Circulating Long Non-Coding RNA GAS5 Is Overexpressed in Serum from Osteoporotic Patients and Is Associated with Increased Risk of Bone Fragility

 , , , ,
, , , ,  ,
, 


Abstract
1. Introduction
2. Results
2.1. Clinical Characteristics of Individuals Included in the Study
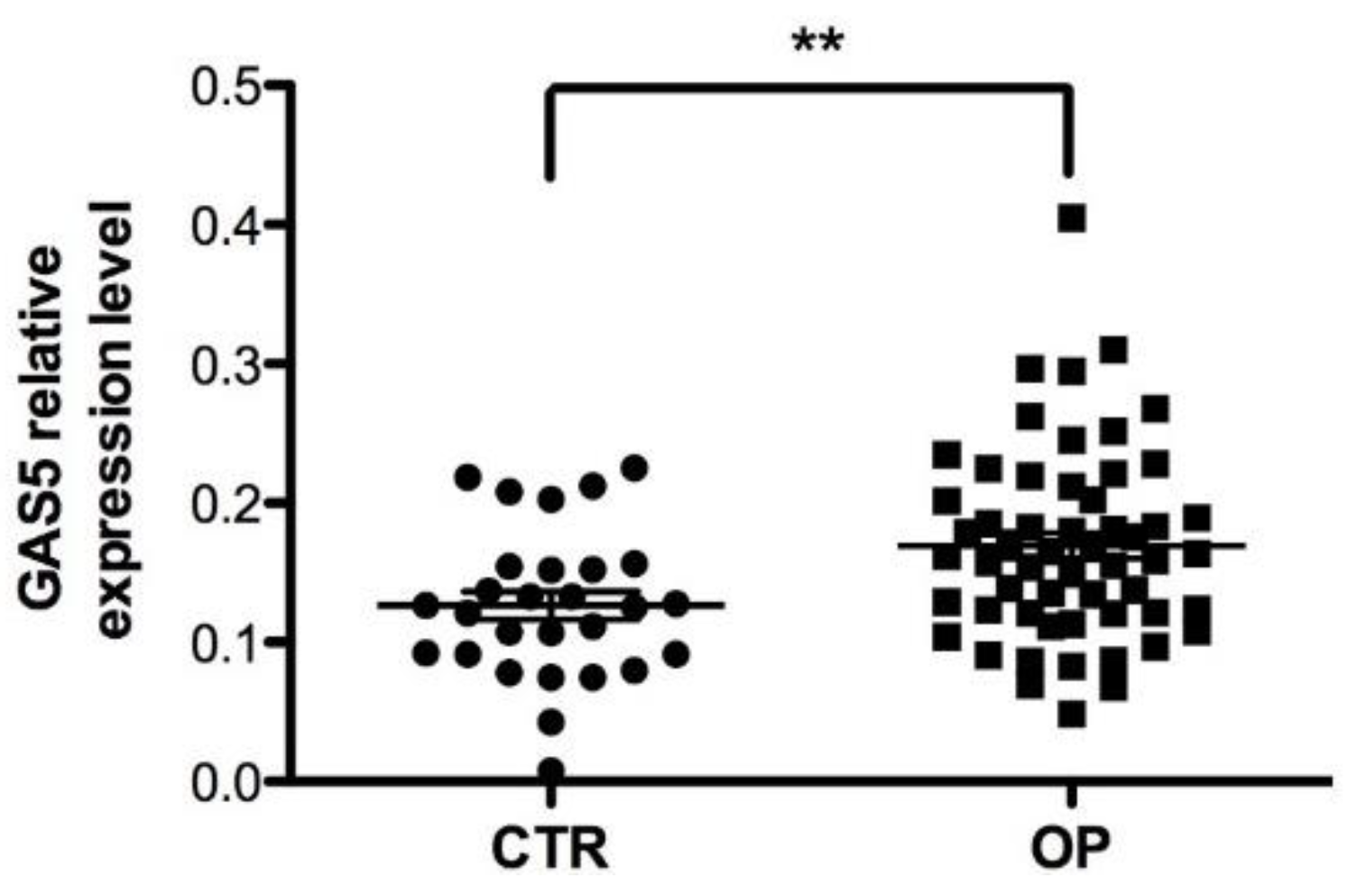
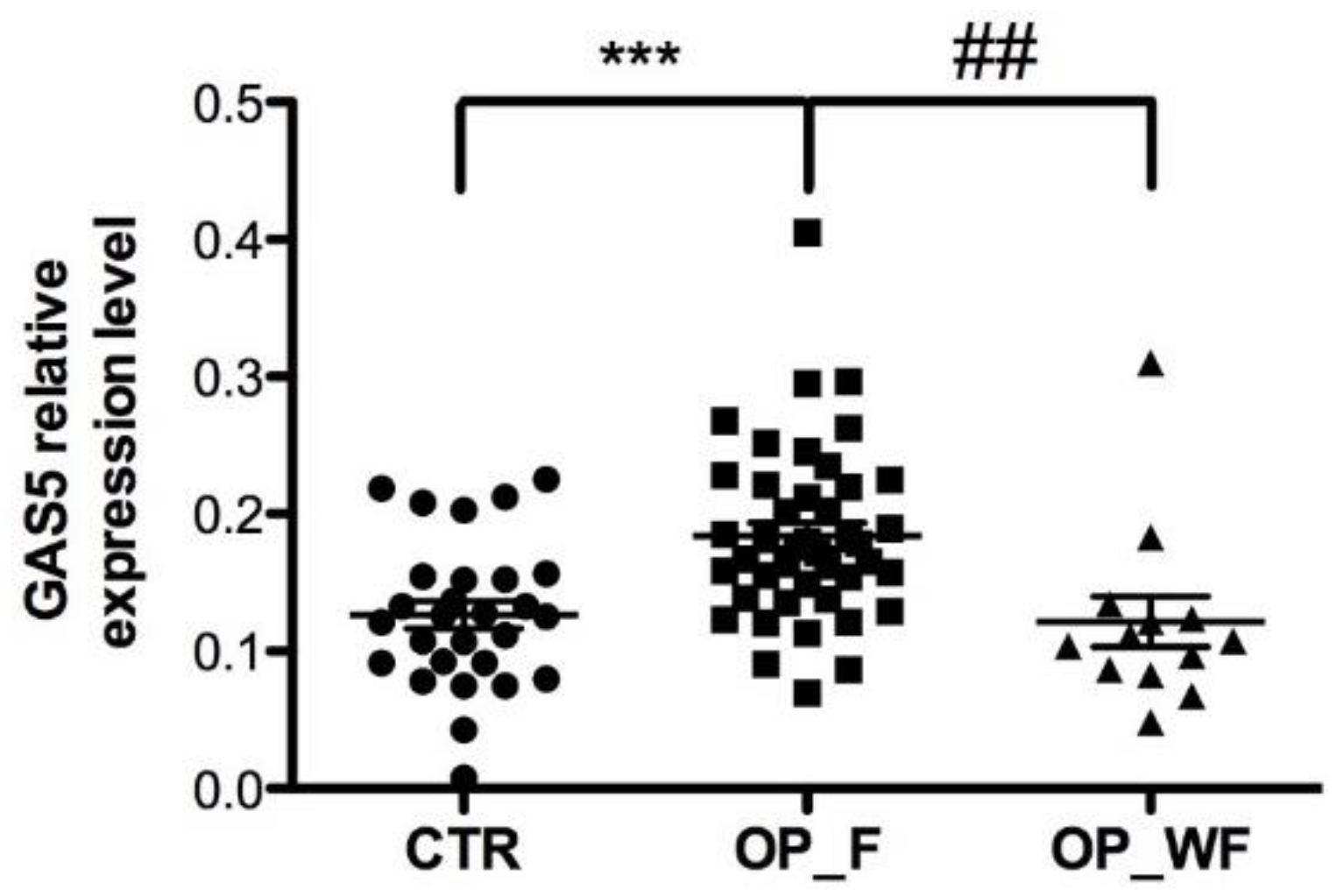
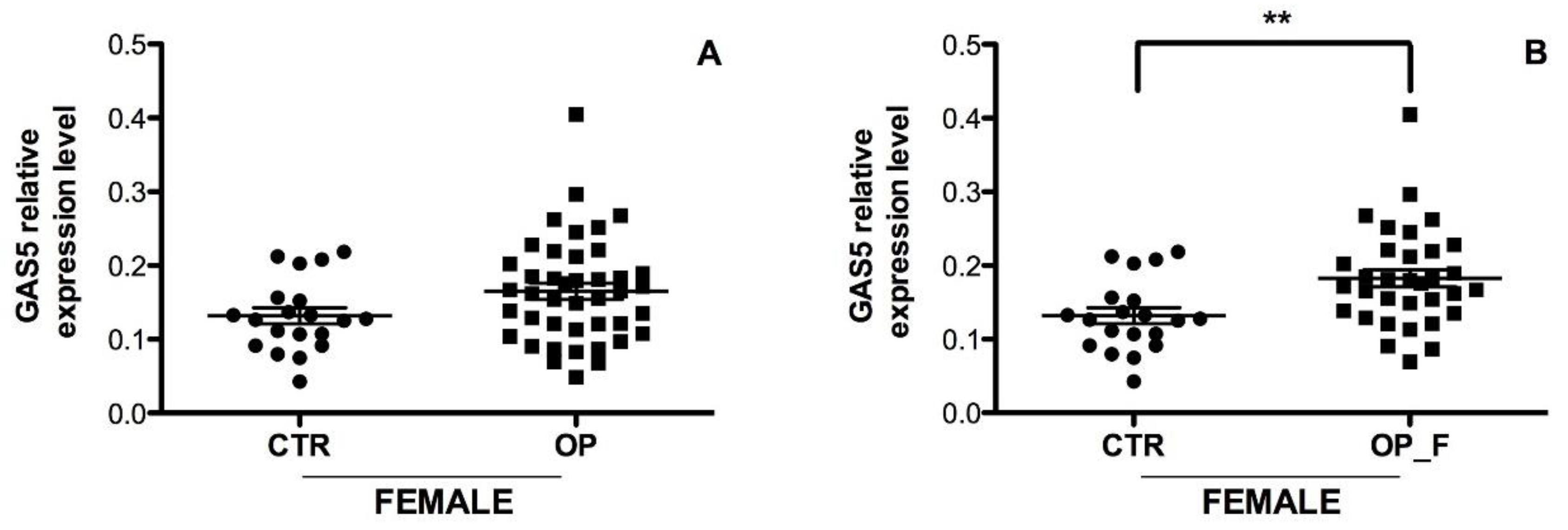
2.2. GAS5 Expression Level Is Significantly Upregulated in Serum from OP Patients with Fragility Fractures
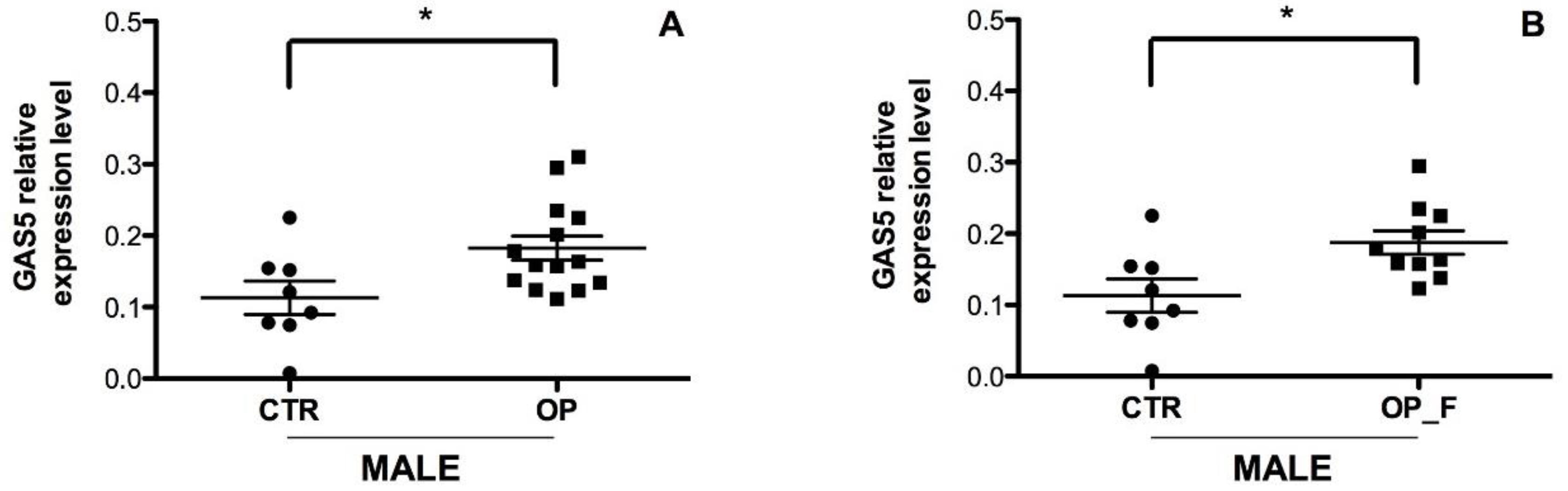
2.3. Correlation between Serum Circulating GAS5 with Clinicopathological Characteristics of Osteoporotic Patients
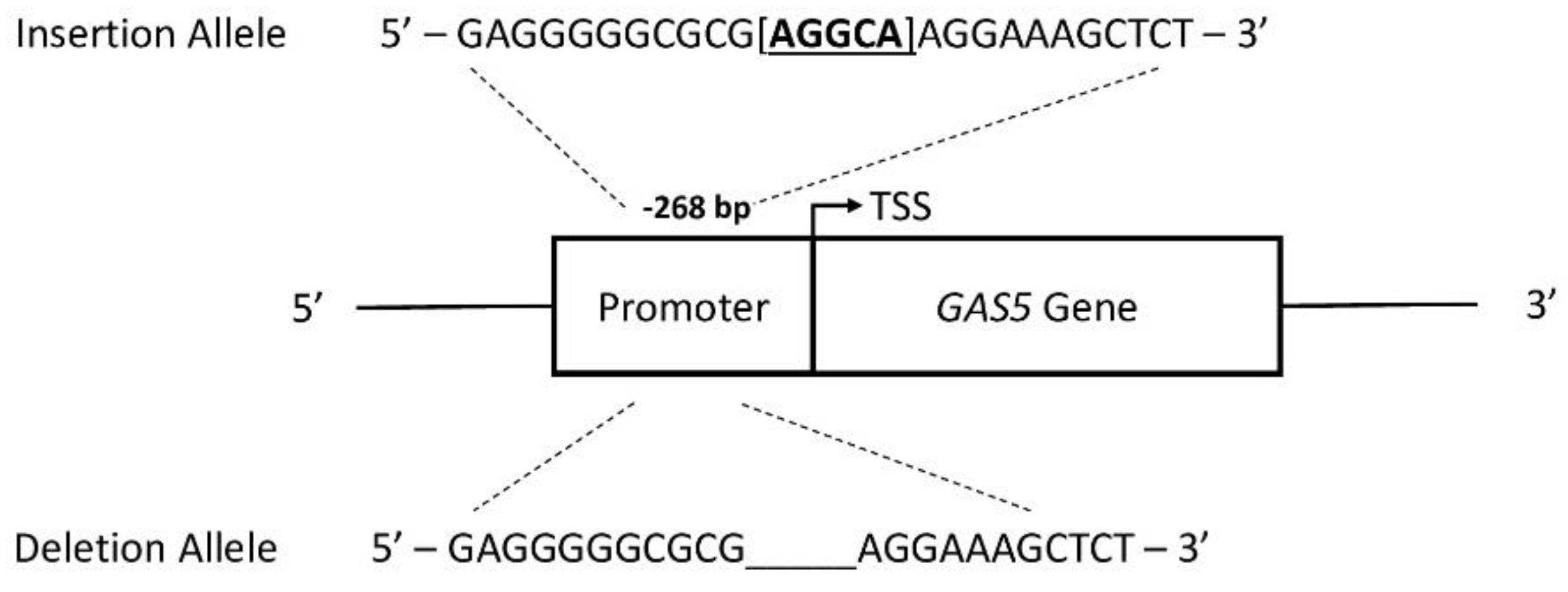
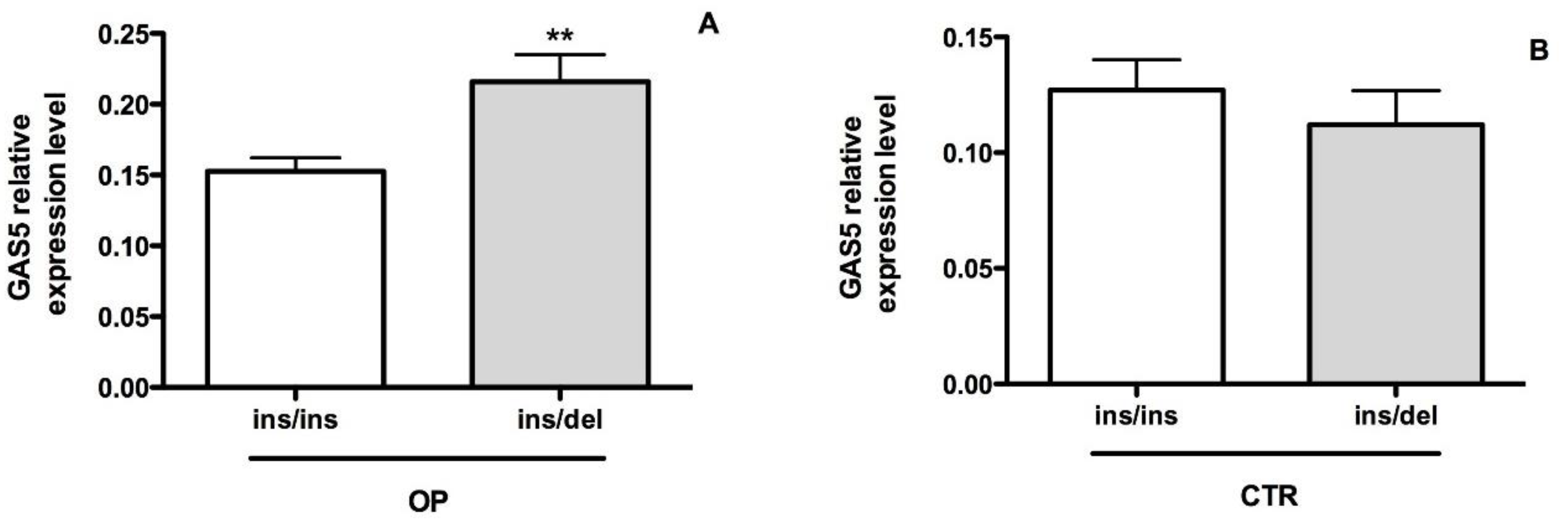
2.4. The rs145204276 Polymorphism Is Associated with the Expression Levels of GAS5
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Specimen Collection
4.3. Clinical and Biochemical Parameters
4.4. Haemolysis Evaluation
4.5. RNA Extraction and RT-qPCR Analysis of Circulating GAS5 Expression
4.6. Rs145204276 Genotyping Assay
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OP | Osteoporosis |
| CTR | Control |
| ncRNA | non-coding RNA |
| LncRNA | long non-coding RNA |
| OP_VF | OP vertebral fractures |
| OP_FF | OP femoral fractures |
| OP_WF | OP without fracture |
| BMD | bone mineral density |
| GAS5 | growth arrest-specific transcript 5 |
| ceRNA | competing endogenous RNA |
| hMSC | human multi-potential mesenchymal stem cell |
| DEXA | dual-energy x-ray absorptiometry |
| 25-(OH)-Vit D | 25-hydroxyvitamin D |
| PTH | parathyroid hormone |
| ALP | alkaline phosphatase |
| BALP | bone alkaline phosphatase |
| CTX | serum C-terminal telopeptide |
| BMI | body mass index |
| BMSC | bone marrow mesenchymal stem cell |
References
- Sözen, T.; Özışık, L.; Başaran, N.Ç. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, R.S.; Manolagas, S.C. Apoptosis and osteoporosis. Am. J. Med. 2000, 108, 153–164. [Google Scholar] [CrossRef]
- Marini, F.; Cianferotti, L.; Brandi, M. Epigenetic Mechanisms in Bone Biology and Osteoporosis: Can They Drive Therapeutic Choices? Int. J. Mol. Sci. 2016, 17, 1329. [Google Scholar] [CrossRef] [PubMed]
- Del Real, A.; Riancho-Zarrabeitia, L.; López-Delgado, L.; Riancho, J.A. Epigenetics of Skeletal Diseases. Curr. Osteoporos. Rep. 2018, 16, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Ghayor, C.; Weber, F.E. Epigenetic regulation of bone remodeling and its impacts in osteoporosis. Int. J. Mol. Sci. 2016, 17, 1446. [Google Scholar] [CrossRef]
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long non-coding RNAs in the regulation of gene expression: Physiology and disease. Non-Coding RNA 2019, 5, 17. [Google Scholar] [CrossRef]
- Yavropoulou, M.P.; Anastasilakis, A.D.; Makras, P.; Tsalikakis, D.G.; Grammatiki, M.; Yovos, J.G. Expression of microRNAs that regulate bone turnover in the serum of postmenopausal women with low bone mass and vertebral fractures. Eur. J. Endocrinol. 2017, 176, 169–176. [Google Scholar] [CrossRef]
- Silva, A.M.; Moura, S.R.; Teixeira, J.H.; Barbosa, M.A.; Santos, S.G.; Almeida, M.I. Long noncoding RNAs: A missing link in osteoporosis. Bone Res. 2019, 7, 10. [Google Scholar] [CrossRef]
- Centofanti, F.; Santoro, M.; Marini, M.; Visconti, V.V.; Rinaldi, A.M.; Celi, M.; D’arcangelo, G.; Novelli, G.; Orlandi, A.; Tancredi, V.; et al. Identification of Aberrantly-Expressed Long Non-Coding RNAs in Osteoblastic Cells from Osteoporotic Patients. Biomedicines 2020, 8, 65. [Google Scholar] [CrossRef]
- Luo, G.; Liu, D.; Huang, C.; Wang, M.; Xiao, X.; Zeng, F.; Wang, L.; Jiang, G. LncRNA GAS5 inhibits cellular proliferation by targeting P27Kip1. Mol. Cancer Res. 2017, 15, 789–799. [Google Scholar] [CrossRef]
- Goustin, A.S.; Thepsuwan, P.; Kosir, M.A.; Lipovich, L. The Growth-Arrest-Specific (GAS)-5 Long Non-Coding RNA: A Fascinating lncRNA Widely Expressed in Cancers. Non-Coding RNA 2019, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Li, Y.; Jiang, F.; Wei, C.; Chen, G.; Zhang, W.; Zhao, W.; Yu, D. LncRNA GAS5 suppresses proliferation, migration, invasion, and epithelial-mesenchymal transition in oral squamous cell carcinoma by regulating the miR-21/PTEN axis. Exp. Cell Res. 2019, 374, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Pickard, M.R.; Williams, G.T. Molecular and cellular mechanisms of action of tumour suppressor GAS5 LncRNA. Genes 2015, 6, 484–499. [Google Scholar] [CrossRef]
- Song, J.; Ahn, C.; Chun, C.-H.; Jin, E.-J. A long non-coding RNA, GAS5, plays a critical role in the regulation of miR-21 during osteoarthritis. J. Orthop. Res. 2014, 32, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhang, N.; Zheng, Y.; Chen, Y.D.; Liu, J.; Yang, M. LncRNA GAS5 Indel Genetic Polymorphism Contributes to Glioma Risk Through Interfering Binding of Transcriptional Factor TFAP2A. DNA Cell Biol. 2018, 37, 750–757. [Google Scholar] [CrossRef]
- Li, Q.; Ma, G.; Sun, S.; Xu, Y.; Wang, B. Polymorphism in the promoter region of lncRNA GAS5 is functionally associated with the risk of gastric cancer. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Aminian, K.; Mashayekhi, F.; Mirzanejad, L.; Salehi, Z. A functional genetic variant in GAS5 lncRNA (rs145204276) modulates p27Kip1 expression and confers risk for gastric cancer. Br. J. Biomed. Sci. 2019, 76, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Mayama, T.; Marr, A.K.; Kino, T. Differential Expression of Glucocorticoid Receptor Noncoding RNA Repressor Gas5 in Autoimmune and Inflammatory Diseases. Horm. Metab. Res. 2016, 48, 550–557. [Google Scholar] [CrossRef]
- Wang, Y.; Song, Q.; Huang, X.; Chen, Z.; Zhang, F.; Wang, K.; Huang, G.; Shen, H. Long noncoding RNA GAS5 promotes apoptosis in primary nucleus pulposus cells derived from the human intervertebral disc via Bcl-2 downregulation and caspase-3 upregulation. Mol. Med. Rep. 2019, 19, 2164–2172. [Google Scholar] [CrossRef]
- Ji, J.; Dai, X.; Yeung, S.C.J.; He, X. The role of long non-coding RNA GAS5 in cancers. Cancer Manag. Res. 2019, 11, 2729–2737. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, Z.; Watabe, K.; Zhang, X.; Bai, C.; Xu, M.; Wu, F.; Mo, Y.-Y. Negative regulation of lncRNA GAS5 by miR-21. Cell Death Differ. 2013, 20, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; She, Y.; Wu, H.; Zhong, D.; Zhang, J. Long non-coding RNA Gas5 regulates proliferation and apoptosis in HCS-2/8 cells and growth plate chondrocytes by controlling FGF1 expression via miR-21 regulation. J. Biomed. Sci. 2018, 25, 18. [Google Scholar] [CrossRef] [PubMed]
- Fei, Q.; Bai, X.; Lin, J.; Meng, H.; Yang, Y.; Guo, A. Identification of aberrantly expressed long non-coding RNAs in postmenopausal osteoporosis. Int. J. Mol. Med. 2018, 41, 3537–3550. [Google Scholar] [CrossRef]
- Feng, J.; Wang, J.-X.; Li, C.-H. LncRNA GAS5 overexpression alleviates the development of osteoporosis through promoting osteogenic differentiation of MSCs via targeting microRNA-498 to regulate RUNX2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7757–7765. [Google Scholar] [PubMed]
- Wang, X.; Zhao, D.; Zhu, Y.; Dong, Y.; Liu, Y. Long non-coding RNA GAS5 promotes osteogenic differentiation of bone marrow mesenchymal stem cells by regulating the miR-135a-5p/FOXO1 pathway. Mol. Cell. Endocrinol. 2019, 496, 110534. [Google Scholar] [CrossRef]
- Gezer, U.; Özgür, E.; Cetinkaya, M.; Isin, M.; Dalay, N. Long non-coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol. Int. 2014, 38, 1076–1079. [Google Scholar] [CrossRef]
- Koldemir, O.; Özgür, E.; Gezer, U. Accumulation of GAS5 in exosomes is a marker of apoptosis induction. Biomed. Rep. 2017, 6, 358–362. [Google Scholar] [CrossRef]
- Dragomir, M.; Chen, B.; Calin, G.A. Exosomal lncRNAs as new players in cell-to-cell communication. Transl. Cancer Res. 2018, 7, S243–S252. [Google Scholar] [CrossRef]
- Lu, X.; Bai, D.; Liu, X.; Zhou, C.; Yang, G. Sedentary lifestyle related exosomal release of Hotair from gluteal-femoral fat promotes intestinal cell proliferation. Sci. Rep. 2017, 7, 45648. [Google Scholar] [CrossRef]
- Li, B.; Hu, X.; Yang, Y.; Zhu, M.; Zhang, J.; Wang, Y.; Pei, X.; Zhou, H.; Wu, J. GAS5/miR-21 Axis as a Potential Target to Rescue ZCL-082-Induced Autophagy of Female Germline Stem Cells In Vitro. Mol. Ther. Nucleic Acids 2019, 17, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zou, Y.; Jiang, D. Honokiol suppresses proliferation and induces apoptosis via regulation of the miR-21/PTEN/PI3K/AKT signaling pathway in human osteosarcoma cells. Int. J. Mol. Med. 2018, 41, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, B.; Li, C. Knockdown of long noncoding RNA GAS5 protects human cardiomyocyte-like AC16 cells against high glucose-induced inflammation by inhibiting miR-21-5p-mediated TLR4/NF-κB signaling. Naunyn. Schmiedebergs. Arch. Pharmacol. 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Z.; Fu, Q.; Zhang, J. Plasma miRNA levels correlate with sensitivity to bone mineral density in postmenopausal osteoporosis patients. Biomarkers 2014, 19, 553–556. [Google Scholar] [CrossRef]
- Chen, P.; Krege, J.H.; Adachi, J.D.; Prior, J.C.; Tenenhouse, A.; Brown, J.P.; Papadimitropoulos, E.; Kreiger, N.; Olszynski, W.P.; Josse, R.G.; et al. Vertebral fracture status and the World Health Organization risk factors for predicting osteoporotic fracture risk. J. Bone Miner. Res. 2009, 24, 495–502. [Google Scholar] [CrossRef]
- Shen, Z.; She, Q. Association between the Deletion Allele of Ins/Del Polymorphism (Rs145204276) in the Promoter Region of GAS5 with the Risk of Atherosclerosis. Cell. Physiol. Biochem. 2018, 49, 1431–1443. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, U.; Iolascon, G.; Cianferotti, L.; Masi, L.; Marcucci, G.; Giusti, F.; Marini, F.; Parri, S.; Feola, M.; Rao, C.; et al. Clinical guidelines for the prevention and treatment of osteoporosis: Summary statements and recommendations from the Italian Society for Orthopaedics and Traumatology. J. Orthop. Traumatol. 2017, 18, 3–36. [Google Scholar] [CrossRef]
- Heaney, R.P. Toward a physiological referent for the vitamin D requirement. J. Endocrinol. Investig. 2014, 37, 1127–1130. [Google Scholar] [CrossRef]
- Cawthon, P.M. Gender differences in osteoporosis and fractures. Clin. Orthop. Relat. Res. 2011, 469, 1900–1905. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | OP (14 Males; 42 Females) | CTRs (8 Males; 20 Females) | T-test (Mann-Whitney Test) |
|---|---|---|---|
| Age (years) | 68.16 ± 4.90 | 67.00 ± 4.95 | NS (p = 0.304) |
| BMI (Kg/cm2) | 24.38 ± 3.29 | 28.72 ± 5.70 | *** (p < 0.001) |
| t-score L1-L4 | −2.87 ± 1.09 | 0.23 ± 1.35 | *** (p < 0.001) |
| t-score FN | −2.46 ± 0.82 | 0.58 ± 0.62 | *** (p < 0.001) |
| Calcium (mg/dL) | 9.21 ± 1.02 | 9.30 ± 0.48 | NS (p = 0.675) |
| Phosphorus (mg/dL) | 3.41 ± 0.65 | 3.28 ± 0.63 | NS (p = 0.312) |
| PTH (pg/mL) | 45.39 ± 24.66 | 45.29 ± 19.65 | NS (p = 0.628) |
| 25-(OH)-Vit D (ng/mL) | 28.04 ± 11.01 | 27.02 ± 11.57 | NS (p = 0.888) |
| ALP (U/L) | 90.80 ± 36.63 | 70.77 ± 14.86 | ** (p = 0.0096) |
| BALP (μg/L) | 16.67 ± 11.30 | 12.85 ± 4.80 | NS (p = 0.169) |
| s-CTX (ng/mL) | 3.45 ± 4.20 | 1.62 ± 2.80 | ** (p = 0.0013) |
| Genotype | p | Allele | p | Odds Ratio (95% CI) | ||||
|---|---|---|---|---|---|---|---|---|
| Del/Del | Del/Ins | Ins/Ins | Del | Ins | ||||
| OP (n = 56) | 0 | 17 (30.4%) | 39 (69.6%) | 0.61 | 17 (15.2%) | 95 (84.8%) | 0.64 | 1.25 (0.48–3.22) |
| CTRs (n = 28) | 0 | 7 (25%) | 21 (75%) | 7 (12.5%) | 49 (87.5%) | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visconti, V.V.; Fittipaldi, S.; Ciuffi, S.; Marini, F.; Isaia, G.; D’Amelio, P.; Migliaccio, S.; Marcocci, C.; Minisola, S.; Nuti, R.; et al. Circulating Long Non-Coding RNA GAS5 Is Overexpressed in Serum from Osteoporotic Patients and Is Associated with Increased Risk of Bone Fragility. Int. J. Mol. Sci. 2020, 21, 6930. https://doi.org/10.3390/ijms21186930
Visconti VV, Fittipaldi S, Ciuffi S, Marini F, Isaia G, D’Amelio P, Migliaccio S, Marcocci C, Minisola S, Nuti R, et al. Circulating Long Non-Coding RNA GAS5 Is Overexpressed in Serum from Osteoporotic Patients and Is Associated with Increased Risk of Bone Fragility. International Journal of Molecular Sciences. 2020; 21(18):6930. https://doi.org/10.3390/ijms21186930
Chicago/Turabian StyleVisconti, Virginia Veronica, Simona Fittipaldi, Simone Ciuffi, Francesca Marini, Giancarlo Isaia, Patrizia D’Amelio, Silvia Migliaccio, Claudio Marcocci, Salvatore Minisola, Ranuccio Nuti, and et al. 2020. "Circulating Long Non-Coding RNA GAS5 Is Overexpressed in Serum from Osteoporotic Patients and Is Associated with Increased Risk of Bone Fragility" International Journal of Molecular Sciences 21, no. 18: 6930. https://doi.org/10.3390/ijms21186930
APA StyleVisconti, V. V., Fittipaldi, S., Ciuffi, S., Marini, F., Isaia, G., D’Amelio, P., Migliaccio, S., Marcocci, C., Minisola, S., Nuti, R., Novelli, G., Brandi, M. L., Botta, A., & Tarantino, U. (2020). Circulating Long Non-Coding RNA GAS5 Is Overexpressed in Serum from Osteoporotic Patients and Is Associated with Increased Risk of Bone Fragility. International Journal of Molecular Sciences, 21(18), 6930. https://doi.org/10.3390/ijms21186930