Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Structure and Mechanism of Adiponectin Action
2.1. Structure of Adiponectin Gene and Proteins
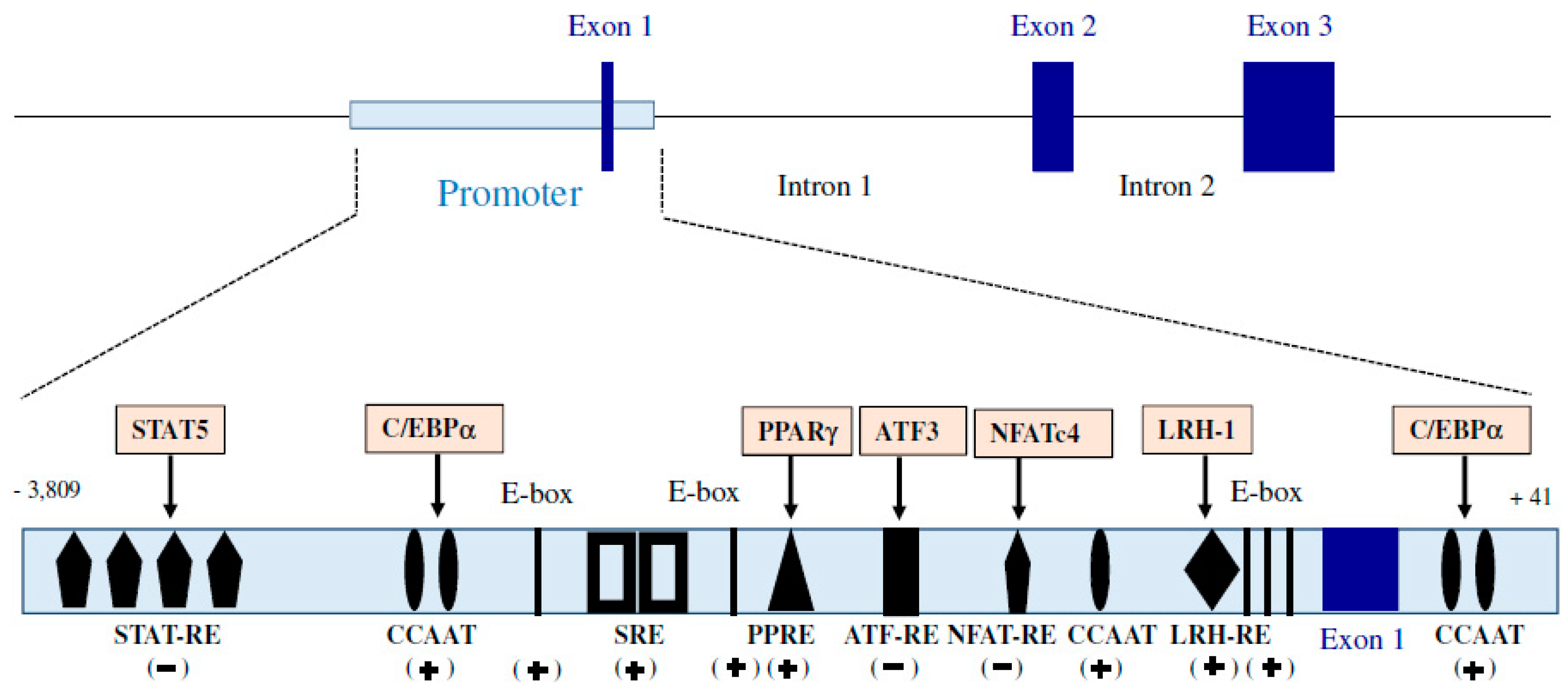
2.1.1. Adiponectin Gene
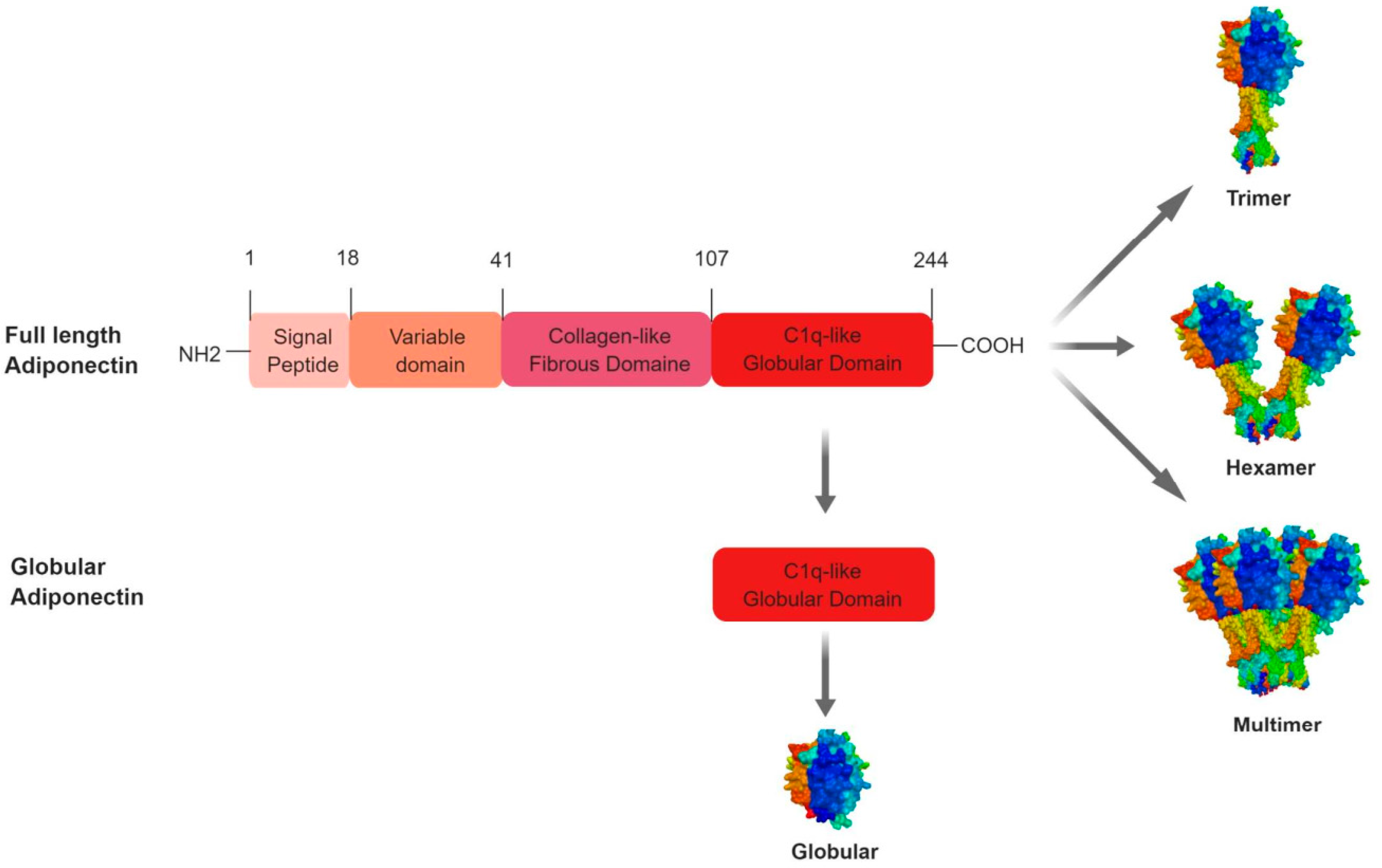
2.1.2. Adiponectin Protein
2.1.3. Regulation of Adiponectin Expression
2.2. Adiponectin Receptors and Adiponectin Signalling Pathways
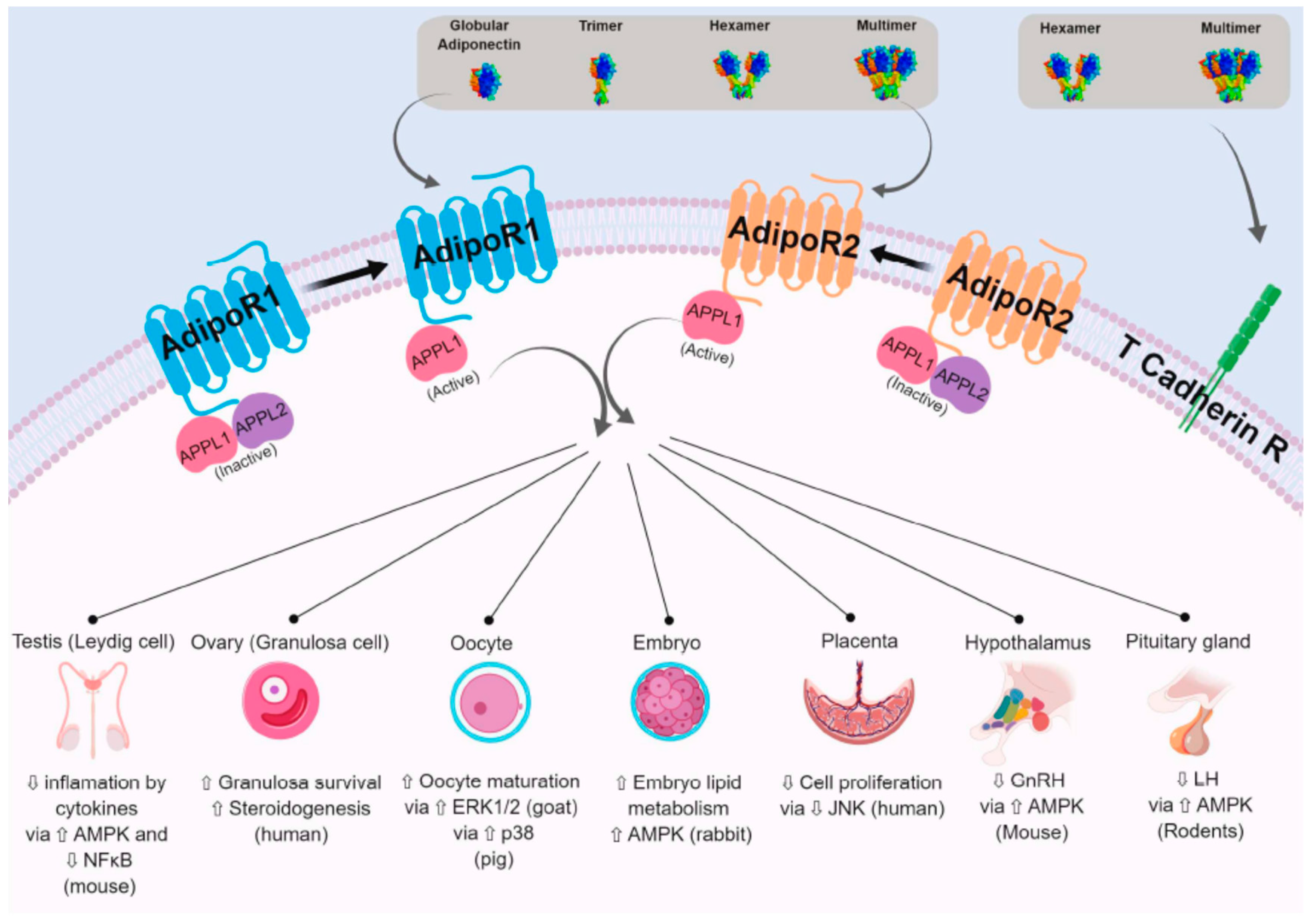
2.2.1. AdipoR1 and AdipoR2
2.2.2. The Other Adiponectin Receptors
2.2.3. APPL1 and APPL2
2.2.4. Signalling Pathways Regulated by Adiponectin
3. Expression, Regulation and Effect of Adiponectin and Adiponectin Receptors in the Hypothalamic–Pituitary Axis
3.1. Adiponectin and Hypothalamus: A Role in the Fertility Regulation?
3.2. Adiponectin and Pituitary: A Role in the Fertility Regulation?
4. Expression, Regulation and Effect of Adiponectin in Gonads
4.1. Expression, Regulation and Effect of Adiponectin System in Ovary
4.1.1. Plasma and Follicular Fluids Profiles
4.1.2. Expression in Ovarian Cells
4.1.3. Regulation by Physiologic Status
4.1.4. Regulation by Hormones
4.1.5. Effect on Steroidogenesis
4.2. Expression, Regulation and Effect of Adiponectin System in Testis
4.2.1. Blood Plasma and Seminal Fluid Profiles
4.2.2. Expression in Testicular Cells
4.2.3. Regulation by Physiologic Status
4.2.4. Regulation by Hormones
4.2.5. Effect on Steroidogenesis, Lactate Production and Cytokine-Mediated Cytotoxicity
5. Expression, Regulation and Effect of Adiponectin System in Gametogenesis (Oocyte and Spermatozoa)
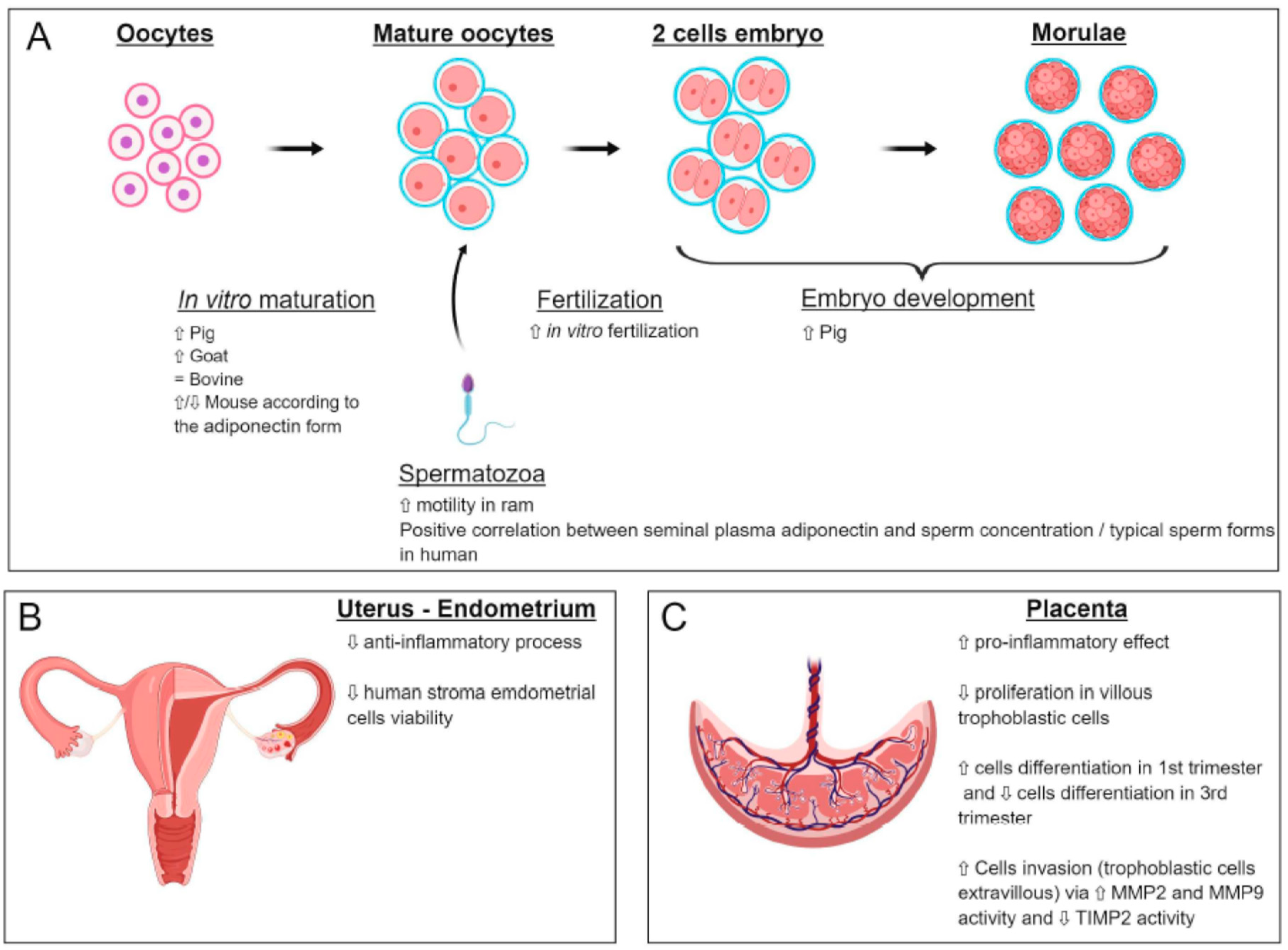
5.1. Oocyte
5.2. Spermatozoa
5.2.1. Localisation of Adiponectin and Its Receptor
5.2.2. Role of Adiponectin on Sperm Motility and Capacitation
6. Expression, Regulation and Effect of Adiponectin System in Embryo Development and Implantation: The Evolution of the Adiponectin System during Pregnancy
6.1. Adiponectin System during Embryo Development
6.2. Evolution of Serum Adiponectin during Pregnancy
7. Expression, Regulation and Effect of Adiponectin System in Endometrium, Placenta and Relation between the Foetus and Mother
7.1. Expression and Effects of Adiponectin on Uterine Functions
7.2. Expression and Effects of Adiponectin on the Placenta
7.3. Relation between Foetus and Mother
8. Foetus Growth
9. Adiponectin and Reproductive Diseases:
9.1. Ovarian Pathologies
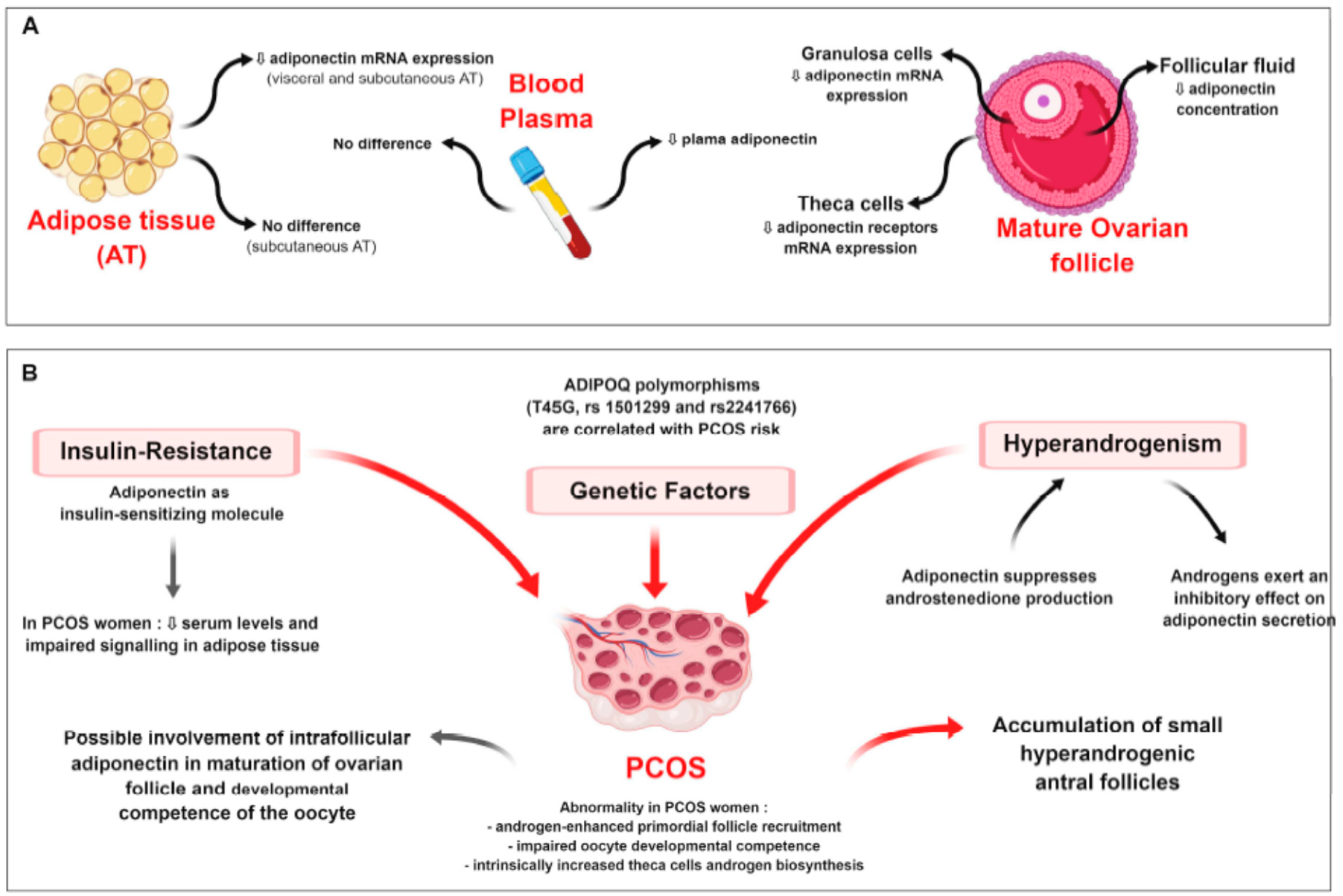
9.1.1. Polycystic Ovary Syndrome (PCOS)
9.1.2. Ovarian Cancers
9.2. Uterine/Endometrial Diseases
9.2.1. Endometriosis and Endometrial Cancer
9.2.2. Endometrial Cancer
9.3. Gestational Pathologies
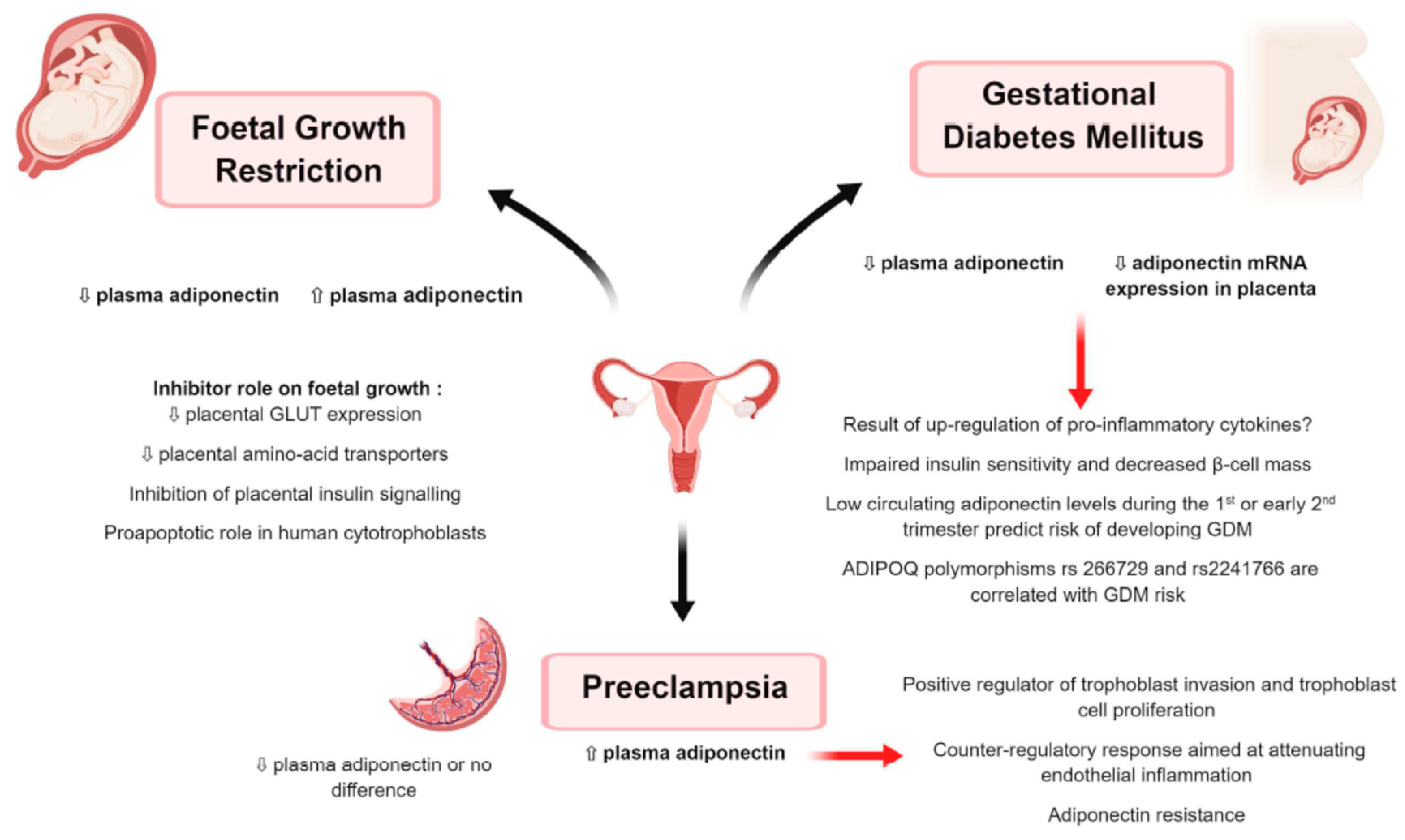
9.3.1. Gestational Diabetes Mellitus (Figure 6)
9.3.2. Preeclampsia (Figure 6)
9.3.3. Foetal Growth Restriction (Figure 6)
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| STAT | Signal Transducers and Activators of Transcription |
| CCAAT | CCAAT box is a distinct pattern of nucleotides with GGCCAATCT consensus sequence |
| SRE | Serum Response Element |
| PPRE | Peroxisome Proliferator Response Element |
| AP-1 | Activator Protein 1 |
| LRH-RE | Liver Receptor Homolog 1 Response Element |
| LMW | Low Molecular Weight |
| MMW | Medium Molecular Weight |
| HMW | High Molecular Weight |
| PCOS | Polycystic Ovary Syndrome |
| CSF | CerebroSpinal Fluid |
| HPG | Hypothalamic–Pituitary–Gonadal axis |
| AMH | Anti-Mullerian Hormone |
| MMP | Matrix MetalloProteinases |
| TIMP | Tissue Inhibitors of MetalloProteinases |
| PE | PreEclempsia |
| IGFBP-1 | Insulin-like Growth Factor Binding Protein 1 |
| hCG | human Chorionic Gonadotropin |
| FF | Follicular Fluid |
| TNF | Tumour Necrosis Factor |
| IFN | InterFeroN |
| PKA | Protein Kinase A |
| SF | Seminal Fluid |
| IVM | In Vitro Maturation |
| IGF-1 | Insulin like Growth Factor 1 |
| IR | Insulin Resistance |
| IS | Insulin Sensitivity |
| FFA | Free Fatty Acid |
| BMI | Body Mass Index |
| StAR | Steroid Acute Regulatory protein |
| T2DM | Type 2 Diabetes Mellitus |
| DHEA | DeHydroEpiAndrosterone |
| PTEN | Phosphatase and TENsin homolog |
| LKB1 | Liver Kinase B1 |
| NFkB | Transcription factor Nuclear Factor-kappa B |
| IL8/1 | InterLeukin 8/1 |
| ERK1/2 | Extracellular signal-Regulated Kinases 1 & 2 |
| MAPK | Mitogen-Activated Protein Kinases |
| PPAR | Peroxisome Proliferator-Activated Receptor |
| AMPK | AMP-activated Protein Kinase |
| APPL1/2 | Adaptor Protein, Phosphotyrosine interacting with PH domain and Leucine zipper 1/2 |
| BMAL1 | Brain and Muscle Arnt-Like protein-1 |
| CLOCK | Circadian Locomoter Output Cycles protein Kaput |
| Siglec10 | Sialic acid binding ig like lectin 10 |
| SNAT | Sodium-coupled Neutral Amino acid Transporters |
| mTORC1 | Mammalian Target Of Rapamycin Complex 1 or mechanistic target of rapamycin complex 1 |
| GLUT | GLUcose Transporter |
| MCT2/4 | MonoCarboxylate Transporter (lactate transporter) |
References
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K.; et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem. Biophys. Res. Commun. 1999, 257, 79–83. [Google Scholar] [CrossRef]
- Takahashi, M.; Arita, Y.; Yamagata, K.; Matsukawa, Y.; Okutomi, K.; Horie, M.; Shimomura, I.; Hotta, K.; Kuriyama, H.; Kihara, S.; et al. Genomic structure and mutations in adipose-specific gene, adiponectin. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, F. Transcriptional and post-translational regulation of adiponectin. Biochem. J. 2009, 425, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Funahashi, T.; Shimomura, I. Pathophysiological significance of adiponectin. Med. Mol. Morphol. 2007, 40, 55–67. [Google Scholar] [CrossRef]
- Waki, H.; Yamauchi, T.; Kamon, J.; Kita, S.; Ito, Y.; Hada, Y.; Uchida, S.; Tsuchida, A.; Takekawa, S.; Kadowaki, T. Generation of globular fragment of adiponectin by leukocyte elastase secreted by monocytic cell line THP-1. Endocrinology 2005, 146, 790–796. [Google Scholar] [CrossRef]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef]
- Hada, Y.; Yamauchi, T.; Waki, H.; Tsuchida, A.; Hara, K.; Yago, H.; Miyazaki, O.; Ebinuma, H.; Kadowaki, T. Selective purification and characterization of adiponectin multimer species from human plasma. Biochem. Biophys. Res. Commun. 2007, 356, 487–493. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, K.S.; Yau, M.H.; Xu, A. Post-translational modifications of adiponectin: Mechanisms and functional implications. Biochem. J. 2008, 409, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.; Scherer, P.E. The crystal structure of a complement-1q family protein suggests an evolutionary link to tumor necrosis factor. Curr. Biol. 1998, 8, 335–338. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, K.S.; Chan, L.; Chan, K.W.; Lam, J.B.; Lam, M.C.; Hoo, R.C.; Mak, W.W.; Cooper, G.J.; Xu, A. Post-translational modifications of the four conserved lysine residues within the collagenous domain of adiponectin are required for the formation of its high molecular weight oligomeric complex. J. Biol. Chem. 2006, 281, 16391–16400. [Google Scholar] [CrossRef] [PubMed]
- Waki, H.; Yamauchi, T.; Kamon, J.; Ito, Y.; Uchida, S.; Kita, S.; Hara, K.; Hada, Y.; Vasseur, F.; Froguel, P.; et al. Impaired multimerization of human adiponectin mutants associated with diabetes. Molecular structure and multimer formation of adiponectin. J. Biol. Chem. 2003, 278, 40352–40363. [Google Scholar] [CrossRef]
- Wedellova, Z.; Kovacova, Z.; Tencerova, M.; Vedral, T.; Rossmeislova, L.; Siklova-Vitkova, M.; Stich, V.; Polak, J. The Impact of Full-Length, Trimeric and Globular Adiponectin on Lipolysis in Subcutaneous and Visceral Adipocytes of Obese and Non-Obese Women. PLoS ONE 2013, 8, e66783. [Google Scholar] [CrossRef]
- Nakano, Y.; Tajima, S.; Yoshimi, A.; Akiyama, H.; Tsushima, M.; Tanioka, T.; Negoro, T.; Tomita, M.; Tobe, T. A novel enzyme-linked immunosorbent assay specific for high-molecular-weight adiponectin. J. Lipid Res. 2006, 47, 1572–1582. [Google Scholar] [CrossRef]
- Tishinsky, J.M.; Robinson, L.E.; Dyck, D.J. Insulin-sensitizing properties of adiponectin. Biochimie 2012, 94, 2131–2136. [Google Scholar] [CrossRef]
- Swarbrick, M.M.; Havel, P.J. Physiological, pharmacological, and nutritional regulation of circulating adiponectin concentrations in humans. Metab. Syndr. Relat. Disord. 2008, 6, 87–102. [Google Scholar] [CrossRef]
- Chabrolle, C.; Tosca, L.; Dupont, J. Regulation of adiponectin and its receptors in rat ovary by human chorionic gonadotrophin treatment and potential involvement of adiponectin in granulosa cell steroidogenesis. Reproduction 2007, 133, 719–731. [Google Scholar] [CrossRef]
- Diot, M.; Reverchon, M.; Rame, C.; Froment, P.; Brillard, J.P.; Briere, S.; Leveque, G.; Guillaume, D.; Dupont, J. Expression of adiponectin, chemerin and visfatin in plasma and different tissues during a laying season in turkeys. Reprod. Biol. Endocrinol. 2015, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Maleszka, A.; Smolinska, N.; Nitkiewicz, A.; Kiezun, M.; Chojnowska, K.; Dobrzyn, K.; Szwaczek, H.; Kaminski, T. Adiponectin Expression in the Porcine Ovary during the Oestrous Cycle and Its Effect on Ovarian Steroidogenesis. Int. J. Endocrinol. 2014, 2014, 957076. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Shimomura, I.; Kishida, K.; Maeda, N.; Kuriyama, H.; Nagaretani, H.; Matsuda, M.; Kondo, H.; Furuyama, N.; Kihara, S.; et al. Androgens decrease plasma adiponectin, an insulin-sensitising adipocyte-derived protein. Diabetes 2002, 51, 2734–2741. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin receptors: A review of their structure, function and how they work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, G.L., 3rd; Hadley, J.A.; Krzysik-Walker, S.M.; Prabhu, K.S.; Vasilatos-Younken, R.; Ramachandran, R. Unique profile of chicken adiponectin, a predominantly heavy molecular weight multimer, and relationship to visceral adiposity. Endocrinology 2009, 150, 3092–3100. [Google Scholar] [CrossRef]
- Combs, T.P.; Marliss, E.B. Adiponectin signaling in the liver. Rev. Endocr. Metab. Disord. 2014, 15, 137–147. [Google Scholar] [CrossRef]
- Singh, S.P.; Haussler, S.; Gross, J.J.; Schwarz, F.J.; Bruckmaier, R.M.; Sauerwein, H. Short communication: Circulating and milk adiponectin change differently during energy deficiency at different stages of lactation in dairy cows. J. Dairy Sci. 2014, 97, 1535–1542. [Google Scholar] [CrossRef]
- Giesy, S.L.; Yoon, B.; Currie, W.B.; Kim, J.W.; Boisclair, Y.R. Adiponectin deficit during the precarious glucose economy of early lactation in dairy cows. Endocrinology 2012, 153, 5834–5844. [Google Scholar] [CrossRef]
- Ohtani, Y.; Takahashi, T.; Sato, K.; Ardiyanti, A.; Song, S.H.; Sato, R.; Onda, K.; Wada, Y.; Obara, Y.; Suzuki, K.; et al. Changes in circulating adiponectin and metabolic hormone concentrations during periparturient and lactation periods in Holstein dairy cows. Anim. Sci. J. 2012, 83, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Mielenz, M.; Mielenz, B.; Singh, S.P.; Kopp, C.; Heinz, J.; Haussler, S.; Sauerwein, H. Development, validation, and pilot application of a semiquantitative Western blot analysis and an ELISA for bovine adiponectin. Domest. Anim. Endocrinol. 2013, 44, 121–130. [Google Scholar] [CrossRef]
- Mellouk, N.; Rame, C.; Touze, J.L.; Briant, E.; Ma, L.; Guillaume, D.; Lomet, D.; Caraty, A.; Ntallaris, T.; Humblot, P.; et al. Involvement of plasma adipokines in metabolic and reproductive parameters in Holstein dairy cows fed with diets with differing energy levels. J. Dairy Sci. 2017, 100, 8518–8533. [Google Scholar] [CrossRef]
- Heinz, J.F.; Singh, S.P.; Janowitz, U.; Hoelker, M.; Tesfaye, D.; Schellander, K.; Sauerwein, H. Characterization of adiponectin concentrations and molecular weight forms in serum, seminal plasma, and ovarian follicular fluid from cattle. Theriogenology 2015, 83, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Yano, W.; Kubota, T.; Yamauchi, T.; Itoh, S.; Kumagai, H.; Kozono, H.; Takamoto, I.; Okamoto, S.; Shiuchi, T.; et al. Adiponectin stimulates AMP-activated protein kinase in the hypothalamus and increases food intake. Cell Metab. 2007, 6, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Kuang, M.; Nie, H.; Bai, W.; Sun, L.; Wang, F.; Mao, D.; Wang, Z. Impact of Food Restriction on the Expression of the Adiponectin System and Genes in the Hypothalamic-Pituitary-Ovarian Axis of Pre-Pubertal Ewes. Reprod. Domest. Anim. 2016, 51, 657–664. [Google Scholar] [CrossRef]
- Cnop, M.; Havel, P.J.; Utzschneider, K.M.; Carr, D.B.; Sinha, M.K.; Boyko, E.J.; Retzlaff, B.M.; Knopp, R.H.; Brunzell, J.D.; Kahn, S.E. Relationship of adiponectin to body fat distribution, insulin sensitivity and plasma lipoproteins: Evidence for independent roles of age and sex. Diabetologia 2003, 46, 459–469. [Google Scholar] [CrossRef]
- Combs, T.P.; Berg, A.H.; Rajala, M.W.; Klebanov, S.; Iyengar, P.; Jimenez-Chillaron, J.C.; Patti, M.E.; Klein, S.L.; Weinstein, R.S.; Scherer, P.E. Sexual differentiation, pregnancy, calorie restriction, and aging affect the adipocyte-specific secretory protein adiponectin. Diabetes 2003, 52, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Walsh, K. Obesity, adiponectin and vascular inflammatory disease. Curr. Opin. Lipidol. 2003, 14, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, M.; Matsuda, M.; Maeda, N.; Funahashi, T.; Matsuzawa, Y.; Makishima, M.; Shimomura, I. Induction of adiponectin, a fat-derived antidiabetic and antiatherogenic factor, by nuclear receptors. Diabetes 2003, 52, 1655–1663. [Google Scholar] [CrossRef]
- Berg, A.H.; Combs, T.P.; Scherer, P.E. ACRP30/adiponectin: An adipokine regulating glucose and lipid metabolism. Trends Endocrinol. Metab. 2002, 13, 84–89. [Google Scholar] [CrossRef]
- Qiao, L.; Maclean, P.S.; Schaack, J.; Orlicky, D.J.; Darimont, C.; Pagliassotti, M.; Friedman, J.E.; Shao, J. C/EBPalpha regulates human adiponectin gene transcription through an intronic enhancer. Diabetes 2005, 54, 1744–1754. [Google Scholar] [CrossRef]
- Qiao, L.; Shao, J. SIRT1 regulates adiponectin gene expression through Foxo1-C/enhancer-binding protein alpha transcriptional complex. J. Biol. Chem. 2006, 281, 39915–39924. [Google Scholar] [CrossRef]
- Seo, J.B.; Moon, H.M.; Noh, M.J.; Lee, Y.S.; Jeong, H.W.; Yoo, E.J.; Kim, W.S.; Park, J.; Youn, B.S.; Kim, J.W.; et al. Adipocyte determination- and differentiation-dependent factor 1/sterol regulatory element-binding protein 1c regulates mouse adiponectin expression. J. Biol. Chem. 2004, 279, 22108–22117. [Google Scholar] [CrossRef]
- Kim, H.B.; Kong, M.; Kim, T.M.; Suh, Y.H.; Kim, W.H.; Lim, J.H.; Song, J.H.; Jung, M.H. NFATc4 and ATF3 negatively regulate adiponectin gene expression in 3T3-L1 adipocytes. Diabetes 2006, 55, 1342–1352. [Google Scholar] [CrossRef] [PubMed]
- Doran, A.C.; Meller, N.; Cutchins, A.; Deliri, H.; Slayton, R.P.; Oldham, S.N.; Kim, J.B.; Keller, S.R.; McNamara, C.A. The helix-loop-helix factors Id3 and E47 are novel regulators of adiponectin. Circ. Res. 2008, 103, 624–634. [Google Scholar] [CrossRef]
- White, U.A.; Maier, J.; Zhao, P.; Richard, A.J.; Stephens, J.M. The modulation of adiponectin by STAT5-activating hormones. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E129–E136. [Google Scholar] [CrossRef]
- Barnea, M.; Chapnik, N.; Genzer, Y.; Froy, O. The circadian clock machinery controls adiponectin expression. Mol. Cell. Endocrinol. 2015, 399, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, H.; Fujii, Y.; Okada-Iwabu, M.; Iwabu, M.; Nakamura, Y.; Hosaka, T.; Motoyama, K.; Ikeda, M.; Wakiyama, M.; Terada, T.; et al. Crystal structures of the human adiponectin receptors. Nature 2015, 520, 312–316. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K.; Tobe, K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Investig. 2006, 116, 1784–1792. [Google Scholar] [CrossRef]
- Hug, C.; Wang, J.; Ahmad, N.S.; Bogan, J.S.; Tsao, T.S.; Lodish, H.F. T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc. Natl. Acad. Sci. USA 2004, 101, 10308–10313. [Google Scholar] [CrossRef] [PubMed]
- Denzel, M.S.; Scimia, M.C.; Zumstein, P.M.; Walsh, K.; Ruiz-Lozano, P.; Ranscht, B. T-cadherin is critical for adiponectin-mediated cardioprotection in mice. J. Clin. Investig. 2010, 120, 4342–4352. [Google Scholar] [CrossRef]
- Ruan, H.; Dong, L.Q. Adiponectin signaling and function in insulin target tissues. J. Mol. Cell Biol. 2016, 8, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Kikani, C.K.; Riojas, R.A.; Langlais, P.; Wang, L.; Ramos, F.J.; Fang, Q.; Christ-Roberts, C.Y.; Hong, J.Y.; Kim, R.Y.; et al. APPL1 binds to adiponectin receptors and mediates adiponectin signalling and function. Nat. Cell Biol. 2006, 8, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xin, X.; Xiang, R.; Ramos, F.J.; Liu, M.; Lee, H.J.; Chen, H.; Mao, X.; Kikani, C.K.; Liu, F.; et al. Yin-Yang regulation of adiponectin signaling by APPL isoforms in muscle cells. J. Biol. Chem. 2009, 284, 31608–31615. [Google Scholar] [CrossRef] [PubMed]
- Kos, K.; Harte, A.L.; da Silva, N.F.; Tonchev, A.; Chaldakov, G.; James, S.; Snead, D.R.; Hoggart, B.; O’Hare, J.P.; McTernan, P.G.; et al. Adiponectin and resistin in human cerebrospinal fluid and expression of adiponectin receptors in the human hypothalamus. J. Clin. Endocrinol. Metab. 2007, 92, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Psilopanagioti, A.; Papadaki, H.; Kranioti, E.F.; Alexandrides, T.K.; Varakis, J.N. Expression of adiponectin and adiponectin receptors in human pituitary gland and brain. Neuroendocrinology 2009, 89, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Shi, H.; Jin, Y.; Li, X.; Pan, J.; Lai, Y.; Lin, Y.; Jin, Y.; Roy, G.; Zhao, A.; et al. Adiponectin Deficiency Leads to Female Subfertility and Ovarian Dysfunctions in Mice. Endocrinology 2016, 157, 4875–4887. [Google Scholar] [CrossRef] [PubMed]
- Guillod-Maximin, E.; Roy, A.F.; Vacher, C.M.; Aubourg, A.; Bailleux, V.; Lorsignol, A.; Penicaud, L.; Parquet, M.; Taouis, M. Adiponectin receptors are expressed in hypothalamus and colocalized with proopiomelanocortin and neuropeptide Y in rodent arcuate neurons. J. Endocrinol. 2009, 200, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, T.; Smolinska, N.; Maleszka, A.; Kiezun, M.; Dobrzyn, K.; Czerwinska, J.; Szeszko, K.; Nitkiewicz, A. Expression of adiponectin and its receptors in the porcine hypothalamus during the oestrous cycle. Reprod. Domest. Anim. 2014, 49, 378–386. [Google Scholar] [CrossRef]
- Kusminski, C.M.; McTernan, P.G.; Schraw, T.; Kos, K.; O’Hare, J.P.; Ahima, R.; Kumar, S.; Scherer, P.E. Adiponectin complexes in human cerebrospinal fluid: Distinct complex distribution from serum. Diabetologia 2007, 50, 634–642. [Google Scholar] [CrossRef]
- Neumeier, M.; Weigert, J.; Buettner, R.; Wanninger, J.; Schaffler, A.; Muller, A.M.; Killian, S.; Sauerbruch, S.; Schlachetzki, F.; Steinbrecher, A.; et al. Detection of adiponectin in cerebrospinal fluid in humans. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E965–E969. [Google Scholar] [CrossRef]
- Qi, Y.; Takahashi, N.; Hileman, S.M.; Patel, H.R.; Berg, A.H.; Pajvani, U.B.; Scherer, P.E.; Ahima, R.S. Adiponectin acts in the brain to decrease body weight. Nat. Med. 2004, 10, 524–529. [Google Scholar] [CrossRef]
- Cheng, X.B.; Wen, J.P.; Yang, J.; Yang, Y.; Ning, G.; Li, X.Y. GnRH secretion is inhibited by adiponectin through activation of AMP-activated protein kinase and extracellular signal-regulated kinase. Endocrine 2011, 39, 6–12. [Google Scholar] [CrossRef]
- Wen, J.P.; Liu, C.; Bi, W.K.; Hu, Y.T.; Chen, Q.; Huang, H.; Liang, J.X.; Li, L.T.; Lin, L.X.; Chen, G. Adiponectin inhibits KISS1 gene transcription through AMPK and specificity protein-1 in the hypothalamic GT1-7 neurons. J. Endocrinol. 2012, 214, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.P.; Lv, W.S.; Yang, J.; Nie, A.F.; Cheng, X.B.; Yang, Y.; Ge, Y.; Li, X.Y.; Ning, G. Globular adiponectin inhibits GnRH secretion from GT1-7 hypothalamic GnRH neurons by induction of hyperpolarization of membrane potential. Biochem. Biophys. Res. Commun. 2008, 371, 756–761. [Google Scholar] [CrossRef]
- Klenke, U.; Taylor-Burds, C.; Wray, S. Metabolic influences on reproduction: Adiponectin attenuates GnRH neuronal activity in female mice. Endocrinology 2014, 155, 1851–1863. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Pacheco, F.; Martinez-Fuentes, A.J.; Tovar, S.; Pinilla, L.; Tena-Sempere, M.; Dieguez, C.; Castano, J.P.; Malagon, M.M. Regulation of pituitary cell function by adiponectin. Endocrinology 2007, 148, 401–410. [Google Scholar] [CrossRef]
- Ramachandran, R.; Ocon-Grove, O.M.; Metzger, S.L. Molecular cloning and tissue expression of chicken AdipoR1 and AdipoR2 complementary deoxyribonucleic acids. Domest. Anim. Endocrinol. 2007, 33, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Kiezun, M.; Smolinska, N.; Maleszka, A.; Dobrzyn, K.; Szeszko, K.; Kaminski, T. Adiponectin expression in the porcine pituitary during the estrous cycle and its effect on LH and FSH secretion. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E1038–E1046. [Google Scholar] [CrossRef]
- Lu, M.; Tang, Q.; Olefsky, J.M.; Mellon, P.L.; Webster, N.J. Adiponectin activates adenosine monophosphate-activated protein kinase and decreases luteinizing hormone secretion in LbetaT2 gonadotropes. Mol. Endocrinol. 2008, 22, 760–771. [Google Scholar] [CrossRef]
- Sarmento-Cabral, A.; Peinado, J.R.; Halliday, L.C.; Malagon, M.M.; Castano, J.P.; Kineman, R.D.; Luque, R.M. Adipokines (Leptin, Adiponectin, Resistin) Differentially Regulate All Hormonal Cell Types in Primary Anterior Pituitary Cell Cultures from Two Primate Species. Sci. Rep. 2017, 7, 43537. [Google Scholar] [CrossRef]
- Rak, A.; Mellouk, N.; Froment, P.; Dupont, J. Adiponectin and resistin: Potential metabolic signals affecting hypothalamo-pituitary gonadal axis in females and males of different species. Reproduction 2017, 153, R215–R226. [Google Scholar] [CrossRef]
- Kawwass, J.F.; Summer, R.; Kallen, C.B. Direct effects of leptin and adiponectin on peripheral reproductive tissues: A critical review. Mol. Hum. Reprod. 2015, 21, 617–632. [Google Scholar] [CrossRef]
- Chabrolle, C.; Tosca, L.; Rame, C.; Lecomte, P.; Royere, D.; Dupont, J. Adiponectin increases insulin-like growth factor I-induced progesterone and estradiol secretion in human granulosa cells. Fertil. Steril. 2009, 92, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Bersinger, N.A.; Wunder, D.M. Adiponectin isoform distribution in serum and in follicular fluid of women undergoing treatment by ICSI. Acta Obstet. Gynecol. Scand. 2010, 89, 782–788. [Google Scholar] [CrossRef]
- Li, L.; Ferin, M.; Sauer, M.V.; Lobo, R.A. Ovarian adipocytokines are associated with early in vitro human embryo development independent of the action of ovarian insulin. J. Assist. Reprod. Genet. 2012, 29, 1397–1404. [Google Scholar] [CrossRef]
- Bersinger, N.A.; Birkhauser, M.H.; Wunder, D.M. Adiponectin as a marker of success in intracytoplasmic sperm injection/embryo transfer cycles. Gynecol. Endocrinol. 2006, 22, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Meng, B.; Fan, R.; Liu, M.; Gao, M.; Xing, Z.; Luan, X. Comparative proteomic analysis of ovaries from Huoyan geese between pre-laying and laying periods using an iTRAQ-based approach. Poult. Sci. 2018, 97, 2170–2182. [Google Scholar] [CrossRef]
- Mellouk, N.; Rame, C.; Marchand, M.; Staub, C.; Touze, J.L.; Venturi, E.; Mercerand, F.; Travel, A.; Chartrin, P.; Lecompte, F.; et al. Effect of different levels of feed restriction and fish oil fatty acid supplementation on fat deposition by using different techniques, plasma levels and mRNA expression of several adipokines in broiler breeder hens. PLoS ONE 2018, 13, e0191121. [Google Scholar] [CrossRef]
- Tabandeh, M.R.; Hosseini, A.; Saeb, M.; Kafi, M.; Saeb, S. Changes in the gene expression of adiponectin and adiponectin receptors (AdipoR1 and AdipoR2) in ovarian follicular cells of dairy cow at different stages of development. Theriogenology 2010, 73, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Wyskida, K.; Franik, G.; Wikarek, T.; Owczarek, A.; Delroba, A.; Chudek, J.; Sikora, J.; Olszanecka-Glinianowicz, M. The levels of adipokines in relation to hormonal changes during the menstrual cycle in young, normal-weight women. Endocr. Connect. 2017, 6, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Galvan, R.E.; Basurto, L.; Saucedo, R.; Campos, S.; Hernandez, M.; Zarate, A. Adiponectin concentrations during menstrual cycle. Ginecol. Obstet. Mexico 2007, 75, 435–438. [Google Scholar]
- Gutman, G.; Barak, V.; Maslovitz, S.; Amit, A.; Lessing, J.B.; Geva, E. Recombinant luteinizing hormone induces increased production of ovarian follicular adiponectin in vivo: Implications for enhanced insulin sensitivity. Fertil. Steril. 2009, 91, 1837–1841. [Google Scholar] [CrossRef]
- Ledoux, S.; Campos, D.B.; Lopes, F.L.; Dobias-Goff, M.; Palin, M.F.; Murphy, B.D. Adiponectin induces periovulatory changes in ovarian follicular cells. Endocrinology 2006, 147, 5178–5186. [Google Scholar] [CrossRef]
- Lagaly, D.V.; Aad, P.Y.; Grado-Ahuir, J.A.; Hulsey, L.B.; Spicer, L.J. Role of adiponectin in regulating ovarian theca and granulosa cell function. Mol. Cell. Endocrinol. 2008, 284, 38–45. [Google Scholar] [CrossRef]
- Pierre, P.; Froment, P.; Negre, D.; Rame, C.; Barateau, V.; Chabrolle, C.; Lecomte, P.; Dupont, J. Role of adiponectin receptors, AdipoR1 and AdipoR2, in the steroidogenesis of the human granulosa tumor cell line, KGN. Hum. Reprod. 2009, 24, 2890–2901. [Google Scholar] [CrossRef] [PubMed]
- Maillard, V.; Uzbekova, S.; Guignot, F.; Perreau, C.; Rame, C.; Coyral-Castel, S.; Dupont, J. Effect of adiponectin on bovine granulosa cell steroidogenesis, oocyte maturation and embryo development. Reprod. Biol. Endocrinol. 2010, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Comim, F.V.; Gutierrez, K.; Bridi, A.; Bochi, G.; Chemeris, R.; Rigo, M.L.; Dau, A.M.; Cezar, A.S.; Moresco, R.N.; Goncalves, P.B. Effects of Adiponectin Including Reduction of Androstenedione Secretion and Ovarian Oxidative Stress Parameters In Vivo. PLoS ONE 2016, 11, e0154453. [Google Scholar] [CrossRef] [PubMed]
- Sepilian, V.; Nagamani, M. Adiponectin levels in women with polycystic ovary syndrome and severe insulin resistance. J. Soc. Gynecol. Investig. 2005, 12, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Kratzsch, D.; Schaab, M.; Scholz, M.; Grunewald, S.; Thiery, J.; Paasch, U.; Kratzsch, J. Seminal plasma adipokine levels are correlated with functional characteristics of spermatozoa. Fertil. Steril. 2013, 99, 1256–1263e3. [Google Scholar] [CrossRef] [PubMed]
- Civitarese, A.E.; Jenkinson, C.P.; Richardson, D.; Bajaj, M.; Cusi, K.; Kashyap, S.; Berria, R.; Belfort, R.; DeFronzo, R.A.; Mandarino, L.J.; et al. Adiponectin receptors gene expression and insulin sensitivity in non-diabetic Mexican Americans with or without a family history of Type 2 diabetes. Diabetologia 2004, 47, 816–820. [Google Scholar] [CrossRef]
- Caminos, J.E.; Nogueiras, R.; Gaytan, F.; Pineda, R.; Gonzalez, C.R.; Barreiro, M.L.; Castano, J.P.; Malagon, M.M.; Pinilla, L.; Toppari, J.; et al. Novel expression and direct effects of adiponectin in the rat testis. Endocrinology 2008, 149, 3390–3402. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J. Implications of adiponectin in linking metabolism to testicular function. Endocrine 2014, 46, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Bjursell, M.; Ahnmark, A.; Bohlooly, Y.M.; William-Olsson, L.; Rhedin, M.; Peng, X.R.; Ploj, K.; Gerdin, A.K.; Arnerup, G.; Elmgren, A.; et al. Opposing effects of adiponectin receptors 1 and 2 on energy metabolism. Diabetes 2007, 56, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Choubey, M.; Ranjan, A.; Bora, P.S.; Baltazar, F.; Martin, L.J.; Krishna, A. Role of adiponectin as a modulator of testicular function during aging in mice. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Ocon-Grove, O.M.; Krzysik-Walker, S.M.; Maddineni, S.R.; Hendricks, G.L., 3rd; Ramachandran, R. Adiponectin and its receptors are expressed in the chicken testis: Influence of sexual maturation on testicular ADIPOR1 and ADIPOR2 mRNA abundance. Reproduction 2008, 136, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Pfaehler, A.; Nanjappa, M.K.; Coleman, E.S.; Mansour, M.; Wanders, D.; Plaisance, E.P.; Judd, R.L.; Akingbemi, B.T. Regulation of adiponectin secretion by soy isoflavones has implication for endocrine function of the testis. Toxicol. Lett. 2012, 209, 78–85. [Google Scholar] [CrossRef]
- Gui, Y.; Silha, J.V.; Murphy, L.J. Sexual dimorphism and regulation of resistin, adiponectin, and leptin expression in the mouse. Obes. Res. 2004, 12, 1481–1491. [Google Scholar] [CrossRef]
- Yarrow, J.F.; Beggs, L.A.; Conover, C.F.; McCoy, S.C.; Beck, D.T.; Borst, S.E. Influence of androgens on circulating adiponectin in male and female rodents. PLoS ONE 2012, 7, e47315. [Google Scholar] [CrossRef] [PubMed]
- Lanfranco, F.; Zitzmann, M.; Simoni, M.; Nieschlag, E. Serum adiponectin levels in hypogonadal males: Influence of testosterone replacement therapy. Clin. Endocrinol. (Oxf.) 2004, 60, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Kadooka, K.; Sato, M.; Matsumoto, T.; Kuhara, S.; Katakura, Y.; Fujimura, T. Pig testis extract augments adiponectin expression and secretion through the peroxisome proliferator-activated receptor signaling pathway in 3T3-L1 adipocytes. Cytotechnology 2018, 70, 983–992. [Google Scholar] [CrossRef]
- Choubey, M.; Ranjan, A.; Bora, P.S.; Baltazar, F.; Krishna, A. Direct actions of adiponectin on changes in reproductive, metabolic, and anti-oxidative enzymes status in the testis of adult mice. Gen. Comp. Endocrinol. 2018. [Google Scholar] [CrossRef]
- Ahn, S.W.; Gang, G.T.; Tadi, S.; Nedumaran, B.; Kim, Y.D.; Park, J.H.; Kweon, G.R.; Koo, S.H.; Lee, K.; Ahn, R.S.; et al. Phosphoenolpyruvate carboxykinase and glucose-6-phosphatase are required for steroidogenesis in testicular Leydig cells. J. Biol. Chem. 2012, 287, 41875–41887. [Google Scholar] [CrossRef]
- Carmina, E.; Chu, M.C.; Moran, C.; Tortoriello, D.; Vardhana, P.; Tena, G.; Preciado, R.; Lobo, R. Subcutaneous and omental fat expression of adiponectin and leptin in women with polycystic ovary syndrome. Fertil. Steril. 2008, 89, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.; Mazumder, S.; Bhattacharya, S.; Sinha, A.K. The sex specific effects of extraneous testosterone on ADP induced platelet aggregation in platelet-rich plasma from male and female subjects. Int. J. Lab. Hematol. 2014, 36, e74–e77. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Xu, B.; Fan, W.; Zhu, X.; Wang, G.; Zhang, A. Adiponectin protects Leydig cells against proinflammatory cytokines by suppressing the nuclear factor-kappaB signaling pathway. FEBS J. 2013, 280, 3920–3927. [Google Scholar] [CrossRef] [PubMed]
- Chappaz, E.; Albornoz, M.S.; Campos, D.; Che, L.; Palin, M.F.; Murphy, B.D.; Bordignon, V. Adiponectin enhances in vitro development of swine embryos. Domest. Anim. Endocrinol. 2008, 35, 198–207. [Google Scholar] [CrossRef]
- Gomes, E.T.; Costa, J.A.S.; Silva, D.M.F.; Al Shebli, W.; Azevedo, M.L.; Monteiro, P.L.J., Jr.; Araujo Silva, R.A.J.; Santos Filho, A.S.; Guerra, M.M.P.; Bartolomeu, C.C.; et al. Effects of adiponectin during in vitro maturation of goat oocytes: MEK 1/2 pathway and gene expression pattern. Reprod. Domest. Anim. 2018, 53, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.S.; Liu, Z.; Kawai, T.; Tabata, K.; Watanabe, H.; Suresh, D.; Kuo, F.T.; Pisarska, M.D.; Shimada, M. Adiponectin and its receptors modulate granulosa cell and cumulus cell functions, fertility, and early embryo development in the mouse and human. Fertil. Steril. 2012, 98, 471–479. [Google Scholar] [CrossRef]
- Oliveira, B.S.P.; Costa, J.A.S.; Gomes, E.T.; Silva, D.M.F.; Torres, S.M.; Monteiro, P.L.J., Jr.; Santos Filho, A.S.; Guerra, M.M.P.; Carneiro, G.F.; Wischral, A.; et al. Expression of adiponectin and its receptors (AdipoR1 and AdipoR2) in goat ovary and its effect on oocyte nuclear maturation in vitro. Theriogenology 2017, 104, 127–133. [Google Scholar] [CrossRef]
- Wang, Q.; Tang, S.B.; Song, X.B.; Deng, T.F.; Zhang, T.T.; Yin, S.; Luo, S.M.; Shen, W.; Zhang, C.L.; Ge, Z.J. High-glucose concentrations change DNA methylation levels in human IVM oocytes. Hum. Reprod. 2018, 33, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, V.R.; Kasimanickam, R.K.; Kastelic, J.P.; Stevenson, J.S. Associations of adiponectin and fertility estimates in Holstein bulls. Theriogenology 2013, 79, 766–777.e3. [Google Scholar] [CrossRef]
- Kadivar, A.; Heidari Khoei, H.; Hassanpour, H.; Golestanfar, A.; Ghanaei, H. Correlation of Adiponectin mRNA Abundance and Its Receptors with Quantitative Parameters of Sperm Motility in Rams. Int. J. Fertil. Steril. 2016, 10, 127–135. [Google Scholar] [PubMed]
- Mellouk, N.; Rame, C.; Delaveau, J.; Rat, C.; Maurer, E.; Froment, P.; Dupont, J. Adipokines expression profile in liver, adipose tissue and muscle during chicken embryo development. Gen. Comp. Endocrinol. 2018, 267, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Nishio, S.; Gibert, Y.; Bernard, L.; Brunet, F.; Triqueneaux, G.; Laudet, V. Adiponectin and adiponectin receptor genes are coexpressed during zebrafish embryogenesis and regulated by food deprivation. Dev. Dyn. 2008, 237, 1682–1690. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.T.; Marquard, K.; Stephens, S.; Louden, E.; Allsworth, J.; Moley, K.H. Adiponectin and adiponectin receptors in the mouse preimplantation embryo and uterus. Hum. Reprod. 2011, 26, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Cikos, S.; Burkus, J.; Bukovska, A.; Fabian, D.; Rehak, P.; Koppel, J. Expression of adiponectin receptors and effects of adiponectin isoforms in mouse preimplantation embryos. Hum. Reprod. 2010, 25, 2247–2255. [Google Scholar] [CrossRef]
- Zglejc-Waszak, K.; Waszkiewicz, E.M.; Franczak, A. Periconceptional undernutrition affects the levels of DNA methylation in the peri-implantation pig endometrium and in embryos. Theriogenology 2019, 123, 185–193. [Google Scholar] [CrossRef]
- Sturmey, R.G.; Reis, A.; Leese, H.J.; McEvoy, T.G. Role of fatty acids in energy provision during oocyte maturation and early embryo development. Reprod. Domest. Anim. 2009, 44 (Suppl. 3), 50–58. [Google Scholar] [CrossRef]
- Stubbs, C.D.; Smith, A.D. The modification of mammalian membrane polyunsaturated fatty acid composition in relation to membrane fluidity and function. Biochim. Biophys. Acta 1984, 779, 89–137. [Google Scholar] [CrossRef]
- Schindler, M.; Pendzialek, M.; Grybel, K.J.; Seeling, T.; Gurke, J.; Fischer, B.; Navarrete Santos, A. Adiponectin stimulates lipid metabolism via AMPK in rabbit blastocysts. Hum. Reprod. 2017, 32, 1382–1392. [Google Scholar] [CrossRef] [PubMed]
- Benaitreau, D.; Dos Santos, E.; Leneveu, M.C.; Alfaidy, N.; Feige, J.J.; de Mazancourt, P.; Pecquery, R.; Dieudonne, M.N. Effects of adiponectin on human trophoblast invasion. J. Endocrinol. 2010, 207, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Benaitreau, D.; Dos Santos, E.; Leneveu, M.C.; De Mazancourt, P.; Pecquery, R.; Dieudonne, M.N. Adiponectin promotes syncytialisation of BeWo cell line and primary trophoblast cells. Reprod. Biol. Endocrinol. 2010, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Mazaki-Tovi, S.; Kanety, H.; Pariente, C.; Hemi, R.; Wiser, A.; Schiff, E.; Sivan, E. Maternal serum adiponectin levels during human pregnancy. J. Perinatol. 2007, 27, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Mazaki-Tovi, S.; Romero, R.; Kusanovic, J.P.; Erez, O.; Vaisbuch, E.; Gotsch, F.; Mittal, P.; Than, G.N.; Nhan-Chang, C.; Chaiworapongsa, T.; et al. Adiponectin multimers in maternal plasma. J. Matern. Fetal Neonatal Med. 2008, 21, 796–815. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Osuga, Y.; Yamauchi, T.; Kobayashi, M.; Harada, M.; Hirata, T.; Morimoto, C.; Hirota, Y.; Yoshino, O.; Koga, K.; et al. Expression of adiponectin receptors and its possible implication in the human endometrium. Endocrinology 2006, 147, 3203–3210. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, E.; Pecquery, R.; de Mazancourt, P.; Dieudonne, M.N. Adiponectin and reproduction. Vitam. Horm. 2012, 90, 187–209. [Google Scholar] [CrossRef]
- Bohlouli, S.; Khazaei, M.; Teshfam, M.; Hassanpour, H. Adiponectin effect on the viability of human endometrial stromal cells and mRNA expression of adiponectin receptors. Int. J. Fertil. Steril. 2013, 7, 43–48. [Google Scholar]
- McDonald, E.A.; Wolfe, M.W. The pro-inflammatory role of adiponectin at the maternal-fetal interface. Am. J. Reprod. Immunol. 2011, 66, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Lappas, M.; Permezel, M.; Rice, G.E. Leptin and adiponectin stimulate the release of proinflammatory cytokines and prostaglandins from human placenta and maternal adipose tissue via nuclear factor-kappaB, peroxisomal proliferator-activated receptor-gamma and extracellularly regulated kinase 1/2. Endocrinology 2005, 146, 3334–3342. [Google Scholar] [CrossRef]
- Benaitreau, D.; Dieudonne, M.N.; Dos Santos, E.; Leneveu, M.C.; Mazancourt, P.; Pecquery, R. Antiproliferative effects of adiponectin on human trophoblastic cell lines JEG-3 and BeWo. Biol. Reprod. 2009, 80, 1107–1114. [Google Scholar] [CrossRef]
- Chen, H.; Chen, H.; Wu, Y.; Liu, B.; Li, Z.; Wang, Z. Adiponectin exerts antiproliferative effect on human placenta via modulation of the JNK/c-Jun pathway. Int. J. Clin. Exp. Pathol. 2014, 7, 2894–2904. [Google Scholar] [PubMed]
- McDonald, E.A.; Wolfe, M.W. Adiponectin attenuation of endocrine function within human term trophoblast cells. Endocrinology 2009, 150, 4358–4365. [Google Scholar] [CrossRef]
- Li, W.; Geng, L.; Liu, X.; Gui, W.; Qi, H. Recombinant adiponectin alleviates abortion in mice by regulating Th17/Treg imbalance via p38MAPK-STAT5 pathway. Biol. Reprod. 2018. [Google Scholar] [CrossRef] [PubMed]
- Duval, F.; Dos Santos, E.; Maury, B.; Serazin, V.; Fathallah, K.; Vialard, F.; Dieudonne, M.N. Adiponectin regulates glycogen metabolism at the human fetal-maternal interface. J. Mol. Endocrinol. 2018, 61, 139–152. [Google Scholar] [CrossRef]
- Rosario, F.J.; Schumacher, M.A.; Jiang, J.; Kanai, Y.; Powell, T.L.; Jansson, T. Chronic maternal infusion of full-length adiponectin in pregnant mice down-regulates placental amino acid transporter activity and expression and decreases fetal growth. J. Physiol. 2012, 590, 1495–1509. [Google Scholar] [CrossRef]
- Aye, I.L.; Powell, T.L.; Jansson, T. Review: Adiponectin—The missing link between maternal adiposity, placental transport and fetal growth? Placenta 2013, 34, S40–S45. [Google Scholar] [CrossRef]
- Jones, H.N.; Jansson, T.; Powell, T.L. Full-length adiponectin attenuates insulin signaling and inhibits insulin-stimulated amino Acid transport in human primary trophoblast cells. Diabetes 2010, 59, 1161–1170. [Google Scholar] [CrossRef]
- Caminos, J.E.; Nogueiras, R.; Gallego, R.; Bravo, S.; Tovar, S.; Garcia-Caballero, T.; Casanueva, F.F.; Dieguez, C. Expression and regulation of adiponectin and receptor in human and rat placenta. J. Clin. Endocrinol. Metab. 2005, 90, 4276–4286. [Google Scholar] [CrossRef] [PubMed]
- Aye, I.L.; Gao, X.; Weintraub, S.T.; Jansson, T.; Powell, T.L. Adiponectin inhibits insulin function in primary trophoblasts by PPARalpha-mediated ceramide synthesis. Mol. Endocrinol. 2014, 28, 512–524. [Google Scholar] [CrossRef]
- Aye, I.L.; Rosario, F.J.; Powell, T.L.; Jansson, T. Adiponectin supplementation in pregnant mice prevents the adverse effects of maternal obesity on placental function and fetal growth. Proc. Natl. Acad. Sci. USA 2015, 112, 12858–12863. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shang, L.X.; Dong, X.; Wang, X.; Wu, N.; Wang, S.H.; Zhang, F.; Xu, L.M.; Xiao, Y. Relationship of adiponectin and resistin levels in umbilical serum, maternal serum and placenta with neonatal birth weight. Aust. N. Z. J. Obstet. Gynaecol. 2010, 50, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Nanda, S.; Yu, C.K.; Giurcaneanu, L.; Akolekar, R.; Nicolaides, K.H. Maternal serum adiponectin at 11-13 weeks of gestation in preeclampsia. Fetal Diagn. Ther. 2011, 29, 208–215. [Google Scholar] [CrossRef]
- Worda, C.; Leipold, H.; Gruber, C.; Kautzky-Willer, A.; Knofler, M.; Bancher-Todesca, D. Decreased plasma adiponectin concentrations in women with gestational diabetes mellitus. Am. J. Obstet. Gynecol. 2004, 191, 2120–2124. [Google Scholar] [CrossRef]
- Maayan-Metzger, A.; Schushan-Eisen, I.; Strauss, T.; Globus, O.; Leibovitch, L. Gestational weight gain and body mass indexes have an impact on the outcomes of diabetic mothers and infants. Acta Paediatr. 2015, 104, 1150–1155. [Google Scholar] [CrossRef]
- Qiao, L.; Wattez, J.S.; Lee, S.; Guo, Z.; Schaack, J.; Hay, W.W., Jr.; Zita, M.M.; Parast, M.; Shao, J. Knockout maternal adiponectin increases fetal growth in mice: Potential role for trophoblast IGFBP-1. Diabetologia 2016, 59, 2417–2425. [Google Scholar] [CrossRef]
- Teede, H.; Deeks, A.; Moran, L. Polycystic ovary syndrome: A complex condition with psychological, reproductive and metabolic manifestations that impacts on health across the lifespan. BMC Med. 2010, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Teede, H.J.; Misso, M.L.; Boyle, J.A.; Garad, R.M.; McAllister, V.; Downes, L.; Gibson, M.; Hart, R.J.; Rombauts, L.; Moran, L.; et al. Translation and implementation of the Australian-led PCOS guideline: Clinical summary and translation resources from the International Evidence-based Guideline for the Assessment and Management of Polycystic Ovary Syndrome. Med. J. Aust. 2018, 209, S3–S8. [Google Scholar] [CrossRef]
- Toulis, K.A.; Goulis, D.G.; Farmakiotis, D.; Georgopoulos, N.A.; Katsikis, I.; Tarlatzis, B.C.; Papadimas, I.; Panidis, D. Adiponectin levels in women with polycystic ovary syndrome: A systematic review and a meta-analysis. Hum. Reprod. Update 2009, 15, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Benrick, A.; Chanclon, B.; Micallef, P.; Wu, Y.; Hadi, L.; Shelton, J.M.; Stener-Victorin, E.; Wernstedt Asterholm, I. Adiponectin protects against development of metabolic disturbances in a PCOS mouse model. Proc. Natl. Acad. Sci. USA 2017, 114, E7187–E7196. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. Positions statement: Criteria for defining polycystic ovary syndrome as a predominantly hyperandrogenic syndrome: An Androgen Excess Society guideline. J. Clin. Endocrinol. Metab. 2006, 91, 4237–4245. [Google Scholar] [CrossRef] [PubMed]
- Groth, S.W. Adiponectin and polycystic ovary syndrome. Biol. Res. Nurs. 2010, 12, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, Z.; Hao, C.; Tian, Y.; Fu, J. Effects of ADIPOQ polymorphisms on PCOS risk: A meta-analysis. Reprod. Biol. Endocrinol. 2018, 16, 120. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, X.; Zhong, H.; Peng, Q.; Chen, S.; Xie, Y.; Qin, X.; Qin, A. Low circulating adiponectin levels in women with polycystic ovary syndrome: An updated meta-analysis. Tumour Biol. 2014, 35, 3961–3973. [Google Scholar] [CrossRef]
- O’Connor, A.; Phelan, N.; Tun, T.K.; Boran, G.; Gibney, J.; Roche, H.M. High-molecular-weight adiponectin is selectively reduced in women with polycystic ovary syndrome independent of body mass index and severity of insulin resistance. J. Clin. Endocrinol. Metab. 2010, 95, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Panidis, D.; Kourtis, A.; Farmakiotis, D.; Mouslech, T.; Rousso, D.; Koliakos, G. Serum adiponectin levels in women with polycystic ovary syndrome. Hum. Reprod. 2003, 18, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Orio, F., Jr.; Palomba, S.; Cascella, T.; Milan, G.; Mioni, R.; Pagano, C.; Zullo, F.; Colao, A.; Lombardi, G.; Vettor, R. Adiponectin levels in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2003, 88, 2619–2623. [Google Scholar] [CrossRef]
- Lecke, S.B.; Mattei, F.; Morsch, D.M.; Spritzer, P.M. Abdominal subcutaneous fat gene expression and circulating levels of leptin and adiponectin in polycystic ovary syndrome. Fertil. Steril. 2011, 95, 2044–2049. [Google Scholar] [CrossRef]
- Svendsen, P.F.; Christiansen, M.; Hedley, P.L.; Nilas, L.; Pedersen, S.B.; Madsbad, S. Adipose expression of adipocytokines in women with polycystic ovary syndrome. Fertil. Steril. 2012, 98, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Tao, T.; Xu, B.; Liu, W. Ovarian HMW adiponectin is associated with folliculogenesis in women with polycystic ovary syndrome. Reprod. Biol. Endocrinol. 2013, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Artimani, T.; Saidijam, M.; Aflatoonian, R.; Ashrafi, M.; Amiri, I.; Yavangi, M.; SoleimaniAsl, S.; Shabab, N.; Karimi, J.; Mehdizadeh, M. Downregulation of adiponectin system in granulosa cells and low levels of HMW adiponectin in PCOS. J. Assist. Reprod. Genet. 2016, 33, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Inal, H.A.; Yilmaz, N.; Gorkem, U.; Oruc, A.S.; Timur, H. The impact of follicular fluid adiponectin and ghrelin levels based on BMI on IVF outcomes in PCOS. J. Endocrinol. Investig. 2016, 39, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Dehghan, R.; Saidijam, M.; Mehdizadeh, M.; Shabab, N.; Yavangi, M.; Artimani, T. Evidence for decreased expression of APPL1 associated with reduced insulin and adiponectin receptors expression in PCOS patients. J. Endocrinol. Investig. 2016, 39, 1075–1082. [Google Scholar] [CrossRef]
- Xu, A.; Chan, K.W.; Hoo, R.L.; Wang, Y.; Tan, K.C.; Zhang, J.; Chen, B.; Lam, M.C.; Tse, C.; Cooper, G.J.; et al. Testosterone selectively reduces the high molecular weight form of adiponectin by inhibiting its secretion from adipocytes. J. Biol. Chem. 2005, 280, 18073–18080. [Google Scholar] [CrossRef]
- Tan, B.K.; Chen, J.; Digby, J.E.; Keay, S.D.; Kennedy, C.R.; Randeva, H.S. Upregulation of adiponectin receptor 1 and 2 mRNA and protein in adipose tissue and adipocytes in insulin-resistant women with polycystic ovary syndrome. Diabetologia 2006, 49, 2723–2728. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.B.; Palin, M.F.; Bordignon, V.; Murphy, B.D. The ‘beneficial’ adipokines in reproduction and fertility. Int. J. Obes. (Lond.) 2008, 32, 223–231. [Google Scholar] [CrossRef]
- Ranjzad, F.; Mahmoudi, T.; Irani Shemirani, A.; Mahban, A.; Nikzamir, A.; Vahedi, M.; Ashrafi, M.; Gourabi, H. A common variant in the adiponectin gene and polycystic ovary syndrome risk. Mol. Biol. Rep. 2012, 39, 2313–2319. [Google Scholar] [CrossRef] [PubMed]
- Alfaqih, M.A.; Khader, Y.S.; Al-Dwairi, A.N.; Alzoubi, A.; Al-Shboul, O.; Hatim, A. Lower Levels of Serum Adiponectin and the T Allele of rs1501299 of the ADIPOQ Gene Are Protective against Polycystic Ovarian Syndrome in Jordan. Korean J. Fam. Med. 2018, 39, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, E.; Rafraf, M.; Farzadi, L.; Asghari-Jafarabadi, M.; Sabour, S. Effects of omega-3 fatty acids supplementation on serum adiponectin levels and some metabolic risk factors in women with polycystic ovary syndrome. Asia Pac. J. Clin. Nutr. 2012, 21, 511–518. [Google Scholar] [PubMed]
- Yang, K.; Zeng, L.; Bao, T.; Ge, J. Effectiveness of Omega-3 fatty acid for polycystic ovary syndrome: A systematic review and meta-analysis. Reprod. Biol. Endocrinol. 2018, 16, 27. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Bora, P.; Krishna, A. Direct action of adiponectin ameliorates increased androgen synthesis and reduces insulin receptor expression in the polycystic ovary. Biochem. Biophys. Res. Commun. 2017, 488, 509–515. [Google Scholar] [CrossRef]
- Singh, A.; Bora, P.; Krishna, A. Systemic adiponectin treatment reverses polycystic ovary syndrome-like features in an animal model. Reprod. Fertil. Dev. 2017. [Google Scholar] [CrossRef]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, I.; Kameda, S.; Hoshi, K. Early detection of ovarian and fallopian tube cancer by examination of cytological samples from the endometrial cavity. Br. J. Cancer 2013, 109, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Romero, I.; Bast, R.C., Jr. Minireview: Human ovarian cancer: Biology, current management, and paths to personalizing therapy. Endocrinology 2012, 153, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Men, X.; Zhang, W.; Lei, P. Advances in tumor markers of ovarian cancer for early diagnosis. Indian J. Cancer 2014, 51 (Suppl. 3), e72–e76. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, Y.; Funahashi, T.; Kihara, S.; Taguchi, T.; Tamaki, Y.; Matsuzawa, Y.; Noguchi, S. Association of serum adiponectin levels with breast cancer risk. Clin. Cancer Res. 2003, 9, 5699–5704. [Google Scholar] [PubMed]
- Goktas, S.; Yilmaz, M.I.; Caglar, K.; Sonmez, A.; Kilic, S.; Bedir, S. Prostate cancer and adiponectin. Urology 2005, 65, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Oritani, K.; Takahashi, I.; Ishikawa, J.; Matsuyama, A.; Ouchi, N.; Kihara, S.; Funahashi, T.; Tenner, A.J.; Tomiyama, Y.; et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood 2000, 96, 1723–1732. [Google Scholar]
- Brakenhielm, E.; Veitonmaki, N.; Cao, R.; Kihara, S.; Matsuzawa, Y.; Zhivotovsky, B.; Funahashi, T.; Cao, Y. Adiponectin-induced antiangiogenesis and antitumor activity involve caspase-mediated endothelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 2476–2481. [Google Scholar] [CrossRef]
- Hoffmann, M.; Gogola, J.; Ptak, A. Adiponectin Reverses the Proliferative Effects of Estradiol and IGF-1 in Human Epithelial Ovarian Cancer Cells by Downregulating the Expression of Their Receptors. Horm. Cancer 2018, 9, 166–174. [Google Scholar] [CrossRef]
- Li, X.; Yu, Z.; Fang, L.; Liu, F.; Jiang, K. Expression of Adiponectin Receptor-1 and Prognosis of Epithelial Ovarian Cancer Patients. Med. Sci. Monit. 2017, 23, 1514–1521. [Google Scholar] [CrossRef]
- Nagaraju, G.P.; Rajitha, B.; Aliya, S.; Kotipatruni, R.P.; Madanraj, A.S.; Hammond, A.; Park, D.; Chigurupati, S.; Alam, A.; Pattnaik, S. The role of adiponectin in obesity-associated female-specific carcinogenesis. Cytokine Growth Factor Rev. 2016, 31, 37–48. [Google Scholar] [CrossRef]
- Takemura, Y.; Osuga, Y.; Harada, M.; Hirata, T.; Koga, K.; Yoshino, O.; Hirota, Y.; Morimoto, C.; Yano, T.; Taketani, Y. Concentration of adiponectin in peritoneal fluid is decreased in women with endometriosis. Am. J. Reprod. Immunol. 2005, 54, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Soliman, P.T.; Wu, D.; Tortolero-Luna, G.; Schmeler, K.M.; Slomovitz, B.M.; Bray, M.S.; Gershenson, D.M.; Lu, K.H. Association between adiponectin, insulin resistance, and endometrial cancer. Cancer 2006, 106, 2376–2381. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.W.; Shin, J.H.; Park, H.T.; Kim, T.; Kim, S.H.; Hur, J.Y. Resistin concentration is increased in the peritoneal fluid of women with endometriosis. Am. J. Reprod. Immunol. 2010, 64, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Kriplani, A.; Yadav, R.K.; Lyngdoh, B.T.; Mahapatra, S.C. Peritoneal fluid leptin levels are increased but adiponectin levels are not changed in infertile patients with pelvic endometriosis. Gynecol. Endocrinol. 2010, 26, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Oh, H.K.; Choi, J.H. Expression of adiponectin, leptin, and their receptors in ovarian endometrioma. Fertil. Steril. 2013, 100, 135–141.e2. [Google Scholar] [CrossRef] [PubMed]
- Ashizawa, N.; Yahata, T.; Quan, J.; Adachi, S.; Yoshihara, K.; Tanaka, K. Serum leptin-adiponectin ratio and endometrial cancer risk in postmenopausal female subjects. Gynecol. Oncol. 2010, 119, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Petridou, E.; Mantzoros, C.; Dessypris, N.; Koukoulomatis, P.; Addy, C.; Voulgaris, Z.; Chrousos, G.; Trichopoulos, D. Plasma adiponectin concentrations in relation to endometrial cancer: A case-control study in Greece. J. Clin. Endocrinol. Metab. 2003, 88, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Dal Maso, L.; Augustin, L.S.; Karalis, A.; Talamini, R.; Franceschi, S.; Trichopoulos, D.; Mantzoros, C.S.; La Vecchia, C. Circulating adiponectin and endometrial cancer risk. J. Clin. Endocrinol. Metab. 2004, 89, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Cust, A.E.; Kaaks, R.; Friedenreich, C.; Bonnet, F.; Laville, M.; Lukanova, A.; Rinaldi, S.; Dossus, L.; Slimani, N.; Lundin, E.; et al. Plasma adiponectin levels and endometrial cancer risk in pre- and postmenopausal women. J. Clin. Endocrinol. Metab. 2007, 92, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.S.; Chamberland, J.P.; Aronis, K.; Tseleni-Balafouta, S.; Mantzoros, C.S. Direct role of adiponectin and adiponectin receptors in endometrial cancer: In vitro and ex vivo studies in humans. Mol. Cancer Ther. 2011, 10, 2234–2243. [Google Scholar] [CrossRef] [PubMed]
- Taliaferro-Smith, L.; Nagalingam, A.; Zhong, D.; Zhou, W.; Saxena, N.K.; Sharma, D. LKB1 is required for adiponectin-mediated modulation of AMPK-S6K axis and inhibition of migration and invasion of breast cancer cells. Oncogene 2009, 28, 2621–2633. [Google Scholar] [CrossRef]
- American Diabetes, A. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42, S13–S28. [Google Scholar] [CrossRef] [PubMed]
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Reynolds, C.M.; Vickers, M.H. The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3342. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.A.; Brelje, T.C.; Sorenson, R.L. Adaptation of islets of Langerhans to pregnancy: Increased islet cell proliferation and insulin secretion correlates with the onset of placental lactogen secretion. Endocrinology 1992, 130, 1459–1466. [Google Scholar] [CrossRef]
- Retnakaran, R. Adiponectin and beta-Cell Adaptation in Pregnancy. Diabetes 2017, 66, 1121–1122. [Google Scholar] [CrossRef]
- Buckley, B.S.; Harreiter, J.; Damm, P.; Corcoy, R.; Chico, A.; Simmons, D.; Vellinga, A.; Dunne, F.; Group, D.C.I. Gestational diabetes mellitus in Europe: Prevalence, current screening practice and barriers to screening. A review. Diabet. Med. 2012, 29, 844–854. [Google Scholar] [CrossRef]
- Catalano, P.M.; Hoegh, M.; Minium, J.; Huston-Presley, L.; Bernard, S.; Kalhan, S.; Hauguel-De Mouzon, S. Adiponectin in human pregnancy: Implications for regulation of glucose and lipid metabolism. Diabetologia 2006, 49, 1677–1685. [Google Scholar] [CrossRef]
- Lara-Castro, C.; Luo, N.; Wallace, P.; Klein, R.L.; Garvey, W.T. Adiponectin multimeric complexes and the metabolic syndrome trait cluster. Diabetes 2006, 55, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Wattez, J.S.; Lee, S.; Nguyen, A.; Schaack, J.; Hay, W.W., Jr.; Shao, J. Adiponectin Deficiency Impairs Maternal Metabolic Adaptation to Pregnancy in Mice. Diabetes 2017, 66, 1126–1135. [Google Scholar] [CrossRef]
- Pala, H.G.; Ozalp, Y.; Yener, A.S.; Gerceklioglu, G.; Uysal, S.; Onvural, A. Adiponectin levels in gestational diabetes mellitus and in pregnant women without glucose intolerance. Adv. Clin. Exp. Med. 2015, 24, 85–92. [Google Scholar] [CrossRef]
- Kajantie, E.; Hytinantti, T.; Hovi, P.; Andersson, S. Cord plasma adiponectin: A 20-fold rise between 24 weeks gestation and term. J. Clin. Endocrinol. Metab. 2004, 89, 4031–4036. [Google Scholar] [CrossRef]
- Chan, T.F.; Chung, Y.F.; Chen, H.S.; Su, J.H.; Yuan, S.S. Elevated amniotic fluid leptin levels in early second trimester are associated with earlier delivery and lower birthweight in twin pregnancy. Acta Obstet. Gynecol. Scand. 2004, 83, 707–710. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tan, B.; Karteris, E.; Zervou, S.; Digby, J.; Hillhouse, E.W.; Vatish, M.; Randeva, H.S. Secretion of adiponectin by human placenta: Differential modulation of adiponectin and its receptors by cytokines. Diabetologia 2006, 49, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Ategbo, J.M.; Grissa, O.; Yessoufou, A.; Hichami, A.; Dramane, K.L.; Moutairou, K.; Miled, A.; Grissa, A.; Jerbi, M.; Tabka, Z.; et al. Modulation of adipokines and cytokines in gestational diabetes and macrosomia. J. Clin. Endocrinol. Metab. 2006, 91, 4137–4143. [Google Scholar] [CrossRef]
- Horosz, E.; Bomba-Opon, D.A.; Szymanska, M.; Wielgos, M. Third trimester plasma adiponectin and leptin in gestational diabetes and normal pregnancies. Diabetes Res. Clin. Pract. 2011, 93, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Retnakaran, R.; Qi, Y.; Connelly, P.W.; Sermer, M.; Hanley, A.J.; Zinman, B. Low adiponectin concentration during pregnancy predicts postpartum insulin resistance, beta cell dysfunction and fasting glycaemia. Diabetologia 2010, 53, 268–276. [Google Scholar] [CrossRef]
- Radaelli, T.; Varastehpour, A.; Catalano, P.; Hauguel-de Mouzon, S. Gestational diabetes induces placental genes for chronic stress and inflammatory pathways. Diabetes 2003, 52, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Lodish, H.F. Insulin resistance in adipose tissue: Direct and indirect effects of tumor necrosis factor-alpha. Cytokine Growth Factor Rev. 2003, 14, 447–455. [Google Scholar] [CrossRef]
- Kern, P.A.; Di Gregorio, G.B.; Lu, T.; Rassouli, N.; Ranganathan, G. Adiponectin expression from human adipose tissue: Relation to obesity, insulin resistance, and tumor necrosis factor-alpha expression. Diabetes 2003, 52, 1779–1785. [Google Scholar] [CrossRef]
- Bao, W.; Baecker, A.; Song, Y.; Kiely, M.; Liu, S.; Zhang, C. Adipokine levels during the first or early second trimester of pregnancy and subsequent risk of gestational diabetes mellitus: A systematic review. Metabolism 2015, 64, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Iliodromiti, S.; Sassarini, J.; Kelsey, T.W.; Lindsay, R.S.; Sattar, N.; Nelson, S.M. Accuracy of circulating adiponectin for predicting gestational diabetes: A systematic review and meta-analysis. Diabetologia 2016, 59, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Qiu, C.; Muy-Rivera, M.; Vadachkoria, S.; Song, T.; Luthy, D.A. Plasma adiponectin concentrations in early pregnancy and subsequent risk of gestational diabetes mellitus. J. Clin. Endocrinol. Metab. 2004, 89, 2306–2311. [Google Scholar] [CrossRef] [PubMed]
- Lain, K.Y.; Daftary, A.R.; Ness, R.B.; Roberts, J.M. First trimester adipocytokine concentrations and risk of developing gestational diabetes later in pregnancy. Clin. Endocrinol. (Oxf.) 2008, 69, 407–411. [Google Scholar] [CrossRef]
- Lacroix, M.; Battista, M.C.; Doyon, M.; Menard, J.; Ardilouze, J.L.; Perron, P.; Hivert, M.F. Lower adiponectin levels at first trimester of pregnancy are associated with increased insulin resistance and higher risk of developing gestational diabetes mellitus. Diabetes Care 2013, 36, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, A.; Teler, J.; Maciejewska, A.; Sawczuk, M.; Safranow, K.; Dziedziejko, V. Adiponectin and leptin gene polymorphisms in women with gestational diabetes mellitus. J. Assist. Reprod. Genet. 2017, 34, 511–516. [Google Scholar] [CrossRef]
- Beltcheva, O.; Boyadzhieva, M.; Angelova, O.; Mitev, V.; Kaneva, R.; Atanasova, I. The rs266729 single-nucleotide polymorphism in the adiponectin gene shows association with gestational diabetes. Arch. Gynecol. Obstet. 2014, 289, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zheng, Y.L.; Fan, Y.P.; Liu, M.H.; Lu, X.Y.; Tao, Q. Association of adiponectin gene polymorphism 45TG with gestational diabetes mellitus diagnosed on the new IADPSG criteria, plasma adiponectin levels and adverse pregnancy outcomes. Clin. Exp. Med. 2015, 15, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Takhshid, M.A.; Haem, Z.; Aboualizadeh, F. The association of circulating adiponectin and + 45 T/G polymorphism of adiponectin gene with gestational diabetes mellitus in Iranian population. J. Diabetes Metab. Disord. 2015, 14, 30. [Google Scholar] [CrossRef]
- Low, C.F.; Mohd Tohit, E.R.; Chong, P.P.; Idris, F. Adiponectin SNP45TG is associated with gestational diabetes mellitus. Arch. Gynecol. Obstet. 2011, 283, 1255–1260. [Google Scholar] [CrossRef]
- Ye, R.; Wang, M.; Wang, Q.A.; Scherer, P.E. Adiponectin-mediated antilipotoxic effects in regenerating pancreatic islets. Endocrinology 2015, 156, 2019–2028. [Google Scholar] [CrossRef]
- Retnakaran, R.; Connelly, P.W.; Maguire, G.; Sermer, M.; Zinman, B.; Hanley, A.J. Decreased high-molecular-weight adiponectin in gestational diabetes: Implications for the pathophysiology of Type 2 diabetes. Diabet. Med. 2007, 24, 245–252. [Google Scholar] [CrossRef]
- Retnakaran, R.; Hanley, A.J.; Raif, N.; Hirning, C.R.; Connelly, P.W.; Sermer, M.; Kahn, S.E.; Zinman, B. Adiponectin and beta cell dysfunction in gestational diabetes: Pathophysiological implications. Diabetologia 2005, 48, 993–1001. [Google Scholar] [CrossRef]
- Abalos, E.; Cuesta, C.; Grosso, A.L.; Chou, D.; Say, L. Global and regional estimates of preeclampsia and eclampsia: A systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mayrink, J.; Costa, M.L.; Cecatti, J.G. Preeclampsia in 2018: Revisiting Concepts, Physiopathology, and Prediction. Sci. World J. 2018, 2018, 6268276. [Google Scholar] [CrossRef]
- Song, Y.; Gao, J.; Qu, Y.; Wang, S.; Wang, X.; Liu, J. Serum levels of leptin, adiponectin and resistin in relation to clinical characteristics in normal pregnancy and preeclampsia. Clin. Chim. Acta 2016, 458, 133–137. [Google Scholar] [CrossRef]
- Demir, B.C.; Atalay, M.A.; Ozerkan, K.; Doster, Y.; Ocakoglu, G.; Kucukkomurcu, S. Maternal adiponectin and visfatin concentrations in normal and complicated pregnancies. Clin. Exp. Obstet. Gynecol. 2013, 40, 261–267. [Google Scholar]
- Miehle, K.; Stepan, H.; Fasshauer, M. Leptin, adiponectin and other adipokines in gestational diabetes mellitus and pre-eclampsia. Clin. Endocrinol. (Oxf.) 2012, 76, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Mannisto, T.; Mendola, P.; Vaarasmaki, M.; Jarvelin, M.R.; Hartikainen, A.L.; Pouta, A.; Suvanto, E. Elevated blood pressure in pregnancy and subsequent chronic disease risk. Circulation 2013, 127, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, J.E.; Jamieson, N.; Greer, I.A.; Sattar, N. Paradoxical elevation in adiponectin concentrations in women with preeclampsia. Hypertension 2003, 42, 891–894. [Google Scholar] [CrossRef]
- Naruse, K.; Yamasaki, M.; Umekage, H.; Sado, T.; Sakamoto, Y.; Morikawa, H. Peripheral blood concentrations of adiponectin, an adipocyte-specific plasma protein, in normal pregnancy and preeclampsia. J. Reprod. Immunol. 2005, 65, 65–75. [Google Scholar] [CrossRef]
- Haugen, F.; Ranheim, T.; Harsem, N.K.; Lips, E.; Staff, A.C.; Drevon, C.A. Increased plasma levels of adipokines in preeclampsia: Relationship to placenta and adipose tissue gene expression. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E326–E333. [Google Scholar] [CrossRef] [PubMed]
- Eleuterio, N.M.; Palei, A.C.; Rangel Machado, J.S.; Tanus-Santos, J.E.; Cavalli, R.C.; Sandrim, V.C. Relationship between adiponectin and nitrite in healthy and preeclampsia pregnancies. Clin. Chim. Acta 2013, 423, 112–115. [Google Scholar] [CrossRef]
- Hendler, I.; Blackwell, S.C.; Mehta, S.H.; Whitty, J.E.; Russell, E.; Sorokin, Y.; Cotton, D.B. The levels of leptin, adiponectin, and resistin in normal weight, overweight, and obese pregnant women with and without preeclampsia. Am. J. Obstet. Gynecol. 2005, 193, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Osuga, Y.; Koga, K.; Tajima, T.; Hirota, Y.; Hirata, T.; Morimoto, C.; Harada, M.; Yano, T.; Taketani, Y. Selective increase in high molecular weight adiponectin concentration in serum of women with preeclampsia. J. Reprod. Immunol. 2007, 73, 60–65. [Google Scholar] [CrossRef]
- D’Anna, R.; Baviera, G.; Corrado, F.; Giordano, D.; De Vivo, A.; Nicocia, G.; Di Benedetto, A. Adiponectin and insulin resistance in early- and late-onset pre-eclampsia. BJOG 2006, 113, 1264–1269. [Google Scholar] [CrossRef]
- Cortelazzi, D.; Corbetta, S.; Ronzoni, S.; Pelle, F.; Marconi, A.; Cozzi, V.; Cetin, I.; Cortelazzi, R.; Beck-Peccoz, P.; Spada, A. Maternal and foetal resistin and adiponectin concentrations in normal and complicated pregnancies. Clin. Endocrinol. (Oxf.) 2007, 66, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Dalamaga, M.; Srinivas, S.K.; Elovitz, M.A.; Chamberland, J.; Mantzoros, C.S. Serum adiponectin and leptin in relation to risk for preeclampsia: Results from a large case-control study. Metabolism 2011, 60, 1539–1544. [Google Scholar] [CrossRef]
- Mazaki-Tovi, S.; Romero, R.; Vaisbuch, E.; Kusanovic, J.P.; Erez, O.; Gotsch, F.; Chaiworapongsa, T.; Than, N.G.; Kim, S.K.; Nhan-Chang, C.L.; et al. Maternal serum adiponectin multimers in preeclampsia. J. Perinat. Med. 2009, 37, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.S.; Palei, A.C.; Amaral, L.M.; Bueno, A.C.; Antonini, S.R.; Duarte, G.; Tanus-Santos, J.E.; Sandrim, V.C.; Cavalli, R.C. Polymorphisms of the adiponectin gene in gestational hypertension and pre-eclampsia. J. Hum. Hypertens. 2014, 28, 128–132. [Google Scholar] [CrossRef]
- Saarela, T.; Hiltunen, M.; Helisalmi, S.; Heinonen, S.; Laakso, M. Adiponectin gene haplotype is associated with preeclampsia. Genet. Test. 2006, 10, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Cheng, K.K.; Vanhoutte, P.M.; Lam, K.S.; Xu, A. Vascular effects of adiponectin: Molecular mechanisms and potential therapeutic intervention. Clin. Sci. (Lond.) 2008, 114, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef]
- Malhotra, A.; Allison, B.J.; Castillo-Melendez, M.; Jenkin, G.; Polglase, G.R.; Miller, S.L. Neonatal Morbidities of Fetal Growth Restriction: Pathophysiology and Impact. Front. Endocrinol. (Lausanne) 2019, 10, 55. [Google Scholar] [CrossRef]
- Ghidini, A. Idiopathic fetal growth restriction: A pathophysiologic approach. Obstet. Gynecol. Surv. 1996, 51, 376–382. [Google Scholar] [CrossRef]
- Sharp, A.N.; Heazell, A.E.; Crocker, I.P.; Mor, G. Placental apoptosis in health and disease. Am. J. Reprod. Immunol. 2010, 64, 159–169. [Google Scholar] [CrossRef]
- Straszewski-Chavez, S.L.; Abrahams, V.M.; Mor, G. The role of apoptosis in the regulation of trophoblast survival and differentiation during pregnancy. Endocr. Rev. 2005, 26, 877–897. [Google Scholar] [CrossRef] [PubMed]
- Gaccioli, F.; Lager, S. Placental Nutrient Transport and Intrauterine Growth Restriction. Front. Physiol. 2016, 7, 40. [Google Scholar] [CrossRef]
- Gardosi, J.; Kady, S.M.; McGeown, P.; Francis, A.; Tonks, A. Classification of stillbirth by relevant condition at death (ReCoDe): Population based cohort study. BMJ 2005, 331, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Shastri, S.; Sharma, P. Intrauterine Growth Restriction: Antenatal and Postnatal Aspects. Clin. Med. Insights Pediatr. 2016, 10, 67–83. [Google Scholar] [CrossRef]
- Goto, E. Blood adiponectin concentration at birth in small for gestational age neonates: A meta-analysis. Diabetes Metab. Syndr. 2019, 13, 183–188. [Google Scholar] [CrossRef]
- Evagelidou, E.N.; Giapros, V.I.; Challa, A.S.; Kiortsis, D.N.; Tsatsoulis, A.A.; Andronikou, S.K. Serum adiponectin levels, insulin resistance, and lipid profile in children born small for gestational age are affected by the severity of growth retardation at birth. Eur. J. Endocrinol. 2007, 156, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Buke, B.; Topcu, H.O.; Engin-Ustun, Y.; Danisman, N. Comparison of serum maternal adiponectin concentrations in women with isolated intrauterine growth retardation and intrauterine growth retardation concomitant with pre-eclampsia. J. Turk. Ger. Gynecol. Assoc. 2014, 15, 173–176. [Google Scholar] [CrossRef]
- Kyriakakou, M.; Malamitsi-Puchner, A.; Militsi, H.; Boutsikou, T.; Margeli, A.; Hassiakos, D.; Kanaka-Gantenbein, C.; Papassotiriou, I.; Mastorakos, G. Leptin and adiponectin concentrations in intrauterine growth restricted and appropriate for gestational age foetuses, neonates, and their mothers. Eur. J. Endocrinol. 2008, 158, 343–348. [Google Scholar] [CrossRef]
- Visentin, S.; Lapolla, A.; Londero, A.P.; Cosma, C.; Dalfra, M.; Camerin, M.; Faggian, D.; Plebani, M.; Cosmi, E. Adiponectin levels are reduced while markers of systemic inflammation and aortic remodelling are increased in intrauterine growth restricted mother-child couple. Biomed. Res. Int. 2014, 2014, 401595. [Google Scholar] [CrossRef] [PubMed]
- Lekva, T.; Roland, M.C.P.; Michelsen, A.E.; Friis, C.M.; Aukrust, P.; Bollerslev, J.; Henriksen, T.; Ueland, T. Large Reduction in Adiponectin During Pregnancy Is Associated with Large-for-Gestational-Age Newborns. J. Clin. Endocrinol. Metab. 2017, 102, 2552–2559. [Google Scholar] [CrossRef] [PubMed]
- Duval, F.; Santos, E.D.; Poidatz, D.; Serazin, V.; Gronier, H.; Vialard, F.; Dieudonne, M.N. Adiponectin Inhibits Nutrient Transporters and Promotes Apoptosis in Human Villous Cytotrophoblasts: Involvement in the Control of Fetal Growth. Biol. Reprod. 2016, 94, 111. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbe, A.; Bongrani, A.; Mellouk, N.; Estienne, A.; Kurowska, P.; Grandhaye, J.; Elfassy, Y.; Levy, R.; Rak, A.; Froment, P.; et al. Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions. Int. J. Mol. Sci. 2019, 20, 1526. https://doi.org/10.3390/ijms20071526
Barbe A, Bongrani A, Mellouk N, Estienne A, Kurowska P, Grandhaye J, Elfassy Y, Levy R, Rak A, Froment P, et al. Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions. International Journal of Molecular Sciences. 2019; 20(7):1526. https://doi.org/10.3390/ijms20071526
Chicago/Turabian StyleBarbe, Alix, Alice Bongrani, Namya Mellouk, Anthony Estienne, Patrycja Kurowska, Jérémy Grandhaye, Yaelle Elfassy, Rachel Levy, Agnieszka Rak, Pascal Froment, and et al. 2019. "Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions" International Journal of Molecular Sciences 20, no. 7: 1526. https://doi.org/10.3390/ijms20071526
APA StyleBarbe, A., Bongrani, A., Mellouk, N., Estienne, A., Kurowska, P., Grandhaye, J., Elfassy, Y., Levy, R., Rak, A., Froment, P., & Dupont, J. (2019). Mechanisms of Adiponectin Action in Fertility: An Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions. International Journal of Molecular Sciences, 20(7), 1526. https://doi.org/10.3390/ijms20071526