovo, L.; Rivière, J.; Tissier, F.; Jouanno, E.; Jeunemaitre, X.; Bénit, P.; Tzagoloff, A.; et al. SDHA is a tumor suppressor gene causing paraganglioma. Hum. Mol. Genet. 2010, 19, 3011–3020. [Google Scholar] [CrossRef] [PubMed]
- Dwight, T.; Mann, K.; Benn, D.E.; Robinson, B.G.; McKelvie, P.; Gill, A.J.; Winship, I.; Clifton-Bligh, R. FamilialSDHAMutation Associated With Pituitary Adenoma and Pheochromocytoma/Paraganglioma. J. Clin. Endocrinol. Metab. 2013, 98, E1103–E1108. [Google Scholar] [CrossRef] [PubMed]
- Snezhkina, A.V.; Lukyanova, E.; Kalinin, D.V.; Pokrovsky, A.V.; Dmitriev, A.A.; Koroban, N.V.; Pudova, E.A.; Fedorova, M.S.; Volchenko, N.N.; Stepanov, O.A.; et al. Exome analysis of carotid body tumor. BMC Med Genom. 2018, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Kudryavtseva, A.V.; Lukyanova, E.; Kalinin, D.V.; Zaretsky, A.R.; Pokrovsky, A.V.; Golovyuk, A.L.; Fedorova, M.S.; Pudova, E.A.; Kharitonov, S.L.; Pavlov, V.S.; et al. Mutational load in carotid body tumor. BMC Med Genom. 2019, 12, 39. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]
- The 1000 Genomes Project Consortium A global reference for human genetic variation. Nature 2015, 526, 68–74. [CrossRef]
- Karczewski, K.; Weisburd, B.; Thomas, B.; Solomonson, M.; Ruderfer, D.M.; Kavanagh, D.; Hamamsy, T.; Lek, M.; Samocha, K.E.; Cummings, B.B.; et al. The ExAC browser: Displaying reference data information from over 60,000 exomes. Nucleic Acids Res. 2016, 45, D840–D845. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, K.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Glusman, G.; Caballero, J.; Mauldin, D.E.; Hood, L.; Roach, J.C. Kaviar: An accessible system for testing SNV novelty. Bioinformatics 2011, 27, 3216–3217. [Google Scholar] [CrossRef]
- Mitchell, A.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2018, 47, D351–D360. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).