Association of Maternal Regulatory Single Nucleotide Polymorphic CD99 Genotype with Preeclampsia in Pregnancies Carrying Male Fetuses in Ethiopian Women

 ,
, 
Abstract
1. Introduction
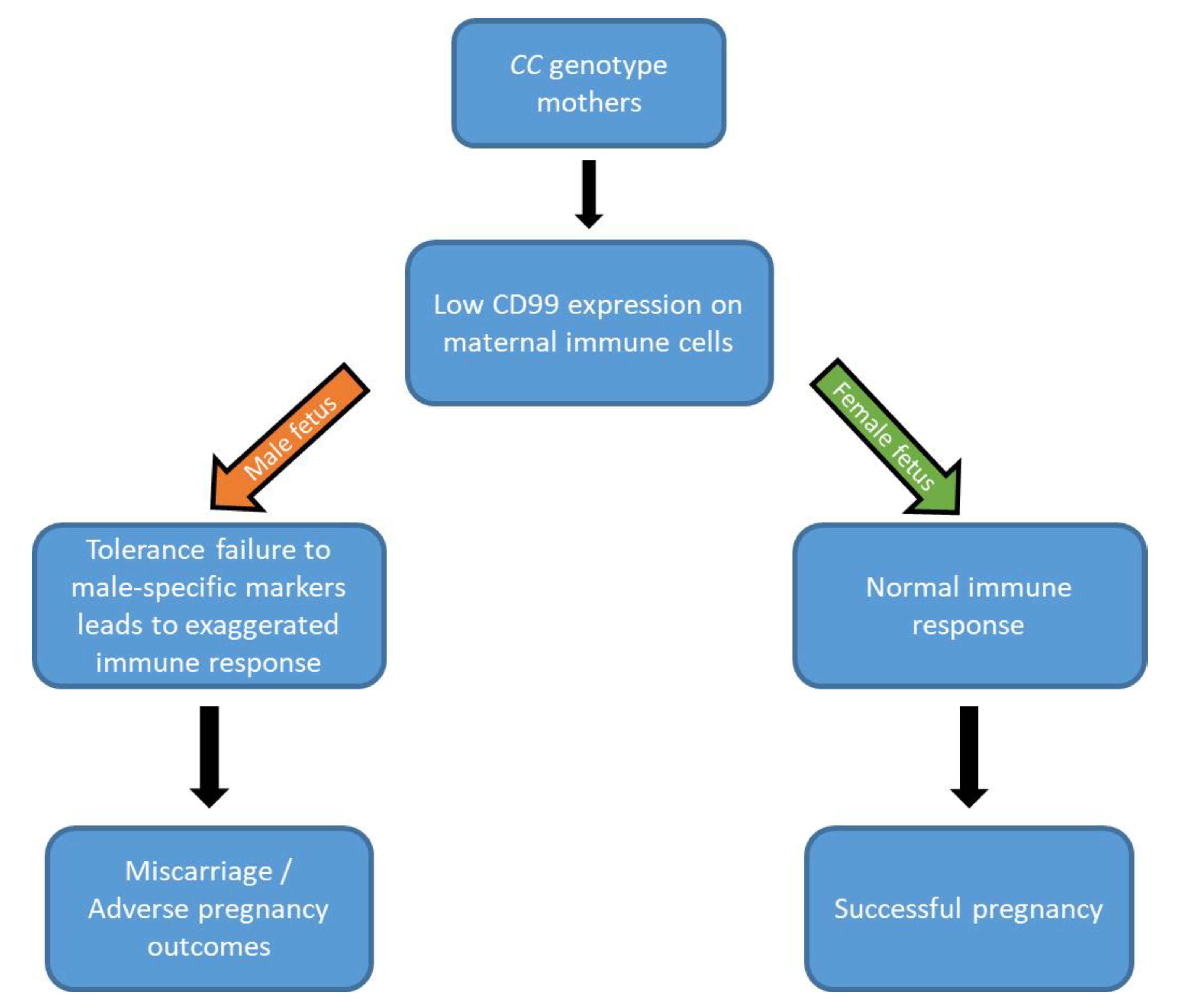
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Design and Study Participants
4.2. Ethical Considerations
4.3. Inclusion Criteria
4.4. DNA Extraction and Genotyping
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
Abbreviations
| PE | preeclampsia |
| GA | gestational age |
| sBP | systolic blood pressure |
| dBP | diastolic blood pressure |
| SNP | single nucleotide polymorphism |
| uNK | uterine natural killer cells |
| Th | T helper cells |
References
- Hogan, M.C.; Foreman, K.J.; Naghavi, M.; Ahn, S.Y.; Wang, M.; Makela, S.M.; Lopez, A.D.; Lozano, R.; Murray, C.J. Maternal mortality for 181 countries, 1980–2008: A systematic analysis of progress towards Millennium Development Goal 5. Lancet 2010, 375, 1609–1623. [Google Scholar] [CrossRef]
- Ghulmiyyah, L.; Sibai, B. Maternal mortality from preeclampsia/eclampsia. Semin. Perinatol. 2012, 36, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Duley, L. The global impact of pre-eclampsia and eclampsia. Semin. Perinatol. 2009, 33, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. The hypertensive disorders of pregnancy: ISSHP classification, diagnosis & management recommendations for international practice. Pregnancy Hypertens. 2018, 13, 291–310. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.; Salonen Ros, H.; Cnattingius, S.; Lichtenstein, P. The importance of genetic and environmental effects for pre-eclampsia and gestational hypertension: A family study. BJOG Int. J. Obstet. Gynaecol. 2004, 111, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Erlandsson, L.; Lindgren, R.; Naav, A.; Krais, A.M.; Strandberg, B.; Lundh, T.; Boman, C.; Isaxon, C.; Hansson, S.R.; Malmqvist, E. Exposure to wood smoke particles leads to inflammation, disrupted proliferation and damage to cellular structures in a human first trimester trophoblast cell line. Envion. Poll. 2020, 264, 114790. [Google Scholar] [CrossRef]
- Roberts, J.M.; Hubel, C.A. The two stage model of preeclampsia: Variations on the theme. Placenta 2009, 30 (Suppl. A), S32–S37. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I.L. Latest advances in understanding preeclampsia. Science 2005, 308, 1592–1594. [Google Scholar] [CrossRef]
- Magee, L.A.; Pels, A.; Helewa, M.; Rey, E.; von Dadelszen, P. Diagnosis, evaluation, and management of the hypertensive disorders of pregnancy. Pregnancy Hypertens. 2014, 4, 105–145. [Google Scholar] [CrossRef]
- Bell, M.J. A historical overview of preeclampsia-eclampsia. J. Obs. Gynecol. Neonatal. Nurs. 2010, 39, 510–518. [Google Scholar] [CrossRef]
- Anderson, U.D.; Gram, M.; Akerstrom, B.; Hansson, S.R. First trimester prediction of preeclampsia. Curr. Hypertens. Rep. 2015, 17, 584. [Google Scholar] [CrossRef] [PubMed]
- Lagana, A.S.; Favilli, A.; Triolo, O.; Granese, R.; Gerli, S. Early serum markers of pre-eclampsia: Are we stepping forward? J. Matern. Fetal. Neonatal. Med. 2016, 29, 3019–3023. [Google Scholar] [CrossRef] [PubMed]
- Lagana, A.S.; Giordano, D.; Loddo, S.; Zoccali, G.; Vitale, S.G.; Santamaria, A.; Buemi, M.; D’Anna, R. Decreased Endothelial Progenitor Cells (EPCs) and increased Natural Killer (NK) cells in peripheral blood as possible early markers of preeclampsia: A case-control analysis. Arch. Gynecol. Obstet. 2017, 295, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, R. The male/female ratio of fetal deaths and births in Japan. Lancet 2000, 356, 738–739. [Google Scholar] [CrossRef]
- Al-Qaraghouli, M.; Fang, Y.M.V. Effect of Fetal Sex on Maternal and Obstetric Outcomes. Front. Pediatr. 2017, 5, 144. [Google Scholar] [CrossRef]
- Masoumi, Z.; Maes, G.E.; Herten, K.; Cortes-Calabuig, A.; Alattar, A.G.; Hanson, E.; Erlandsson, L.; Mezey, E.; Magnusson, M.; Vermeesch, J.R.; et al. Preeclampsia is Associated with Sex-Specific Transcriptional and Proteomic Changes in Fetal Erythroid Cells. Int. J. Mol. Sci. 2019, 20, 38. [Google Scholar] [CrossRef]
- Ellis, N.A.; Ye, T.Z.; Patton, S.; German, J.; Goodfellow, P.N.; Weller, P. Cloning of PBDX, an MIC2-related gene that spans the pseudoautosomal boundary on chromosome Xp. Nat. Genet. 1994, 6, 394–400. [Google Scholar] [CrossRef]
- Reid, M.E.; Lomas-Francis, C.; Olsson, M.L. The Blood Group Antigen Factsbook, 3rd ed.; Elsevier/AP: Amsterdam, The Netherlands, 2012; p. 745. ISBN 9780124158498. [Google Scholar]
- Gelin, C.; Aubrit, F.; Phalipon, A.; Raynal, B.; Cole, S.; Kaczorek, M.; Bernard, A. The E2 antigen, a 32 kd glycoprotein involved in T-cell adhesion processes, is the MIC2 gene product. EMBO J. 1989, 8, 3253–3259. [Google Scholar] [CrossRef]
- Pasello, M.; Manara, M.C.; Scotlandi, K. CD99 at the crossroads of physiology and pathology. J. Cell Commun. Signal. 2018, 12, 55–68. [Google Scholar] [CrossRef]
- Pettersen, R.D.; Bernard, G.; Olafsen, M.K.; Pourtein, M.; Lie, S.O. CD99 signals caspase-independent T cell death. J. Immunol. (Baltim. Md. 1950) 2001, 166, 4931–4942. [Google Scholar] [CrossRef]
- Bernard, G.; Breittmayer, J.P.; de Matteis, M.; Trampont, P.; Hofman, P.; Senik, A.; Bernard, A. Apoptosis of immature thymocytes mediated by E2/CD99. J. Immunol. (Baltim. Md. 1950) 1997, 158, 2543–2550. [Google Scholar]
- Sciandra, M.; Marino, M.T.; Manara, M.C.; Guerzoni, C.; Grano, M.; Oranger, A.; Lucarelli, E.; Lollini, P.L.; Dozza, B.; Pratelli, L.; et al. CD99 drives terminal differentiation of osteosarcoma cells by acting as a spatial regulator of ERK 1/2. J. Bone Min. Res. 2014, 29, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Imbert, A.M.; Belaaloui, G.; Bardin, F.; Tonnelle, C.; Lopez, M.; Chabannon, C. CD99 expressed on human mobilized peripheral blood CD34+ cells is involved in transendothelial migration. Blood 2006, 108, 2578–2586. [Google Scholar] [CrossRef] [PubMed]
- Dufour, E.M.; Deroche, A.; Bae, Y.; Muller, W.A. CD99 is essential for leukocyte diapedesis in vivo. Cell Commun. Adhes. 2008, 15, 351–363. [Google Scholar] [CrossRef]
- Lou, O.; Alcaide, P.; Luscinskas, F.W.; Muller, W.A. CD99 is a key mediator of the transendothelial migration of neutrophils. J. Immunol. (Baltim. Md. 1950) 2007, 178, 1136–1143. [Google Scholar] [CrossRef]
- Wilson, M.D.; Cheung, J.; Martindale, D.W.; Scherer, S.W.; Koop, B.F. Comparative analysis of the paired immunoglobulin-like receptor (PILR) locus in six mammalian genomes: Duplication, conversion, and the birth of new genes. Physiol. Genom. 2006, 27, 201–218. [Google Scholar] [CrossRef]
- Takheaw, N.; Earwong, P.; Laopajon, W.; Pata, S.; Kasinrerk, W. Interaction of CD99 and its ligand upregulates IL-6 and TNF-alpha upon T cell activation. PLoS ONE 2019, 14, e0217393. [Google Scholar] [CrossRef]
- Petit, C.; Levilliers, J.; Weissenbach, J. Physical mapping of the human pseudo-autosomal region; comparison with genetic linkage map. EMBO J. 1988, 7, 2369–2376. [Google Scholar] [CrossRef]
- Goodfellow, P.N.; Pym, B.; Pritchard, C.; Ellis, N.; Palmer, M.; Smith, M.; Goodfellow, P.J. MIC2: A human pseudoautosomal gene. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1988, 322, 145–154. [Google Scholar] [CrossRef]
- Moller, M.; Lee, Y.Q.; Vidovic, K.; Kjellstrom, S.; Bjorkman, L.; Storry, J.R.; Olsson, M.L. Disruption of a GATA1-binding motif upstream of XG/PBDX abolishes Xg(a) expression and resolves the Xg blood group system. Blood 2018, 132, 334–338. [Google Scholar] [CrossRef]
- Yeh, C.C.; Chang, C.J.; Twu, Y.C.; Chu, C.C.; Liu, B.S.; Huang, J.T.; Hung, S.T.; Chan, Y.S.; Tsai, Y.J.; Lin, S.W.; et al. The molecular genetic background leading to the formation of the human erythroid-specific Xg(a)/CD99 blood groups. Blood Adv. 2018, 2, 1854–1864. [Google Scholar] [CrossRef] [PubMed]
- Darlison, M. Investigating the Molecular Genetic Basis of the Xg Blood Group System and Its Red Blood Cell Phenotypes. Master’s Thesis, Lund University, Lund, Sweden, 2018. [Google Scholar]
- Gao, J.; Chen, Y.H.; Peterson, L.C. GATA family transcriptional factors: Emerging suspects in hematologic disorders. Exp. Hematol. Oncol. 2015, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Tindemans, I.; Serafini, N.; Di Santo, J.P.; Hendriks, R.W. GATA-3 function in innate and adaptive immunity. Immunity 2014, 41, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Skapenko, A.; Leipe, J.; Niesner, U.; Devriendt, K.; Beetz, R.; Radbruch, A.; Kalden, J.R.; Lipsky, P.E.; Schulze-Koops, H. GATA-3 in human T cell helper type 2 development. J. Exp. Med. 2004, 199, 423–428. [Google Scholar] [CrossRef]
- Elsmen, E.; Kallen, K.; Marsal, K.; Hellstrom-Westas, L. Fetal gender and gestational-age-related incidence of pre-eclampsia. Acta Obstet. Gynecol. Scand. 2006, 85, 1285–1291. [Google Scholar] [CrossRef]
- Vatten, L.J.; Skjaerven, R. Offspring sex and pregnancy outcome by length of gestation. Early Hum. Dev. 2004, 76, 47–54. [Google Scholar] [CrossRef]
- Ghidini, A.; Salafia, C.M. Gender differences of placental dysfunction in severe prematurity. BJOG Int. J. Obstet. Gynaecol. 2005, 112, 140–144. [Google Scholar] [CrossRef]
- Byrne, J.; Warburton, D. Male excess among anatomically normal fetuses in spontaneous abortions. Am. J. Med. Genet. 1987, 26, 605–611. [Google Scholar] [CrossRef]
- Redman, C.W.G.; Sargent, I.L. Preeclampsia and the systemic inflammatory response. Semin. Nephrol. 2004, 24, 565–570. [Google Scholar] [CrossRef]
- Enninga, E.A.; Nevala, W.K.; Creedon, D.J.; Markovic, S.N.; Holtan, S.G. Fetal sex-based differences in maternal hormones, angiogenic factors, and immune mediators during pregnancy and the postpartum period. Am. J. Reprod. Immunol. 2015, 73, 251–262. [Google Scholar] [CrossRef]
- Veenstra van Nieuwenhoven, A.L.; Heineman, M.J.; Faas, M.M. The immunology of successful pregnancy. Hum. Reprod. Update 2003, 9, 347–357. [Google Scholar] [CrossRef] [PubMed]
- McCracken, S.A.; Gallery, E.; Morris, J.M. Pregnancy-specific down-regulation of NF-kappa B expression in T cells in humans is essential for the maintenance of the cytokine profile required for pregnancy success. J. Immunol. 2004, 172, 4583–4591. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, T.G.; Lin, H.; Guilbert, L.; Mosmann, T.R. Bidirectional cytokine interactions in the maternal-fetal relationship: Is successful pregnancy a TH2 phenomenon? Immunol. Today 1993, 14, 353–356. [Google Scholar] [CrossRef]
- Loke, Y.W.; King, A.; Burrows, T.D. Decidua in human implantation. Hum. Reprod. 1995, 10, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Moffett, A.; Hiby, S.E.; Sharkey, A.M. The role of the maternal immune system in the regulation of human birthweight. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 20140071. [Google Scholar] [CrossRef] [PubMed]
- Fukui, A.; Yokota, M.; Funamizu, A.; Nakamua, R.; Fukuhara, R.; Yamada, K.; Kimura, H.; Fukuyama, A.; Kamoi, M.; Tanaka, K.; et al. Changes of NK cells in preeclampsia. Am. J. Reprod. Immunol. (New York N. Y. 1989) 2012, 67, 278–286. [Google Scholar] [CrossRef]
- Kelemu, T.; Erlandsson, L.; Seifu, D.; Hansson, E.; Abebe, M.; Teklu, S.; Girma, S.; Traherne, J.A.; Moffett, A.; Hansson, S.R. Polymorphism in killer cell immunoglobulin-like receptors and human leukocyte antigen-c and predisposition to preeclampsia in Ethiopian pregnant women population. J. Reprod. Immunol. 2020, 141, 103169. [Google Scholar] [CrossRef]
- Tranquilli, A.L.; Dekker, G.; Magee, L.; Roberts, J.; Sibai, B.M.; Steyn, W.; Zeeman, G.G.; Brown, M.A. The classification, diagnosis and management of the hypertensive disorders of pregnancy: A revised statement from the ISSHP. Pregnancy Hypertens. 2014, 4, 97–104. [Google Scholar] [CrossRef]


{kind=link}
| Characteristics | Controls (n = 136) | PE Cases (n = 105) |
|---|---|---|
| Age | 25.3 ± 5 | 26.6 ± 5 |
| sBP | 114.0 ± 9 | 149.9 ± 17.2 |
| dBP | 72.4 ± 8.1 | 97.6 ± 10.1 |
| GA | 38 + 5 weeks | 35 + 6 weeks |
| Early-onset PE Late-onset PE | n.a n.a | 23.8% (25) 76.2% (80) |
| Maternal Genotype | PE Mothers of Males % (n = 51) | Control Mothers of Males % (n = 77) | PE Mothers of Females % (n = 54) | Control Mothers of Females % (n = 59) |
|---|---|---|---|---|
| G+ | 92.2 (47) | 79.2 (61) | 77.8 (42) | 81.4 (48) |
| CC | 7.8 (4) | 20.8 (16) | 22.2 (12) | 18.6 (11) |
| Fetal Genotype | Males of PE Mothers % (n = 51) | Males of Control Mothers % (n = 77) | Females of PE Mothers % (n = 54) | Females of Control Mothers % (n = 59) |
|---|---|---|---|---|
| G+ | 80.4 (41) | 76.6 (59) | 75.9 (41) | 79.7 (47) |
| CC | 19.6 (10) | 23.4 (18) | 24.1 (13) | 20.3 (12) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelemu, T.; Erlandsson, L.; Seifu, D.; Abebe, M.; Teklu, S.; Storry, J.R.; Hansson, S.R. Association of Maternal Regulatory Single Nucleotide Polymorphic CD99 Genotype with Preeclampsia in Pregnancies Carrying Male Fetuses in Ethiopian Women. Int. J. Mol. Sci. 2020, 21, 5837. https://doi.org/10.3390/ijms21165837
Kelemu T, Erlandsson L, Seifu D, Abebe M, Teklu S, Storry JR, Hansson SR. Association of Maternal Regulatory Single Nucleotide Polymorphic CD99 Genotype with Preeclampsia in Pregnancies Carrying Male Fetuses in Ethiopian Women. International Journal of Molecular Sciences. 2020; 21(16):5837. https://doi.org/10.3390/ijms21165837
Chicago/Turabian StyleKelemu, Tsehayneh, Lena Erlandsson, Daniel Seifu, Markos Abebe, Sisay Teklu, Jill R. Storry, and Stefan R. Hansson. 2020. "Association of Maternal Regulatory Single Nucleotide Polymorphic CD99 Genotype with Preeclampsia in Pregnancies Carrying Male Fetuses in Ethiopian Women" International Journal of Molecular Sciences 21, no. 16: 5837. https://doi.org/10.3390/ijms21165837
APA StyleKelemu, T., Erlandsson, L., Seifu, D., Abebe, M., Teklu, S., Storry, J. R., & Hansson, S. R. (2020). Association of Maternal Regulatory Single Nucleotide Polymorphic CD99 Genotype with Preeclampsia in Pregnancies Carrying Male Fetuses in Ethiopian Women. International Journal of Molecular Sciences, 21(16), 5837. https://doi.org/10.3390/ijms21165837