Upregulation of the MYB2 Transcription Factor is Associated with Increased Accumulation of Anthocyanin in the Leaves of Dendrobium bigibbum

 , ,
, , 
 and
and
Abstract
1. Introduction
2. Results
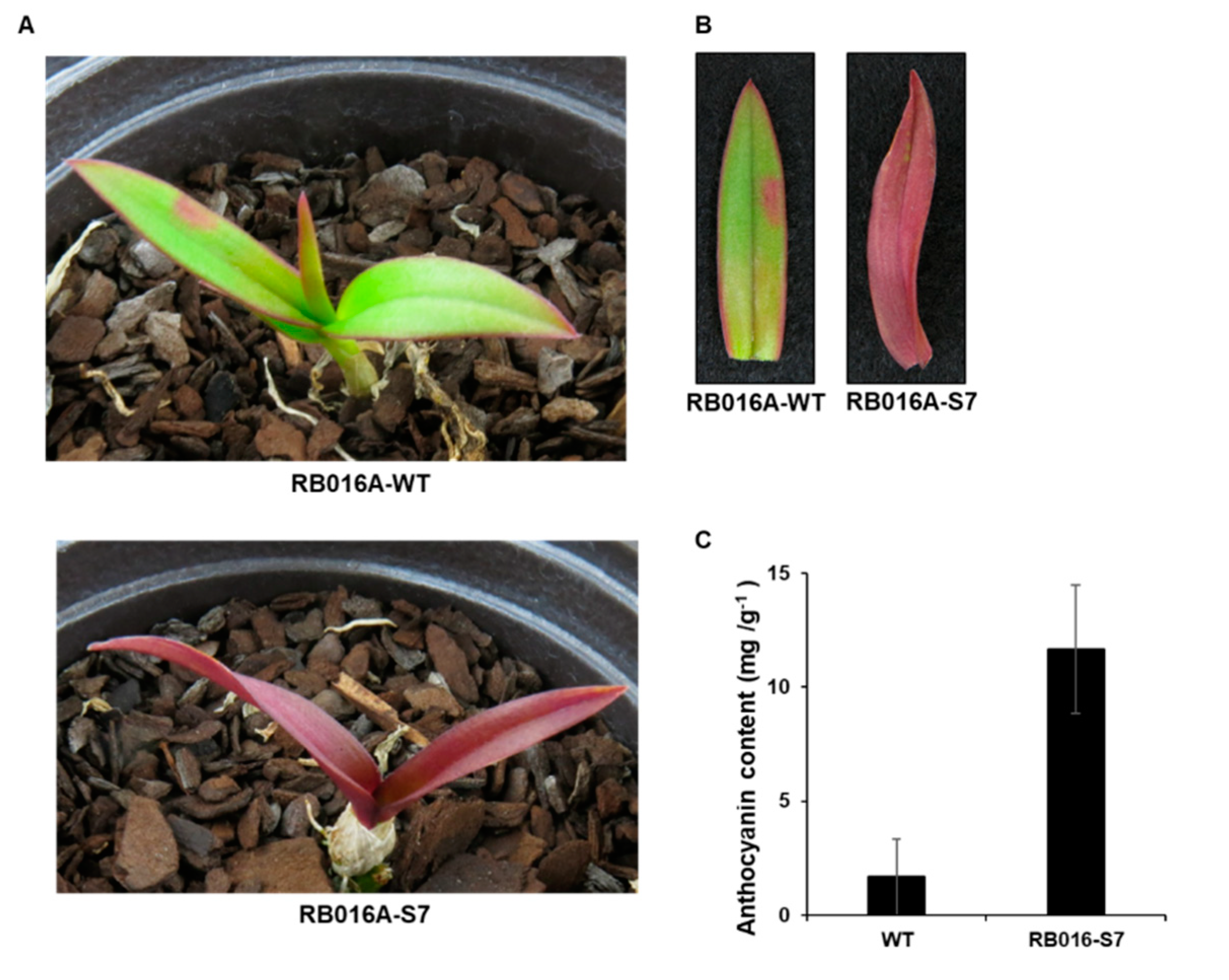
2.1. The Purple Mutant of D. Bigibbum Accumulates Higher Levels of Anthocyanin
2.2. Functional Annotation and Classification
2.3. Analysis of Differentially Expressed Genes (DEGs) Associated with Anthocyanin Biosynthesis
2.4. Analysis of Anthocyanin Biosynthetic Genes in Identified DEGs
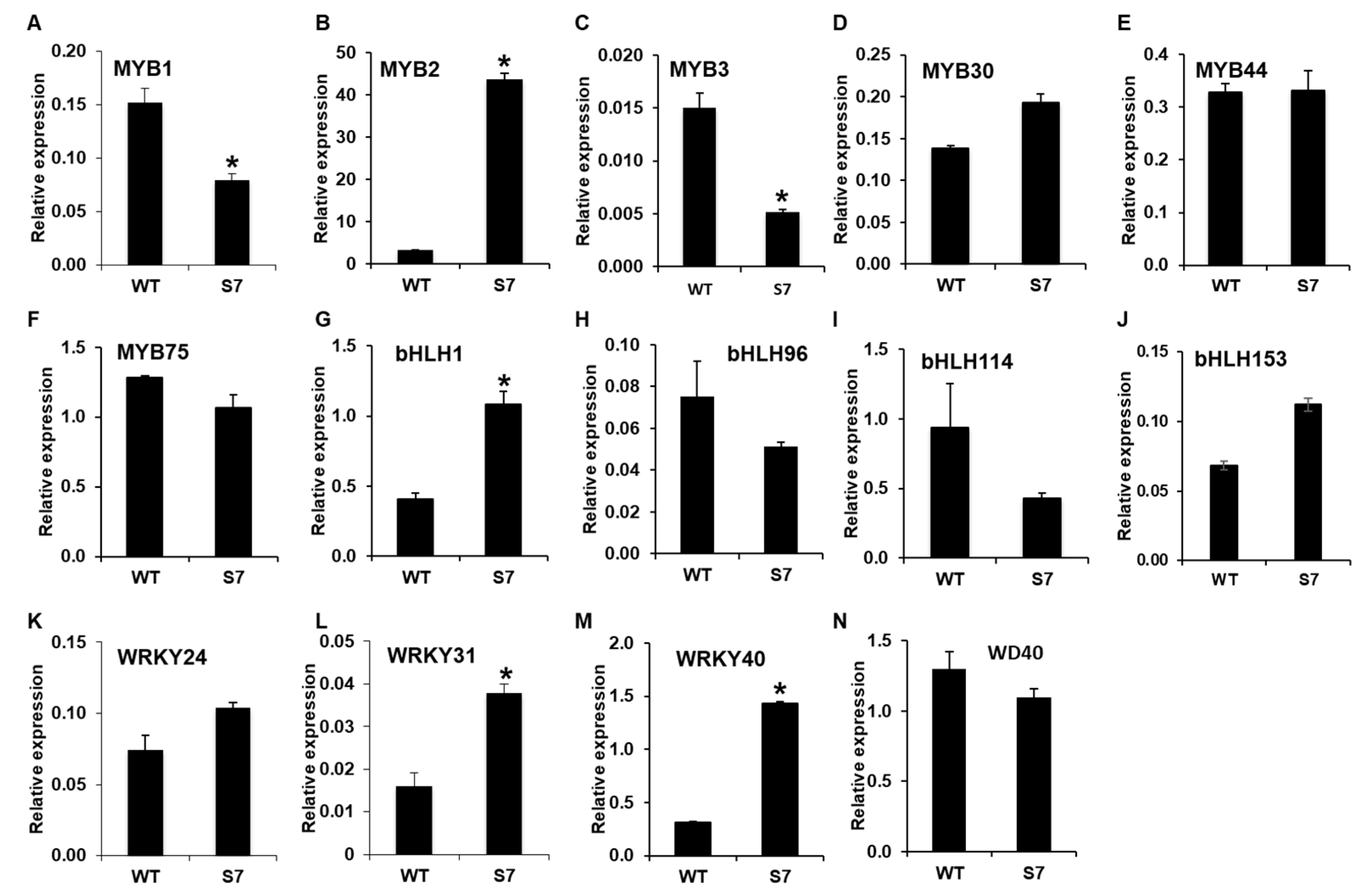
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis of the Genes Involved in Anthocyanin Biosynthesis
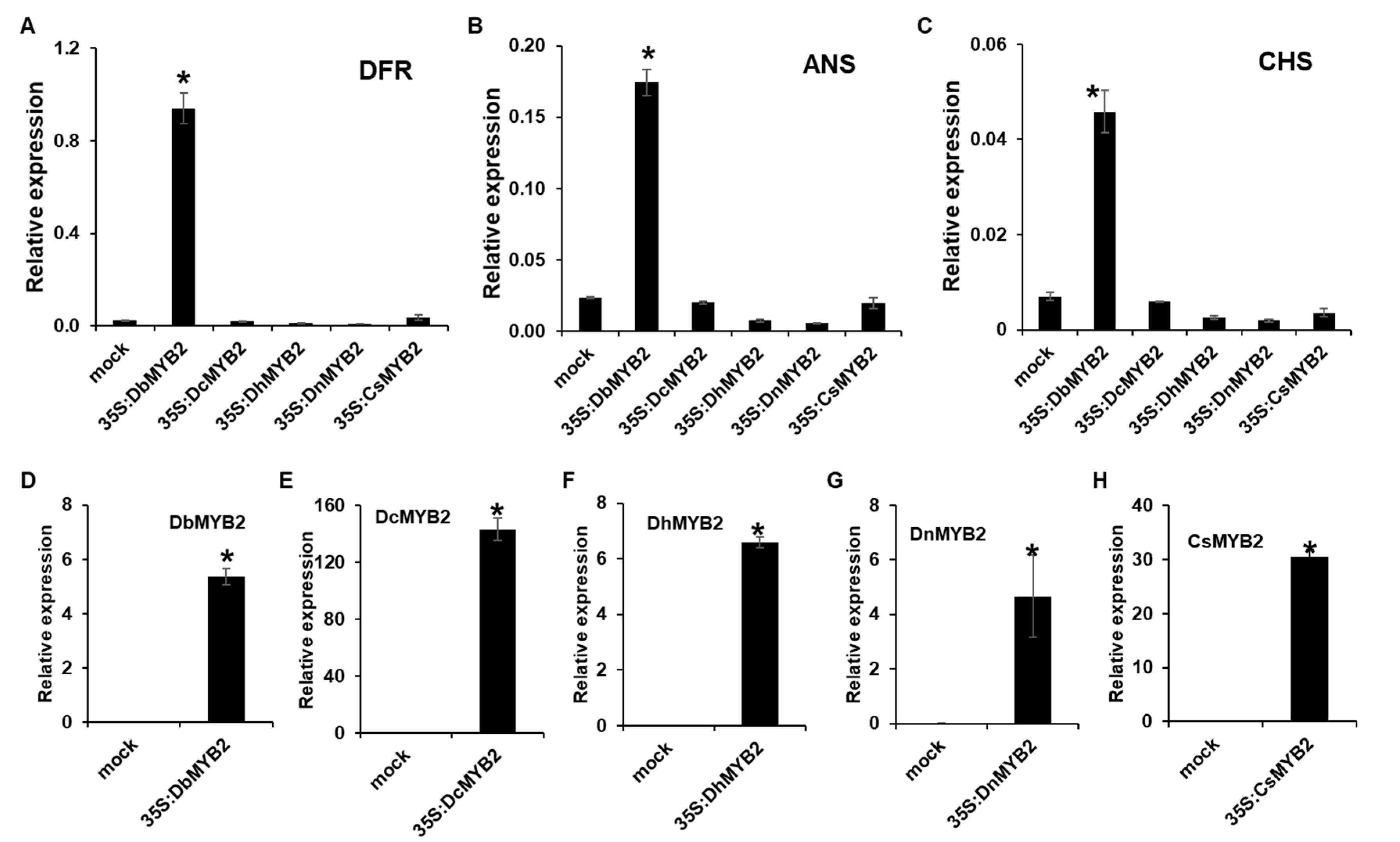
2.6. DbMYB2 Positively Regulates Anthocyanin Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. γ-Irradiation of in Vitro Shoot Cultures
4.3. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR) Analysis
4.4. Measurement of Total Anthocyanin Content
4.5. RNA-Seq Analysis, De Novo Assembly, and Unigene Generation
4.6. Functional Annotation of Unigenes
4.7. Expression Profiling of Unigenes
4.8. Identification of DEGs between the WT and RB016-S7 Mutant
4.9. Analysis of Unigenes Involved in Anthocyanin Biosynthesis
4.10. Cloning of Orchid MYB Genes
4.11. Agroinfiltration of N. Benthamiana
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANS | anthocyanidin synthase |
| 4CL | 4-coumarate CoA ligase |
| C4H | cinnamic acid 4-hydroxylase |
| CHI | chalcone isomerase |
| CHS | chalcone synthase |
| DFR | dihydroflavonol 4-reductase |
| F3H | flavanone 3-hydroxylase |
| F3′H | flavonoid 3′-monooxygenase |
| GO | gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| PAL | phenylalanine ammonia-lyase |
| WT | wild -type |
References
- Wang, H.Z.; Feng, S.G.; Lu, J.J.; Shi, N.N.; Liu, J.J. Phylogenetic study and molecular identification of 31 Dendrobium species using inter-simple sequence repeat (ISSR) markers. Sci. Hortic. 2009, 122, 440–447. [Google Scholar] [CrossRef]
- Feng, S.G.; Lu, J.J.; Gao, L.; Liu, J.J.; Wang, H.-Z. Molecular phylogeny analysis and species identification of Dendrobium (Orchidaceae) in China. Biochem. Genet. 2014, 52, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Bulpitt, C.J.; Li, Y.; Bulpitt, P.F. The use of orchids in Chinese medicine. Genome Natl. Res. Counc. Can. 2007, 100, 558–563. [Google Scholar]
- Wang, H.; Zhang, T.; Sun, W.; Wang, Z.; Zuo, D.; Zhou, Z.; Li, S.; Xu, J.; Yin, F.; Hua, Y. Erianin induces G2/M-phase arrest, apoptosis, and autophagy via the ROS/JNK signaling pathway in human osteosarcoma cells in vitro and in vivo. Cell Death Dis. 2016, 7, e2247. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Muddathir, A.M.; Yamauchi, K.; Batubara, I.; Mohieldin, E.A.M.; Mitsunaga, T. Anti-tyrosinase, total phenolic content and antioxidant activity of selected Sudanese medicinal plants. S. Afr. J. Bot. 2017, 109, 9–15. [Google Scholar] [CrossRef]
- Szymanowska, U.; Baraniak, B. Antioxidant and Potentially Anti-Inflammatory Activity of Anthocyanin Fractions from Pomace Obtained from Enzymatically Treated Raspberries. Antioxidants 2019, 8, 299. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant. Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef]
- Sadilova, E.; Stintzing, F.C.; Carle, R. Anthocyanins, colour and antioxidant properties of eggplant (Solanum melongena L.) and violet pepper (Capsicum annuum L.) peel extracts. Zeitschrift für Naturforschung C 2006, 61, 527–535. [Google Scholar] [CrossRef]
- Goodrich, J.; Carpenter, R.; Coen, E.S. A common gene regulates pigmentation pattern in diverse plant species. Cell 1992, 68, 955–964. [Google Scholar] [CrossRef]
- Wang, L.M.; Zhang, J.; Dong, X.Y.; Fu, Z.Z.; Jiang, H.; Zhang, H.C. Identification and functional analysis of anthocyanin biosynthesis genes in Phalaenopsis hybrids. Biol. Plant. 2018, 62, 45–54. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Wing, J.F.; Va, K.; Mol, J.N.M.; Koes, R. Analysis of bHLH and MYB domain proteins: Species-specific regulatory differences are caused by divergent evolution of target anthocyanin genes. Plant. J. 1998, 13, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Cone, K.C.; Cocciolone, S.M.; Burr, F.A.; Burr, B. Maize anthocyanin regulatory gene pl is a duplicate of c1 that functions in the plant. Plant. Cell 1993, 5, 1795–1805. [Google Scholar] [PubMed]
- Hanson, M.A.; Gaut, B.S.; Stec, A.O.; Fuerstenberg, S.I.; Goodman, M.M.; Coe, E.H.; Doebley, J.F. Evolution of anthocyanin biosynthesis in maize kernels: The role of regulatory and enzymatic loci. Genetics 1996, 143, 1395–1407. [Google Scholar]
- Kim, C.; Park, S.; Kikuchi, S.; Kwon, S.; Park, S.; Yoon, U.; Park, D.; Seol, Y.; Hahn, J.; Park, S. Genetic analysis of gene expression for pigmentation in Chinese cabbage (Brassica rapa). Biochip J. 2010, 4, 123–128. [Google Scholar] [CrossRef]
- Guo, N.; Cheng, F.; Wu, J.; Liu, B.; Zheng, S.; Liang, J.; Wang, X. Anthocyanin biosynthetic genes in Brassica rapa. BMC Genom. 2014, 15, 426. [Google Scholar] [CrossRef]
- Borevitz, J.O.; Xia, Y.; Blount, J.; Dixon, R.A.; Lamb, C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant. Cell 2000, 12, 2383–2393. [Google Scholar] [CrossRef]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 encodes a bHLH protein that regulates trichome development in Arabidopsis through interaction with GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar]
- Holton, T.A.; Cornish, E.C. Genetics and biochemistry of anthocyanin biosynthesis. Plant. Cell 1995, 7, 1071. [Google Scholar] [CrossRef]
- Britsch, L.; Ruhnau-Brich, B.; Forkmann, G. Molecular cloning, sequence analysis, and in vitro expression of flavanone 3 beta-hydroxylase from Petunia hybrida. J. Biol. Chem. 1992, 267, 5380–5387. [Google Scholar]
- Franken, P.; Niesbach-Klösgen, U.; Weydemann, U.; Maréchal-Drouard, L.; Saedler, H.; Wienand, U. The duplicated chalcone synthase genes C2 and Whp (white pollen) of Zea mays are independently regulated; evidence for translational control of Whp expression by the anthocyanin intensifying gene in. EMBO J. 1991, 10, 2605–2612. [Google Scholar] [CrossRef]
- Inagaki, Y.; Hisatomi, Y.; Iida, S. Somatic mutations caused by excision of the transposable element, Tpn1, from the DFR gene for pigmentation in sub-epidermal layer of periclinally chimeric flowers of Japanese morning glory and their germinal transmission to their progeny. Theor. Appl. Genet. 1996, 92, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, T.; Nishihara, M.; Mishiba, K.; Yamamura, S. Temporal expression of flavonoid biosynthesis-related genes regulates flower pigmentation in gentian plants. Plant. Sci. 2005, 168, 1309–1318. [Google Scholar] [CrossRef]
- Napoli, C.A.; Fahy, D.; Wang, H.-Y.; Taylor, L.P. White anther: A petunia mutant that abolishes pollen flavonol accumulation, induces male sterility, and is complemented by a chalcone synthase transgene. Plant Physiol. 1999, 120, 615–622. [Google Scholar] [CrossRef]
- Yu, Z.; Liao, Y.; Teixeira da Silva, J.A.; Yang, Z.; Duan, J. Differential accumulation of anthocyanins in dendrobium officinale stems with red and green peels. Int J. Mol. Sci. 2018, 19, 2857. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.C.; Hellens, R.P.; Laing, W.A. MYB transcription factors that colour our fruit. Trends Plant Sci. 2008, 13, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Schaart, J.G.; Dubos, C.; Romero De La Fuente, I.; van Houwelingen, A.M.M.L.; de Vos, R.C.H.; Jonker, H.H.; Xu, W.; Routaboul, J.M.; Lepiniec, L.; Bovy, A.G. Identification and characterization of MYB-b HLH-WD 40 regulatory complexes controlling proanthocyanidin biosynthesis in strawberry (F ragaria× ananassa) fruits. New Phytol. 2013, 197, 454–467. [Google Scholar] [CrossRef]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. A protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant. J. 2008, 55, 954–967. [Google Scholar] [CrossRef]
- Jun, J.H.; Liu, C.; Xiao, X.; Dixon, R.A. The transcriptional repressor MYB2 regulates both spatial and temporal patterns of proanthocyandin and anthocyanin pigmentation in Medicago truncatula. Plant Cell 2015, 27, 2860–2879. [Google Scholar]
- Hsu, C.-C.; Chen, Y.-Y.; Tsai, W.-C.; Chen, W.-H.; Chen, H.-H. Three R2R3-MYB transcription factors regulate distinct floral pigmentation patterning in Phalaenopsis spp. Plant Physiol. 2015, 168, 175–191. [Google Scholar] [CrossRef]
- Lu, H.-C.; Hsieh, M.-H.; Chen, C.-E.; Chen, H.-H.; Wang, H.-I.; Yeh, H.-H. A high-throughput virus-induced gene-silencing vector for screening transcription factors in virus-induced plant defense response in orchid. Mol. Plant Microbe Interact. 2012, 25, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Schrader, A. TRANSPARENT TESTA GLABRA 1-dependent regulation of flavonoid biosynthesis. Plants 2017, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Qiu, J.; Ding, L.; Huang, M.; Huang, S.; Yang, G.; Yin, J. Anthocyanin biosynthesis regulation of DhMYB2 and DhbHLH1 in Dendrobium hybrids petals. Plant Physiol. Biochem. 2017, 112, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. Overview of plant pigments. In Pigments in Fruits and Vegetables; Springer: New York, NY, USA, 2015; pp. 1–7. [Google Scholar]
- Veberic, R.; Slatnar, A.; Bizjak, J.; Stampar, F.; Mikulic-Petkovsek, M. Anthocyanin composition of different wild and cultivated berry species. LWT 2015, 60, 509–517. [Google Scholar] [CrossRef]
- Noda, N.; Yoshioka, S.; Kishimoto, S.; Nakayama, M.; Douzono, M.; Tanaka, Y.; Aida, R. Generation of blue chrysanthemums by anthocyanin B-ring hydroxylation and glucosylation and its coloration mechanism. Sci. Adv. 2017, 3, e1602785. [Google Scholar] [CrossRef]
- Lloyd, A.; Brockman, A.; Aguirre, L.; Campbell, A.; Bean, A.; Cantero, A.; Gonzalez, A. Advances in the MYB-bHLH-WD Repeat (MBW) Pigment Regulatory Model: Addition of a WRKY Factor and Co-option of an Anthocyanin MYB for Betalain Regulation. Plant Cell Physiol. 2017, 58, 1431–1441. [Google Scholar] [CrossRef]
- Albert, N.W.; Lewis, D.H.; Zhang, H.; Schwinn, K.E.; Jameson, P.E.; Davies, K.M. Members of an R2R3-MYB transcription factor family in Petunia are developmentally and environmentally regulated to control complex floral and vegetative pigmentation patterning. Plant J. 2011, 65, 771–784. [Google Scholar] [CrossRef]
- Gonzalez, A.; Brown, M.; Hatlestad, G.; Akhavan, N.; Smith, T.; Hembd, A.; Moore, J.; Montes, D.; Mosley, T.; Resendez, J. TTG2 controls the developmental regulation of seed coat tannins in Arabidopsis by regulating vacuolar transport steps in the proanthocyanidin pathway. Dev. Biol. 2016, 419, 54–63. [Google Scholar] [CrossRef]
- Ramsay, N.A.; Glover, B.J. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005, 10, 63–70. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Han, Y.; Wei, G.; Korban, S.S. An apple myb transcription factor, MdMYB3, is involved in regulation of anthocyanin biosynthesis and flower development. BMC Plant Biol. 2013, 13, 176. [Google Scholar] [CrossRef]
- Jo, Y.D.; Kim, J.-B. Frequency and Spectrum of Radiation-Induced Mutations Revealed by Whole-Genome Sequencing Analyses of Plants. Quantum Beam Sci. 2019, 3, 7. [Google Scholar] [CrossRef]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome. Bio. 2010, 11, R106–R110. [Google Scholar] [CrossRef]
- Ren, M.; Wang, Z.; Xue, M.; Wang, X.; Zhang, F.; Zhang, Y.; Zhang, W.; Wang, M. Constitutive expression of an A-5 subgroup member in the DREB transcription factor subfamily from Ammopiptanthus mongolicus enhanced abiotic stress tolerance and anthocyanin accumulation in transgenic Arabidopsis. PLoS ONE 2019, 14, e0224296. [Google Scholar]
- Hellens, R.; Mullineaux, P.; Klee, H. Technical focus: A guide to Agrobacterium binary Ti vectors. Trends Plant Sci. 2000, 5, 446–451. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequences | Control 1 | Control 2 | Control 3 | RB016-S7-1 | RB016-S7-2 | RB016-S7-3 |
|---|---|---|---|---|---|---|
| BEFORE TRIMMING | ||||||
| Total nucleotides (bp) | 4,962,911,568 | 5,845,449,712 | 5,115,158,264 | 4,956,765,904 | 5,110,844,048 | 4,343,315,032 |
| Number of raw reads | 65,301,468 | 76,913,812 | 67,304,714 | 65,220,604 | 67,247,948 | 57,148,882 |
| AFTER TRIMMING | ||||||
| Total nucleotides (bp) | 4,288,037,900 | 4,994,465,471 | 4,370,988,523 | 4,215,819,862 | 4,288,137,551 | 3,561,476,236 |
| Number of clean reads | 56,856,166 | 66,205,490 | 57,973,956 | 55,934,410 | 56,856,134 | 47,287,332 |
| GC content (%) | 46.42 | 44.22 | 45.73 | 47.06 | 46.79 | 46.55 |
| Q30 percentage (%) | 95.67 | 95.86 | 95.56 | 95.43 | 95.68 | 95.22 |
| AFTER ASSEMBLY | ||||||
| Number of transcripts in the combined data | 110,104 | |||||
| Number of unigenes in the combined data | 32,575 | |||||
| Total nucleotides of transcripts (bp) | 122,947,955 | |||||
| Total nucleotides of unigenes (bp) | 34,155,642 | |||||
| Mean length of transcripts (bp) | 1,116 | |||||
| Mean length of unigenes (bp) | 1,048 | |||||
| N50 of unigenes (bp) | 1350 | |||||
| Pathway | DEG Number | Pathway ID |
|---|---|---|
| Ribosome | 58 | ko03010 |
| MAPK signaling pathway-plant | 40 | ko04016 |
| Plant-pathogen interaction | 40 | ko04626 |
| Plant hormone signal transduction | 37 | ko04075 |
| Phenylpropanoid biosynthesis | 28 | ko00940 |
| Starch and sucrose metabolism | 25 | ko00500 |
| Flavonoid biosynthesis | 23 | ko00941 |
| Fluid shear stress and atherosclerosis | 23 | ko05418 |
| Phenylalanine metabolism | 19 | ko00360 |
| Cancer-related pathways | 18 | ko05200 |
| Protein processing in the endoplasmic reticulum | 18 | ko04141 |
| Cellular senescence | 16 | ko04218 |
| Endocytosis | 16 | ko04144 |
| Glycolysis/Gluconeogenesis | 16 | ko00010 |
| β-Alanine metabolism | 15 | ko00410 |
| Calcium signaling pathway | 14 | ko04020 |
| Oxytocin signaling pathway | 14 | ko04921 |
| Phagosome | 14 | ko04145 |
| Amino sugar and nucleotide sugar metabolism | 13 | ko00520 |
| Arginine and proline metabolism | 13 | ko00330 |
| Pathway | DEG Number | Pathway ID |
|---|---|---|
| Folate biosynthesis | 13 | ko00790 |
| Starch and sucrose metabolism | 8 | ko00500 |
| Plant hormone signal transduction | 7 | ko04075 |
| Brassinosteroid biosynthesis | 6 | ko00905 |
| Phenylpropanoid biosynthesis | 6 | ko00940 |
| Circadian rhythm - plant | 5 | ko04712 |
| Cyanoamino acid metabolism | 5 | ko00460 |
| Glyoxylate and dicarboxylate metabolism | 5 | ko00630 |
| Protein processing in the endoplasmic reticulum | 5 | ko04141 |
| Renin-angiotensin system | 5 | ko04614 |
| β-Alanine metabolism | 4 | ko00410 |
| Glycine, serine and threonine metabolism | 4 | ko00260 |
| Lysosome | 4 | ko04142 |
| Phenylalanine metabolism | 4 | ko00360 |
| Photosynthesis | 4 | ko00195 |
| Photosynthesis - antenna proteins | 4 | ko00196 |
| Platinum drug resistance | 4 | ko01524 |
| Protein digestion and absorption | 4 | ko04974 |
| Purine metabolism | 4 | ko00230 |
| Tropane, piperidine, and pyridine alkaloid biosynthesis | 4 | ko00960 |
| Gene Name | Unigene ID | Gene Length | FPKM | Fold Change | log2Fold Change | |
|---|---|---|---|---|---|---|
| Wild Type | S7 Mutant | |||||
| PAL1 | denphalae05809 | 2223 | 3.98 | 89.98 | 21.44 | 4.42 |
| PAL2 | denphalae05806 | 2093 | 12.98 | 196.34 | 14.13 | 3.82 |
| PAL3 | denphalae05808 | 2139 | 8.59 | 71.56 | 7.45 | 2.90 |
| C4H | denphalae10925 | 1518 | 1.92 | 18.63 | 8.97 | 3.16 |
| denphalae10926 * | 1518 | 6.09 | 36.67 | 5.50 | 2.46 | |
| 4CL | denphalae18583 * | 1731 | 15.16 | 79.17 | 4.78 | 2.26 |
| denphalae22607 | 1698 | 3.97 | 13.60 | 2.89 | 1.53 | |
| denphalae27156 | 1473 | 0.63 | 3.23 | 5.15 | 2.36 | |
| denphalae27157 | 1695 | 2.91 | 6.02 | - | - | |
| CHS | denphalae02657 * | 1173 | 2.52 | 9.64 | 3.40 | 1.77 |
| denphalae02658 | 1170 | 10.50 | 36.49 | 3.15 | 1.65 | |
| denphalae05188 | 1092 | 0.52 | 1.62 | - | - | |
| denphalae11910 | 1188 | 102.38 | 659.26 | 5.62 | 2.49 | |
| F3H | denphalae02991 | 1137 | 98.69 | 245.15 | 2.07 | 1.05 |
| F3’H | denphalae11915 | 1563 | 31.60 | 152.97 | 4.28 | 2.10 |
| DFR | denphalae11241 | 1059 | 102.75 | 311.67 | 2.54 | 1.34 |
| ANS | denphalae18276 | 1083 | 70.86 | 180.97 | 2.22 | 1.15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, G.-H.; Kim, S.W.; Ryu, J.; Kang, S.-Y.; Kim, J.-B.; Kim, S.H. Upregulation of the MYB2 Transcription Factor is Associated with Increased Accumulation of Anthocyanin in the Leaves of Dendrobium bigibbum. Int. J. Mol. Sci. 2020, 21, 5653. https://doi.org/10.3390/ijms21165653
Lim G-H, Kim SW, Ryu J, Kang S-Y, Kim J-B, Kim SH. Upregulation of the MYB2 Transcription Factor is Associated with Increased Accumulation of Anthocyanin in the Leaves of Dendrobium bigibbum. International Journal of Molecular Sciences. 2020; 21(16):5653. https://doi.org/10.3390/ijms21165653
Chicago/Turabian StyleLim, Gah-Hyun, Se Won Kim, Jaihyunk Ryu, Si-Yong Kang, Jin-Baek Kim, and Sang Hoon Kim. 2020. "Upregulation of the MYB2 Transcription Factor is Associated with Increased Accumulation of Anthocyanin in the Leaves of Dendrobium bigibbum" International Journal of Molecular Sciences 21, no. 16: 5653. https://doi.org/10.3390/ijms21165653
APA StyleLim, G.-H., Kim, S. W., Ryu, J., Kang, S.-Y., Kim, J.-B., & Kim, S. H. (2020). Upregulation of the MYB2 Transcription Factor is Associated with Increased Accumulation of Anthocyanin in the Leaves of Dendrobium bigibbum. International Journal of Molecular Sciences, 21(16), 5653. https://doi.org/10.3390/ijms21165653