Antimicrobial Effects of Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy in Pseudomonas aeruginosa Induced Keratitis

 and
and
Abstract
1. Introduction
2. Methods
2.1. Human Corneal Epithelial Cell Culture
2.2. Minimum Inhibitory Concentration (MIC) Assay
2.3. Measurement of Antimicrobial Peptides (AMPs)
2.4. Statistics
3. Results
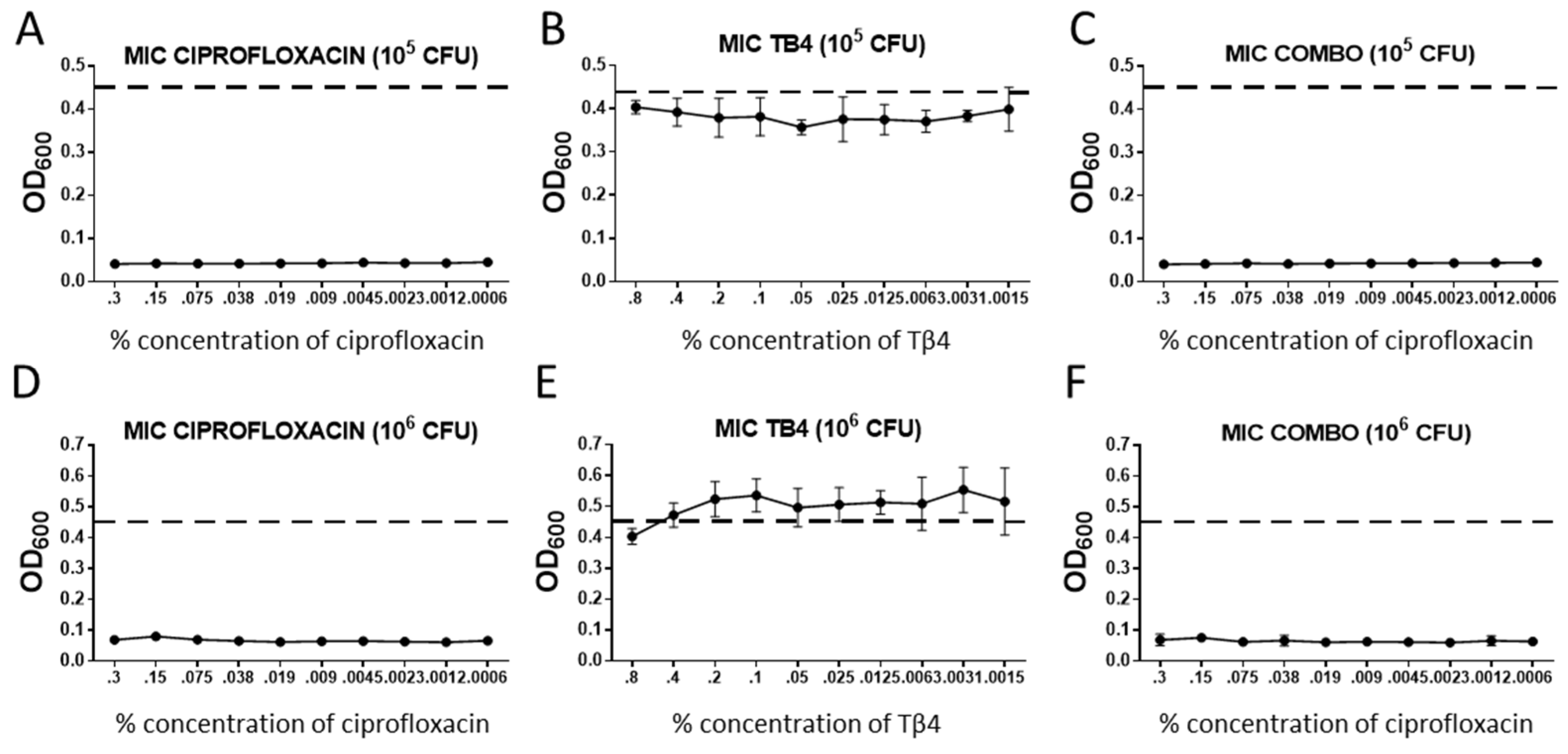
3.1. Tβ4 Does Not Appear to Directly Influence Bactericidal Activity
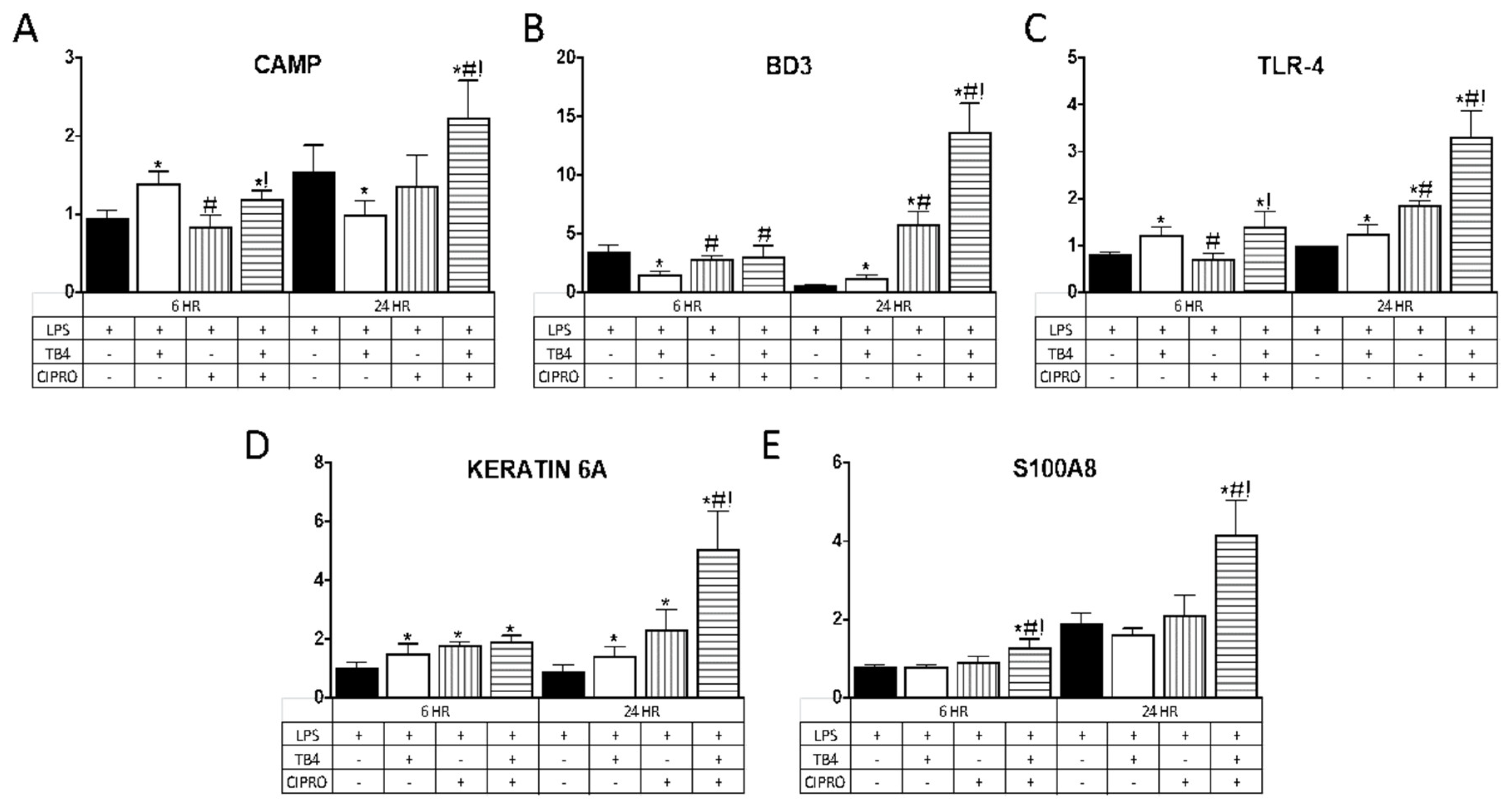
3.2. Tβ4 Upregulates AMP Expression in Human Telomerase-Immortalized Corneal Epithelial Cells HUCLs
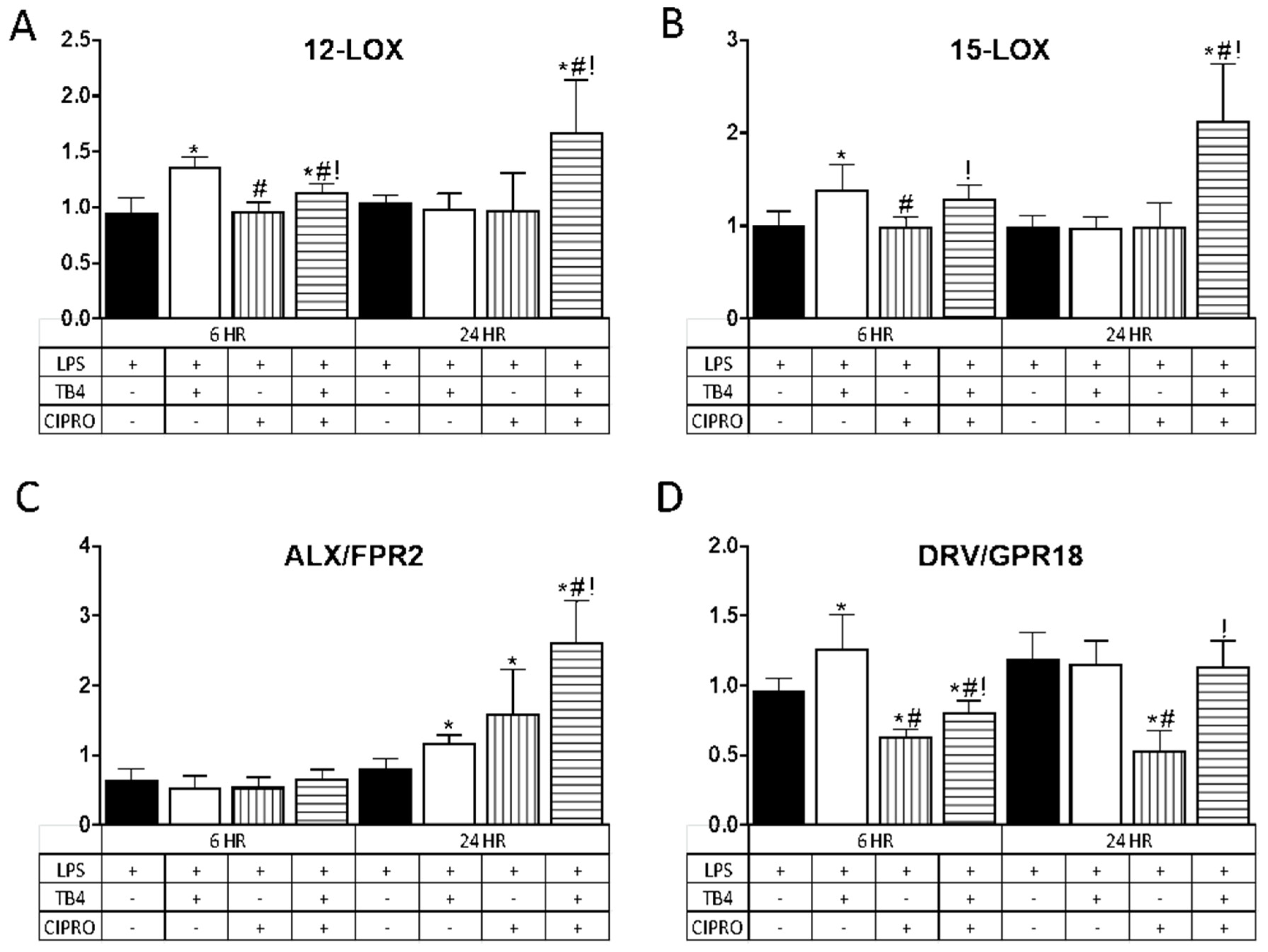
3.3. Tβ4 Influences Specialized Pro-Resolving Molecules (SPM)-Related Enzymes and Receptors in HUCLs
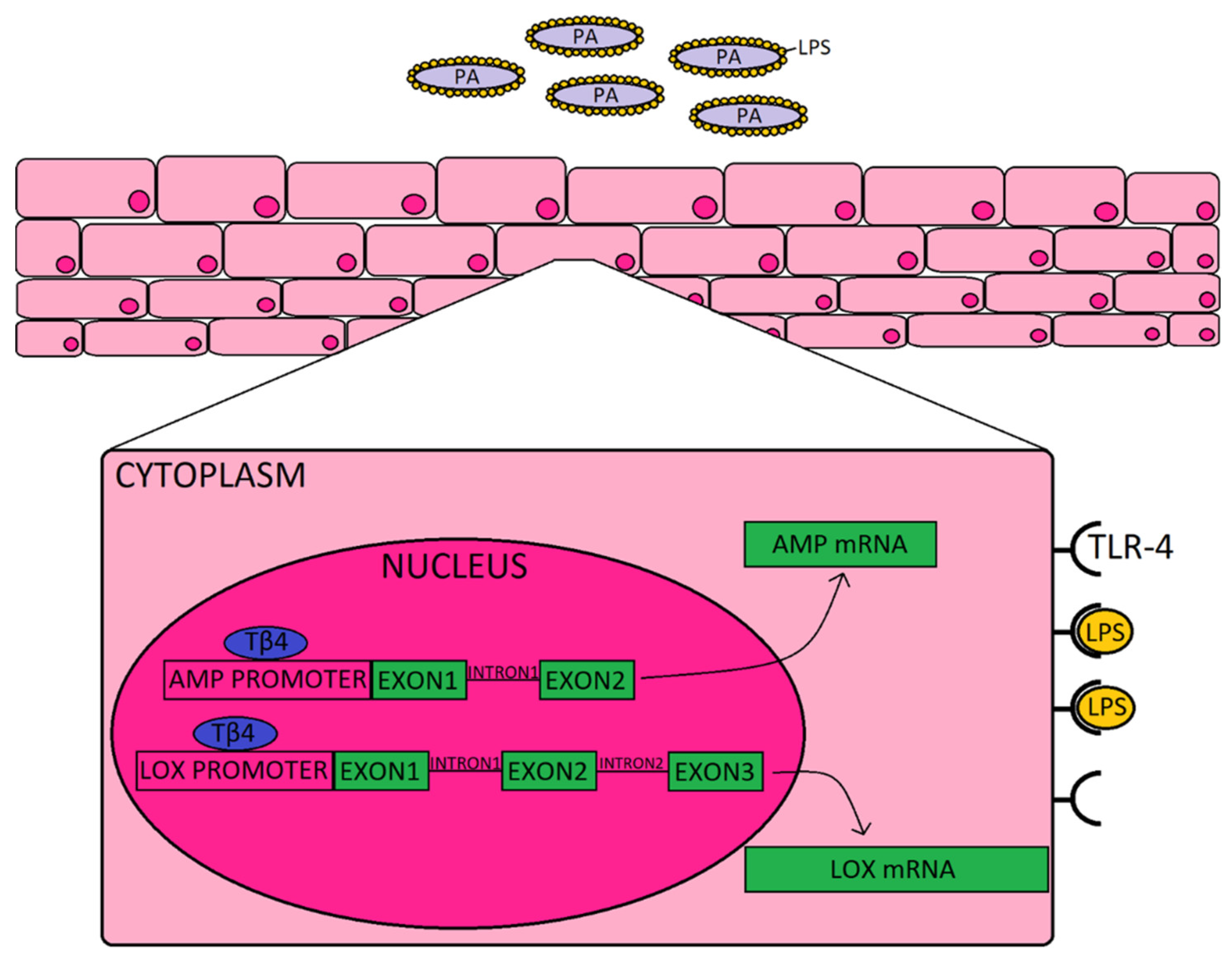
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Rattanatam, T.; Heng, W.J.; Rapuano, C.J.; Laibson, P.R.; Cohen, E.J. Trends in Contact Lens–related Corneal Ulcers. Cornea 2001, 20, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Pinna, A.; Usai, D.; Sechi, L.A.; Molicotti, P.; Zanetti, S.; Carta, A. Detection of Virulence Factors in Pseudomonas aeruginosa Strains Isolated From Contact Lens-Associated Corneal Ulcers. Cornea 2008, 27, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Lim, J.-A.; Choi, J.-S.; Kim, K.-A.; Joo, C.-K. The Resistance Patterns of Normal Ocular Bacterial Flora to 4 Fluoroquinolone Antibiotics. Cornea 2009, 28, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Sosa, A.B.; Epstein, S.P.; Asbell, P. Evaluation of Toxicity of Commercial Ophthalmic Fluoroquinolone Antibiotics as Assessed on Immortalized Corneal and Conjunctival Epithelial Cells. Cornea 2008, 27, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmus, K.R. Indecision about corticosteroids for bacterial keratitis. Ophthalmology 2002, 109, 835–842. [Google Scholar] [CrossRef]
- Low, T.L.; Hu, S.K.; Goldstein, A.L. Complete amino acid sequence of bovine thymosin beta 4: A thymic hormone that induces terminal deoxynucleotidyl transferase activity in thymocyte populations. Proc. Natl. Acad. Sci. USA 1981, 78, 1162–1166. [Google Scholar] [CrossRef]
- Huff, T.; Müller, C.S.G.; Hannappel, E. Thymosin beta4 Is Not Always the Main beta-Thymosin in Mammalian Platelets. Ann. N. Y. Acad. Sci. 2007, 1112, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.L.; Hannappel, E.; Sosne, G.; Kleinman, H.K. Thymosin beta4: A multi-functional regenerative peptide. Basic properties and clinical applications. Expert Opin. Biol. Ther. 2012, 12, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Freeman, K.W.; Bowman, B.R.; Zetter, B.R. Regenerative protein thymosin beta-4 is a novel regulator of purinergic signaling. FASEB J. 2011, 25, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Carion, T.W.; Ebrahim, A.S.; Kracht, D.; Agrawal, A.; Strand, E.; Kaddurah, O.; McWhirter, C.R.; Sosne, G.; Berger, E. Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy Improves Pseudomonas aeruginosa-Induced Keratitis. Cells 2018, 7, 145. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-Q.; Yeaman, M.R.; Selsted, M.E. Antimicrobial Peptides from Human Platelets. Infect. Immun. 2002, 70, 6524–6533. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.D.; Gilroy, D.W.; Serhan, C.N. Proresolving lipid mediators and mechanisms in the resolution of acute inflammation. Immunity 2014, 40, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of Growth and Differentiation in a Telomerase-Immortalized Human Corneal Epithelial Cell Line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, H.; Wang, J.; Dong, Z.; Mian, S.; Yu, F.-S.X. Role of EGFR Transactivation in Preventing Apoptosis inPseudomonas aeruginosa–Infected Human Corneal Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2569–2576. [Google Scholar] [CrossRef] [PubMed]
- Carion, T.W.; Greenwood, M.; Ebrahim, A.S.; Jerome, A.; Suvas, S.; Gronert, K.; Berger, E. Immunoregulatory role of 15-lipoxygenase in the pathogenesis of bacterial keratitis. FASEB J. 2018, 32, 5026–5038. [Google Scholar] [CrossRef]
- Roux, K.H. Optimization and troubleshooting in PCR. Genome Res. 1995, 4, S185–S194. [Google Scholar] [CrossRef]
- Kahlenberg, J.M.; Kaplan, M.J. Little Peptide, Big Effects: The Role of LL-37 in Inflammation and Autoimmune Disease. J. Immunol. 2013, 191, 4895–4901. [Google Scholar] [CrossRef]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: Biological and therapeutic considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in Inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef]
- Clark, H.L.; Jhingran, A.; Sun, Y.; Vareechon, C.; de Jesus, S.C.; Skaar, E.P.; Chazin, W.J.; Calera, J.A.; Hohl, T.M.; Pearlman, E. Zinc and Manganese Chelation by Neutrophil S100A8/A9 (Calprotectin) Limits Extracellular Aspergillus fumigatus Hyphal Growth and Corneal Infection. J. Immunol. 2016, 196, 336–344. [Google Scholar] [CrossRef]
- Wu, M.; McClellan, S.A.; Barrett, R.P.; Zhang, Y.; Hazlett, L.D. Beta-defensins 2 and 3 together promote resistance to Pseudomonas aeruginosa keratitis. J. Immunol. 2009, 183, 8054–8060. [Google Scholar] [CrossRef]
- Donnarumma, G.; Paoletti, I.; Fusco, A.; Perfetto, B.; Buommino, E.; de Gregorio, V.; Baroni, A. Beta-Defensins: Work in Progress. Adv. Exp. Med. Biol. 2016, 901, 59–76. [Google Scholar] [PubMed]
- Tam, C.; Mun, J.J.; Evans, D.J.; Fleiszig, S.M.J. Cytokeratins mediate epithelial innate defense through their antimicrobial properties. J. Clin. Investig. 2012, 122, 3665–3677. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Du, W.; McClellan, S.A.; Barrett, R.P.; Hazlett, L.D. TLR4 Is Required for Host Resistance inPseudomonas aeruginosaKeratitis. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4910–4916. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; McClellan, S.A.; Barrett, R.P.; Zhang, Y.; Hazlett, L.D. Vasoactive intestinal peptide downregulates proinflammatory TLRs while upregulating anti-inflammatory TLRs in the infected cornea. J. Immunol. 2012, 189, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.; Apopa, P.L.; Vesci, J.; Kenyon, V.; Rai, G.; Jadhav, A.; Simeonov, A.; Holman, T.R.; Maloney, D.J.; Boutaud, O.; et al. Protein Kinase C Regulation of 12-Lipoxygenase-Mediated Human Platelet Activation. Mol. Pharmacol. 2011, 81, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.; Chen, X.S.; Takahashi, Y.; Yamamoto, S.; Funk, C.D.; Serhan, C.N. Lipoxin synthase activity of human platelet 12-lipoxygenase. Biochem. J. 1993, 296, 127–133. [Google Scholar] [CrossRef]
- Serhan, C.N.; Jain, A.; Marleau, S.; Clish, C.B.; Kantarci, A.; Behbehani, B.; Colgan, S.P.; Stahl, G.L.; Merched, A.; Petasis, N.A.; et al. Reduced Inflammation and Tissue Damage in Transgenic Rabbits Overexpressing 15-Lipoxygenase and Endogenous Anti-inflammatory Lipid Mediators. J. Immunol. 2003, 171, 6856–6865. [Google Scholar] [CrossRef]
- Gronert, K.; Maheshwari, N.; Khan, N.; Hassan, I.R.; Dunn, M.; Schwartzman, M.L. A role for the mouse 12/15-lipoxygenase pathway in promoting epithelial wound healing and host defense. J. Biol. Chem. 2005, 280, 15267–15278. [Google Scholar] [CrossRef]
- Carion, T.W.; Kracht, D.; Strand, E.; David, E.; McWhirter, C.; Ebrahim, A.S.; Berger, E. VIP modulates the ALX/FPR2 receptor axis toward inflammation resolution in a mouse model of bacterial keratitis. Prostaglandins Other Lipid Mediat. 2019, 140, 18–25. [Google Scholar] [CrossRef]
- Chiang, N.; De La Rosa, X.; Libreros, S.; Serhan, C.N. Novel Resolvin D2 Receptor Axis in Infectious Inflammation. J. Immunol. 2016, 198, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Jean, D.; Proske, R.J.; Reins, R.Y.; McDermott, A.M. Ocular Surface Expression andIn VitroActivity of Antimicrobial Peptides. Curr. Eye Res. 2007, 32, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Wheater, M.K.; Qiu, Y.; Sosne, G. Thymosin beta4 inhibits TNF-alpha-induced NF-kappaB activation, IL-8 expression, and the sensitizing effects by its partners PINCH-1 and ILK. FASEB J. 2011, 25, 1815–1826. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Qiu, P.; Christopherson, P.L.; Wheater, M.K. Thymosin beta 4 suppression of corneal NFkappaB: A potential anti-inflammatory pathway. Exp. Eye Res. 2007, 84, 663–669. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Background | Function |
|---|---|---|
| LL-37/CAMP [17,18] | LL-37/CAMP is a 37 amino acid AMP that is produced in many cells including macrophages, secondary neutrophils, NK cells, and select epithelial cells. | LL-37/CAMP has been shown to have many functions including antimicrobial activity and as a chemoattractant. It also possesses anti-inflammatory properties, modulates TLRs, and regulates IFN. |
| S100A8/A9 [19,20] | S100A8/A9 are Ca2+ binding proteins that are generally present in their heterodimeric form, which is called calprotectin. These AMPs are constitutively present in PMNs and monocytes and are heavily upregulated during trauma and inflammation. | The active release of S100A8/A9 during inflammation helps induce cytokine release and promote leukocyte recruitment. S100 proteins are often involved in the regulation of cell cycle progression and differentiation. When S100A8 complexes with S100A14, they work to regulate myeloid cell function by binding to TLR4. |
| hBD-1/-2/-3 [21,22] | Defensins are 2–6 kDa, cationic, microbicidal AMPs containing three pairs of intramolecular disulfide bonds. | Human beta defensins (hBDs) often work synergistically to promote bactericidal and bacteriostatic activity during infection. Specifically, they promote chemotaxis, activation, and degranulation of mast cells during inflammation. |
| Keratin 6A [23] | Keratin 6A codes for a 564 amino acid, 60 kDa AMP. It is a type II cytokeratin, one of a number of isoforms of keratin 6 encoded by separate genes from the gene cluster on human chromosome 12q. | Epidermis-specific keratin is involved in wound healing and shown to have antimicrobial properties. It is the main antimicrobial factor in the eye. It is involved in the activation of follicular keratinocytes after wounding, although it does not play a major role in keratinocyte proliferation or migration. |
| TLR4 [24,25] | TLR4 is a 95 kDa transmembrane protein of the Toll-like receptor family. | They recognize PAMPs on microbes and activates the NF-κB pathway of inflammation and the innate immune response. Cooperates with LY96 and CD14 to mediate the innate immune response to gram negative bacterial lipopolysaccharide (LPS). |
| 12-LOX [26,27] | 12-LOX is a 75 kDa enzyme composed of 663 amino acids. This protein is a lipoxygenase-type enzyme that is coded by the ALOX12 gene on chromosome 17p13.3. | Regulation of 12-LOX principally relies on the availability of its polyunsaturated fatty acids (PUFAs), which are released from lipid membranes during inflammation. This enzyme participates in arachidonic acid metabolism which can result in the activation of specialized pro-resolving molecules (SPMs). |
| 15-LOX [28,29] | 15-LOX is a 75 kDa enzyme composed of 662 amino acids. This protein is a lipoxygenase-type enzyme that is coded by the ALOX15 gene on chromosome 17p13.3. | 15-LOX functions to metabolize PUFAs into SPMs that facilitate inflammation resolution. |
| ALX/FPR2 [30] | The ALX/FPR2 receptor is a 351 amino acid receptor that is coded for by the FPR2 gene located on chromosome 19q.13.3. | FPR2 is G protein-coupled cell surface receptor. One of the primary functions of FPR2 is its binding of SPMs, which help mediate a pro-resolving response. |
| DRV2/GPR18 [31] | The DRV2/GPR18 receptor is a 38 kDa protein composed of 331 amino acids. This receptor is coded for by the GPR18 gene located on chromosome 13q32.3. | GPR18 is a G protein-coupled receptor for endogenous lipid neurotransmitters. Resolvin D2, a DHA-derived SPM, is also an activating ligand for the GPR18 receptor. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carion, T.W.; Ebrahim, A.S.; Alluri, S.; Ebrahim, T.; Parker, T.; Burns, J.; Sosne, G.; Berger, E.A. Antimicrobial Effects of Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy in Pseudomonas aeruginosa Induced Keratitis. Int. J. Mol. Sci. 2020, 21, 6840. https://doi.org/10.3390/ijms21186840
Carion TW, Ebrahim AS, Alluri S, Ebrahim T, Parker T, Burns J, Sosne G, Berger EA. Antimicrobial Effects of Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy in Pseudomonas aeruginosa Induced Keratitis. International Journal of Molecular Sciences. 2020; 21(18):6840. https://doi.org/10.3390/ijms21186840
Chicago/Turabian StyleCarion, Thomas W., Abdul Shukkur Ebrahim, Spandana Alluri, Thanzeela Ebrahim, Tressa Parker, Julia Burns, Gabriel Sosne, and Elizabeth A. Berger. 2020. "Antimicrobial Effects of Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy in Pseudomonas aeruginosa Induced Keratitis" International Journal of Molecular Sciences 21, no. 18: 6840. https://doi.org/10.3390/ijms21186840
APA StyleCarion, T. W., Ebrahim, A. S., Alluri, S., Ebrahim, T., Parker, T., Burns, J., Sosne, G., & Berger, E. A. (2020). Antimicrobial Effects of Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy in Pseudomonas aeruginosa Induced Keratitis. International Journal of Molecular Sciences, 21(18), 6840. https://doi.org/10.3390/ijms21186840