Rosiglitazone Enhances Browning Adipocytes in Association with MAPK and PI3-K Pathways During the Differentiation of Telomerase-Transformed Mesenchymal Stromal Cells into Adipocytes


Abstract
1. Introduction
2. Results
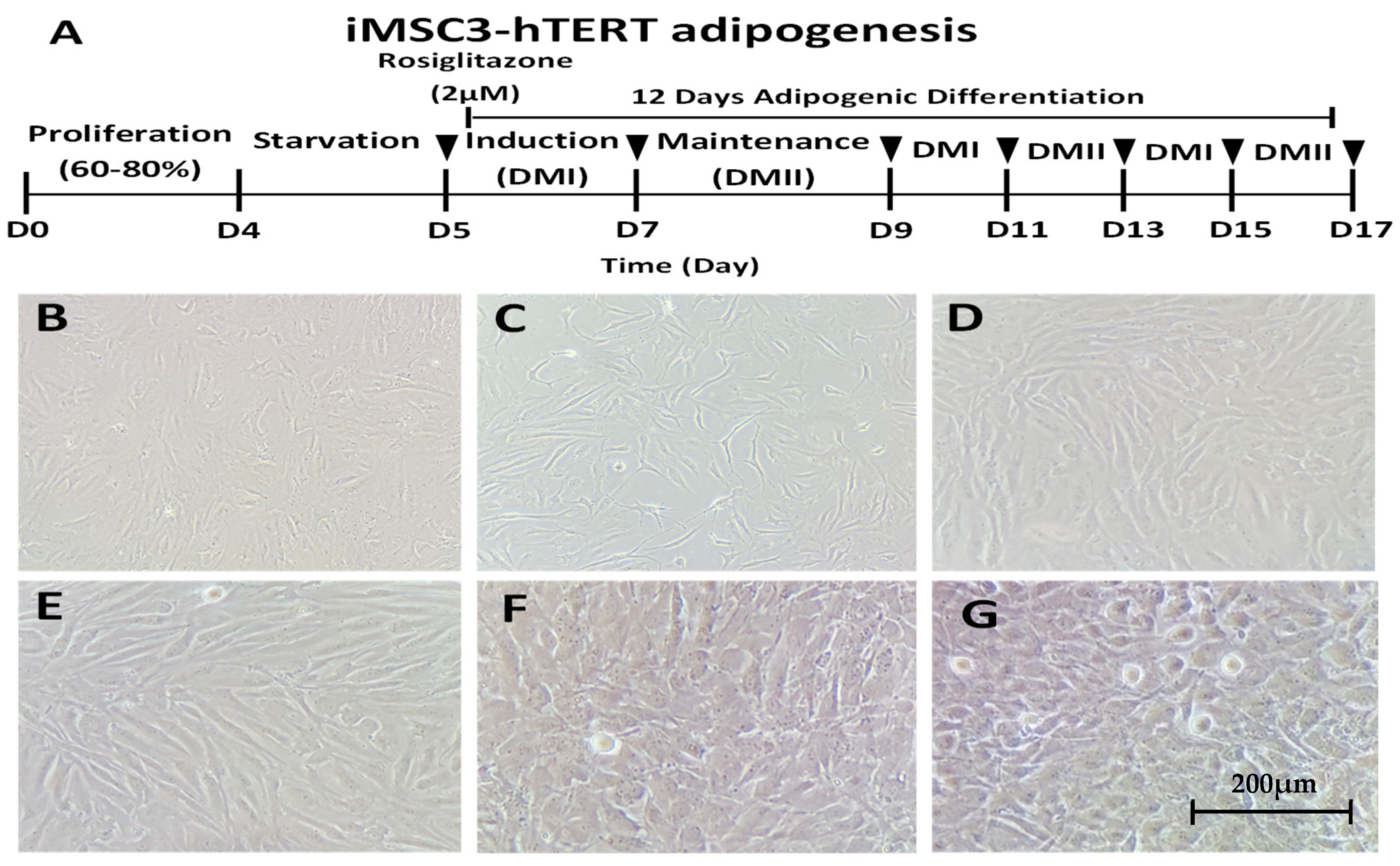
2.1. In Vitro Differentiation of Telomerase-Transformed MSCs into Adipocytes
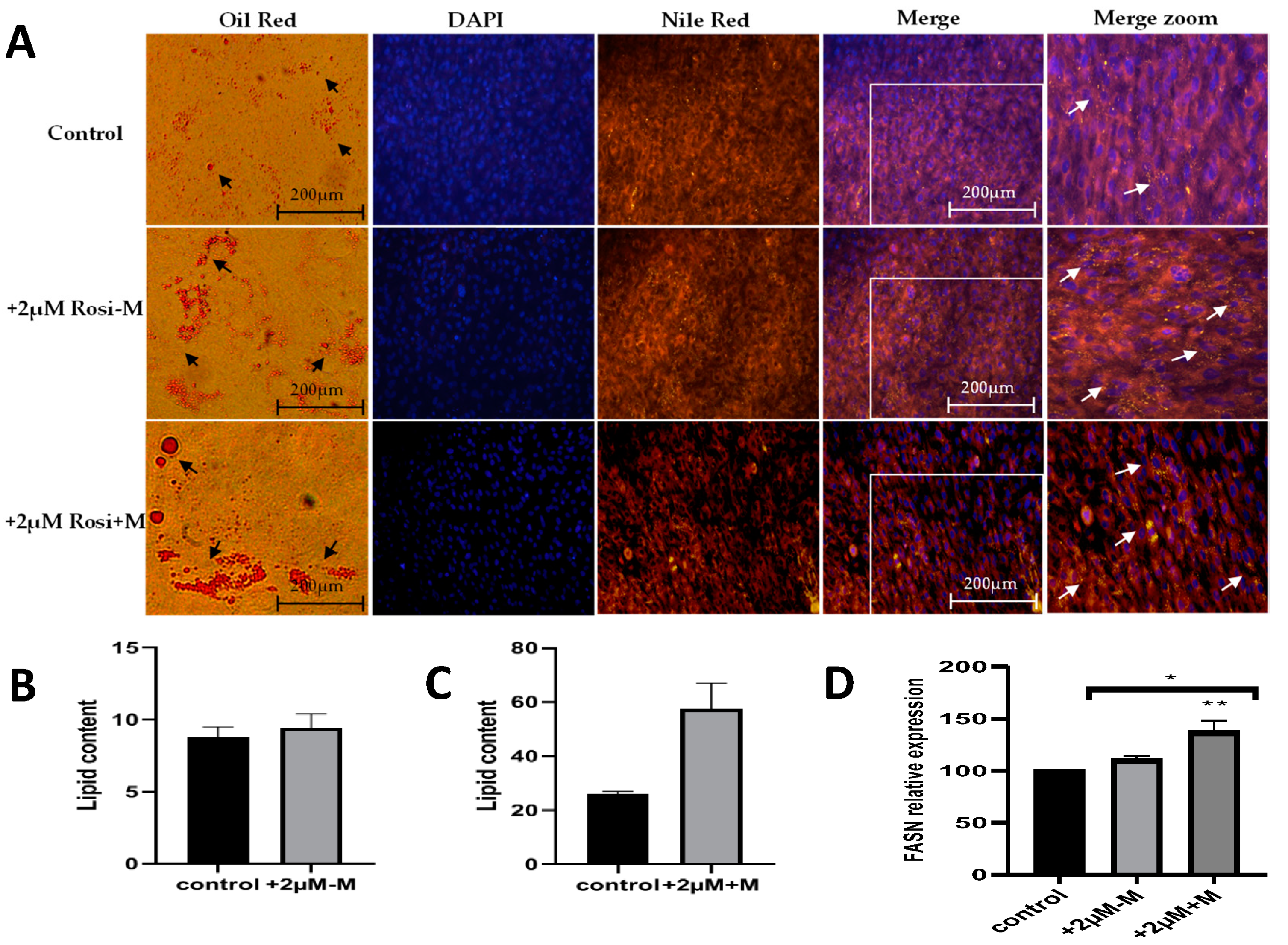
2.2. Rosiglitazone Increases Fatty Acid Synthesis and Thus Lipid Content in Differentiated Adipocytes
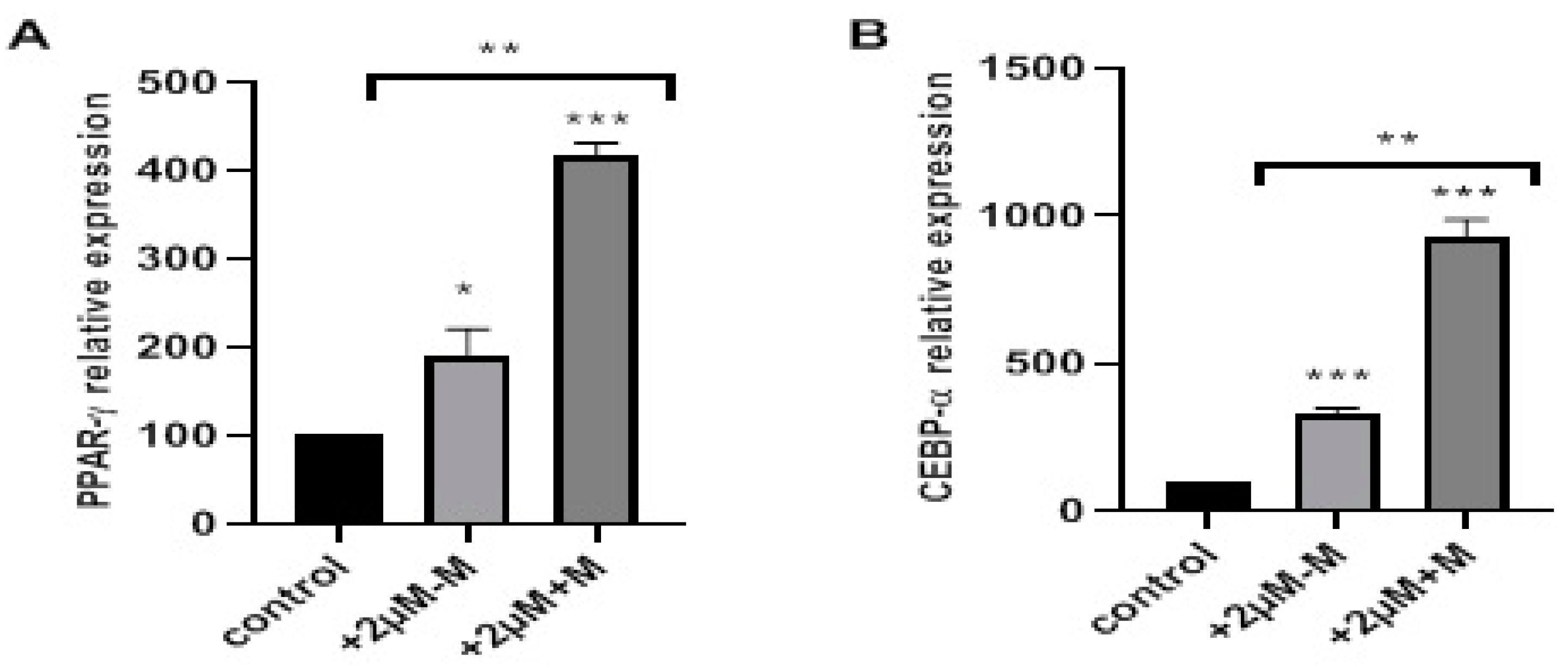
2.3. Rosiglitazone Enhances Adipogenic Differentiation by Targeting PPAR-γ and C/EBP-α in a Time-Dependent Manner
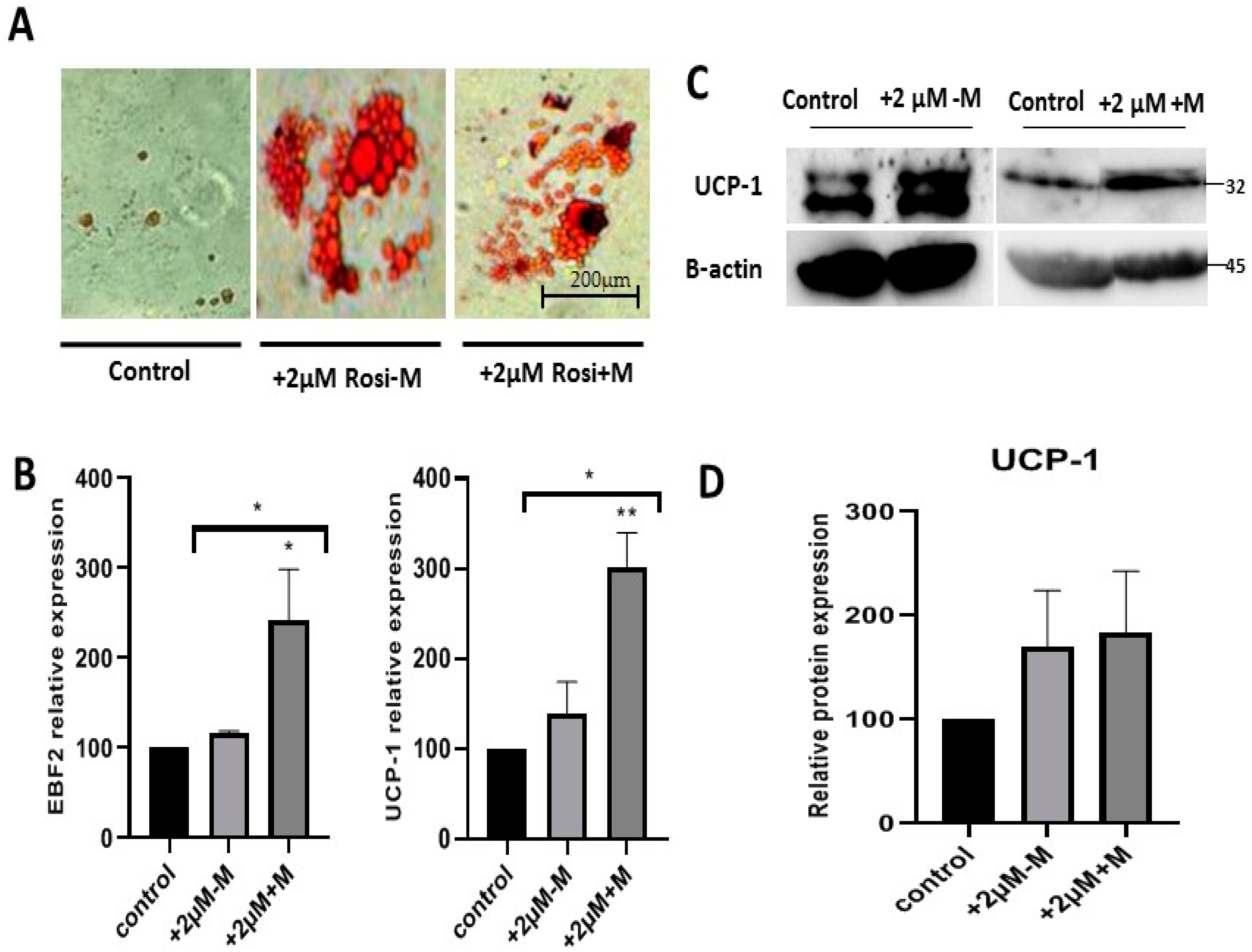
2.4. Rosiglitazone Enhances Browning Characteristics and Markers in Differentiated Adipocytes in a Time-Dependent Manner
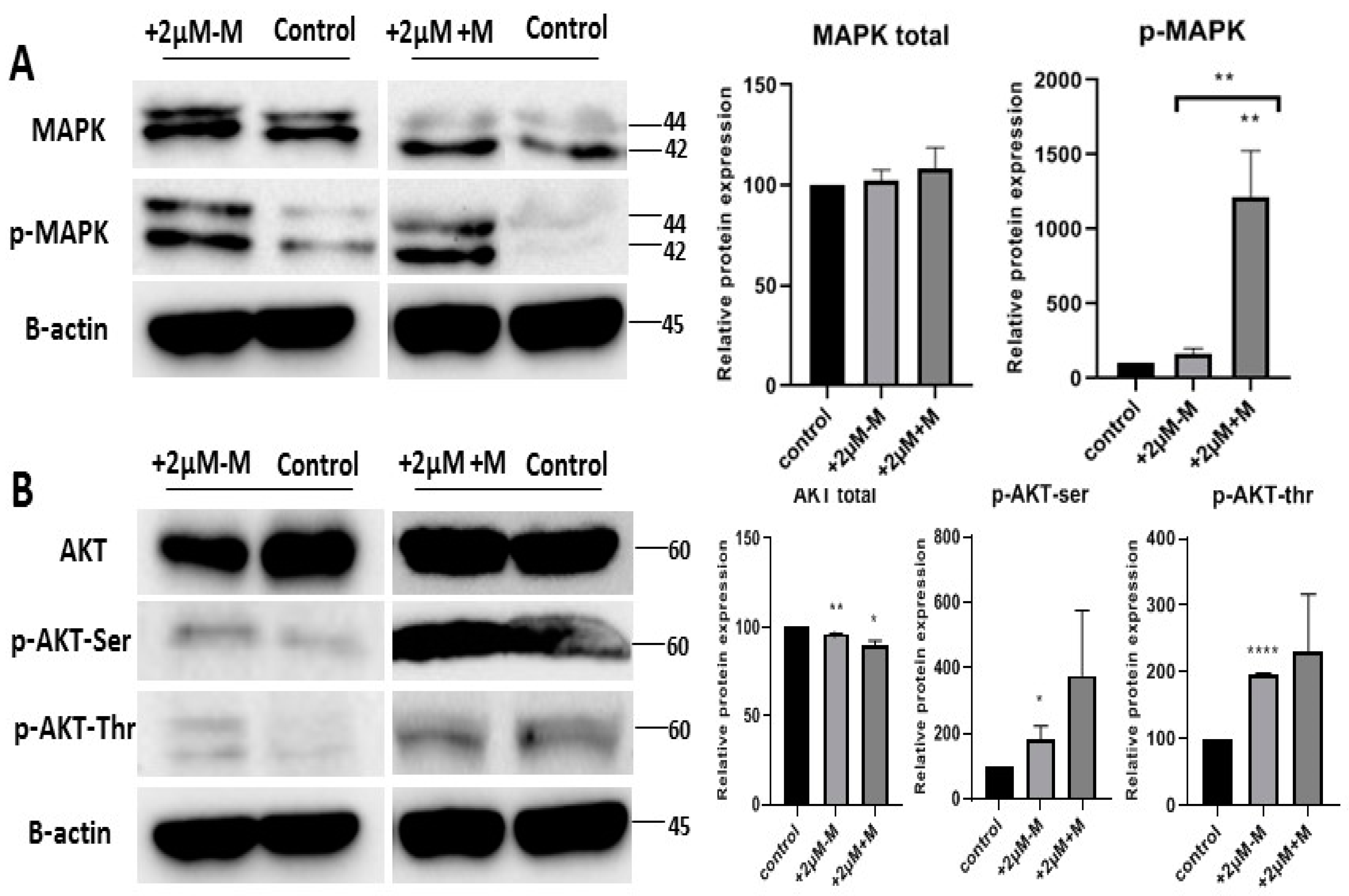
2.5. Rosiglitazone Enhances the Browning Effect by Activation of MAPK and PI3K Pathways
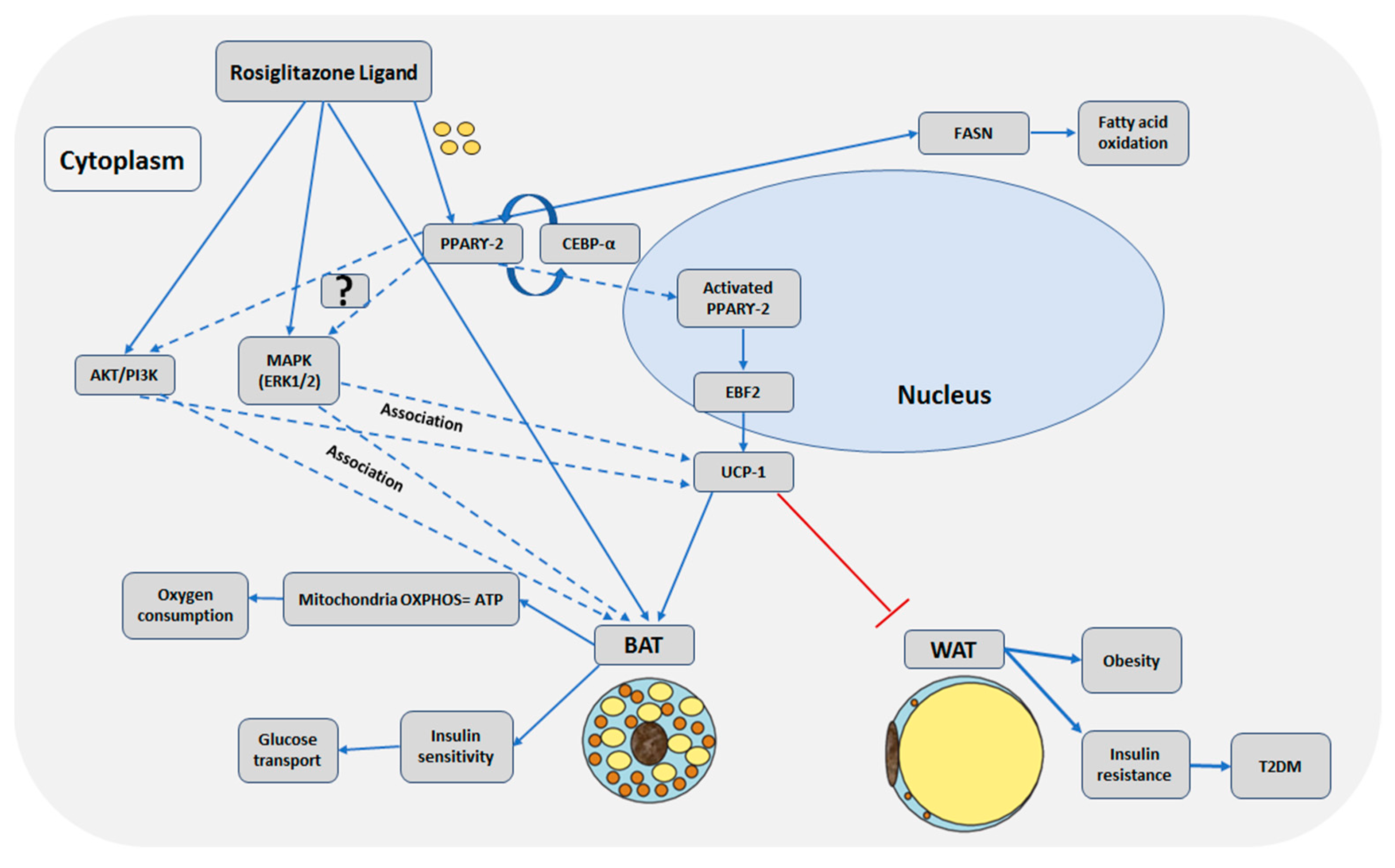
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture
4.2. Differentiation of MSCs into Adipocytes
4.3. Nile Red and DAPI Staining
4.4. Oil-O Red Staining
4.5. Lipid Content Determination
4.6. RNA Extraction and cDNA Synthesis
4.7. Gene Expression Study by RT-PCR
4.8. Protein Extraction and Immunoblotting
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal Stromal Cells |
| iMSC3 | Immortalized Human Bone Marrow Mesenchymal Stromal Cells- hTERT |
| PPAR-γ | Peroxisome proliferator-activated receptor gamma 2 |
| C/EBP-α | CCAAT/enhancer binding protein alpha |
| EBF2 | Early B cell factor 2 |
| UCP-1 | Uncoupling protein 1 |
| FASN | Fatty acid synthase |
| MAPK | Mitogen-activated protein kinases |
| PI3K/AKT | Phosphoinositide 3-kinase/Protein Kinase B |
References
- Kwak, D.H.; Lee, J.H.; Kim, T.; Ahn, H.S.; Cho, W.K.; Ha, H.; Hwang, Y.H.; Ma, J.Y. Aristolochia Manshuriensis Kom Inhibits Adipocyte Differentiation by Regulation of ERK1/2 and Akt Pathway. PLoS ONE 2012, 7, e49530. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, J.; Ali, M.; Jung, H.; Min, B.; Choi, R.; Kim, G.; Jung, H. Anti-Adipogenic Effect of Epiberberine is Mediated by Regulation of the Raf/MEK1/2/ERK1/2 and AMPKα/Akt Pathways. Arch. Pharm. Res. 2015, 38, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Koppen, A.; Kalkhoven, E. Brown vs. White Adipocytes: The PPARc Coregulator Story. Koppen, A.; Kalkhoven, E. Brown vs white adipocytes: The PPARγ coregulator story. FEBS Lett. 2010, 584, 3250–3259. [Google Scholar] [CrossRef]
- Lu, Y.; Fujioka, H.; Joshi, D.; Li, Q.; Sangwung, P.; Hsieh, P.; Zhu, J.; Torio, J.; Sweet, D.; Wang, L. Mitophagy is Required for Brown Adipose Tissue Mitochondrial Homeostasis during Cold Challenge. Sci. Rep. 2018, 8, 8251. [Google Scholar] [CrossRef]
- Cedikova, M.; Kripnerová, M.; Dvorakova, J.; Pitule, P.; Grundmanova, M.; Babuska, V.; Mullerova, D.; Kuncova, J. Mitochondria in White, Brown, and Beige Adipocytes. Stem Cells Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Shabalina, I.; Petrovic, N.; de Jong, J.A.; Kalinovich, A.; Cannon, B.; Nedergaard, J. UCP1 in Brite/Beige Adipose Tissue Mitochondria is Functionally Thermogenic. Cell Rep. 2013, 5, 1196–1203. [Google Scholar] [CrossRef]
- Merlin, J.; Sato, M.; Chia, L.Y.; Fahey, R.; Pakzad, M.; Nowell, C.J.; Summers, R.J.; Bengtsson, T.; Evans, B.A.; Hutchinson, D.S. Rosiglitazone and a Β3-Adrenoceptor Agonist are both Required for Functional Browning of White Adipocytes in Culture. Front. Endocrinol. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Than, A.; Xu, S.; Li, R.; Leow, M.; Sun, L.; Chen, P. Angiotensin Type 2 Receptor Activation Promotes Browning of White Adipose Tissue and Brown Adipogenesis. Signal Transduct Target Ther. 2017, 2, 17022. [Google Scholar] [CrossRef] [PubMed]
- Ridderstråle, M.; Groop, L. Genetic Dissection of Type 2 Diabetes. Mol. Cell. Endocrinol. 2009, 297, 10–17. [Google Scholar] [CrossRef]
- Groop, L.; Pociot, F. Genetics of Diabetes – are we Missing the Genes Or the Disease? Mol. Cell. Endocrinol. 2014, 382, 726–739. [Google Scholar] [CrossRef]
- Scioli, M.G.; Bielli, A.; Gentile, P.; Mazzaglia, D.; Cervelli, V.; Orlandi, A. The Biomolecular Basis of Adipogenic Differentiation of Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2014, 15, 6517–6526. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Genever, P. Regulation of Mesenchymal Stem Cell Differentiation. Adv. Exp. Med. Biol. 2013, 786, 213. [Google Scholar] [PubMed]
- Janderová, L.; McNeil, M.; Murrell, A.N.; Mynatt, R.L.; Smith, S.R. Human Mesenchymal Stem Cells as an in Vitro Model for Human Adipogenesis. Obes. Res. 2003, 11, 65–74. [Google Scholar] [CrossRef] [PubMed]
- MacDougald, O.A.; Rosen, E.D. Adipocyte Differentiation from the Inside Out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar]
- Morrison, R.F.; Farmer, S.R. Hormonal Signaling and Transcriptional Control of Adipocyte Differentiation. J. Nutr. 2000, 130, 3121S. [Google Scholar] [CrossRef]
- Muruganandan, S.; Roman, A.; Sinal, C. Adipocyte Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells: Cross Talk with the Osteoblastogenic Program. Cell. Mol. Life Sci. 2009, 66, 236–253. [Google Scholar] [CrossRef]
- Ntambi, J.M.; Young-Cheul, K. Adipocyte Differentiation and Gene Expression. J. Nutr. 2000, 130, 3126S. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.W.; Li, X.; Zhang, Y.Y.; Huang, H.Y.; Liu, Y.; Sun, X.; Tang, Q.Q. Characterization of Adipocyte Differentiation from Human Mesenchymal Stem Cells in Bone Marrow. BMC Dev. Biol. 2010, 10, 47. [Google Scholar] [CrossRef]
- Sauer, S. Ligands for the Nuclear Peroxisome Proliferator-Activated Receptor Gamma. Trends Pharmacol. Sci. 2015, 36, 688–704. [Google Scholar] [CrossRef]
- Kudoh, A.; Satoh, H.; Hirai, H.; Watanabe, T. Pioglitazone Upregulates Adiponectin Receptor 2 in 3T3-L1 Adipocytes. Life Sci. 2011, 88, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Bou, M.; Todorčević, M.; Rodríguez, J.; Capilla, E.; Gutiérrez, J.; Navarro, I. Interplay of Adiponectin, TNFα and Insulin on Gene Expression, Glucose Uptake and PPARγ, AKT and TOR Pathways in Rainbow Trout Cultured Adipocytes. Gen. Comp. Endocrinol. 2014, 205, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Choi, H.Y.; Dayem, A.A.; Kim, K.; Yang, G.; Won, J.; Do, S.H.; Kim, J.; Jeong, K.; Cho, S. Regulation of Adipogenesis through Differential Modulation of ROS and Kinase Signaling Pathways by 3,4′-Dihydroxyflavone Treatment. J. Cell. Biochem. 2017, 118, 1065–1077. [Google Scholar] [CrossRef]
- Kim, H.; Sakamoto, K. (−)-Epigallocatechin Gallate Suppresses Adipocyte Differentiation through the MEK/ERK and PI3K/Akt Pathways. Cell Biol. Int. 2012, 36, 147–153. [Google Scholar] [CrossRef]
- Wijesekara, N.; Krishnamurthy, M.; Bhattacharjee, A.; Suhail, A.; Sweeney, G.; Wheeler, M.B. Adiponectin-Induced ERK and Akt Phosphorylation Protects Against Pancreatic Beta Cell Apoptosis and Increases Insulin Gene Expression and Secretion. J. Biol. Chem. 2010, 285, 33623–33631. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, F.M.C.; dos Santos, J.C.; Campos, J.L.O.; Mafud, A.C.; Polikarpov, I.; Figueira, A.C.M.; Nascimento, A.S. Structure-Based Identification of Novel PPAR Gamma Ligands. Bioorg. Med. Chem. Lett. 2013, 23, 5795–5802. [Google Scholar] [CrossRef]
- Zoete, V.; Grosdidier, A.; Michielin, O. Peroxisome Proliferator-Activated Receptor Structures: Ligand Specificity, Molecular Switch and Interactions with Regulators. Biochim. Biophys. Acta. 2007, 1771, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Festuccia, W.T.; Blanchard, P.; Deshaies, Y. Control of Brown Adipose Tissue Glucose and Lipid Metabolism by PPARγ. Front. Endocrinol. 2011, 2, 84. [Google Scholar] [CrossRef]
- Choi, J.H.; Banks, A.S.; Estall, J.L.; Kajimura, S.; Bostrom, P.; Laznik, D.; Ruas, J.L.; Chalmers, M.J.; Kamenecka, T.M.; Bluher, M. Obesity-Linked Phosphorylation of PPARγ by Cdk5 is a Direct Target of the Anti-Diabetic PPARγ Ligands. Nature 2010, 466, 451–456. [Google Scholar] [CrossRef]
- Siersbaek, R.; Nielsen, R.; Mandrup, S. PPARγ in adipocyte differentiation and metabolism - Novel insights from genome-wide studies. FEBS Lett. 2010, 584, 3242–3249. [Google Scholar] [CrossRef]
- Hansson, B.; Rippe, C.; Kotowska, D.; Wasserstrom, S.; Säll, J.; Göransson, O.; Swärd, K.; Stenkula, K.G. Rosiglitazone Drives Cavin-2/SDPR Expression in Adipocytes in a CEBPα-Dependent Manner. PLoS ONE 2017, 12, e0173412. [Google Scholar] [CrossRef] [PubMed]
- Madsen, M.S.; Siersbaek, R.; Boergesen, M.; Nielsen, R.; Mandrup, S. Peroxisome Proliferator-Activated Receptor and C/EBP Synergistically Activate Key Metabolic Adipocyte Genes by Assisted Loading. Mol. Cell. Biol. 2014, 34, 939–954. [Google Scholar] [CrossRef]
- Ambele, M.A.; Dessels, C.; Durandt, C.; Pepper, M.S. Genome-Wide Analysis of Gene Expression during Adipogenesis in Human Adipose-Derived Stromal Cells Reveals Novel Patterns of Gene Expression during Adipocyte Differentiation. Stem Cell Res. 2016, 16, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Loft, A.; Forss, I.; Siersbæk, M.S.; Schmidt, S.F.; Larsen, A.B.; Madsen, J.G.S.; Pisani, D.F.; Nielsen, R.; Aagaard, M.M.; Mathison, A. Browning of Human Adipocytes Requires KLF11 and Reprogramming of PPARγ Superenhancers. Genes Dev. 2015, 29, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Rajakumari, S.; Wu, J.; Ishibashi, J.; Lim, H.; Giang, A.; Won, K.; Reed, R.; Seale, P. EBF2 Determines and Maintains Brown Adipocyte Identity. Cell Metab. 2013, 17, 562–574. [Google Scholar] [CrossRef]
- Shapira, S.N.; Lim, H.; Rajakumari, S.; Sakers, A.P.; Ishibashi, J.; Harms, M.J.; Won, K.; Seale, P. EBF2 Transcriptionally Regulates Brown Adipogenesis Via the Histone Reader DPF3 and the BAF Chromatin Remodeling Complex. Genes Dev. 2017, 31, 660–673. [Google Scholar] [CrossRef]
- Wang, W.; Kissig, M.; Rajakumari, S.; Huang, L.; Lim, H.W.; Won, K.J.; Seale, P. Ebf2 is a Selective Marker of Brown and Beige Adipogenic Precursor Cells. Proc. Natl. Acad. Sci. USA 2014, 111, 14466–14471. [Google Scholar] [CrossRef]
- Sidossis, L.; Porter, C.; Saraf, M.; Børsheim, E.; Radhakrishnan, R.; Chao, T.; Ali, A.; Chondronikola, M.; Mlcak, R.; Finnerty, C. Browning of Subcutaneous White Adipose Tissue in Humans After Severe Adrenergic Stress. Cell Metab. 2015, 22, 219–227. [Google Scholar] [CrossRef]
- Chen, Y.C.; Yu, Y.H. The Potential of Brown Adipogenesis and Browning in Porcine Bone Marrow-Derived Mesenchymal Stem Cells 1. J. Animal Sci. 2018, 96, 3635–3644. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Jiang, T.; Fan, J.; Zheng, X.; Shi, X.; Yu, T.; Chu, G.; Yang, G. An Additive Effect of Promoting Thermogenic Gene Expression in Mice Adipose-Derived Stromal Vascular Cells by Combination of Rosiglitazone and CL316,243. Int. J. of Mol. Sci. 2017, 18, 1002. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, J.; Wang, Y.; Cannon, B. Cell Proliferation and Apoptosis Inhibition: Essential Processes for Recruitment of the Full Thermogenic Capacity of Brown Adipose Tissue. Biochim. Biophys. Acta. 2019, 1864, 51–58. [Google Scholar] [CrossRef]
- Sanchez-Gurmaches, J.; Tang, Y.; Jespersen, N.Z.; Wallace, M.; Martinez Calejman, C.; Gujja, S.; Li, H.; Edwards, Y.J.K.; Wolfrum, C.; Metallo, C.M. Brown Fat AKT2 is a Cold-Induced Kinase that Stimulates ChREBP-Mediated De Novo Lipogenesis to Optimize Fuel Storage and Thermogenesis. Cell Metab. 2018, 27, 209.e6. [Google Scholar] [CrossRef] [PubMed]
- Marquez, M.P.; Alencastro, F.; Madrigal, A.; Jimenez, J.L.; Blanco, G.; Gureghian, A.; Keagy, L.; Lee, C.; Liu, R.; Tan, L. The Role of Cellular Proliferation in Adipogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2017, 26, 1578–1595. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, Y.; Chen, H.; Chuang, W.; Lin, A.; Tsai, C.; Huang, C.; Lii, C. Andrographolide Inhibits Adipogenesis of 3T3-L1 Cells by Suppressing C/EBPβ Expression and Activation. Toxicol. Appl. Pharmacol. 2016, 307, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Song, W.; Zushin, P.H.; Liu, B.; Jedrychowski, M.P.; Mina, A.I.; Deng, Z.; Cabarkapa, D.; Hall, J.A.; Palmer, C.J. Phosphorylation of Beta-3 Adrenergic Receptor at Serine 247 by ERK MAP Kinase Drives Lipolysis in Obese Adipocytes. Mol. Metab. 2018, 12, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.F.; Bargut, T.C.L.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Thermogenesis, Fatty Acid Synthesis with Oxidation, and Inflammation in the Brown Adipose Tissue of Ob/Ob (−/−) Mice. Ann. Anat. 2017, 210, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Li, H.; Jeong, J.H.; Noh, M.; Ryu, J. Kazinol B from Broussonetia Kazinoki Improves Insulin Sensitivity Via Akt and AMPK Activation in 3T3-L1 Adipocytes. Fitoterapia 2016, 112, 90–96. [Google Scholar] [CrossRef]
- Choi, K.H.; Lee, H.A.; Park, M.H.; Han, J.S. Cyanidin-3-Rutinoside Increases Glucose Uptake by Activating the PI3K/Akt Pathway in 3T3-L1 Adipocytes. Environ. Toxicol. Pharmacol. 2017, 54, 1–6. [Google Scholar] [CrossRef]
- Soundharrajan, I.; Kim, D.H.; Srisesharam, S.; Kuppusamy, P.; Choi, K.C. Limonene Enhances Differentiation and 2-Deoxy-D-Glucose Uptake in 3T3-L1 Preadipocytes by Activating the Akt Signaling Pathway. Evid. Based Complement Alternat Med. 2018, 2018, 4573254. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Kim, S.; Kim, C.D.; Ha, J.M.; Yun, S.J.; Chung, S.W.; Hong, K.W.; Bae, S.S. Transcriptional Activation of Peroxisome Proliferator-Activated Receptor-Γ Requires Activation of both Protein Kinase A and Akt during Adipocyte Differentiation. Biochem. Biophys. Res. Commun. 2010, 399, 55–59. [Google Scholar] [CrossRef]
- Chernogubova, E.; Cannon, B.; Bengtsson, T. Norepinephrine Increases Glucose Transport in Brown Adipocytes Via Β3-Adrenoceptors through a cAMP, PKA, and PI3-Kinase-Dependent Pathway Stimulating Conventional and Novel PKCs. Endocrinology 2004, 145, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Standaert, M.L. Cbl, IRS-1, and IRS-2 Mediate Effects of Rosiglitazone on PI3K, PKC-, and Glucose Transport in 3T3/L1 Adipocytes. Endocrinology 2002, 143, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.; Teruel, T.; Lorenzo, M. Rosiglitazone Produces Insulin Sensitisation by Increasing Expression of the Insulin Receptor and its Tyrosine Kinase Activity in Brown Adipocytes. Diabetologia 2003, 46, 1618–1628. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.M.; Sato, M.; Dallner, O.S.; Sandström, A.L.; Pisani, D.F.; Chambard, J.C.; Amri, E.Z.; Hutchinson, D.S.; Bengtsson, T. Glucose Uptake in Brown Fat Cells is Dependent on mTOR Complex 2–promoted GLUT1 Translocation. J. Cell Biol. 2014, 207, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, K.; Voigt, V.; Wabitsch, M.; Brandsch, M. Protocol for Effective Differentiation of 3T3-L1 Cells to Adipocytes. Anal. Biochem. 2012, 425, 88–90. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Wu, Y.; Fried, S.K. A Modified Protocol to Maximize Differentiation of Human Preadipocytes and Improve Metabolic Phenotypes. Obesity (Silver Spring, Md.) 2012, 20, 2334–2340. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z. Growth Arrest Induction of 3T3-L1 Preadipocytes by Serum Starvation and their Differentiation by the Hormonal Adipogenic Cocktail. J. Cell Animal Biol. 2012, 6, 57–65. [Google Scholar]
- Abdallah, B.M.; Haack-Sørensen, M.; Burns, J.S.; Elsnab, B.; Jakob, F.; Hokland, P.; Kassem, M. Maintenance of Differentiation Potential of Human Bone Marrow Mesenchymal Stem Cells Immortalized by Human Telomerase Reverse Transcriptase Gene Despite of Extensive Proliferation. Biochem. Biophys. Res. Commun. 2005, 326, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Contador, D.; Ezquer, F.; Espinosa, M.; Arango-Rodriguez, M.; Puebla, C.; Sobrevia, L.; Conget, P. Dexamethasone and Rosiglitazone are Sufficient and Necessary for Producing Functional Adipocytes from Mesenchymal Stem Cells. Exp. Biol. Med. (Maywood) 2015, 240, 1235. [Google Scholar] [CrossRef] [PubMed]
- Skårn, M.; Noordhuis, P.; Wang, M.; Veuger, M.; Kresse, S.H.; Egeland, E.V.; Micci, F.; Namløs, H.M.; Håkelien, A.; Olafsrud, S.M. Generation and Characterization of an Immortalized Human Mesenchymal Stromal Cell Line. Stem Cells Dev. 2014, 23, 2377–2389. [Google Scholar] [CrossRef]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile Red: A Selective Fluorescent Stain for Intracellular Lipid Droplets. J. Cell Biol. 1985, 100, 965–973. [Google Scholar] [CrossRef]
- Aguena, M.; Fanganiello, R.D.; Tissiani, L.A.; Ishiy, F.A.; Atique, R.; Alonso, N.; Passos-Bueno, M.R. Optimization of Parameters for a More Efficient use of Adipose-Derived Stem Cells in Regenerative Medicine Therapies. Stem Cells Int. 2012, 2012, 303610. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| PPAR-γ | 5′-TTCTCCTATTGACCCAGAAAGC-3′ | 5′-CTCCACTTTGATTGCACTTTGG-3′ |
| C/EBP-α | 5′-TCGGTGGACAAGAACAG-3′ | 5′-GCAGGCGGTCATT-3′ |
| FASN | 5′AAGGACCTGTCTAGGTTTGATGC-3′ | 5′- TGGCTTCATAGGTGACTTCCA -3′ |
| EBF2 | 5′-TAGGAAGAGGACCAACTCTGAAA-3′ | 5′-CGACATTAGCGTCCACCACTC-3′ |
| UCP-1 | 5′-AGGTCCAAGGTGAATGCCC-3′ | 5′-TTACCACAGCGGTGATTGTTC-3′ |
| GAPDH | 5′-AGGGCTGCTTTTAACTCTGGT-3′ | 5′- CCCCACTTGATTTTGGAGGGA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayyad, A.M.; Khan, A.A.; Abdallah, S.H.; Alomran, S.S.; Bajou, K.; Khattak, M.N.K. Rosiglitazone Enhances Browning Adipocytes in Association with MAPK and PI3-K Pathways During the Differentiation of Telomerase-Transformed Mesenchymal Stromal Cells into Adipocytes. Int. J. Mol. Sci. 2019, 20, 1618. https://doi.org/10.3390/ijms20071618
Fayyad AM, Khan AA, Abdallah SH, Alomran SS, Bajou K, Khattak MNK. Rosiglitazone Enhances Browning Adipocytes in Association with MAPK and PI3-K Pathways During the Differentiation of Telomerase-Transformed Mesenchymal Stromal Cells into Adipocytes. International Journal of Molecular Sciences. 2019; 20(7):1618. https://doi.org/10.3390/ijms20071618
Chicago/Turabian StyleFayyad, Abeer Maher, Amir Ali Khan, Sallam Hasan Abdallah, Sara Sultan Alomran, Khalid Bajou, and Muhammad Nasir Khan Khattak. 2019. "Rosiglitazone Enhances Browning Adipocytes in Association with MAPK and PI3-K Pathways During the Differentiation of Telomerase-Transformed Mesenchymal Stromal Cells into Adipocytes" International Journal of Molecular Sciences 20, no. 7: 1618. https://doi.org/10.3390/ijms20071618
APA StyleFayyad, A. M., Khan, A. A., Abdallah, S. H., Alomran, S. S., Bajou, K., & Khattak, M. N. K. (2019). Rosiglitazone Enhances Browning Adipocytes in Association with MAPK and PI3-K Pathways During the Differentiation of Telomerase-Transformed Mesenchymal Stromal Cells into Adipocytes. International Journal of Molecular Sciences, 20(7), 1618. https://doi.org/10.3390/ijms20071618