CD20-Mimotope Peptides: A Model to Define the Molecular Basis of Epitope Spreading

Abstract
1. Introduction
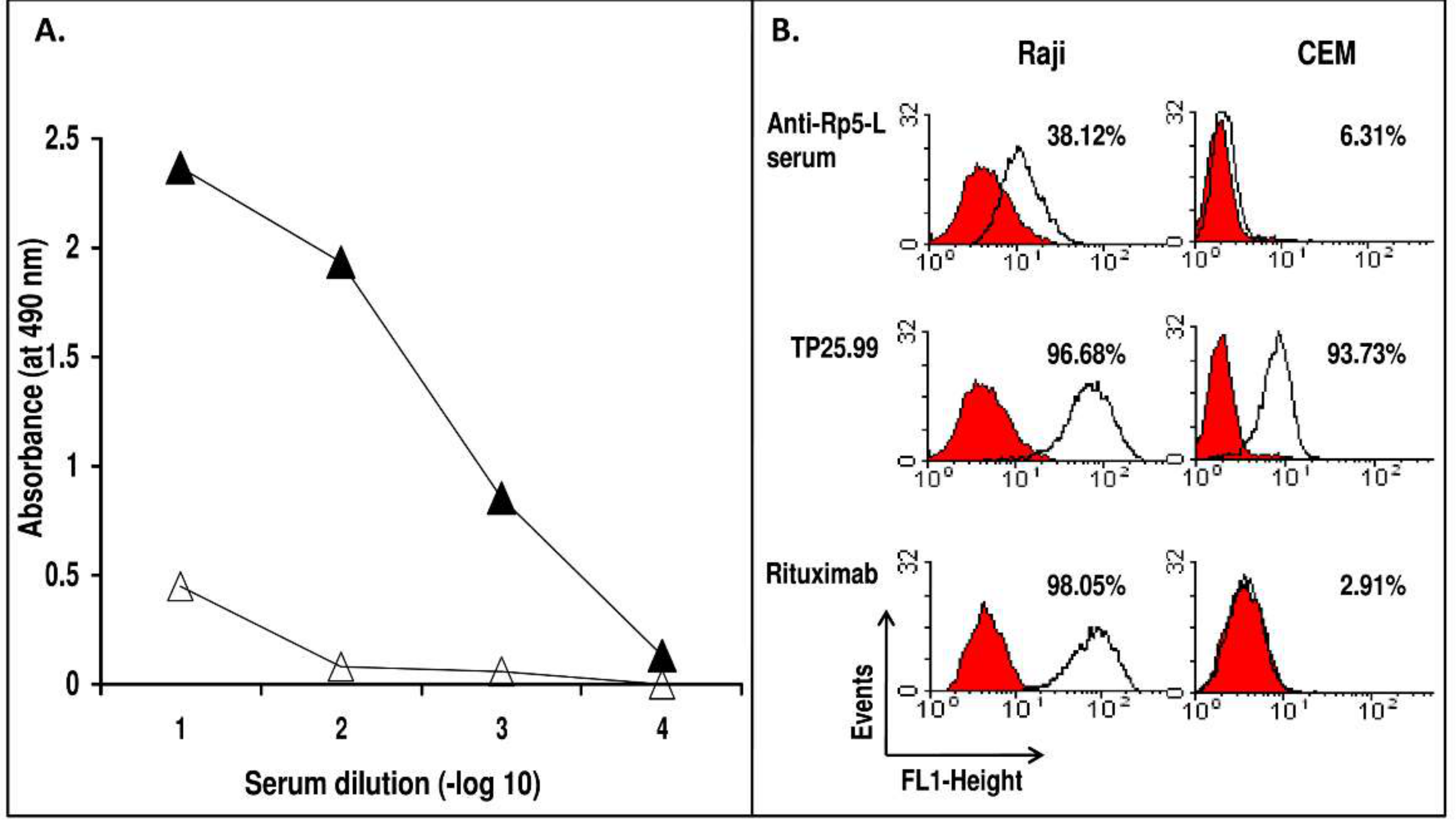
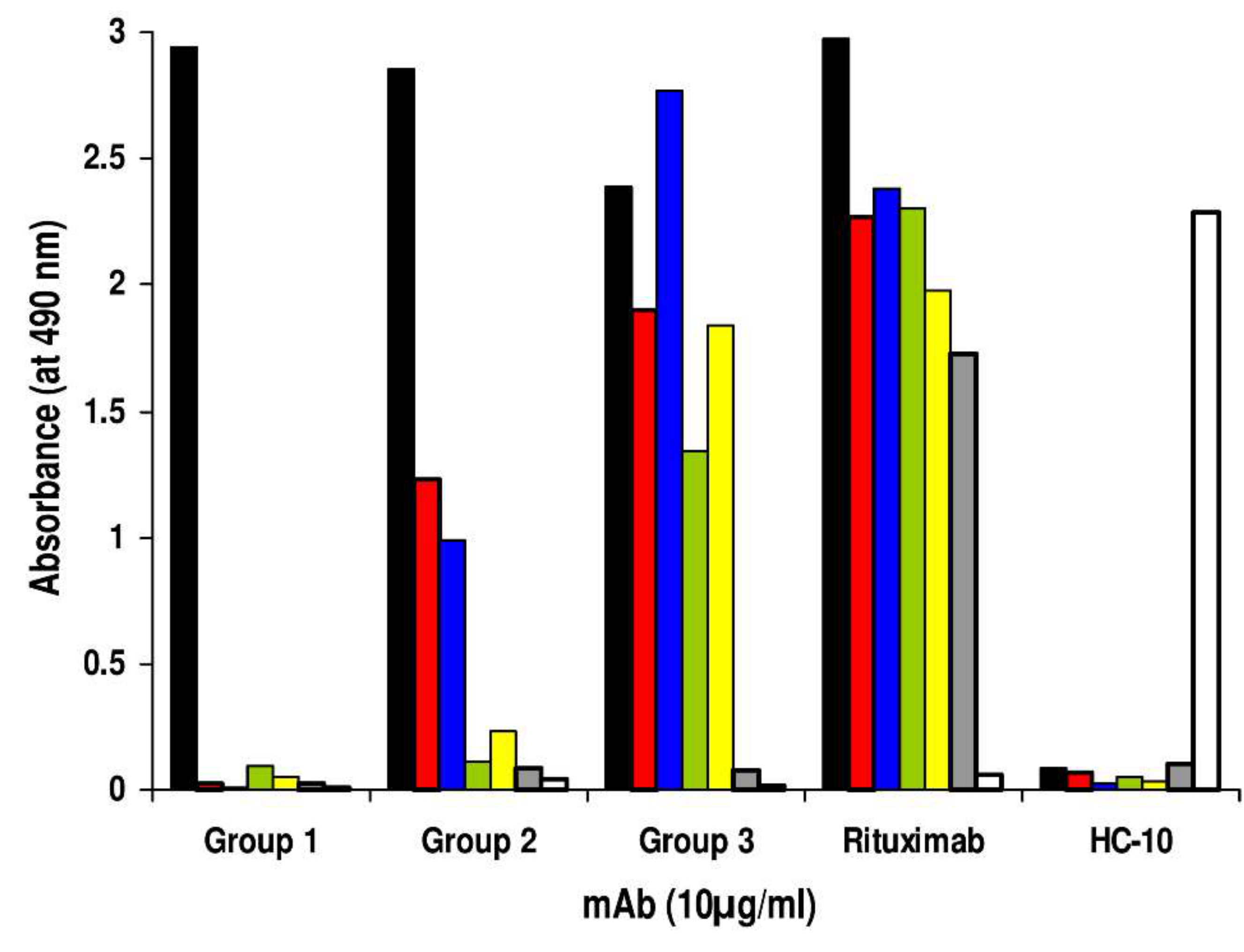
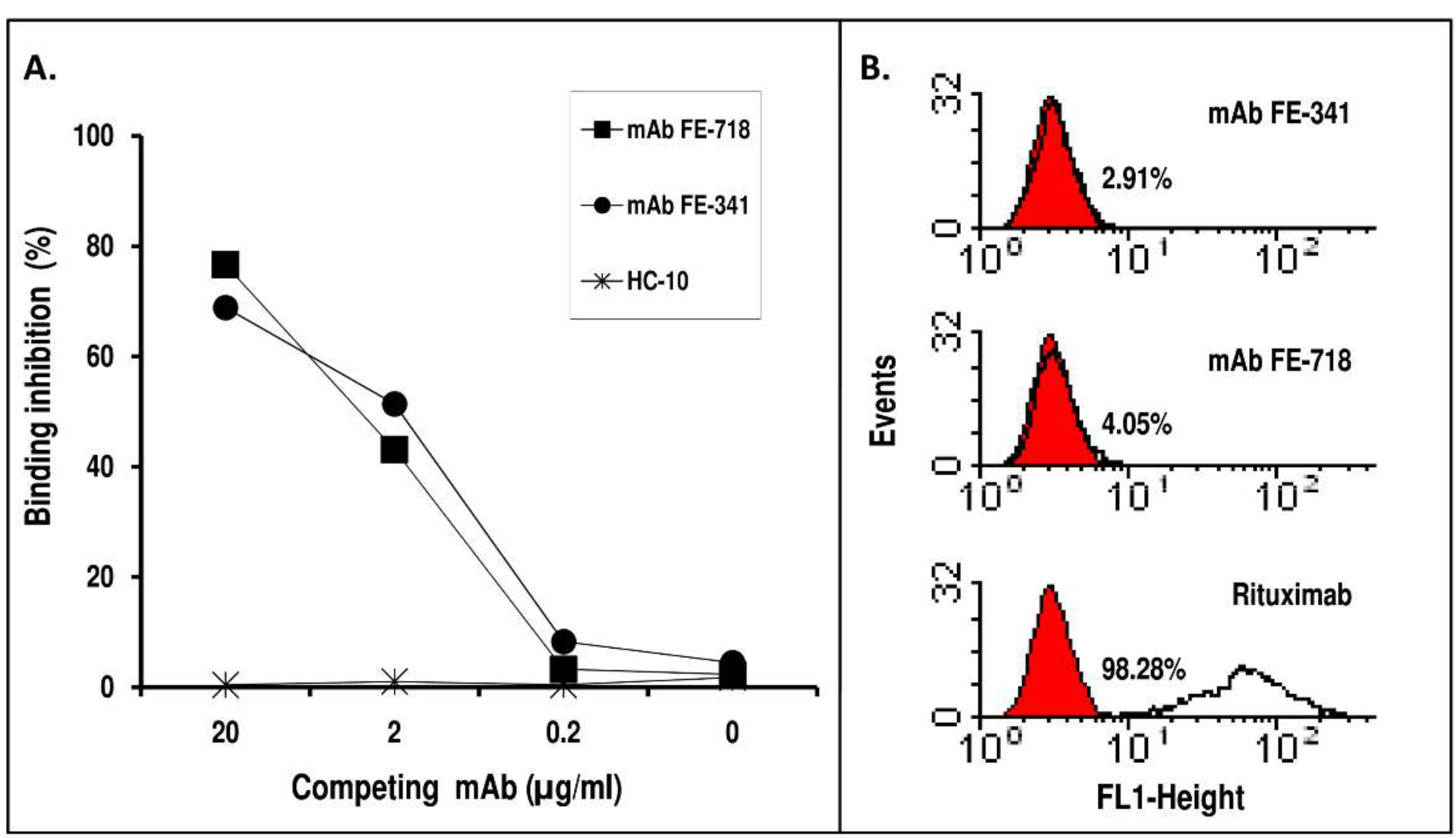
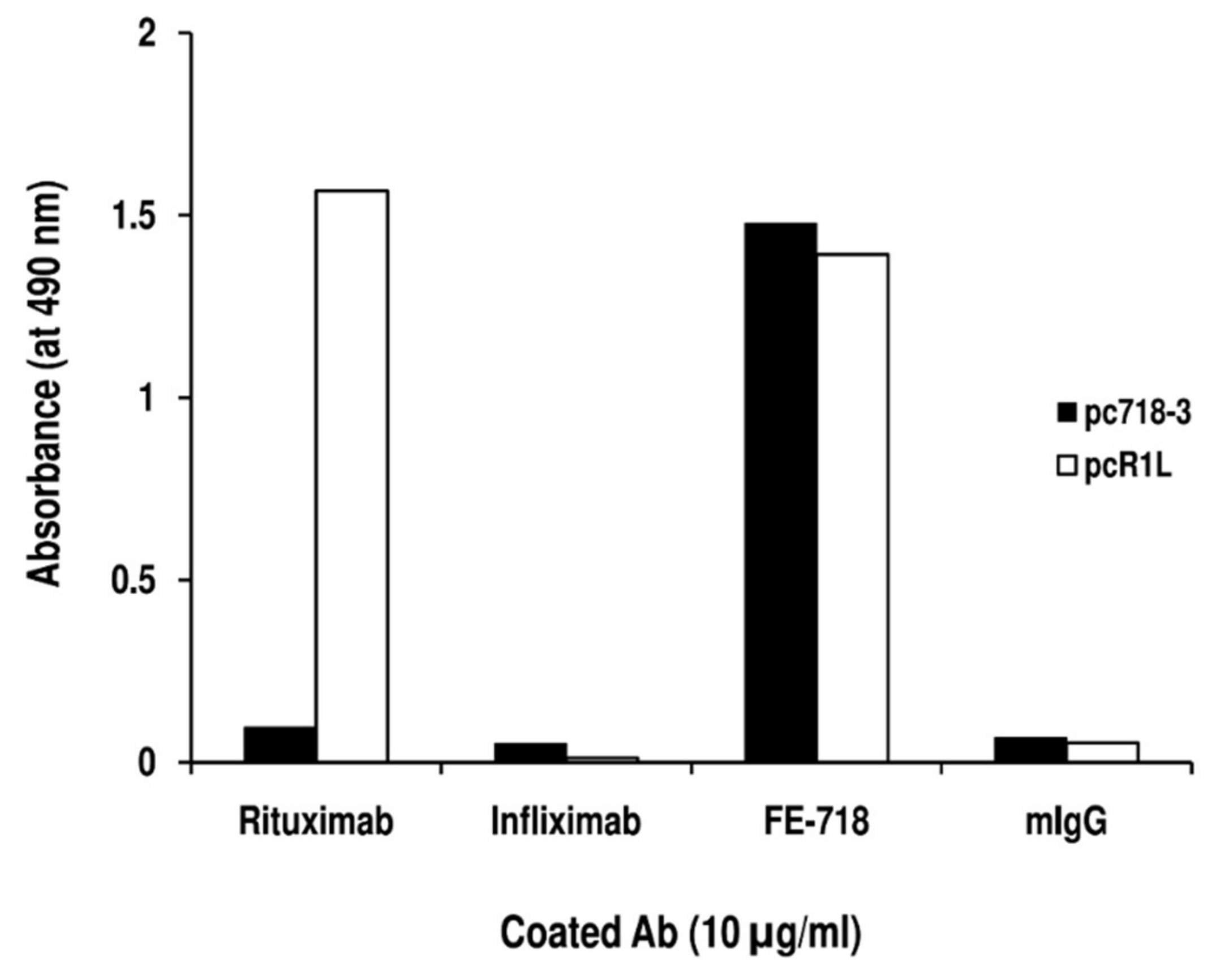
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cells
4.3. Conventional Reagents, mAb, Peptides and PDPL
4.4. Preparation of Anti-Rp5-L mAb
4.5. Binding Assay
4.6. Flow Cytometry
4.7. Biopanning, Immunoscreening, and Sequence Analysis
Author Contributions
Funding
Acknowledgments
Conflict of Interest
References
- Bucktrout, S.L.; Bluestone, J.A.; Ramsdell, F. Recent advances in immunotherapies: From infection and autoimmunity, to cancer, and back again. Genome Med. 2018, 10, 79. [Google Scholar] [CrossRef]
- Alfonso, M.; Díaz, A.; Hernández, A.M.; Pérez, A.; Rodríguez, E.; Bitton, R.; Pérez, R.; Vázquez, A.M. An anti-idiotype vaccine elicits a specific response to N-glycolyl sialic acid residues of glycoconjugates in melanoma patients. J. Immunol. 2002, 168, 2523–2529. [Google Scholar] [CrossRef]
- De Cerio, A.L.; Zabalegui, N.; Rodriguez-Calvillo, M.; Inoges, S.; Bendandi, M. Anti-idiotype antibodies in cancer treatment. Oncogene 2007, 26, 3594–3602. [Google Scholar] [CrossRef]
- Kieber-Emmons, T.; Monzavi-Karbassi, B.; Pashov, A.; Saha, S.; Murali, R.; Kohler, H. The promise of the anti-idiotype concept. Front. Oncol. 2012, 2, 196. [Google Scholar] [CrossRef]
- Ladjemi, M.Z. Anti-idiotypic antibodies as cancer vaccines: Achievements and future improvements. Front. Oncol. 2012, 2, 158. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, A.M.; Rodreguez-Zhurbenko, N.; Lopez, A.M. Anti-ganglioside anti-idiotypic vaccination: More than molecular mimicry. Front. Oncol. 2012, 2, 170. [Google Scholar] [CrossRef] [PubMed]
- Cacciavillano, W.; Sampor, C.; Venier, C.; Gabri, M.R.; de Dávila, M.T.; Galluzzo, M.L.; Guthmann, M.D.; Fainboim, L.; Alonso, D.F.; Chantada, G.L. A Phase I Study of the Anti-Idiotype Vaccine Racotumomab in Neuroblastoma and Other Pediatric Refractory Malignancies. Pediatr. Blood Cancer 2015, 62, 2120–2124. [Google Scholar] [CrossRef] [PubMed]
- Wada, S.; Yada, E.; Ohtake, J.; Fujimoto, Y.; Uchiyama, H.; Yoshida, S.; Sasada, T. Current status and future prospects of peptide-based cancer vaccines. Immunotherapy 2016, 8, 1321–1333. [Google Scholar] [CrossRef]
- Klausen, U.; Holmberg, S.; Holmström, M.O.; Jørgensen, N.G.D.; Grauslund, J.H.; Svane, I.M.; Andersen, M.H. Novel Strategies for Peptide-Based Vaccines in Hematological Malignancies. Front. Immunol. 2018, 9, 2264. [Google Scholar] [CrossRef] [PubMed]
- Obara, W.; Kanehira, M.; Katagiri, T.; Kato, R.; Kato, Y.; Takata, R. Present status and future perspective of peptide-based vaccine therapy for urological cancer. Cancer Sci. 2018, 109, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Shi, J.; Yang, C. HER2-Based Immunotherapy for Breast Cancer. Cancer Biother. Radiopharm. 2018, 33, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Prete, M.; Perosa, F.; Favoino, E.; Dammacco, F. Biological therapy with monoclonal antibodies: A novel treatment approach to autoimmune disease. Clin. Exp. Med. 2005, 5, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Perosa, F.; Prete, M.; Racanelli, V.; Dammacco, F. CD20-depleting therapy in autoimmune diseases: From basic research to the clinic. J. Intern. Med. 2010, 267, 260–277. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.K.; Livingston, P.O.; Agus, D.B.; Pinilla-Ibarz, J.; Zelenetz, A.; Scheinberg, D.A. Vaccination with CD20 peptides induces a biologically active, specific immune response in mice. Blood 2002, 99, 3748–3755. [Google Scholar] [CrossRef] [PubMed]
- Perosa, F.; Favoino, E.; Caragnano, M.A.; Prete, M.; Dammacco, F. CD20: A target antigen for immunotherapy of autoimmune diseases. Autoimmun. Rev. 2005, 4, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Perosa, F.; Favoino, E.; Caragnano, M.A.; Dammacco, F. CD20 mimicry by a mAb rituximab-specific linear peptide: A potential tool for active immunotherapy of autoimmune diseases. Ann. N. Y. Acad. Sci. 2005, 1051, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Perosa, F.; Favoino, E.; Caragnano, M.A.; Dammacco, F. Generation of biologically active linear and cyclic peptides has revealed a unique fine specificity of rituximab and its possible cross-reactivity with acid sphingomyelinase-like phosphodiesterase 3b precursor. Blood 2006, 107, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Perosa, F.; Favoino, E.; Vicenti, C.; Guarnera, A.; Racanelli, V.; De Pinto, V.; Dammacco, F. Two structurally different rituximab-specific CD20 mimotope peptides reveal that rituximab recognizes two different CD20-associated epitopes. J. Immunol. 2009, 182, 416–423. [Google Scholar] [CrossRef]
- Fornoni, A.; Sageshima, J.; Wei, C.; Merscher-Gomez, S.; Aguillon-Prada, R.; Jauregui, A.N.; Li, J.; Mattiazzi, A.; Ciancio, G.; Chen, L.; et al. Rituximab targets podocytes in recurrent focal segmental glomerulosclerosis. Sci. Transl. Med. 2011, 3, 85ra46. [Google Scholar] [CrossRef]
- Perosa, F.; Favoino, E.; Vicenti, C.; Merchionne, F.; Dammacco, F. Identification of an antigenic and immunogenic motif expressed by two 7-mer rituximab-specific cyclic peptide mimotopes: Implication for peptide-based active immunotherapy. J. Immunol. 2007, 179, 7967–7974. [Google Scholar] [CrossRef]
- Favoino, E.; Prete, M.; Marzullo, A.; Millo, E.; Shoenfeld, Y.; Perosa, F. CD20-Mimotope Peptide Active Immunotherapy in Systemic Lupus Erythematosus and a Reappraisal of Vaccination Strategies in Rheumatic Diseases. Clin. Rev. Allergy Immunol. 2017, 52, 217–233. [Google Scholar] [CrossRef]
- Hou, Y.; Gu, X.X. Development of peptide mimotopes of lipooligosaccharide from nontypeable Haemophilus influenzae as vaccine candidates. J. Immunol. 2003, 170, 4373–4379. [Google Scholar] [CrossRef]
- Wagner, S.; Hafner, C.; Allwardt, D.; Jasinska, J.; Ferrone, S.; Zielinski, C.C.; Scheiner, O.; Wiedermann, U.; Pehamberger, H.; Breiteneder, H. Vaccination with a human high molecular weight melanoma-associated antigen mimotope induces a humoral response inhibiting melanoma cell growth in vitro. J. Immunol. 2005, 174, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Krepler, C.; Allwardt, D.; Latzka, J.; Strommer, S.; Scheiner, O.; Pehamberger, H.; Wiedermann, U.; Hafner, C.; Breiteneder, H. Reduction of human melanoma tumor growth in severe combined immunodeficient mice by passive transfer of antibodies induced by a high molecular weight melanoma-associated antigen mimotope vaccine. Clin. Cancer Res. 2008, 14, 8178–8183. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vanderlugt, C.L.; Miller, S.D. Epitope spreading in immune-mediated diseases: Implications for immunotherapy. Nat. Rev. Immunol. 2002, 2, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Disis, M.L.; Goodell, V.; Schiffman, K.; Knutson, K.L. Humoral epitope-spreading following immunization with a HER-2/neu peptide based vaccine in cancer patients. J. Clin. Immunol. 2004, 24, 571–578. [Google Scholar] [CrossRef]
- Walter, S.; Weinschenk, T.; Stenzl, A.; Zdrojowy, R.; Pluzanska, A.; Szczylik, C.; Staehler, M.; Brugger, W.; Dietrich, P.-Y.; Mendrzyk, R.; et al. Multipeptide immune response to cancer vaccine IMA901 after single-dose cyclophosphamide associates with longer patient survival. Nat. Med. 2012, 18, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Gulley, J.L.; Madan, R.A.; Pachynski, R.; Mulders, P.; Sheikh, N.A.; Trager, J.; Drake, C.G. Role of Antigen Spread and Distinctive Characteristics of Immunotherapy in Cancer Treatment. J. Natl. Cancer Inst. 2017, 109, djw261. [Google Scholar] [CrossRef]
- Vanderlugt, C.J.; Miller, S.D. Epitope spreading. Curr. Opin. Immunol. 1996, 8, 831–836. [Google Scholar] [CrossRef]
- Latzka, J.; Gaier, S.; Hofstetter, G.; Balazs, N.; Smole, U.; Ferrone, S.; Scheiner, O.; Breiteneder, H.; Pehamberger, H.; Wagner, S. Specificity of mimotope-induced anti-high molecular weight-melanoma associated antigen (HMW-MAA) antibodies does not ensure biological activity. PLoS ONE 2011, 6, e19383. [Google Scholar] [CrossRef]
- Beenhouwer, D.O.; May, R.J.; Valadon, P.; Scharff, M.D. High affinity mimotope of the polysaccharide capsule of Cryptococcus neoformans identified from an evolutionary phage peptide library. J. Immunol. 2002, 169, 6992–6999. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Hsu, J.C.; Kieber-Emmons, T.; Wang, X.; Ferrone, S. Human tumor associated antigen mimicry by xenoantigens, anti-idiotypic antibodies and peptide mimics: Implications for immunotherapy of malignant diseases. Cancer Chemother. Biol. Response Modif. 2005, 22, 769–787. [Google Scholar] [PubMed]
- Van Regenmortel, M.H. Molecular design versus empirical discovery in peptide-based vaccines. Coming to terms with fuzzy recognition sites and ill-defined structure-function relationships in immunology. Vaccine 1999, 18, 216–221. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H. Antigenicity and immunogenicity of synthetic peptides. Biologicals 2001, 29, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.J.; Katsumata, Y.; Ascherman, D.P. Structural and thermodynamic approach to peptide immunogenicity. PLoS Comput. Biol. 2008, 4, e1000231. [Google Scholar] [CrossRef] [PubMed]
- Disis, M.L.; Schiffman, K.; Guthrie, K.; Salazar, L.G.; Knutson, K.L.; Goodell, V.; dela Rosa, C.; Cheever, M.A. Effect of dose on immune response in patients vaccinated with an her-2/neu intracellular domain protein—Based vaccine. J. Clin. Oncol. 2004, 22, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Polyak, M.J.; Deans, J.P. Alanine-170 and proline-172 are critical determinants for extracellular CD20 epitopes; heterogeneity in the fine specificity of CD20 monoclonal antibodies is defined by additional requirements imposed by both amino acid sequence and quaternary structure. Blood 2002, 99, 3256–3262. [Google Scholar] [CrossRef]
- Niederfellner, G.; Lammens, A.; Mundigl, O.; Georges, G.J.; Schaefer, W.; Schwaiger, M.; Franke, A.; Wiechmann, K.; Jenewein, S.; Slootstra, J.W.; et al. Epitope characterization and crystal structure of GA101 provide insights into the molecular basis for type I/II distinction of CD20 antibodies. Blood 2011, 118, 358–367. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, Y.; He, W.; Wang, W.; Chen, Y.; Zhang, S.; Ma, Y.; Gohda, J.; Ishida, T.; Walter, T.S.; et al. A RANKL mutant used as an inter-species vaccine for efficient immunotherapy of osteoporosis. Sci. Rep. 2015, 5, 14150. [Google Scholar] [CrossRef]
- Perosa, F.; Luccarelli, G.; Prete, M.; Favoino, E.; Ferrone, S.; Dammacco, F. Beta 2-microglobulin-free HLA class I heavy chain epitope mimicry by monoclonal antibody HC-10-specific peptide. J. Immunol. 2003, 171, 1918–1926. [Google Scholar] [CrossRef]
- Perosa, F.; Carbone, R.; Ferrone, S.; Dammacco, F. Purification of human immunoglobulins by sequential precipitation with caprylic acid and ammonium sulphate. J. Immunol. Methods 1990, 128, 9–16. [Google Scholar] [CrossRef]
- Favoino, E.; Digiglio, L.; Cuomo, G.; Favia, I.E.; Racanelli, V.; Valentini, G.; Perosa, F. Autoantibodies recognizing the amino terminal 1–17 segment of CENP-A display unique specificities in systemic sclerosis. PLoS ONE 2013, 8, e61453. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Origin | Peptide | |
|---|---|---|
| Denomination | Sequence | |
| Rituximab-specific peptides | ||
| Rp5-L | QDKLTQWPKWLEg | |
| Rp1-L | WPRWLEN | |
| Rp10-L | ITPWPHWLERSSg | |
| ASMLPD | ||
| Rev-pASMLPD | 163SLWPKWLEAIQ153 | |
| pASMLPD | 153QIAELWKPWLS163 | |
| Human CD20 | ||
| RpCD20-L | 165YNCEPANPSEKNSPSTQYCY184 |
| Phage Clone Insert # | Clones, n (%) | Deduced Amino Acid Insert Sequence (a) | Specificity of Reactivity (A490nm) | |
|---|---|---|---|---|
| mAb | mIgG | |||
| pc718-1 | 5 (27.7%) | WPHVLPE | 1.754 ± 0.01 | 0.123 ± 0.002 |
| pc718-2 | 2 (11.1%) | KWPQYLS | 1.833 ± 0.14 | 0.144 ± 0.07 |
| pc718-3 | 11 (61.1%) | MWPKWLP | 1.92 ± 0.044 | 0.107 ± 0.02 |
| FE-718 motif | WP—-L | |||
| pc341-1 | 4 (22.2%) | SLKMPHWPHLLP | 1.644 ± 0.01 | 0.167 ± 0.002 |
| pc341-2 | 1 (5.5%) | QHVNLARWPWQL | 1.834 ± 0.021 | 0.111 ± 0.013 |
| pc341-3 | 10 (55.5%) | TQLGWPHSIGDA | 1.421 ± 0.08 | 0.172 ± 0.1 |
| pc341-4 | 2 (11.1%) | HSSWPRHLDPPQ | 1.962 ± 0.013 | 0.069 ± 0.002 |
| pc341-5 | 1 (5.5%) | QWPNELRNSGLS | 1.718 ± 0.032 | 0.098 ± 0.011 |
| FE-341 motif | WP—-l | |||
| Rituximab motif | WP-WLE | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Favoino, E.; Prete, M.; Catacchio, G.; Conteduca, G.; Perosa, F. CD20-Mimotope Peptides: A Model to Define the Molecular Basis of Epitope Spreading. Int. J. Mol. Sci. 2019, 20, 1920. https://doi.org/10.3390/ijms20081920
Favoino E, Prete M, Catacchio G, Conteduca G, Perosa F. CD20-Mimotope Peptides: A Model to Define the Molecular Basis of Epitope Spreading. International Journal of Molecular Sciences. 2019; 20(8):1920. https://doi.org/10.3390/ijms20081920
Chicago/Turabian StyleFavoino, Elvira, Marcella Prete, Giacomo Catacchio, Giuseppina Conteduca, and Federico Perosa. 2019. "CD20-Mimotope Peptides: A Model to Define the Molecular Basis of Epitope Spreading" International Journal of Molecular Sciences 20, no. 8: 1920. https://doi.org/10.3390/ijms20081920
APA StyleFavoino, E., Prete, M., Catacchio, G., Conteduca, G., & Perosa, F. (2019). CD20-Mimotope Peptides: A Model to Define the Molecular Basis of Epitope Spreading. International Journal of Molecular Sciences, 20(8), 1920. https://doi.org/10.3390/ijms20081920