Anticancer Potential of Furanocoumarins: Mechanistic and Therapeutic Aspects

 ,
, 


 and
and 
Abstract
1. Introduction
2. Chemistry of Furocoumarins
3. Bioavailability of Furocoumarins
4. Mechanistic Insights
4.1. NF-κB Inactivation
4.2. PI3K/Akt Inhibition
4.3. p53 Modulation
4.4. Cell Cycle Arrest
4.5. Autophagy
4.6. Antioxidant
5. Role in MDR Cancers
6. Furanocoumarins as Adjuvant with Other Anticancer Agents
7. Furanocoumarins and Cancer Risks
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, S.; Sharma, B.; Kanwar, S.S.; Kumar, A. Lead phytochemicals for anticancer drug development. Front. Plant Sci. 2016, 7, 1667. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A. Phytochemicals as potential anticancer drugs: Time to Ponder Nature’s Bounty. BioMed Res. Int. 2020, 2020, 8602879. [Google Scholar] [CrossRef]
- Khattak, S.; Khan, H. Anti-cancer potential of phyto-alkaloids: A prospective review. Curr. Cancer Ther. Rev. 2016, 12, 66–75. [Google Scholar] [CrossRef]
- Rauf, A.; Khan, R.; Khan, H.; Tokuda, H. Cytotoxic, antitumor-promoting and inhibition of protein denaturation effects of flavonoids, isolated from Potentilla evestita Th. Wolf. Nat. Prod. Res. 2015, 29, 1775–1778. [Google Scholar] [CrossRef]
- Khan, H.; Reale, M.; Ullah, H.; Sureda, A.; Tejada, S.; Wang, Y.; Zhang, Z.-J.; Xiao, J. Anti-cancer effects of polyphenols via targeting p53 signaling pathway: Updates and future directions. Biotechnol. Adv. 2020, 38, 107385. [Google Scholar] [CrossRef] [PubMed]
- Spector, D. Optimizing Cancer Survivors’ Health: The Role of Lifestyle Behaviors. J. Nurse Pract. 2018, 14, 323–329. [Google Scholar] [CrossRef]
- Patridge, E.V.; Gareiss, P.; Kinch, M.; Hoyer, D. An analysis of FDA-approved drugs: Natural products and their derivatives. Drug Discov. Today 2016, 21, 204–207. [Google Scholar] [CrossRef]
- Faraone, I.; Sinisgalli, C.; Ostuni, A.; Armentano, M.F.; Carmosino, M.; Milella, L.; Russo, D.; Labanca, F.; Khan, H. Astaxanthin anticancer effects are mediated through multiple molecular mechanisms: A systematic review. Pharmacol. Res. 2020, 155, 104689. [Google Scholar] [CrossRef]
- Del Río, J.A.; Diaz, L.; García-Bernal, D.; Blanquer, M.; Ortuño, A.; Correal, E.; Moraleda, J.M. Furanocoumarins: Biomolecules of therapeutic interest. In Studies in Natural Products Chemistry; Rahman, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 43, pp. 145–195. [Google Scholar]
- Melough, M.M.; Cho, E.; Chun, O.K. Furocoumarins: A review of biochemical activities, dietary sources and intake, and potential health risks. Food Chem. Toxicol. 2018, 113, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Sumorek-Wiadro, J.; Zając, A.; Maciejczyk, A.; Jakubowicz-Gil, J.; Joanna, S.-W.; Adrian, Z.; Aleksandra, M.; Joanna, J.-G. Furanocoumarins in anticancer therapy-For and against. Fitoterapia 2020, 142, 104492. [Google Scholar] [CrossRef] [PubMed]
- Olomola, T.O.; Mosebi, S.; Klein, R.; Traut-Johnstone, T.; Coates, J.; Hewer, R.; Kaye, P.T. Novel furocoumarins as potential HIV-1 integrase inhibitors. Bioorg. Chem. 2014, 57, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Pynam, H.; Dharmesh, S.M. Antioxidant and anti-inflammatory properties of marmelosin from Bael (Aegle marmelos L.); Inhibition of TNF-α mediated inflammatory/tumor markers. Biomed. Pharmacother. 2018, 106, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Melis, C.; Distinto, S.; Bianco, G.; Meleddu, R.; Cottiglia, F.; Fois, B.; Taverna, D.; Angius, R.; Alcaro, S.; Ortuso, F.; et al. Targeting Tumor Associated Carbonic Anhydrases IX and XII: Highly Isozyme Selective Coumarin and Psoralen Inhibitors. ACS Med. Chem. Lett. 2018, 9, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Lu, X.; Aisa, H.A. Preparation of novel 1,2,3-triazole furocoumarin derivatives via click chemistry and their anti-vitiligo activity. RSC Adv. 2019, 9, 1671–1678. [Google Scholar] [CrossRef]
- Niu, C.; Yin, L.; Aisa, H.A. Novel Furocoumarin Derivatives Stimulate Melanogenesis in B16 Melanoma Cells by Up-Regulation of MITF and TYR Family via Akt/GSK3β/β-Catenin Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 746. [Google Scholar] [CrossRef]
- Wang, F.; Li, J.; Li, R.; Pan, G.; Bai, M.; Huang, Q. Angelicin inhibits liver cancer growth in vitro and in vivo. Mol. Med. Rep. 2017, 16, 5441–5449. [Google Scholar] [CrossRef]
- Guillon, C.; Jan, Y.-H.; Foster, N.; Ressner, J.; Heck, D.E.; Laskin, J.D.; Heindel, N.D. Synthetically modified methoxsalen for enhanced cytotoxicity in light and dark reactions. Bioorg. Med. Chem. Lett. 2019, 29, 619–622. [Google Scholar] [CrossRef]
- Carbone, A.; Montalbano, A.; Spanò, V.; Musante, I.; Galietta, L.J.; Barraja, P. Furocoumarins as multi-target agents in the treatment of cystic fibrosis. Eur. J. Med. Chem. 2019, 180, 283–290. [Google Scholar] [CrossRef]
- Fujioka, T.; Furumi, K.; Fujii, H.; Okabe, H.; Mihashi, K.; Nakano, Y.; Matsunaga, H.; Katano, M.; Mori, M. Antiproliferative constituents from Umbelliferae plants. V. A new furanocoumarin and falcarindiol furanocoumarin ethers from the root of Angelica japonica. Chem. Pharm. Bull. 1999, 47, 96–100. [Google Scholar] [CrossRef]
- Gawron, A.; Glowniak, K. Cytostatic activity of coumarins in vitro. Planta Med. 1987, 53, 526–529. [Google Scholar] [CrossRef]
- Widelski, J.; Kukula-Koch, W.; Baj, T.; Kedzierski, B.; Fokialakis, N.; Magiatis, P.; Pozarowski, P.; Roliński, J.; Graikou, K.; Chinou, I. Rare Coumarins Induce Apoptosis, G1 Cell Block and Reduce RNA Content in HL60 Cells. Open Chem. 2017, 15, 1–6. [Google Scholar] [CrossRef]
- Godwin, P.; Baird, A.-M.; Heavey, S.; Barr, M.; O’Byrne, K.J.; Gately, K.A. Targeting Nuclear Factor-Kappa B to Overcome Resistance to Chemotherapy. Front. Oncol. 2013, 3, 120. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Feldman, G.M.; Tosato, G. Inhibition of STAT3 signaling induces apoptosis and decreases survivin expression in primary effusion lymphoma. Blood 2003, 101, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Bode, A.M.; Dong, Z. Breaking the NF-κB and STAT3 alliance inhibits inflammation and pancreatic tumorigenesis. Cancer Prev. Res. 2010, 3, 1379–1381. [Google Scholar] [CrossRef]
- Luo, W.; Song, Z.; Sun, H.; Liang, J.; Zhao, S. Bergamottin, a natural furanocoumarin abundantly present in grapefruit juice, suppresses the invasiveness of human glioma cells via inactivation of Rac1 signaling. Oncol. Lett. 2018, 15, 3259–3266. [Google Scholar] [CrossRef]
- Wang, Y.; Hong, C.; Zhou, C.; Xu, D.; Qu, H.-B. Screening antitumor compounds psoralen and isopsoralen from Psoralea corylifolia L. seeds. Evid.-Based Complement. Altern. Med. 2011, 2011, 363052. [Google Scholar] [CrossRef]
- Augustin, S.; Berard, M.; Kellaf, S.; Peyri, N.; Fauvel-Lafève, F.; Legrand, C.; He, L.; Crépin, M. Matrix metalloproteinases are involved in both type I (apoptosis) and type II (autophagy) cell death induced by sodium phenylacetate in MDA-MB-231 breast tumour cells. Anticancer Res. 2009, 29, 1335–1343. [Google Scholar]
- Chase, A.J.; Bond, M.; Crook, M.F.; Newby, A. Role of nuclear factor-kappa B activation in metalloproteinase-1, -3, and -9 secretion by human macrophages in vitro and rabbit foam cells produced in vivo. Arter. Thromb. Vasc. Biol. 2002, 22, 765–771. [Google Scholar] [CrossRef]
- Artacho-Cordón, F.; Ríos-Arrabal, S.; Lara, P.; Artacho-Cordón, A.; Calvente, I.; Nuñez, M. Matrix metalloproteinases: Potential therapy to prevent the development of second malignancies after breast radiotherapy. Surg. Oncol. 2012, 21, e143–e151. [Google Scholar] [CrossRef]
- Escárcega, R.O.; Fuentes-Alexandro, S.; García-Carrasco, M.; Gatica, A.; Zamora, A. The Transcription Factor Nuclear Factor-kappa B and Cancer. Clin. Oncol. 2007, 19, 154–161. [Google Scholar] [CrossRef]
- Papa, S.; Zazzeroni, F.; Pham, C.G.; Bubici, C.; Franzoso, G. Linking JNK signaling to NF-B: A key to survival. J. Cell Sci. 2004, 117, 5197–5208. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Song, X.; He, Y.; Yao, J.; Huang, C.; Deng, Y.; Xie, S.; Ren, J.; Jin, M.; Liu, H. Angelicin inhibits human lung carcinoma A549 cell growth and migration through regulating JNK and ERK pathways. Oncol. Rep. 2016, 36, 3504–3512. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Nam, D.; Um, J.Y.; Jung, S.H.; Sethi, G.; Ahn, K.S. Bergamottin Suppresses Metastasis of Lung Cancer Cells through Abrogation of Diverse Oncogenic Signaling Cascades and Epithelial-to-Mesenchymal Transition. Molecules (Basel, Switzerland) 2018, 23, 1601. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-H.; Arfuso, F.; Sethi, G.; Ahn, K.S. Pharmacological Utilization of Bergamottin, Derived from Grapefruits, in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2018, 19, 4048. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Yun, H.J.; Choi, J.H.; Kang, K.W.; Jeong, H.G. Suppression of phorbol-12-myristate-13-acetate-induced tumor cell invasion by bergamottin via the inhibition of protein kinase Cδ/p38 mitogen-activated protein kinase and JNK/nuclear factor-κB-dependent matrix metalloproteinase-9 expression. Mol. Nutr. Food Res. 2009, 54, 977–990. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, K.; Han, Y.; Zhang, G.; Dong, J.; Cui, Y.; Yang, Z. Effects of psoralen as an anti-tumor agent in human breast cancer MCF-7/ADR cells. Biol. Pharm. Bull. 2016, 39, 815–822. [Google Scholar] [CrossRef]
- Embade, N.; Valerón, P.F.; Aznar, S.; López-Collazo, E.; Lacal, J.C. Apoptosis Induced by Rac GTPase Correlates with Induction of FasL and Ceramides Production. Mol. Biol. Cell 2000, 11, 4347–4358. [Google Scholar] [CrossRef]
- Gastonguay, A.; Berg, T.; Hauser, A.D.; Schuld, N.; Lorimer, E.; Williams, C.L. The role of Rac1 in the regulation of NF-kappaB activity, cell proliferation, and cell migration in non-small cell lung carcinoma. Cancer Biol. Ther 2012, 13, 647–656. [Google Scholar] [CrossRef]
- Rayburn, E.R.; Ezell, S.J.; Zhang, R. Anti-Inflammatory Agents for Cancer Therapy. Mol. Cell. Pharmacol. 2009, 1, 29–43. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Castilho, P.C.M.F.; Gomila, A.S.; D’Onofrio, G.; Filosa, R.; Wang, F.; Nabavi, S.M.; Daglia, M.; Silva, A.S.; et al. Targeting NF-kappaB signaling pathway in cancer by dietary polyphenols. Crit. Rev. Food Sci. Nutr. 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb Perspect Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Khan, H.; Fratantonio, D.; Hasan, M.M.; Sharifi, S.; Fathi, N.; Ullah, H.; Rastrelli, L.; Sharifi, S. Apoptosis induced by luteolin in breast cancer: Mechanistic and therapeutic perspectives. Phytomedicine 2019, 59, 152883. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-L.; Liang, Y.C.; Chang, C.W.; Lee, W.-S.; Kuo, C.T.; Wang, C.C.; Lee, H.-M.; Lin, C.-H. Effects of sphondin, isolated from Heracleum laciniatum, on IL-1beta-induced cyclooxygenase-2 expression in human pulmonary epithelial cells. Life Sci. 2002, 72, 199–213. [Google Scholar] [CrossRef]
- Nasser, M.; Zhu, S.; Hu, H.; Huang, H.; Guo, M.; Zhu, P. Effects of imperatorin in the cardiovascular system and cancer. Biomed. Pharmacother. 2019, 120, 109401. [Google Scholar] [CrossRef]
- Zhang, X.; Li, W.; Abudureheman, A.; Cheng, T.; Peng, P. Imperatorin possesses notable anti-inflammatory activity in vitro and in vivo through inhibition of the NF kappa B pathway. Mol. Med. Rep. 2017, 16, 8619–8626. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, K.; Mei, W.; Wang, Y.; Yu, C.; Yu, B.; Deng, S.; Hu, J. Anti-inflammatory and pro resolution activities of bergapten isolated from the roots of Ficus hirta in an in vivo zebrafish model. Biochem. Biophys. Res. Commun. 2018, 496, 763–769. [Google Scholar] [CrossRef]
- Li, X.; Yu, C.; Hu, Y.; Xia, X.; Liao, Y.; Zhang, J.; Chen, H.; Lu, W.; Zhou, W.; Song, Z. New application of psoralen and angelicin on periodontitis with anti-bacterial, anti-inflammatory, and osteogenesis effects. Front. Microbiol. 2018, 8, 178. [Google Scholar] [CrossRef]
- A West, K.; Castillo, S.S.; Dennis, P.A. Activation of the PI3K/Akt pathway and chemotherapeutic resistance. Drug Resist. Updat. 2002, 5, 234–248. [Google Scholar] [CrossRef]
- Nicholson, K.M.; Anderson, N.G. The protein kinase B/Akt signalling pathway in human malignancy. Cell Signal 2002, 14, 381–395. [Google Scholar] [CrossRef]
- Bartnik, M.; Sławińska-Brych, A.; Żurek, A.; Kandefer-Szerszeń, M.; Zdzisińska, B. 8-methoxypsoralen reduces AKT phosphorylation, induces intrinsic and extrinsic apoptotic pathways, and suppresses cell growth of SK-N-AS neuroblastoma and SW620 metastatic colon cancer cells. J. Ethnopharmacol. 2017, 207, 19–29. [Google Scholar] [CrossRef]
- Zheng, Y.M.; Lu, A.X.; Shen, Z.; Kwok, A.H.Y.; Ho, W.S. Imperatorin exhibits anticancer activities in human colon cancer cells via the caspase cascade. Oncol. Rep. 2016, 35, 1995–2002. [Google Scholar] [CrossRef] [PubMed]
- Pattanayak, S.P.; Bose, P.; Sunita, P.; Siddique, M.U.M.; Lapenna, A. Bergapten inhibits liver carcinogenesis by modulating LXR/PI3K/Akt and IDOL/LDLR pathways. Biomed. Pharmacother. 2018, 108, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Toné, S.; Sugimoto, K.; Tanda, K.; Suda, T.; Uehira, K.; Kanouchi, H.; Samejima, K.; Minatogawa, Y.; Earnshaw, W.C. Three distinct stages of apoptotic nuclear condensation revealed by time-lapse imaging, biochemical and electron microscopy analysis of cell-free apoptosis. Exp. Cell Res. 2007, 313, 3635–3644. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S.A. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, Y.-P.; Lin, H.; Zhang, L.-Q.; Wu, L.-J.; Pang, L.-X. Antiproliferative and apoptotic effects of angelicin in highly invasive prostate cancer cells. Trop. J. Pharm. Res. 2015, 14, 1405–1411. [Google Scholar] [CrossRef]
- Wu, H.-J.; Wu, H.-B.; Zhao, Y.-Q.; Chen, L.-J.; Zou, H.-Z. Bergamottin isolated from Citrus bergamia exerts in vitro and in vivo antitumor activity in lung adenocarcinoma through the induction of apoptosis, cell cycle arrest, mitochondrial membrane potential loss and inhibition of cell migration and invasion. Oncol. Rep. 2016, 36, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Am, J.U.; Gong, W.J.; Su, Y.; Mou, Z.B. Imperatorin shows selective antitumor effects in SGC-7901 human gastric adenocarcinoma cells by inducing apoptosis, cell cycle arrest and targeting PI3K/Akt/m-TOR signalling pathway. J. Balk. Union Oncol. 2018, 22, 1471–1476. [Google Scholar]
- Ataur Rahman, M.; Kim, N.-H.; Yang, H.; Huh, S.-O. Angelicin induces apoptosis through intrinsic caspase-dependent pathway in human SH-SY5Y neuroblastoma cells. Mol. Cell. Biochem. 2012, 369, 95–104. [Google Scholar] [CrossRef]
- Min, K.J.; Um, H.J.; Seo, S.U.; Woo, S.M.; Kim, S.; Park, J.W.; Lee, H.S.; Kim, S.H.; Choi, Y.H.; Lee, T.J. Angelicin potentiates TRAIL-induced apoptosis in renal carcinoma Caki cells through activation of caspase 3 and down-regulation of c-FLIP expression. Drug Dev. Res. 2017, 79, 3–10. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, S.; Lin, T. Bergamottin exerts anticancer effects on human colon cancer cells via induction of apoptosis, G2/M cell cycle arrest and deactivation of the Ras/Raf/ERK signalling pathway. Arch. Med. Sci. 2019, 15. [Google Scholar] [CrossRef]
- Panno, M.L.; Giordano, F.; Rizza, P.; Pellegrino, M.; Zito, D.; Giordano, C.; Mauro, L.; Catalano, S.; Aquila, S.; Sisci, D. Bergapten induces ER depletion in breast cancer cells through SMAD4-mediated ubiquitination. Breast Cancer Res. Treat. 2012, 136, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.P.; Lin, C.S.; Lin, H.H.; Li, K.T.; Kao, S.H.; Tsao, S.M. Bergapten induces G1 arrest and pro-apoptotic cascade in colorectal cancer cells associating with p53/p21/PTEN axis. Environ. Toxicol. 2019, 34, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.-C.; Qu, X.; Yu, H.-F.; Zhang, H.-M.; Wang, Z.-H.; Zhang, Z.-T. Antitumor and apoptotic effects of bergaptol are mediated via mitochondrial death pathway and cell cycle arrest in human breast carcinoma cells. Bangladesh J. Pharmacol. 2016, 11, 489–494. [Google Scholar] [CrossRef]
- Yang, H.; Xiong, J.; Luo, W.; Jian, Y.; Xi, T. 8-Methoxypsoralen Induces Intrinsic Apoptosis in HepG2 Cells: Involvement of Reactive Oxygen Species Generation and ERK1/2 Pathway Inhibition. Cell. Physiol. Biochem. 2015, 37, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Pae, H.-O.; Oh, H.; Yun, Y.-G.; Oh, G.-S.; Jang, S.I.; Hwang, K.-M.; Kwon, T.-O.; Lee, H.-S.; Chung, H.-T. Imperatorin, a furanocoumarin from Angelica dahurica (Umbelliferae), induces cytochrome c-dependent apoptosis in human pro myelocytic leukaemia, HL-60 Cells. Pharmacol. Toxicol. 2002, 91, 40–48. [Google Scholar] [CrossRef]
- Rahman, A.; Siddiqui, S.A.; Jakhar, R.; Kang, S.C. Growth inhibition of various human cancer cell lines by imperatorin and limonin from Poncirus trifoliata Rafin. Seeds. Anticancer Agents Med. Chem. 2015, 15, 236–241. [Google Scholar] [CrossRef]
- Luo, K.-W.; Sun, J.-G.; Chan, J.; Yang, L.; Wu, S.; Fung, K.-P.; Liu, F. Anticancer Effects of Imperatorin Isolated from Angelica dahurica: Induction of Apoptosis in HepG2 Cells through both Death-Receptor- and Mitochondria-Mediated Pathways. Chemotherapy 2011, 57, 449–459. [Google Scholar] [CrossRef]
- Tong, K.; Xin, C.; Chen, W. Isoimperatorin induces apoptosis of the SGC-7901 human gastric cancer cell line via the mitochondria-mediated pathway. Oncol. Lett. 2016, 13, 518–524. [Google Scholar] [CrossRef]
- Dong, L.; Xu, W.-W.; Li, H.; Bi, K.-H. In vitro and in vivo anticancer effects of marmesin in U937 human leukemia cells are mediated via mitochondrial-mediated apoptosis, cell cycle arrest, and inhibition of cancer cell migration. Oncol. Rep. 2017, 39, 597–602. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Ryu, S.Y. Antiproliferative Effect of Furanocoumarins from the Root of Angelica dahurica on Cultured Human Tumor Cell Lines. Phytother. Res. 2007, 21, 288–290. [Google Scholar] [CrossRef]
- Patil, J.; Jayaprakasha, G.K.; Kim, J.; Murthy, K.; Chetti, M.; Nam, S.-Y.; Patil, B.S. 5-Geranyloxy-7-Methoxycoumarin inhibits colon cancer (SW480) cells growth by inducing apoptosis. Planta Med. 2013, 79, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Peng, P.; Pan, Z.; Fang, Z.; Lu, W.; Liu, X. Psoralen inhibits malignant proliferation and induces apoptosis through triggering endoplasmic reticulum stress in human SMMC7721 hepatoma cells. Biol. Res. 2019, 52, 34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jiang, H.; Zhang, J.; Zhang, Y.; Liu, A.; Zhao, Y.; Zhu, X.; Lin, Z.; Yuan, X. Liver X receptor activation induces apoptosis of melanoma cell through caspase pathway. Cancer Cell Int. 2014, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, J.; Guo, D. SCAP/SREBPs are Central Players in Lipid Metabolism and Novel Metabolic Targets in Cancer Therapy. Curr. Top. Med. Chem. 2018, 18, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.L.; Levine, A.J. The p53 pathway: Positive and negative feedback loops. Oncogene 2005, 24, 2899–2908. [Google Scholar] [CrossRef]
- Chiang, S.; Lin, C.; Lin, H.; Shieh, P.; Kao, S.-H. Bergapten induces G1 arrest of non-small cell lung cancer cells, associated with the p53-mediated cascade. Mol. Med. Rep. 2019, 19, 1972–1978. [Google Scholar] [CrossRef]
- Fang, H.; Ji, H. Furanocoumarin A: A Novel Anticancer agent on human lung cancer A549 cells from Fructus liquidambaris. Anticancer Agents Med. Chem. 2019, 19, 2091–2096. [Google Scholar] [CrossRef]
- Choochuay, K.; Chunhacha, P.; Pongrakhananon, V.; Luechapudiporn, R.; Chanvorachote, P. Imperatorin sensitizes anoikis and inhibits anchorage-independent growth of lung cancer cells. J. Nat. Med. 2012, 67, 599–606. [Google Scholar] [CrossRef]
- Panno, M.L.; Giordano, F. Effects of psoralens as anti-tumoral agents in breast cancer cells. World J. Clin. Oncol. 2014, 5, 348–358. [Google Scholar] [CrossRef]
- Panno, M.L.; Giordano, F.; Palma, M.G.; Bartella, V.; Rago, V.; Maggiolini, M.; Sisci, D.; Lanzino, M.; De Amicis, F.; Ando, S. Evidence that bergapten, independently of its photoactivation, enhances p53 gene expression and induces apoptosis in human breast cancer cells. Curr. Cancer Drug Targets 2009, 9, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, C.; Hua, Y.; Sun, L.; Cheng, K.; Jia, Z.; Han, Y.; Dong, J.; Cui, Y.; Yang, Z. Exosomes play an important role in the process of psoralen reverse multidrug resistance of breast cancer. J. Exp. Clin. Cancer Res. 2016, 35, 186. [Google Scholar] [CrossRef] [PubMed]
- Simabuco, F.M.; Morale, M.G.; Pavan, I.C.; Morelli, A.P.; Silva, F.R.; Tamura, R.E. p53 and metabolism: From mechanism to therapeutics. Oncotarget 2018, 9, 23780–23823. [Google Scholar] [CrossRef] [PubMed]
- Burroughs, S.K.; Kaluz, S.; Wang, D.; Wang, K.; Van Meir, E.G.; Wang, B. Hypoxia inducible factor pathway inhibitors as anticancer therapeutics. Future Med. Chem. 2013, 5, 553–572. [Google Scholar] [CrossRef]
- Mi, C.; Ma, J.; Wang, K.S.; Zuo, H.X.; Wang, Z.; Li, M.Y.; Piao, L.-X.; Xu, G.-H.; Li, X.; Quan, Z.-S. Imperatorin suppresses proliferation and angiogenesis of human colon cancer cell by targeting HIF-1α via the mTOR/p70S6K/4E-BP1 and MAPK pathways. J. Ethnopharmacol. 2017, 203, 27–38. [Google Scholar] [CrossRef]
- Haseeb, M.; Pirzada, R.H.; Ain, Q.U.; Choi, S. Wnt Signaling in the Regulation of Immune Cell and Cancer Therapeutics. Cells 2019, 8, 1380. [Google Scholar] [CrossRef]
- Ingham, M.; Schwartz, G.K. Cell-Cycle Therapeutics Come of Age. J. Clin. Oncol. 2017, 35, 2949–2959. [Google Scholar] [CrossRef]
- Mills, C.C.; Kolb, E.; Sampson, V.B. Recent Advances of Cell-Cycle Inhibitor Therapies for Pediatric Cancer. Cancer Res. 2017, 77, 6489–6498. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Chen, X.; Liang, Y.; Yang, D.; Dong, J.; Yang, N.; Liang, Z. Angelicin inhibits the malignant behaviors of human cervical cancer potentially via inhibiting autophagy. Exp. Ther. Med. 2019, 18, 3365–3374. [Google Scholar] [CrossRef]
- Lee, Y.; Wu, T.; Chen, S.; Chung, J.-G. Effect of 5-methoxypsoralen (5-MOP) on cell apoptosis and cell cycle in human hepatocellular carcinoma cell line. Toxicol. In Vitro 2003, 17, 279–287. [Google Scholar] [CrossRef]
- Kang, J.-H.; Lee, S.-K.; Yim, D.-S. Effect of isoimperatorin on the proliferation of prostate cancer cell line DU145 cells. Biomol. Ther. 2005, 13, 185–189. [Google Scholar]
- Wang, X.; Xu, C.; Hua, Y.; Cheng, K.; Zhang, Y.; Liu, J.; Han, Y.; Liu, S.; Zhang, G.; Xu, S.; et al. Psoralen induced cell cycle arrest by modulating Wnt/β-catenin pathway in breast cancer cells. Sci. Rep. 2018, 8, 14001. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Karantza, V. Autophagy as a therapeutic target in cancer. Cancer Biol. Ther. 2011, 11, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef]
- Song, M.S.; Salmena, L.; Pandolfi, P.P. The functions and regulation of the PTEN tumour suppressor. Nat. Rev. Mol. Cell Biol. 2012, 13, 283–296. [Google Scholar] [CrossRef]
- De Amicis, F.; Aquila, S.; Morelli, C.; Guido, C.; Santoro, M.; Perrotta, I.; Mauro, L.; Giordano, F.; Nigro, A.; Andò, S.; et al. Bergapten drives autophagy through the up-regulation of PTEN expression in breast cancer cells. Mol. Cancer 2015, 14, 130. [Google Scholar] [CrossRef]
- Kaewpiboon, C.; Surapinit, S.; Malilas, W.; Moon, J.; Phuwapraisirisan, P.; Tip-Pyang, S.; Johnston, R.N.; Koh, S.S.; Assavalapsakul, W.; Chung, Y.-H. Feroniellin A-induced autophagy causes apoptosis in multidrug-resistant human A549 lung cancer cells. Int. J. Oncol. 2014, 44, 1233–1242. [Google Scholar] [CrossRef]
- Haq, S.H.; Al-Ruwaished, G.; Al-Mutlaq, M.A.; Naji, S.A.; Al-Mogren, M.; Al-Rashed, S.; Ain, Q.T.; Al-Amro, A.A.; Al-Mussallam, A. Antioxidant, anticancer activity and phytochemical analysis of Green Algae, Chaetomorpha collected from the Arabian Gulf. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef]
- Hung, W.-L.; Suh, J.H.; Wang, Y. Chemistry and health effects of furanocoumarins in grapefruit. J. Food Drug Anal. 2017, 25, 71–83. [Google Scholar] [CrossRef]
- Karakaya, S.; Koca, M.; Yılmaz, S.V.; Yıldırım, K.; Pınar, N.M.; Demirci, B.; Brestic, M.; Sytar, O. Molecular docking studies of coumarins isolated from extracts and essential oils of Zosima absinthifolia Link as potential inhibitors for Alzheimer’s Disease. Molecules 2019, 24, 722. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, Y.; Wang, N.; He, L. Antioxidant effect of imperatorin from Angelica dahurica in hypertension via inhibiting NADPH oxidase activation and MAPK pathway. J. Am. Soc. Hypertens. 2014, 8, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Razavi, S.; Zahri, S.; Motamed, Z.; Ghasemi, G. Bioscreening of oxypeucedanin, a known Furanocoumarin. Iran J. Basic. Med. Sci. 2010, 13, 133–138. [Google Scholar]
- Dhalwal, K.; Shinde, V.M.; Namdeo, A.G.; Mahadik, K.R. Antioxidant profile and HPTLC-Densitometric analysis of umbelliferone and psoralen in Aegle marmelos. Pharm. Biol. 2008, 46, 266–272. [Google Scholar] [CrossRef]
- Upadhyay, J.; Kesharwani, R.K.; Misra, K. Comparative study of antioxidants as cancer preventives through inhibition of HIF-1 alpha activity. Bioinformation 2009, 4, 233–236. [Google Scholar] [CrossRef][Green Version]
- Kasaian, J.; Mosaffa, F.; Behravan, J.; Masullo, M.; Piacente, S.; Ghandadi, M.; Iranshahi, M. Reversal of P-glycoprotein-mediated multidrug resistance in MCF-7/Adr cancer cells by Sesquiterpene coumarins. Fitoterapia 2015, 103, 149–154. [Google Scholar] [CrossRef]
- Dewanjee, S.; Dua, T.K.; Bhattacharjee, N.; Das, A.; Gangopadhyay, M.; Khanra, R.; Joardar, S.; Riaz, M.; De Feo, V.; Zia-Ul-Haq, M. Natural products as alternative choices for P-Glycoprotein (P-gp) inhibition. Molecules 2017, 22, 871. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, X.; Cheng, K.; Zhao, W.; Hua, Y.; Xu, C.; Yang, Z. Psoralen reverses the P-glycoprotein-mediated multidrug resistance in human breast cancer MCF-7/ADR cells. Mol. Med. Rep. 2016, 13, 4745–4750. [Google Scholar] [CrossRef]
- Kubrak, T.; Bogucka-Kocka, A.; Komsta, Ł.; Załuski, D.; Bogucki, J.; Galkowski, D.; Kaczmarczyk, R.; Feldo, M.; Adamczyk-Cioch, M.; Kocki, J. Modulation of multidrug resistance gene expression by coumarin derivatives in human leukemic cells. Oxid. Med. Cell. Longev. 2017, 2017, 1–13. [Google Scholar] [CrossRef]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. ABC transporters in cancer: More than just drug efflux pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef]
- Nakanishi, T.; Ross, U.D. Breast cancer resistance protein (BCRP/ABCG2): Its role in multidrug resistance and regulation of its gene expression. Chin. J. Cancer 2012, 31, 73–99. [Google Scholar] [CrossRef]
- Deng, L.; Tatebe, S.; Lin-Lee, Y.-C.; Ishikawa, T.; Kuo, M.T. MDR and MRP Gene Families as Cellular Determinant Factors for Resistance to Clinical Anticancer Agents; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2002; Volume 112, pp. 49–66. [Google Scholar]
- Mirzaei, S.A.; Dehkordi, N.G.; Ghamghami, M.; Amiri, A.; Abdolahinia, E.D.; Elahian, F. ABC-transporter blockage mediated by xanthotoxin and bergapten is the major pathway for chemo sensitization of multidrug-resistant cancer cells. Toxicol. Appl. Pharmacol. 2017, 337, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Sui, M.; Huang, Y.; Park, B.H.; Davidson, N.E.; Fan, W. Estrogen receptor α mediates breast cancer cell resistance to paclitaxel through inhibition of apoptotic cell death. Cancer Res. 2007, 67, 5337–5344. [Google Scholar] [CrossRef] [PubMed]
- Acharya, R.; Chacko, S.; Bose, P.; Lapenna, A.; Pattanayak, S.P. Structure Based Multitargeted Molecular docking analysis of selected furanocoumarins against breast cancer. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Yousafzai, N.A.; Wang, H.; Wang, Z.; Zhu, Y.; Zhu, L.; Jin, H.; Wang, X. Exosome mediated multidrug resistance in cancer. Am. J. Cancer Res. 2018, 8, 2210–2226. [Google Scholar]
- Hsieh, M.J.; Chen, M.K.; Yu, Y.Y.; Sheu, G.T.; Chiou, H.L. Psoralen reverses docetaxel-induced multidrug resistance in A549/D16 human lung cancer cells lines. Phytomedicine 2014, 21, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lee, E.J.; Lee, J.H.; Yang, W.M.; Nam, D.; Lee, J.H.; Lee, S.G.; Um, J.Y.; Shim, B.S.; Ahn, K.S. Simvastatin in combination with bergamottin potentiates TNF-induced apoptosis through modulation of NF-κB signalling pathway in human chronic myelogenous leukemia. Pharm. Biol. 2016, 54, 2050–2060. [Google Scholar] [CrossRef]
- Alzahrani, A.M.; Rajendran, P. The Multifarious Link between Cytochrome P450s and Cancer. Oxid. Med. Cell. Longev. 2020, 2020, 3028387. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Chang, C.; Hsu, C.; Tsai, M.; Cheng, H.; Leong, M.K.; Sung, P.; Chen, J.; Weng, C. Natural compounds as potential adjuvants to cancer therapy: Preclinical evidence. Br. J. Pharmacol. 2019, 177, 1409–1423. [Google Scholar] [CrossRef]
- Ohnishi, A.; Matsuo, H.; Yamada, S.; Takanaga, H.; Morimoto, S.; Shoyama, Y.; Ohtani, H.; Sawada, Y. Effect of furanocoumarin derivatives in grapefruit juice on the uptake of vinblastine by Caco-2 cells and on the activity of cytochrome P450 3A4. Br. J. Pharmacol. 2000, 130, 1369–1377. [Google Scholar] [CrossRef]
- Kleiner, H.E.; Reed, M.J.; DiGiovanni, J. Naturally Occurring Coumarins Inhibit Human Cytochromes P450 and Block Benzo[a]pyrene and 7,12-Dimethylbenz[a]anthracene DNA Adduct Formation in MCF-7 Cells. Chem. Res. Toxicol. 2003, 16, 415–422. [Google Scholar] [CrossRef]
- Herynk, M.H.; Fuqua, S.A. Estrogen receptors in resistance to hormone therapy. Retin. Degener. Dis. 2007, 608, 130–143. [Google Scholar] [CrossRef]
- Zheng, Y.; Jiang, K. Antitumor effect of imperatorin enhances cytotoxicity of doxorubicin to HeLa cells. Chin. J. Pathophysiol. 2015, 31, 1578–1583. [Google Scholar]
- Jakubowicz-Gil, J.; Paduch, R.; Ulz, Z.; Badziul, D.; Głowniak, K.; Gawron, A. Cell death in HeLa cells upon imperatorin and cisplatin treatment. Folia Histochem. Cytobiol. 2012, 50, 381–391. [Google Scholar] [CrossRef]
- Hu, J.; Xu, C.; Cheng, B.; Jin, L.; Li, J.; Gong, Y.; Lin, W.; Pan, Z.; Pan, C.-W. Imperatorin acts as a cisplatin sensitizer via downregulating Mcl-1 expression in HCC chemotherapy. Tumor Biol. 2015, 37, 331–339. [Google Scholar] [CrossRef]
- Viola, G.; Fortunato, E.; Cecconet, L.; Disarò, S.; Basso, G. Induction of apoptosis in Jurkat cells by photoexcited psoralen derivatives: Implication of mitochondrial dysfunctions and caspases activation. Toxicol. In Vitro 2007, 21, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Panno, M.L.; Giordano, F.; Mastroianni, F.; Palma, M.G.; Bartella, V.; Carpino, A.; Aquila, S.; Andò, S. Breast cancer cell survival signal is affected by bergapten combined with an ultraviolet irradiation. FEBS Lett. 2010, 584, 2321–2326. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, M.; Sakanaka, M.; Taniguchi, M.; Baba, K.; Kimura, Y. Anti-tumor effects of various furocoumarins isolated from the roots, seeds and fruits of Angelica and Cnidium species under ultraviolet A irradiation. J. Nat. Med. 2013, 68, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.S. The risk of squamous cell and basal cell cancer associated with psoralen and ultraviolet A therapy: A 30-year prospective study. J. Am. Acad. Dermatol. 2012, 66, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Dowdy, J.C.; Sayre, R.M. Melanoma risk from dietary furocoumarins: How much more evidence is required? J. Clin. Oncol. 2016, 34, 636–637. [Google Scholar] [CrossRef] [PubMed]
- DelRosso, G.; Savoia, P. Effectiveness and Safety of Topical Phototherapy in the Treatment of Dermatological Diseases. Photomed. Adv. Clin. Pract. 2017, 137–151. [Google Scholar] [CrossRef]
- Oldham, M.; Yoon, S.W.; Fathi, Z.; Beyer, W.F.; Adamson, J.; Liu, L.; Alcorta, D.; Xia, W.; Osada, T.; Liu, C.; et al. X-Ray Psoralen Activated Cancer Therapy (X-PACT). PLoS ONE 2016, 11, e0162078. [Google Scholar] [CrossRef] [PubMed]
- Melough, M.M.; Chun, O.K. Dietary furocoumarins and skin cancer: A review of current biological evidence. Food Chem. Toxicol. 2018, 122, 163–171. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Hollenberg, P.F.; Woolf, T.F. Compositions Containing Bergamottin for Increasing the Oral Bioavailability of Pharmaceutical Agents. U.S. Patent No. 6,509,371B1, 21 June 2003. [Google Scholar]
- Brigger, I.; Dubernet, C.; Couvreur, P.; Patrick, C. Nanoparticles in cancer therapy and diagnosis. Adv. Drug Deliv. Rev. 2012, 64, 24–36. [Google Scholar] [CrossRef]
- Bor, G.; Azmi, I.D.M.; Yaghmur, A. Nanomedicines for cancer therapy: Current status, challenges and future prospects. Ther. Deliv. 2019, 10, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Cai, T.; Li, Q.; Huang, Y.; Liu, Q.; Wang, B.; Xia, X.; Wang, Q.; Whitney, J.C.C.; Cole, S.P.C.; et al. Preparation of psoralen polymer-lipid hybrid nanoparticles and their reversal of multidrug resistance in MCF-7/ADR cells. Drug Deliv. 2018, 25, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Mottaghipisheh, J.; Nové, M.; Spengler, G.; Kúsz, N.; Hohmann, J.; Csupor, D. Antiproliferative and cytotoxic activities of furocoumarins of Ducrosia anethifolia. Pharm. Biol. 2018, 56, 658–664. [Google Scholar] [CrossRef]



{kind=link}
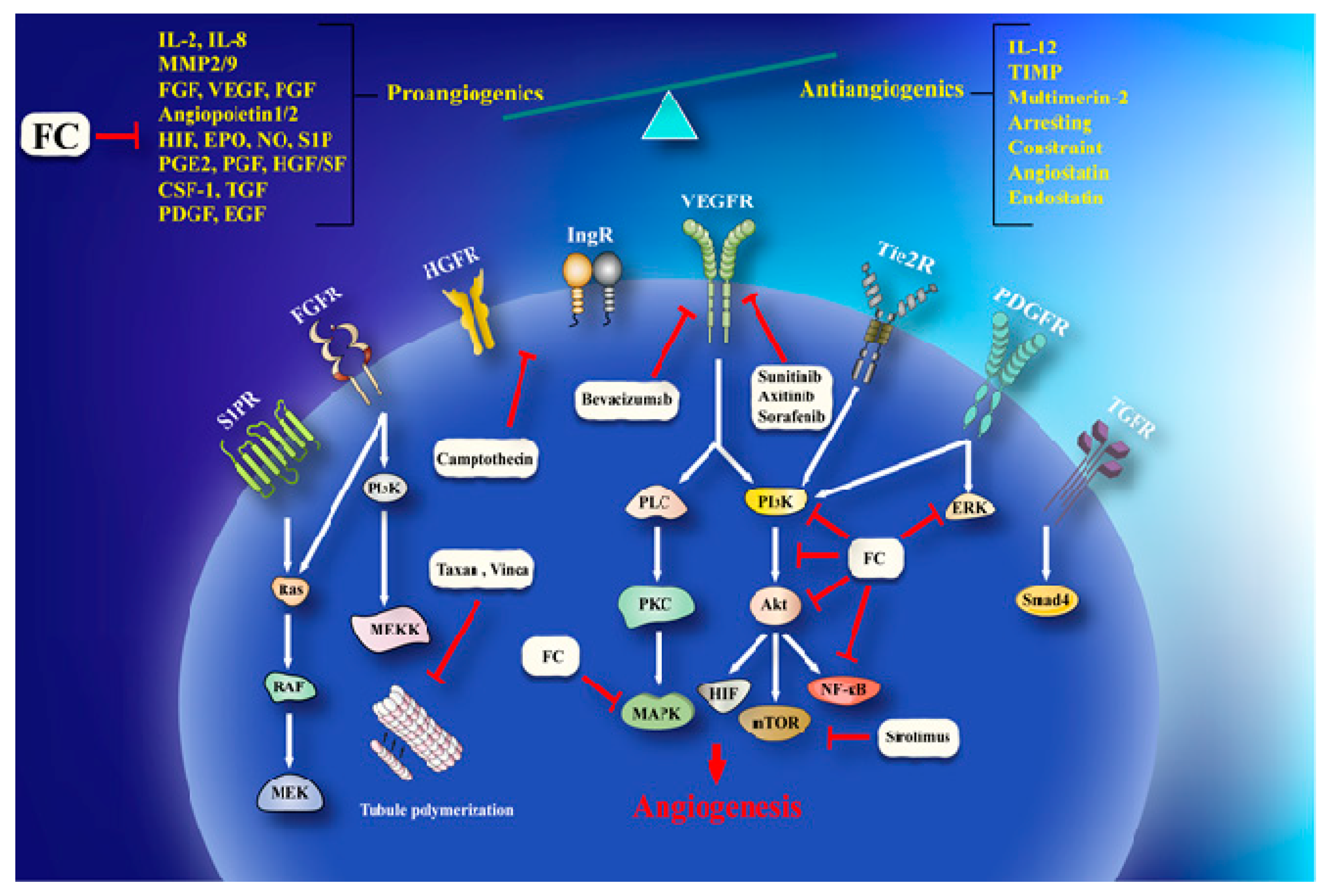{kind=link}
| Furanocoumarins | Dietary Sources [2,3,4] | In Vitro | In Vivo | Anticancer Mechanisms | References | ||
|---|---|---|---|---|---|---|---|
| Cell Lines | Cytotoxic Concentration | Experimental Model | Dose | ||||
 Angelicin | Parsnip | HeLa and SiHa | IC30 = 27.8 µM; IC50 = 38.2 µM | — | — | Atg3,7 and 12-5 ↑, G1/G0 arrest | [5] |
| A549 (NSCLC) | IC50~ 50 µmol | — | — | NF-κB inactivation G2/M phase arrest, cyclinB1 ↓, cyclin E1 ↓, and Cdc2 ↓; Bcl2 ↓; caspase 3,9 ↑, Bax↑ JNK ↑, ERK ↑; MMP2, MMP9 ↓, E-cadherin ↑ | [6] | ||
| SH-SY5Y | IC50 = 49.56 μM | — | — | caspase 3,9 ↑, Bcl-2 ↓, Bcl-xL↓, and Mcl-1↓ | [7] | ||
| Caki | IC50 = angelicin (50–100 μM) and TRAIL (50 ng/mL) | — | — | caspase 3↑, c-FLIP ↓ | [8] | ||
| HepG2 and Huh-7 | IC50 = 90 ± 6.565 (HepG2); 60 ± 4.256 μM (Huh-7) | — | — | PI3K/Akt inhibition, Bcl-2↓ | [9] | ||
| — | — | Mouse liver xenograft model (BALB/c-nu/nu mice) | 50 mg/kg (16 days) | cancer cell growth ↓; p-VEGFR ↓ | [9] | ||
 Bergamottin | Carrot, cumin, dill, fig, grapefruit, lemon, lime, orange, parsley, parsnip | A549, H1299 | IC50 = 50–100 μM | — | — | EMT inhibition, JNK ↑; PI3K ↓, Akt↓, and mTOR kinases ↓ | [10] |
| U87 and U251 | IC50 = 2–10 μM | — | — | NF-κB inactivation, MMP9 ↓, STAT3 inhibition, Rac1 inactivation | [11] | ||
| HT-1080 | IC50 = 5–50 μM | — | — | NF-κB inactivation; MMP2, MMP9↓ | [12] | ||
| HT-29 and RKO | IC50 = 12.5 µM | — | — | caspase-3,8,9↑; PARP ↑; G2/M phase arrest, cyclinA ↓, cyclin B1↓, and Cdc2 ↓ | [13] | ||
| A549 (NSCLC) | IC50 = 50 μM | — | — | G2/M phase arrest, cyclinA ↓, cyclin B1↓, and Cdc2 ↓ | [14] | ||
| — | — | Mouse lung xenograft model (BALB/c nude mice) | 100 mg/kg (18 days) | ↓ cancer cell growth | [14] | ||
 Bergapten or 5-Methoxypsoralen (5-MOP) | Anise, carrot, caraway, celeriac, celery, coriander, cumin, dill, fig, grapefruit, lemon, lime, orange, parsley, parsnip, turnip | DLD-1 and LoVo | IC50 = 30 and 50 μM | — | — | G2/M phase arrest, cyclin E ↓, Cdk2 inhibition; AKT inhibition; p53 ↑, p21↑, PTEN↑ | [15] |
| MCF-7 | IC50 = 50 μM | — | — | G1-phase phase arrest, P53 ↑; AKT inhibition | [16] | ||
| Hep-G2 | IC50 = 25–50 mM | — | — | G2/M phase arrest, Cdk1 inhibition | [17] | ||
| IC50 = 25–100 μM | — | — | G2-M phase arrest, P53 ↑ and P21 ↑ | [18] | |||
| A549 (NSCLC) | 79.1 ± 2.8% (50 μM) | — | — | G1-phase arrest, P53 ↑ | [19] | ||
| NCI-H460 | 74.5 ± 3.1% (50 μM) | — | — | ||||
| MCF7 and ZR-75 | 50 μM | — | — | PTEN ↑, Beclin1 ↑, PI3KIII ↑, UVRAG ↑, AMBRA, LC3-I to LC3-II | [20] | ||
| HepG2 | 50 mM | — | — | PI3K/AKT inhibition, LXR (α and β)↑ | [21] | ||
| — | — | NDEA induced liver cancer (Wistar rats) | 25 and 50 mg/kg (14 days) | ↓ cancer cell growth | |||
 Bergaptol | Carrot, grapefruit, lemon, lime, parsley, parsnip | MCF-7 | IC50 = 52.2 µM | — | — | G1-phase arrest; caspase 3,9 ↑, Bax↑, Bcl2 ↓,MMP2 ↓, MMP9 ↓, cyt c release ↑ | [22] |
 Byakangelicol | Lemon | HCT-15 | IC50 = 18.1 ± 0.3 μg·mL−1 | — | — | ↓ cell viability | [23] |
| A549 (NSCLC) | IC50 =14.3 ± 0.2 μg·mL−1 | — | — | ||||
| SK-OV-3 | IC50 = 20.2 ± 0.3 μg·mL−1 | — | — | ||||
| SK-MEL-2 | IC50 = 21.2 ± 0.3 μg·mL−1 | — | — | ||||
| XF498 | IC50 = 28.4 ± 0.3 μg·mL−1 | — | — | ||||
 Cnidicin | Lemon | HCT-15 | IC50 = 7.0 ± 0.2 μg·mL−1 | — | — | ↓ cell viability | [23] |
| A549 (NSCLC) | IC50 = 6.8 ± 0.1 μg·mL−1 | — | — | ||||
| SK-OV-3 | IC50 = 8.8 ± 0.2 μg·mL−1 | — | — | ||||
| SK-MEL-2 | IC50 = 8.8 ± 0.2 μg·mL−1 | — | — | ||||
| XF498 | IC50 = 7.2 ± 0.3 μg·mL−1 | — | — | ||||
 Feroniellin A | Feroniella lucida roots | A549 | 0.25 mM | — | — | NF-κB inactivation Atg5 ↑, Beclin1 ↑, mTOR inactivation LC3-I to LC3-II | [24] |
 Furanocoumarin A | Fructus liquidambaris | A549 | IC50 = 65.28 ± 5.36 μM | — | — | P53 ↑,Bax↑, Bcl2 ↓, caspase 3 ↑ | [25] |
 Imperatorin | Lime, parsley | SGC-7901 | IC50 = 62.6 μM | — | — | promoting G1-phase arrest; PI3K/Akt/m-TOR signaling ↓ | [26] |
| HT-29 | IC50 = 78 µM | — | — | G1-phase arrest; P53 ↑ and P21 ↑; caspase 3,7 ↑; Bcl2 ↓ | [27] | ||
| HL-60 | 10 μM | — | — | caspase 3,9 ↑; cyt c release ↑; Bcl2 ↓, PARP cleavage | [28] | ||
| H23 | 10 μg/mL | — | — | P53 ↑,Bax↑, Mcl-1↓ | [29] | ||
| HeLa | 200 μM for imperatorin and 5 μM for cisplatin | — | — | Hsp27 and Hsp72 ↓,LC3 cleavage | [30] | ||
| HCT116 | 150 μM | mTOR↓, p70S6K ↓, 4E-BP1↓, MAPK↓, HIF-1α inhibition | [31] | ||||
| HCT-15 | IC50 = 19.4 ± 0.3 μg·mL−1 | — | — | ↓ cell viability | [23] | ||
| A549 | IC50 = 16.4 ± 0.3 μg·mL−1 | — | — | ||||
| SK-OV-3 | IC50 =13.7 ± 0.3 μg·mL−1 | — | — | ||||
| SK-MEL-2 | IC50 = 14.5 ± 0.2 μg·mL−1 | — | — | ||||
| XF498 | IC50 = 12.3 ± 0.5 μg·mL−1 | — | — | ||||
 Isoimperatorin | Lime, parsley | SGC-7901 | IC50 = 18.75 μg·mL−1 | SGC-7901 cell-induced xenograft model (mice) | 10 mg/kg (20 days) | G1-phase arrest; caspase 3,9 ↑; Bax↑, Bcl2 ↓, Survivin↓ | [32] |
| DU145 | 100 µM | — | — | G1-phase arrest | [33] | ||
| HCT-15 | IC50 = 5.6 ± 0.3 μg·mL−1 | — | — | ↓ cell viability | [23] | ||
| A549 (NSCLC) | IC50 = 12.2 ± 0.4 μg·mL−1 | — | — | ||||
| SK-OV-3 | IC50 = 6.8 ± 0.3 μg·mL−1 | — | — | ||||
| SK-MEL-2 | IC50 = 9.9 ± 0.2 μg·mL−1 | — | — | ||||
| XF498 | IC50 = 10.7 ± 0.3 μg·mL−1 | — | — | ||||
 Marmesin | Broad bean, blackberry, raspberry | U937 | IC50 = 40 µM | Mice | 30mg/kg (30 days) | Bax↑, Bcl2 ↓, Bax/Bcl-2 ratio ↑; G2/M-phase arrest | [34] |
 Methoxsalen/Xanthotoxin (8-MOP) | Anise, carrot, caraway celeriac, celery, cumin, dill, grapefruit, lemon, lime, parsley, parsnip, turnip | HepG2 | 100 µM | — | — | Bax/Bcl-2 ratio ↑, MMP2 ↓, MMP9 ↓, ERK1/2 pathway inhibition, cyt c release ↑, AIF transposition ↑ | [35] |
| SK-N-AS and SW620 | IC50 = 56.3 µM for SK-N-AS and 88.5 µM for SW620 | — | — | Caspase-3,8,9↑PI3K/AKT pathway ↓, Bcl2 ↓, Bax/Bcl-2 ratio ↑ | [36] | ||
 Oxypeucedanin | Lime, parsley | L5178Y (mouse T-cell lymphoma cells) | IC50 = 40.33 ± 0.63 µM | — | — | caspase 3,9 ↑; cyt c release ↑; Bcl2 ↓ | [37] |
| HCT-15 | IC50 = 3.4 ± 0.3 μg·mL−1 | — | — | ↓ cell viability | [23] | ||
| A549 (NSCLC) | IC50 = 9.5 ± 0.3 μg·mL−1 | — | — | ||||
| SK-OV-3 | IC50 = 19.3 ± 0.3 μg·mL−1 | — | — | ||||
| SK-MEL-2 | IC50 = 16.5 ± 0.2 μg·mL−1 | — | — | ||||
| XF498 | IC50 = 16.1 ± 0.5 μg·mL−1 | — | — | ||||
 (+)-oxypeucedanin hydrate | Angelica dahurica roots | A549, HCT-15, SK-MEL-2, SK-OV-3, XF498 | IC50 > 30 μg·mL−1 | — | — | ||
 Psoralen | Carrot, celeriac, celery, cilantro, cumin, dill, fig, grapefruit, lemon, lime, parsley, parsnip | KBv200 | 75.3% (80 μg·mL−1) | — | — | NF-κB inactivation G1/S phase arrest, c-FLIP, and IAP inhibition | [38] |
| K562 | 92.4% (80 μg·mL−1) | — | — | ||||
| MCF-7 | 17.32 ± 4.28% (8 μg·mL−1) | — | — | G1/G0 phase arrest | [39] | ||
| SMMC7721 | 40 μM | — | — | G1 phase arrest, cyclin E↓, Bax↑, Bcl2 ↓, ER stress, CHOP induction, GADD34↑, ATF4↑, GRP78↑, GRP94 | [40] | ||
| MCF-7/ADR | IC10 = 8 μg/mL | P-gp efflux function inhibition | [41] | ||||
| IC50 = 25.59 ± 1.74 µg/mL | P-gp ATPase activity inhibition | [42] | |||||
| 43.0 µM | EMT inhibition | ||||||
| 107.5 µM | G0/G1 phase arrest | ||||||
 Sphondin | Parsnip | A549 | 50 μM | — | — | NF-κB inactivation | [43] |
 5-geranyloxy-7-methoxy-coumarin | Lime | SW480 | 25 µM (67%) | — | — | caspase-3,8↑; Bcl2 ↓; p38 MAPK phosphorylation inhibition; G1/G0 phase arrest | [44] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, S.; Khan, H.; Aschner, M.; Mirzae, H.; Küpeli Akkol, E.; Capasso, R. Anticancer Potential of Furanocoumarins: Mechanistic and Therapeutic Aspects. Int. J. Mol. Sci. 2020, 21, 5622. https://doi.org/10.3390/ijms21165622
Ahmed S, Khan H, Aschner M, Mirzae H, Küpeli Akkol E, Capasso R. Anticancer Potential of Furanocoumarins: Mechanistic and Therapeutic Aspects. International Journal of Molecular Sciences. 2020; 21(16):5622. https://doi.org/10.3390/ijms21165622
Chicago/Turabian StyleAhmed, Salman, Haroon Khan, Michael Aschner, Hamed Mirzae, Esra Küpeli Akkol, and Raffaele Capasso. 2020. "Anticancer Potential of Furanocoumarins: Mechanistic and Therapeutic Aspects" International Journal of Molecular Sciences 21, no. 16: 5622. https://doi.org/10.3390/ijms21165622
APA StyleAhmed, S., Khan, H., Aschner, M., Mirzae, H., Küpeli Akkol, E., & Capasso, R. (2020). Anticancer Potential of Furanocoumarins: Mechanistic and Therapeutic Aspects. International Journal of Molecular Sciences, 21(16), 5622. https://doi.org/10.3390/ijms21165622