Structural Insights into the Mechanism of a Nanobody That Stabilizes PAI-1 and Modulates Its Activity


Abstract
1. Introduction
2. Results
2.1. Crystallographic Analysis of the PAI-1/Nb93 Complex
2.2. Nb93 Binds to the Surface-Exposed RCL and Thereby Directly Competes for the PA Binding Site on PAI-1
2.3. The Structure of the PAI-1/Nb93 Complex Supports Previously Obtained Data on the Affinity Profile of Nb93
2.4. The X-Ray Structure Is Valid in Solution
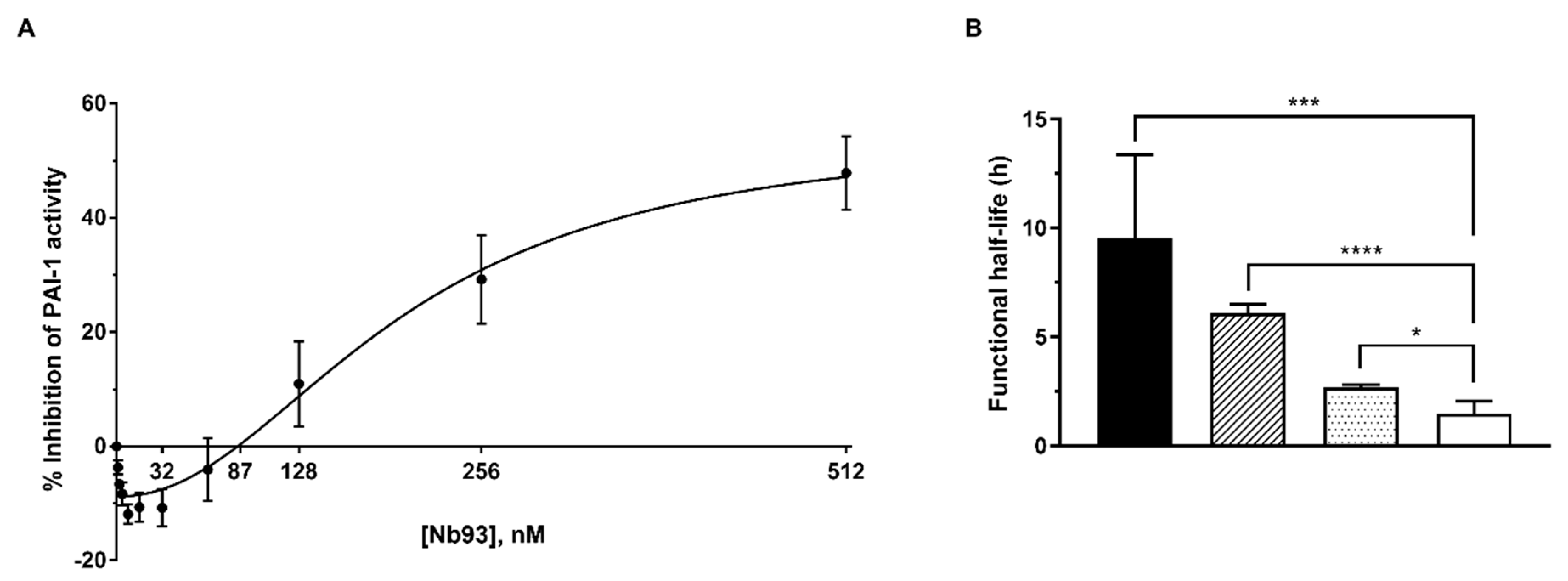
2.5. Nb93 Exhibits a Biphasic Effect on the Inhibition of PAI-1 Activity Due to a Substantial Stabilization of the Active PAI-1 Conformation
3. Discussion
4. Materials and Methods
4.1. Cloning, Expression, and Purification of PAI-1 Variants and Nb93
4.2. Crystallization and Data Collection
4.3. Structure Determination, Refinement, and Analysis
4.4. Size Exclusion Chromatography with Inline Small-Angle X-Ray Scattering
4.5. Activity Profile of Nb93
4.6. Functional Half-Life of PAI-1
4.7. Quantification and Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kruithof, E.K.; Tran-Thang, C.; Ransijn, A.; Bachmann, F. Demonstration of a fast-acting inhibitor of plasminogen activators in human plasma. Blood 1984, 64, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Gettins, P.G. Serpin structure, mechanism, and function. Chem Rev. 2002, 102, 4751–4804. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Liu, M.; Zeng, T.; Shi, X.; Yuan, C.; Andreasen, P.A.; Huang, M. Crystal structure of the michaelis complex between tissue-type plasminogen activator and plasminogen activators inhibitor-1. J. Biol Chem. 2015, 290, 25795–25804. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Jiang, L.; Yuan, C.; Jensen, J.K.; Zhang, X.; Luo, Z.; Furie, B.C.; Furie, B.; Andreasen, P.A.; Huang, M. Structural basis for recognition of urokinase-type plasminogen activator by plasminogen activator inhibitor-1. J. Biol Chem. 2011, 286, 7027–7032. [Google Scholar] [CrossRef] [PubMed]
- van Meijer, M.; Smilde, A.; Tans, G.; Nesheim, M.E.; Pannekoek, H.; Horrevoets, A.J. The suicide substrate reaction between plasminogen activator inhibitor 1 and thrombin is regulated by the cofactors vitronectin and heparin. Blood 1997, 90, 1874–1882. [Google Scholar] [CrossRef]
- Declerck, P.J.; De Mol, M.; Alessi, M.C.; Baudner, S.; Pâques, E.P.; Preissner, K.T.; Müller-Berghaus, G.; Collen, D. Purification and characterization of a plasminogen activator inhibitor 1 binding protein from human plasma. Identification as a multimeric form of S protein (vitronectin). J. Biol. Chem. 1988, 263, 15454–15461. [Google Scholar]
- Lindahl, T.L.; Sigurdardottir, O.; Wiman, B. Stability of plasminogen activator inhibitor 1 (PAI-1). Thromb. Haemost. 1989, 62, 748–751. [Google Scholar] [CrossRef]
- Meltzer, M.E.; Lisman, T.; de Groot, P.G.; Meijers, J.C.; le Cessie, S.; Doggen, C.J.; Rosendaal, F.R. Venous thrombosis risk associated with plasma hypofibrinolysis is explained by elevated plasma levels of TAFI and PAI-1. Blood 2010, 116, 113–121. [Google Scholar] [CrossRef]
- Thögersen, A.M.; Jansson, J.H.; Boman, K.; Nilsson, T.K.; Weinehall, L.; Huhtasaari, F.; Hallmans, G. High plasminogen activator inhibitor and tissue plasminogen activator levels in plasma precede a first acute myocardial infarction in both men and women: Evidence for the fibrinolytic system as an independent primary risk factor. Circulation 1998, 98, 2241–2247. [Google Scholar] [CrossRef]
- Margaglione, M.; Di Minno, G.; Grandone, E.; Vecchione, G.; Celentano, E.; Cappucci, G.; Grilli, M.; Simone, P.; Panico, S.; Mancini, M. Abnormally high circulation levels of tissue plasminogen activator and plasminogen activator inhibitor-1 in patients with a history of ischemic stroke. Arterioscler. Thromb. 1994, 14, 1741–1745. [Google Scholar] [CrossRef]
- Izuhara, Y.; Yamaoka, N.; Kodama, H.; Dan, T.; Takizawa, S.; Hirayama, N.; Meguro, K.; de Strihou, C.v.; Miyata, T. A novel inhibitor of plasminogen activator inhibitor-1 provides antithrombotic benefits devoid of bleeding effect in nonhuman primates. J. Cereb. Blood Flow Metab. 2010, 30, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Van De Craen, B.; Scroyen, I.; Abdelnabi, R.; Brouwers, E.; Lijnen, H.R.; Declerck, P.J.; Gils, A. Characterization of a panel of monoclonal antibodies toward mouse PAI-1 that exert a significant profibrinolytic effect in vivo. Thromb. Res. 2011, 128, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Wyseure, T.; Rubio, M.; Denorme, F.; Martinez de Lizarrondo, S.; Peeters, M.; Gils, A.; De Meyer, S.F.; Vivien, D.; Declerck, P.J. Innovative thrombolytic strategy using a heterodimer diabody against TAFI and PAI-1 in mouse models of thrombosis and stroke. Blood 2015, 125, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Florova, G.; Azghani, A.; Karandashova, S.; Schaefer, C.; Koenig, K.; Stewart-Evans, K.; Declerck, P.J.; Idell, S.; Komissarov, A.A. Targeting of plasminogen activator inhibitor 1 improves fibrinolytic therapy for tetracycline-induced pleural injury in rabbits. Am. J. Respir Cell Mol. Biol. 2015, 52, 429–437. [Google Scholar] [CrossRef]
- Florova, G.; Azghani, A.O.; Karandashova, S.; Schaefer, C.; Yarovoi, S.V.; Declerck, P.J.; Cines, D.B.; Idell, S.; Komissarov, A.A. Targeting plasminogen activator inhibitor-1 in tetracycline-induced pleural injury in rabbits. Am. J. Physiol Lung Cell Mol. Physiol. 2018, 314, L54–L68. [Google Scholar] [CrossRef]
- Placencio, V.R.; DeClerck, Y.A. Plasminogen activator inhibitor-1 in cancer: Rationale and insight for future therapeutic testing. Cancer Res. 2015, 75, 2969–2974. [Google Scholar] [CrossRef]
- Rabieian, R.; Boshtam, M.; Zareei, M.; Kouhpayeh, S.; Masoudifar, A.; Mirzaei, H. Plasminogen activator inhibitor type-1 as a regulator of fibrosis. J. Cell Biochem. 2018, 119, 17–27. [Google Scholar] [CrossRef]
- Sillen, M.; Weeks, S.D.; Zhou, X.; Komissarov, A.A.; Florova, G.; Idell, S.; Strelkov, S.V.; Declerck, P.J. Molecular mechanism of two nanobodies that inhibit PAI-1 activity reveals a modulation at distinct stages of the PAI-1/plasminogen activator interaction. J. Thromb. Haemost. 2020, 18, 681–692. [Google Scholar] [CrossRef]
- Vousden, K.A.; Lundqvist, T.; Popovic, B.; Naiman, B.; Carruthers, A.M.; Newton, P.; Johnson, D.J.D.; Pomowski, A.; Wilkinson, T.; Dufner, P.; et al. Discovery and characterisation of an antibody that selectively modulates the inhibitory activity of plasminogen activator inhibitor-1. Sci Rep. 2019, 9, 1605. [Google Scholar] [CrossRef]
- Bijnens, A.P.; Gils, A.; Stassen, J.M.; Komissarov, A.A.; Knockaert, I.; Brouwers, E.; Shore, J.D.; Declerck, P.J. The distal hinge of the reactive site loop and its proximity: A target to modulate plasminogen activator inhibitor-1 activity. J. Biol. Chem. 2001, 276, 44912–44918. [Google Scholar] [CrossRef]
- Komissarov, A.A.; Declerck, P.J.; Shore, J.D. Mechanisms of conversion of plasminogen activator inhibitor 1 from a suicide inhibitor to a substrate by monoclonal antibodies. J. Biol. Chem. 2002, 277, 43858–43865. [Google Scholar] [CrossRef] [PubMed]
- Verhamme, I.; Kvassman, J.O.; Day, D.; Debrock, S.; Vleugels, N.; Declerck, P.J.; Shore, J.D. Accelerated conversion of human plasminogen activator inhibitor-1 to its latent form by antibody binding. J. Biol. Chem. 1999, 274, 17511–17517. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Hendrickx, M.L.; Hassanzadeh-Ghassabeh, G.; Muyldermans, S.; Declerck, P.J. Generation and in vitro characterisation of inhibitory nanobodies towards plasminogen activator inhibitor 1. Thromb Haemost. 2016, 116, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Conrath, K.E.; Wernery, U.; Muyldermans, S.; Nguyen, V.K. Emergence and evolution of functional heavy-chain antibodies in Camelidae. Dev. Comp. Immunol. 2003, 27, 87–103. [Google Scholar] [CrossRef]
- Zhou, X.; Weeks, S.D.; Ameloot, P.; Callewaert, N.; Strelkov, S.V.; Declerck, P.J. Elucidation of the molecular mechanisms of two nanobodies that inhibit thrombin-activatable fibrinolysis inhibitor activation and activated thrombin-activatable fibrinolysis inhibitor activity. J. Thromb. Haemost. 2016, 14, 1629–1638. [Google Scholar] [CrossRef] [PubMed]
- Aertgeerts, K.; De Bondt, H.L.; De Ranter, C.J.; Declerck, P.J. Mechanisms contributing to the conformational and functional flexibility of plasminogen activator inhibitor-1. Nat. Struct. Biol. 1995, 2, 891–897. [Google Scholar] [CrossRef]
- Jung, R.G.; Motazedian, P.; Ramirez, F.D.; Simard, T.; Di Santo, P.; Visintini, S.; Faraz, M.A.; Labinaz, A.; Jung, Y.; Hibbert, B. Association between plasminogen activator inhibitor-1 and cardiovascular events: A systematic review and meta-analysis. Thromb. J. 2018, 16, 12. [Google Scholar] [CrossRef]
- Flevaris, P.; Vaughan, D. The role of plasminogen activator inhibitor type-1 in fibrosis. Semin. Thromb. Hemost. 2017, 43, 169–177. [Google Scholar] [CrossRef]
- Li, S.H.; Reinke, A.A.; Sanders, K.L.; Emal, C.D.; Whisstock, J.C.; Stuckey, J.A.; Lawrence, D.A. Mechanistic characterization and crystal structure of a small molecule inactivator bound to plasminogen activator inhibitor-1. Proc. Natl. Acad. Sci. USA 2013, 110, E4941–E4949. [Google Scholar] [CrossRef]
- Yarmolinsky, J.; Bordin Barbieri, N.; Weinmann, T.; Ziegelmann, P.K.; Duncan, B.B.; Inês Schmidt, M. Plasminogen activator inhibitor-1 and type 2 diabetes: A systematic review and meta-analysis of observational studies. Sci. Rep. 2016, 6, 17714. [Google Scholar] [CrossRef]
- Fortenberry, Y.M. Plasminogen activator inhibitor-1 inhibitors: A patent review (2006-present). Expert Opin. Ther. Pat. 2013, 23, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Bartha, K.; Declerck, P.J.; Moreau, H.; Nelles, L.; Collen, D. Synthesis and secretion of plasminogen activator inhibitor 1 by human endothelial cells in vitro. Effect of active site mutagenized tissue-type plasminogen activator. J. Biol. Chem. 1991, 266, 792–797. [Google Scholar] [PubMed]
- Götzke, H.; Kilisch, M.; Martínez-Carranza, M.; Sograte-Idrissi, S.; Rajavel, A.; Schlichthaerle, T.; Engels, N.; Jungmann, R.; Stenmark, P.; Opazo, F.; et al. The ALFA-tag is a highly versatile tool for nanobody-based bioscience applications. Nat. Commun. 2019, 10, 4403. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.B.; Traenkle, B.; Koch, P.A.; Emele, F.; Weiss, F.; Poetz, O.; Stehle, T.; Rothbauer, U. Peptides in headlock—A novel high-affinity and versatile peptide-binding nanobody for proteomics and microscopy. Sci. Rep. 2016, 6, 19211. [Google Scholar] [CrossRef] [PubMed]
- Verheesen, P.; ten Haaft, M.R.; Lindner, N.; Verrips, C.T.; de Haard, J.J.W. Beneficial properties of single-domain antibody fragments for application in immunoaffinity purification and immuno-perfusion chromatography. Biochim. Biophys. Acta 2003, 1624, 21–28. [Google Scholar] [CrossRef]
- Tereshko, V.; Uysal, S.; Koide, A.; Margalef, K.; Koide, S.; Kossiakoff, A.A. Toward chaperone-assisted crystallography: Protein engineering enhancement of crystal packing and X-ray phasing capabilities of a camelid single-domain antibody (VHH) scaffold. Protein Sci. 2008, 17, 1175–1187. [Google Scholar] [CrossRef]
- Weeks, S.D.; Drinker, M.; Loll, P.J. Ligation independent cloning vectors for expression of SUMO fusions. Protein Expr. Purif. 2007, 53, 40–50. [Google Scholar] [CrossRef]
- Vonrhein, C.; Flensburg, C.; Keller, P.; Sharff, A.; Smart, O.; Paciorek, W.; Womack, T.; Bricogne, G. Data processing and analysis with the autoPROC toolbox. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 293–302. [Google Scholar] [CrossRef]
- Jensen, J.K.; Thompson, L.C.; Bucci, J.C.; Nissen, P.; Gettins, P.G.; Peterson, C.B.; Andreasen, P.A.; Morth, J.P. Crystal structure of plasminogen activator inhibitor-1 in an active conformation with normal thermodynamic stability. J. Biol. Chem. 2011, 286, 29709–29717. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef]
- Rossey, I.; Gilman, M.S.; Kabeche, S.C.; Sedeyn, K.; Wrapp, D.; Kanekiyo, M.; Chen, M.; Mas, V.; Spitaels, J.; Melero, J.A.; et al. Potent single-domain antibodies that arrest respiratory syncytial virus fusion protein in its prefusion state. Nat. Commun. 2017, 8, 14158. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Svergun, D.; Barberato, C.; Koch, M.H.J. CRYSOL–A program to evaluate X-ray solution scattering of biological macromolecules from atomic coordinates. J. Appl. Crystallogr. 1995, 28, 768–773. [Google Scholar] [CrossRef]
- Rambo, R.P.; Tainer, J.A. Characterizing flexible and intrinsically unstructured biological macromolecules by SAS using the Porod-Debye law. Biopolymers 2011, 95, 559–571. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef]
- Verheijen, J.H.; Chang, G.T.; Kluft, C. Evidence for the occurrence of a fast-acting inhibitor for tissue-type plasminogen activator in human plasma. Thromb. Haemost. 1984, 51, 392–395. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAI-1-W175F/Nb93 | |||
|---|---|---|---|
| Data Collection | Refinement | ||
| Space group | P 41 21 2 | No. of complexes/ASU | 1 |
| Cell dimensions | Reflections used in refinement | 55,639 (5474) | |
| a, b, c (Å) | 117.81, 117.81, 96.48 | Reflections used for Rfree | 2315 (234) |
| α, β, γ (°) | 90, 90, 90 | Rwork | 0.197 |
| Resolution (Å) | 52.69–1.88 (1.95–1.88) | Rfree | 0.233 |
| Rmerge | 0.115 (1.326) | No. of non-hydrogen atoms | 4098 |
| I/σ(I) | 12.6 (2.0) | Protein | 3783 |
| Wilson B-factor (Å2) | 30.88 | Water | 315 |
| CC1/2 | 0.998 (0.839) | Average B-factors (Å2) | |
| Completeness (%) | 100 (100) | Protein | 36.26 |
| Redundancy | 13.0 (13.1) | Water | 41.16 |
| R.m.s. deviations | |||
| Bond lengths (Å) | 0.007 | ||
| Bond angles (°) | 0.79 | ||
| PAI-1-W175F/Nb93 | |||
|---|---|---|---|
| Monomer | Nb93 | PAI-1 | Distance (Å) |
| Hydrogen bonds | Gln39 [HE22]* | Ser337 [OG] | 2.2 |
| Glu44 [OE1]* | Asp181 [H] | 2.1 | |
| Glu44 [OE2]* | Ser182 [H] | 2.0 | |
| Glu44 [OE2]* | Ser182 [HG] | 2.0 | |
| Arg45 [O]* | Ile342 [H] | 2.0 | |
| Arg45 [HE]* | Ser337 [O] | 2.8 | |
| Gly47 [H]* | Ile342 [O] | 1.9 | |
| Asp62 [OD1] | Ser344 [HG] | 2.1 | |
| Trp104 [HE1] | Glu350 [OE1 ] | 2.0 | |
| Thr106 [O] | Thr205 [HG1] | 2.2 | |
| Asn108 [HD22] | Thr205 [O] | 2.0 | |
| Asn108 [HD21] | Val341 [O] | 2.2 | |
| Asn108 [OD1] | Val341 [H] | 1.9 | |
| Salt bridges | Asp62 [OD2] | Arg346 [NH1] | 2.8 |
| Glu65 [OE2] | Arg346 [NE] | 3.9 | |
| Asp99 [OD1] | Lys207 [NZ] | 2.8 | |
| Asp99 [OD2] | Lys207 [NZ] | 2.7 | |
| Hydrophobic contacts a | Phe37, Gly47, Cys50, Tyr59, Thr61, Trp104, Cys105, Phe107 | Thr205, Pro270, Val341, Ile342, Val343, Met347, Ala348, Pro349 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sillen, M.; Weeks, S.D.; Strelkov, S.V.; Declerck, P.J. Structural Insights into the Mechanism of a Nanobody That Stabilizes PAI-1 and Modulates Its Activity. Int. J. Mol. Sci. 2020, 21, 5859. https://doi.org/10.3390/ijms21165859
Sillen M, Weeks SD, Strelkov SV, Declerck PJ. Structural Insights into the Mechanism of a Nanobody That Stabilizes PAI-1 and Modulates Its Activity. International Journal of Molecular Sciences. 2020; 21(16):5859. https://doi.org/10.3390/ijms21165859
Chicago/Turabian StyleSillen, Machteld, Stephen D. Weeks, Sergei V. Strelkov, and Paul J. Declerck. 2020. "Structural Insights into the Mechanism of a Nanobody That Stabilizes PAI-1 and Modulates Its Activity" International Journal of Molecular Sciences 21, no. 16: 5859. https://doi.org/10.3390/ijms21165859
APA StyleSillen, M., Weeks, S. D., Strelkov, S. V., & Declerck, P. J. (2020). Structural Insights into the Mechanism of a Nanobody That Stabilizes PAI-1 and Modulates Its Activity. International Journal of Molecular Sciences, 21(16), 5859. https://doi.org/10.3390/ijms21165859