Specificity of the Metallothionein-1 Response by Cadmium-Exposed Normal Human Urothelial Cells


 , ,
, ,
Abstract
1. Introduction
2. Results
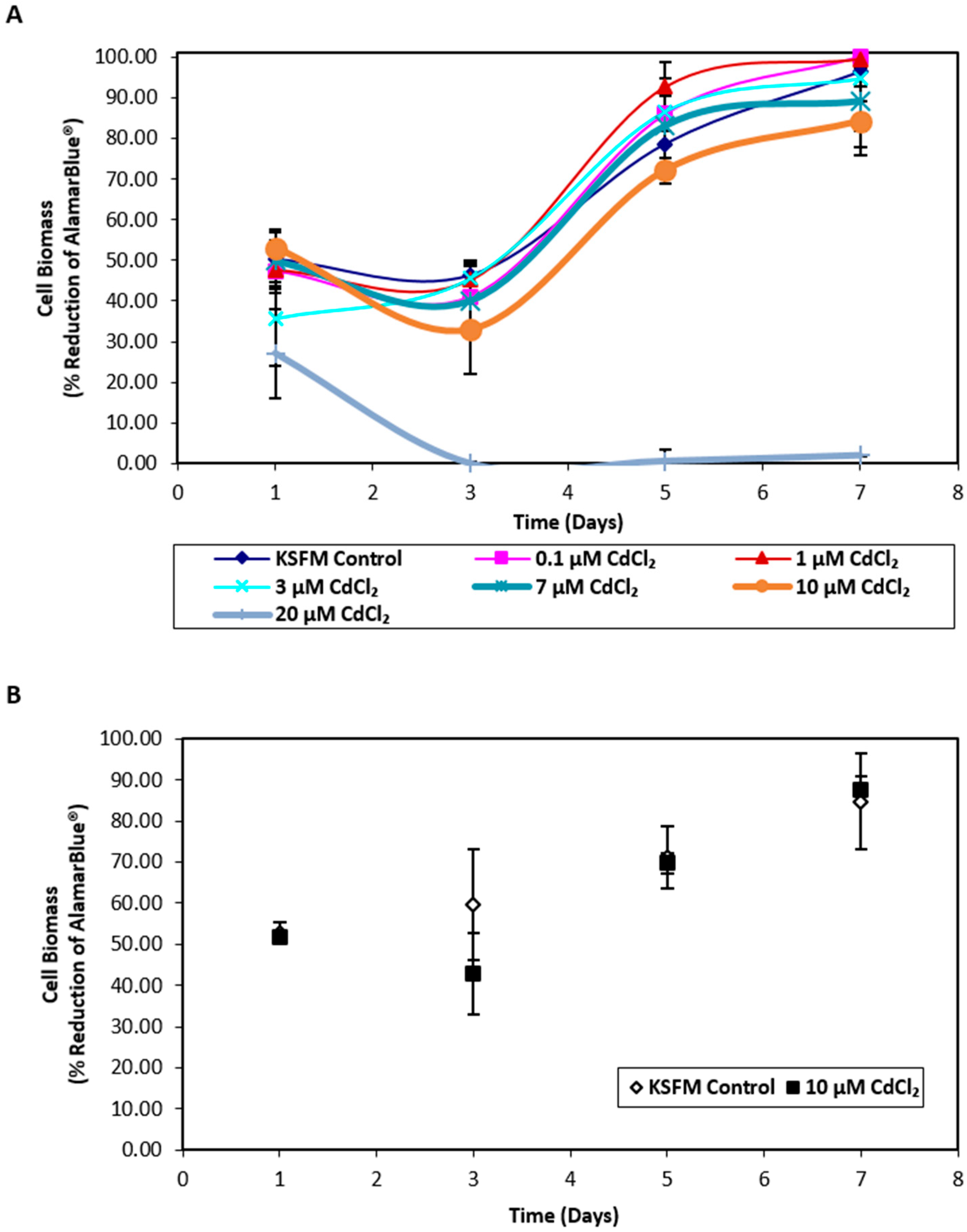
2.1. Influence of Cadmium on NHU Cell Culture Growth and Uptake of Cadmium across an Intact Urothelial Barrier
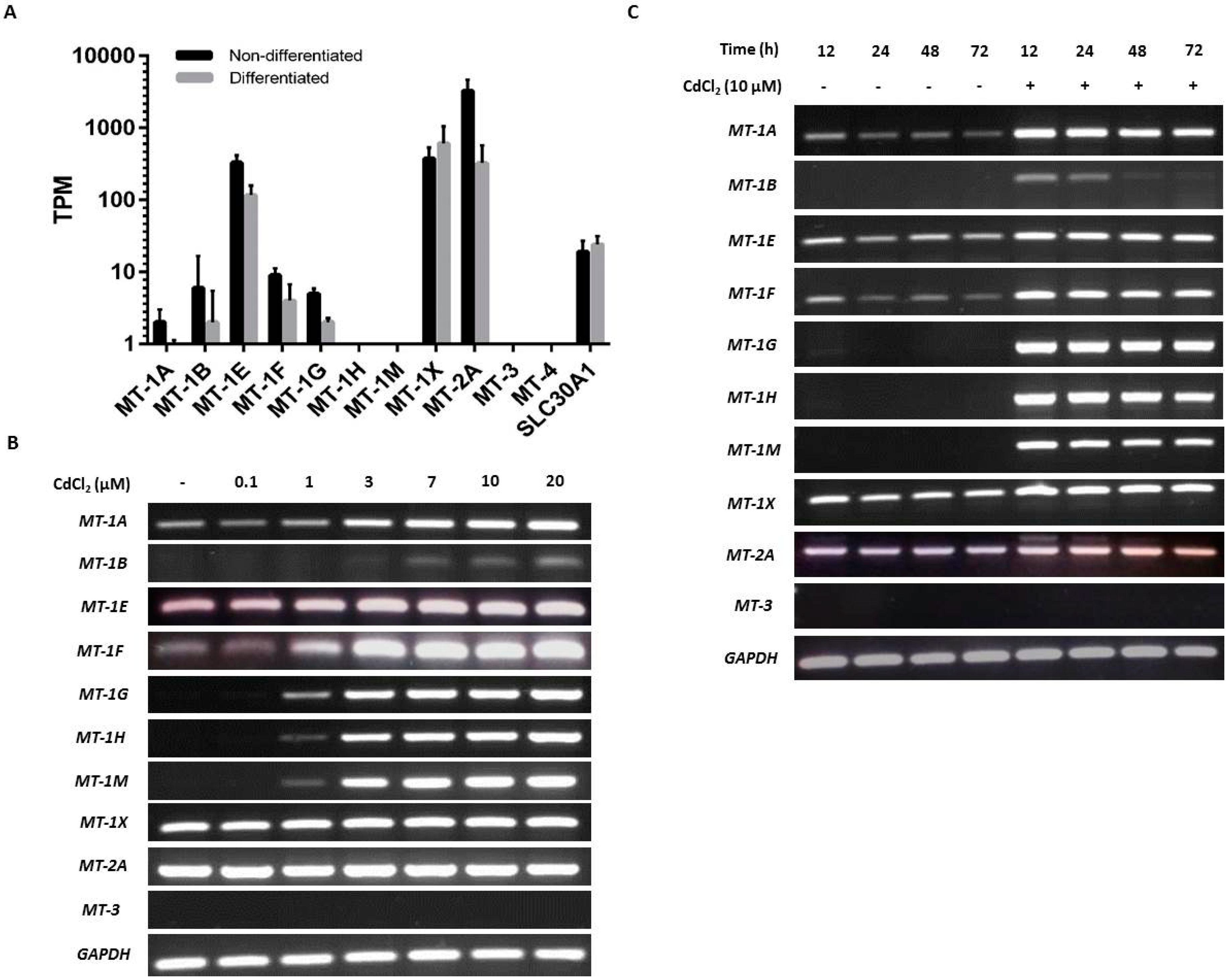
2.2. Baseline and Cadmium-Induced MT Transcription in NHU Cells
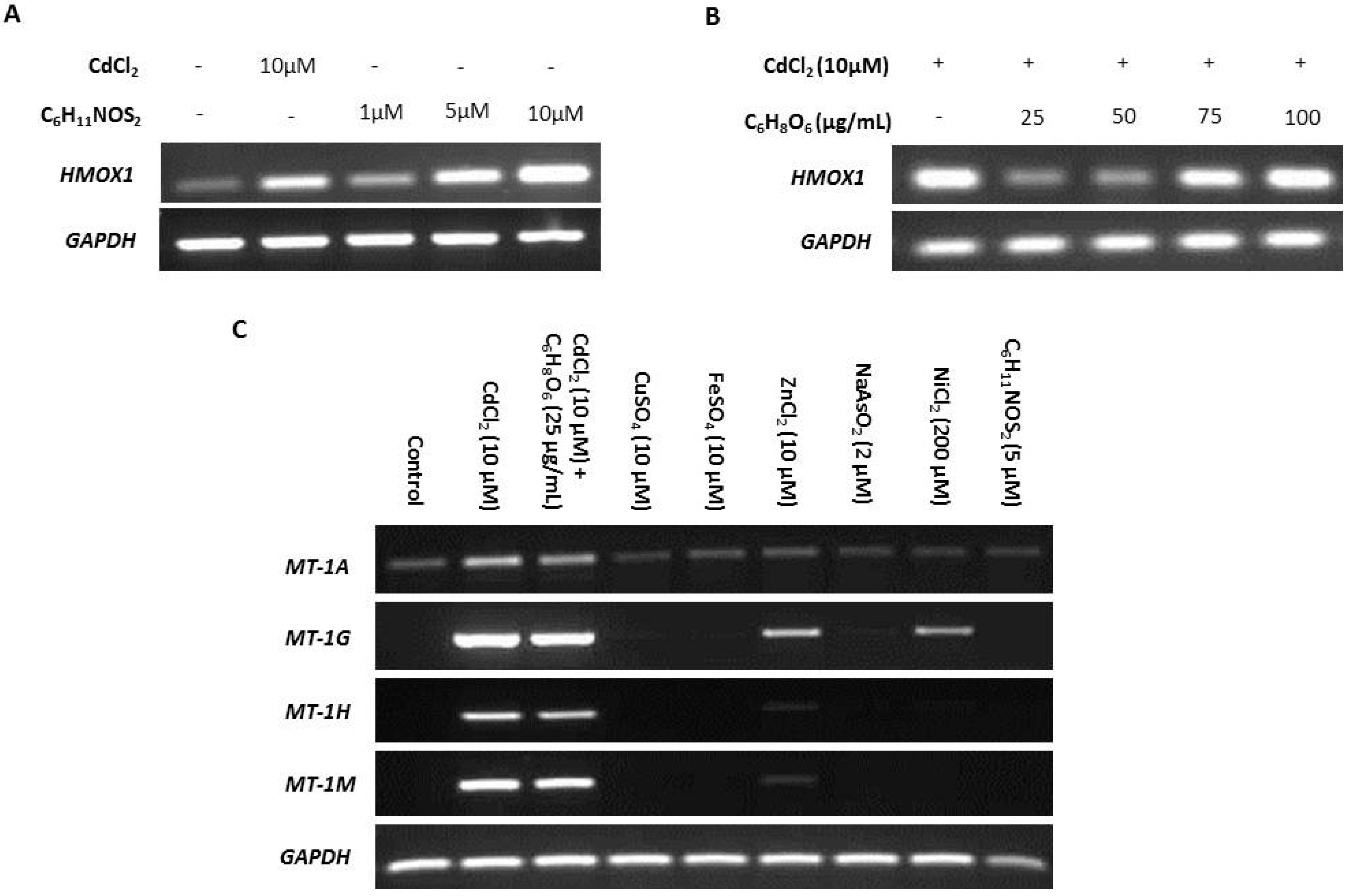
2.3. Specificity of Cadmium-Induced MT Transcription
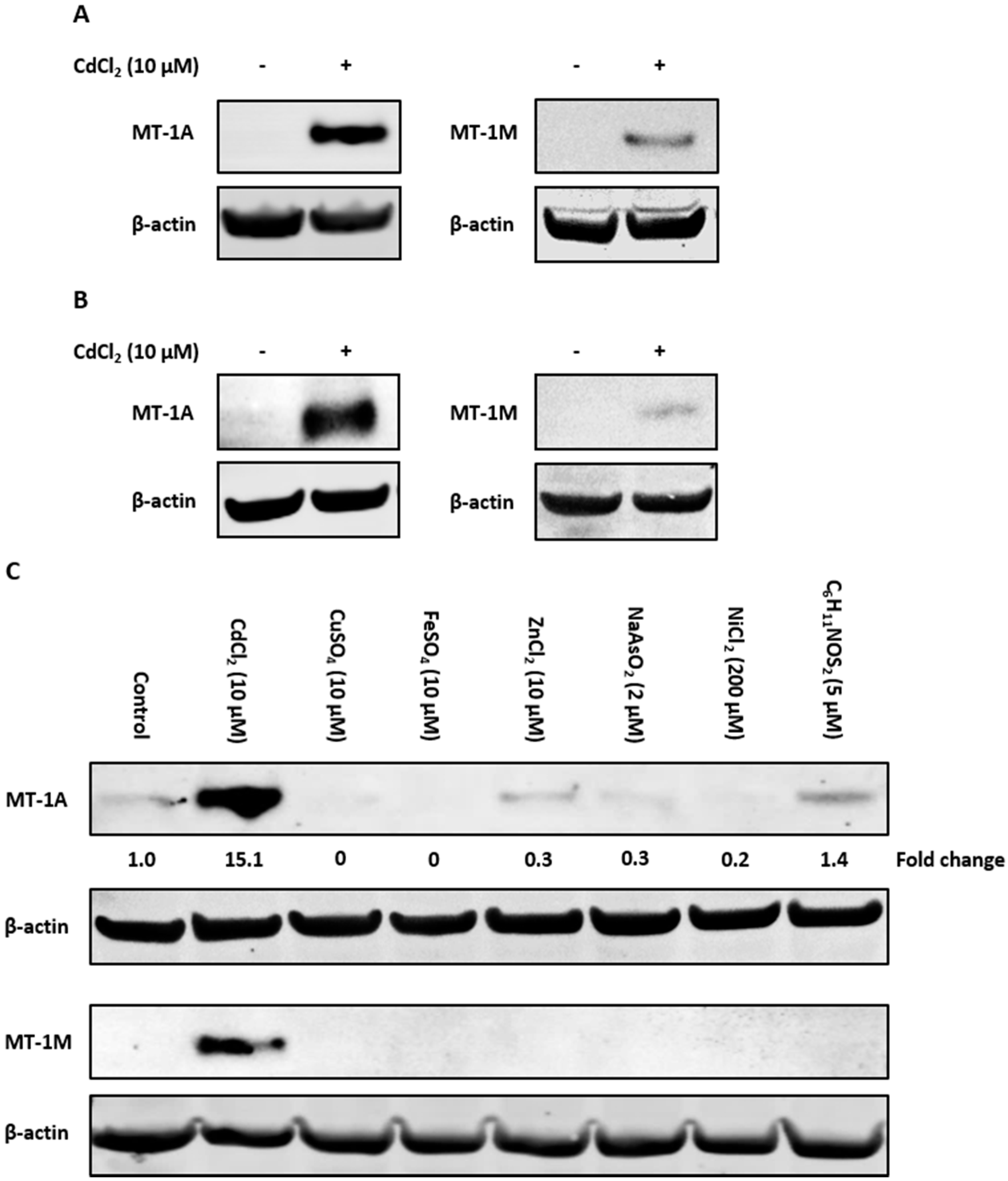
2.4. Immunoblotting With Isoform-Specific Antibodies
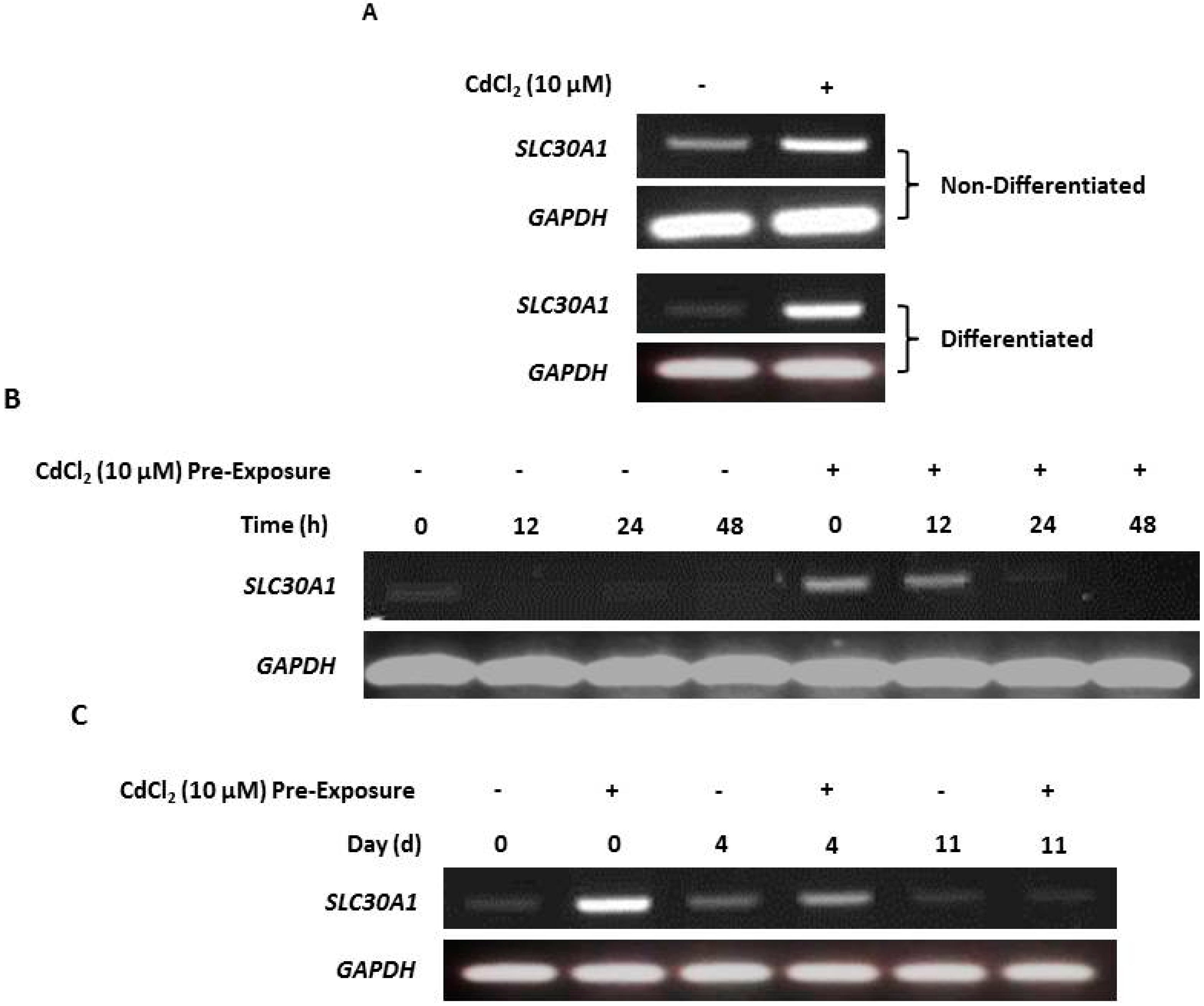
2.5. Upregulation of Zinc Transporter-1 (SLC30A1) Transcription in Cadmium-Exposed NHU Cells
3. Discussion
4. Materials and Methods
4.1. NHU Cell Culture and Exposure to Cadmium and Other Agents
4.2. Measurement of Transepithelial Electrical Resistance
4.3. MT Transcript Abundance Quantification by Next-Generation Sequencing
4.4. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.5. Western Blotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HMOX1 | Heme oxygenase-1 |
| MT | Metallothionein |
| NHU | Normal human urothelial |
| SLC30A1 | Solute carrier family A member 1 |
| TEER | Trans-epithelial electrical resistance |
References
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metals Toxicity and the Environment. EXS 2012, 101, 133–164. [Google Scholar]
- Feki-Tounsi, M.; Hamza-Chaffai, A. Cadmium as a possible cause of bladder cancer: A review of accumulated evidence. Environ. Sci. Pollut. Res. 2014, 21, 10561–10573. [Google Scholar] [CrossRef] [PubMed]
- Waalkes, M.P. Cadmium carcinogenesis. Mutat. Res. 2003, 533, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Agency for Toxic Substances and Disease Registry Toxicological Profile for Cadmium. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp.asp?id=48&tid=15 (accessed on 14 September 2016).
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Sigel, A.; Sigel, H.; Sigel, R.K.O. Metallothioneins and Related Chelators; Metal Ions in Life Sciences; RSC Publishing: Cambridge, UK, 2009; Volume 5. [Google Scholar]
- Klaassen, C.D.; Choudhuri, S.; McKim, J.M.; Lehman-McKeeman, L.D.; Kershaw, W.C. In vitro and in vivo studies on the degradation of metallothionein. Environ. Health Perspect. 1994, 102, 141–146. [Google Scholar] [PubMed]
- Kellen, E.; Zeegers, M.P.; Hond, E.D.; Buntinx, F. Blood cadmium may be associated with bladder carcinogenesis: The Belgian case-control study on bladder cancer. Cancer Detect. Prev. 2007, 31, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Wolf, C.; Strenziok, R.; Kyriakopoulos, A. Elevated metallothionein-bound cadmium concentrations in urine from bladder carcinoma patients, investigated by size exclusion chromatography-inductively coupled plasma mass spectrometry. Anal. Chim. Acta 2009, 631, 218–222. [Google Scholar] [CrossRef]
- Feki-Tounsi, M.; Olmedo, P.; Gil, F.; Khlifi, R.; Mhiri, M.-N.; Rebai, A.; Hamza-Chaffai, A. Cadmium in blood of Tunisian men and risk of bladder cancer: Interactions with arsenic exposure and smoking. Environ. Sci. Pollut. Res. Int. 2013, 20, 7204–7213. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Liu, C.-S.; Liu, H.-J.; Huang, C.-P.; Huang, C.-Y.; Hsu, H.-T.; Liou, S.-H.; Chung, C.-J. Association between levels of urinary heavy metals and increased risk of urothelial carcinoma. Int. J. Urol. 2016, 23, 233–239. [Google Scholar] [CrossRef]
- Achanzar, W.E.; Diwan, B.A.; Liu, J.; Quader, S.T.; Webber, M.M.; Waalkes, M.P. Cadmium-induced Malignant Transformation of Human Prostate Epithelial Cells. Cancer Res. 2001, 61, 455–458. [Google Scholar]
- Takiguchi, M.; Achanzar, W.E.; Qu, W.; Li, G.; Waalkes, M.P. Effects of cadmium on DNA-(Cytosine-5) methyltransferase activity and DNA methylation status during cadmium-induced cellular transformation. Exp. Cell Res. 2003, 286, 355–365. [Google Scholar] [CrossRef]
- Lei, Y.-X.; Wei, L.; Wang, M.; Wu, G.-R.; Li, M. Malignant transformation and abnormal expression of eukaryotic initiation factor in bronchial epithelial cells induced by cadmium chloride. Biomed. Environ. Sci. 2008, 21, 332–338. [Google Scholar] [CrossRef]
- Sens, D.A.; Park, S.; Gurel, V.; Sens, M.A.; Garrett, S.H.; Somji, S. Inorganic Cadmium- and Arsenite-Induced Malignant Transformation of Human Bladder Urothelial Cells. Toxicol. Sci. 2004, 79, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Kreft, M.E.; Hudoklin, S.; Jezernik, K.; Romih, R. Formation and maintenance of blood–urine barrier in urothelium. Protoplasma 2010, 246, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, J.; Gardner, I.; Swales, N. An Introduction to Drug Disposition: The Basic Principles of Absorption, Distribution, Metabolism, and Excretion. Toxicol. Pathol. 1995, 23, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.; Rak-Raszewska, A.; Pietilä, I.; Quaggin, S.E.; Vainio, S. Signaling during Kidney Development. Cells 2015, 4, 112–132. [Google Scholar] [CrossRef]
- Margoshes, M.; Vallee, B.L. A Cadmium Protein from Equine Kidney Cortex. J. Am. Chem. Soc. 1957, 79, 4813–4814. [Google Scholar] [CrossRef]
- Kägi, J.H.; Schäffer, A. Biochemistry of metallothionein. Biochemistry 1988, 27, 8509–8515. [Google Scholar] [CrossRef]
- Hunt, C.T.; Boulanger, Y.; Fesik, S.W.; Armitage, I.M. NMR analysis of the structure and metal sequestering properties of metallothioneins. Environ. Health Perspect. 1984, 54, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, M.; Bofill, R.; Palacios, Ò.; Atrian, S. State-of-the-art of metallothioneins at the beginning of the 21st century. Coord. Chem. Rev. 2012, 256, 46–62. [Google Scholar] [CrossRef]
- Dalton, T.; Fu, K.; Palmiter, R.D.; Andrews, G.K. Transgenic mice that overexpress metallothionein-I resist dietary zinc deficiency. J. Nutr. 1996, 126, 825–833. [Google Scholar] [CrossRef]
- Kelly, E.J.; Quaife, C.J.; Froelick, G.J.; Palmiter, R.D. Metallothionein I and II protect against zinc deficiency and zinc toxicity in mice. J. Nutr. 1996, 126, 1782–1790. [Google Scholar]
- Lee, D.K.; Geiser, J.; Dufner-Beattie, J.; Andrews, G.K. Pancreatic metallothionein-I may play a role in zinc homeostasis during maternal dietary zinc deficiency in mice. J. Nutr. 2003, 133, 45–50. [Google Scholar] [CrossRef]
- Penkowa, M.; Cáceres, M.; Borup, R.; Nielsen, F.C.; Poulsen, C.B.; Quintana, A.; Molinero, A.; Carrasco, J.; Florit, S.; Giralt, M.; et al. Novel roles for metallothionein-I + II (MT-I + II) in defense responses, neurogenesis, and tissue restoration after traumatic brain injury: Insights from global gene expression profiling in wild-type and MT-I + II knockout mice. J. Neurosci. Res. 2006, 84, 1452–1474. [Google Scholar] [CrossRef] [PubMed]
- Lazo, J.S.; Kondo, Y.; Dellapiazza, D.; Michalska, A.E.; Choo, K.H.; Pitt, B.R. Enhanced sensitivity to oxidative stress in cultured embryonic cells from transgenic mice deficient in metallothionein I and II genes. J. Biol. Chem. 1995, 270, 5506–5510. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Haslinger, A.; Holtgreve, H.; Richards, R.I.; Krauter, P.; Westphal, H.M.; Beato, M. Characterization of DNA sequences through which cadmium and glucocorticoid hormones induce human metallothionein-IIA gene. Nature 1984, 308, 513–519. [Google Scholar] [CrossRef]
- Schulkens, I.A.; Castricum, K.C.M.; Weijers, E.M.; Koolwijk, P.; Griffioen, A.W.; Thijssen, V.L. Expression, regulation and function of human metallothioneins in endothelial cells. J. Vasc. Res. 2014, 51, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Yap, X.; Tan, H.-Y.; Huang, J.; Lai, Y.; Yip, G.W.-C.; Tan, P.-H.; Bay, B.-H. Over-expression of metallothionein predicts chemoresistance in breast cancer. J. Pathol. 2009, 217, 563–570. [Google Scholar] [CrossRef]
- Liu, Z.-M.; van Hasselt, C.A.; Song, F.-Z.; Vlantis, A.C.; Cherian, M.G.; Koropatnick, J.; Chen, G.G. Expression of functional metallothionein isoforms in papillary thyroid cancer. Mol. Cell. Endocrinol. 2009, 302, 92–98. [Google Scholar] [CrossRef]
- Ioachim, E.E.; Charchanti, A.V.; Stavropoulos, N.E.; Athanassiou, E.D.; Michael, M.C.; Agnantis, N.J. Localization of metallothionein in urothelial carcinoma of the human urinary bladder: An immunohistochemical study including correlation with HLA-DR antigen, p53, and proliferation indices. Anticancer Res. 2001, 21, 1757–1761. [Google Scholar] [PubMed]
- Jin, R.; Chow, V.T.-K.; Tan, P.-H.; Dheen, S.T.; Duan, W.; Bay, B.-H. Metallothionein 2A expression is associated with cell proliferation in breast cancer. Carcinogenesis 2002, 23, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Varshney, U.; Jahroudi, N.; Foster, R.; Gedamu, L. Structure, organization, and regulation of human metallothionein IF gene: Differential and cell-type-specific expression in response to heavy metals and glucocorticoids. Mol. Cell. Biol. 1986, 6, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Boonprasert, K.; Ruengweerayut, R.; Aunpad, R.; Satarug, S.; Na-Bangchang, K. Expression of metallothionein isoforms in peripheral blood leukocytes from Thai population residing in cadmium-contaminated areas. Environ. Toxicol. Pharmacol. 2012, 34, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Mita, M.; Satoh, M.; Shimada, A.; Okajima, M.; Azuma, S.; Suzuki, J.S.; Sakabe, K.; Hara, S.; Himeno, S. Metallothionein is a crucial protective factor against Helicobacter pylori-induced gastric erosive lesions in a mouse model. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G877–G884. [Google Scholar] [CrossRef]
- Kägi, J.H.; Kojima, Y. Chemistry and biochemistry of metallothionein. Exp. Suppl. 1987, 52, 25–61. [Google Scholar]
- Irvine, G.W.; Pinter, T.B.J.; Stillman, M.J. Defining the metal binding pathways of human metallothionein 1a: Balancing zinc availability and cadmium seclusion. Metallomics 2016, 8, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kambe, T. The Functions of Metallothionein and ZIP and ZnT Transporters: An Overview and Perspective. Int. J. Mol. Sci. 2016, 17, 336. [Google Scholar] [CrossRef]
- Miles, A.T.; Hawksworth, G.M.; Beattie, J.H.; Rodilla, V. Induction, Regulation, Degradation, and Biological Significance of Mammalian Metallothioneins. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 35–70. [Google Scholar] [CrossRef]
- Mehus, A.A.; Muhonen, W.W.; Garrett, S.H.; Somji, S.; Sens, D.A.; Shabb, J.B. Quantitation of Human Metallothionein Isoforms: A Family of Small, Highly Conserved, Cysteine-rich Proteins. Mol. Cell. Proteom. 2014, 13, 1020–1033. [Google Scholar] [CrossRef]
- Conway, D.E.; Lee, S.; Eskin, S.G.; Shah, A.K.; Jo, H.; McIntire, L.V. Endothelial metallothionein expression and intracellular free zinc levels are regulated by shear stress. Am. J. Physiol. Cell. Physiol. 2010, 299, C1461–C1467. [Google Scholar] [CrossRef]
- Jahroudi, N.; Foster, R.; Price-Haughey, J.; Beitel, G.; Gedamu, L. Cell-type specific and differential regulation of the human metallothionein genes. Correlation with DNA methylation and chromatin structure. J. Biol. Chem. 1990, 265, 6506–6511. [Google Scholar]
- Miura, N.; Koizumi, S. Heavy metal responses of the human metallothionein isoform genes. J. Pharm. Soc. Jpn. 2007, 127, 665–673. [Google Scholar] [CrossRef]
- Richards, R.I.; Heguy, A.; Karin, M. Structural and functional analysis of the human metallothionein-IA gene: Differential induction by metal ions and glucocorticoids. Cell 1984, 37, 263–272. [Google Scholar] [CrossRef]
- Selvaraj, A.; Balamurugan, K.; Yepiskoposyan, H.; Zhou, H.; Egli, D.; Georgiev, O.; Thiele, D.J.; Schaffner, W. Metal-responsive transcription factor (MTF-1) handles both extremes, copper load and copper starvation, by activating different genes. Genes Dev. 2005, 19, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Sims, H.I.; Chirn, G.-W.; Marr, M.T. Single nucleotide in the MTF-1 binding site can determine metal-specific transcription activation. PNAS 2012, 109, 16516–16521. [Google Scholar] [CrossRef] [PubMed]
- Sadhu, C.; Gedamu, L. Regulation of human metallothionein (MT) genes. Differential expression of MTI-F, MTI-G, and MTII-A genes in the hepatoblastoma cell line (HepG2). J. Biol. Chem. 1988, 263, 2679–2684. [Google Scholar]
- Lehman-McKeeman, L.D.; Andrews, G.K.; Klaassen, C.D. Mechanisms of regulation of rat hepatic metallothionein-I and metallothionein-II levels following administration of zinc. Toxicol. Appl. Pharmacol. 1988, 92, 1–9. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Lehman-McKeeman, L.D. Regulation of the isoforms of metallothionein. Biol. Trace Elem. Res. 1989, 21, 119–129. [Google Scholar] [CrossRef]
- Kim, J.-H.; Wang, S.-Y.; Kim, I.-C.; Ki, J.-S.; Raisuddin, S.; Lee, J.-S.; Han, K.-N. Cloning of a river pufferfish (Takifugu obscurus) metallothionein cDNA and study of its induction profile in cadmium-exposed fish. Chemosphere 2008, 71, 1251–1259. [Google Scholar] [CrossRef]
- Jasani, B.; Schmid, K.W. Significance of metallothionein overexpression in human tumours. Histopathology 1997, 31, 131–136. [Google Scholar] [CrossRef]
- Chaney, R.L. Cadmium and Zinc. In Trace Elements in Soils; Wiley: New York, NY, USA, 2010; pp. 409–440. [Google Scholar]
- Malgieri, G.; Zaccaro, L.; Leone, M.; Bucci, E.; Esposito, S.; Baglivo, I.; Del Gatto, A.; Russo, L.; Scandurra, R.; Pedone, P.V.; et al. Zinc to cadmium replacement in the A. thaliana SUPERMAN Cys₂ His₂ zinc finger induces structural rearrangements of typical DNA base determinant positions. Biopolymers 2011, 95, 801–810. [Google Scholar] [PubMed]
- Tang, L.; Qiu, R.; Tang, Y.; Wang, S. Cadmium–zinc exchange and their binary relationship in the structure of Zn-related proteins: A mini review. Metallomics 2014, 6, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Eide, D.J. The SLC39 family of zinc transporters. Mol. Asp. Med. 2013, 34, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Southgate, J.; Hutton, K.A.; Thomas, D.F.; Trejdosiewicz, L.K. Normal human urothelial cells in vitro: Proliferation and induction of stratification. Lab. Investig. 1994, 71, 583–594. [Google Scholar] [PubMed]
- Cross, W.R.; Eardley, I.; Leese, H.J.; Southgate, J. A biomimetic tissue from cultured normal human urothelial cells: Analysis of physiological function. Am. J. Physiol. Renal Physiol. 2005, 289, F459–F468. [Google Scholar] [CrossRef]
- Wang, Y.; Fang, J.; Leonard, S.S.; Rao, K.M.K. Cadmium inhibits the electron transfer chain and induces reactive oxygen species. Free Radic. Biol. Med. 2004, 36, 1434–1443. [Google Scholar] [CrossRef]
- Heyno, E.; Klose, C.; Krieger-Liszkay, A. Origin of cadmium-induced reactive oxygen species production: Mitochondrial electron transfer versus plasma membrane NADPH oxidase. New Phytol. 2008, 179, 687–699. [Google Scholar] [CrossRef]
- Durnam, D.M.; Palmiter, R.D. Induction of metallothionein-I mRNA in cultured cells by heavy metals and iodoacetate: Evidence for gratuitous inducers. Mol. Cell. Biol. 1984, 4, 484–491. [Google Scholar] [CrossRef]
- Alam, J.; Smith, A. Heme-hemopexin-mediated induction of metallothionein gene expression. J. Biol. Chem. 1992, 267, 16379–16384. [Google Scholar]
- Baird, S.K.; Kurz, T.; Brunk, U.T. Metallothionein protects against oxidative stress-induced lysosomal destabilization. Biochem. J. 2006, 394, 275–283. [Google Scholar] [CrossRef]
- Palmiter, R.D. Regulation of metallothionein genes by heavy metals appears to be mediated by a zinc-sensitive inhibitor that interacts with a constitutively active transcription factor, MTF-1. Proc. Natl. Acad. Sci. USA 1994, 91, 1219–1223. [Google Scholar] [CrossRef] [PubMed]
- Kheradmand, F.; Nourmohammadi, I.; Modarressi, M.H.; Firoozrai, M.; Ahmadi Faghih, M.A. Differential Gene-Expression of Metallothionein 1M and 1G in Response to Zinc in Sertoli TM4 Cells. Iran Biomed. J. 2010, 14, 9–15. [Google Scholar] [PubMed]
- Chu, A.; Foster, M.; Ward, S.; Zaman, K.; Hancock, D.; Petocz, P.; Samman, S. Zinc-induced upregulation of metallothionein (MT)-2A is predicted by gene expression of zinc transporters in healthy adults. Genes Nutr. 2015, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, L.; Bai, Q.; Zhu, X.; Zhang, J.; Wei, Q.; Li, D.; Gao, C.; Li, J.; Zhang, Z.; et al. Heavy metal-induced metallothionein expression is regulated by specific protein phosphatase 2A complexes. J. Biol. Chem. 2014, 289, 22413–22426. [Google Scholar] [CrossRef] [PubMed]
- Heuchel, R.; Radtke, F.; Georgiev, O.; Stark, G.; Aguet, M.; Schaffner, W. The transcription factor MTF-1 is essential for basal and heavy metal-induced metallothionein gene expression. EMBO J. 1994, 13, 2870–2875. [Google Scholar] [CrossRef]
- Zhou, X.D.; Sens, D.A.; Sens, M.A.; Namburi, V.B.R.K.; Singh, R.K.; Garrett, S.H.; Somji, S. Metallothionein-1 and -2 Expression in Cadmium- or Arsenic-Derived Human Malignant Urothelial Cells and Tumor Heterotransplants and as a Prognostic Indicator in Human Bladder Cancer. Toxicol. Sci. 2006, 91, 467–475. [Google Scholar] [CrossRef]
- Irvine, G.W.; Stillman, M.J. Topographical analysis of As-induced folding of α-MT1a. Biochem. Biophys. Res. Commun. 2013, 441, 208–213. [Google Scholar] [CrossRef]
- McMahon, R.J.; Cousins, R.J. Regulation of the zinc transporter ZnT-1 by dietary zinc. Proc. Natl. Acad. Sci. USA 1998, 95, 4841–4846. [Google Scholar] [CrossRef]
- Holloway, A.F.; Stennard, F.A.; West, A.K. Human metallothionein gene MT1L mRNA is present in several human tissues but is unlikely to produce a metallothionein protein. FEBS Lett. 1997, 404, 41–44. [Google Scholar] [CrossRef]
- Hahn, Y.; Lee, B. Human-specific nonsense mutations identified by genome sequence comparisons. Hum. Genet. 2006, 119, 169–178. [Google Scholar] [CrossRef]
- Ryter, S.W.; Choi, A.M.K. Heme oxygenase-1: Redox regulation of a stress protein in lung and cell culture models. Antioxid. Redox Signal. 2005, 7, 80–91. [Google Scholar] [CrossRef]
- Sarma, S.N.; Kim, Y.-J.; Song, M.; Ryu, J.-C. Induction of apoptosis in human leukemia cells through the production of reactive oxygen species and activation of HMOX1 and Noxa by benzene, toluene, and o-xylene. Toxicology 2011, 280, 109–117. [Google Scholar] [CrossRef]
- Faurskov, B.; Bjerregaard, H.F. Effect of cadmium on active ion transport and cytotoxicity in cultured renal epithelial cells (A6). Toxicol. In Vitro 1997, 11, 717–722. [Google Scholar] [CrossRef]
- Cao, X.; Lin, H.; Muskhelishvili, L.; Latendresse, J.; Richter, P.; Heflich, R.H. Tight junction disruption by cadmium in an in vitro human airway tissue model. Respir. Res. 2015, 16, 30. [Google Scholar] [CrossRef]
- Thirumoorthy, N.; Manisenthil Kumar, K.-T.; Shyam Sundar, A.; Panayappan, L.; Chatterjee, M. Metallothionein: An overview. World J. Gastroenterol. 2007, 13, 993–996. [Google Scholar] [CrossRef]
- Raudenska, M.; Gumulec, J.; Podlaha, O.; Sztalmachova, M.; Babula, P.; Eckschlager, T.; Adam, V.; Kizek, R.; Masarik, M. Metallothionein polymorphisms in pathological processes. Metallomics 2013, 6, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Findley, S.D.; Whitmore, T.E.; Durnam, D.M. MT-III, a brain-specific member of the metallothionein gene family. Proc. Natl. Acad. Sci. USA 1992, 89, 6333–6337. [Google Scholar] [CrossRef] [PubMed]
- Sens, M.A.; Somji, S.; Lamm, D.L.; Garrett, S.H.; Slovinsky, F.; Todd, J.H.; Sens, D.A. Metallothionein isoform 3 as a potential biomarker for human bladder cancer. Environ. Health Perspect. 2000, 108, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.D.; Sens, M.A.; Garrett, S.H.; Somji, S.; Park, S.; Gurel, V.; Sens, D.A. Enhanced expression of metallothionein isoform 3 protein in tumor heterotransplants derived from As+3- and Cd+2-transformed human urothelial cells. Toxicol. Sci. 2006, 93, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Somji, S.; Garrett, S.H.; Toni, C.; Zhou, X.D.; Zheng, Y.; Ajjimaporn, A.; Sens, M.A.; Sens, D.A. Differences in the epigenetic regulation of MT-3 gene expression between parental and Cd+2 or As+3 transformed human urothelial cells. Cancer Cell Int. 2011, 11, 2. [Google Scholar] [CrossRef]
- Brazão-Silva, M.T.; Rodriguez, M.F.S.; Eisenberg, A.L.A.; Dias, F.L.; de Castro, L.M.; Nunes, F.D.; Faria, P.R.; Cardoso, S.V.; Loyola, A.M.; de Sousa, S.C.O.M. Metallothionein gene expression is altered in oral cancer and may predict metastasis and patient outcomes. Histopathology 2015, 67, 358–367. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer Beryllium, cadmium, mercury, and exposures in the glass manufacturing industry. In IARC Monographs on the Evaluation of the Carcinogenic Risks to Humans; IARC: Lyon, France, 1993; Volume 58.
- Salnikow, K.; An, W.G.; Melillo, G.; Blagosklonny, M.V.; Costa, M. Nickel-induced transformation shifts the balance between HIF-1 and p53 transcription factors. Carcinogenesis 1999, 20, 1819–1823. [Google Scholar] [CrossRef]
- Günther, V.; Lindert, U.; Schaffner, W. The taste of heavy metals: Gene regulation by MTF-1. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2012, 1823, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Itoh, N.; Andrews, G.K. Mechanisms of Heavy Metal Sensing by Metal Response Element-binding Transcription Factor-1. J. Health Sci. 2009, 55, 484–494. [Google Scholar] [CrossRef]
- Otsuka, F. Molecular Mechanism of the Metallothionein Gene Expression Mediated by Metal-Responsive Transcription Factor 1. J. Health Sci. 2001, 47, 513–519. [Google Scholar] [CrossRef]
- Langmade, S.J.; Ravindra, R.; Daniels, P.J.; Andrews, G.K. The transcription factor MTF-1 mediates metal regulation of the mouse ZnT1 gene. J. Biol. Chem. 2000, 275, 34803–34809. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Lichten, L.A.; Ryu, M.-S.; Liuzzi, J.P.; Wang, F.; Cousins, R.J. STAT5-glucocorticoid receptor interaction and MTF-1 regulate the expression of ZnT2 (Slc30a2) in pancreatic acinar cells. Proc. Natl. Acad. Sci. USA 2010, 107, 2818–2823. [Google Scholar] [CrossRef]
- Bafaro, E.; Liu, Y.; Xu, Y.; Dempski, R.E. The emerging role of zinc transporters in cellular homeostasis and cancer. Signal Transduct. Target. Ther. 2017, 2, 17029. [Google Scholar] [CrossRef]
- Urani, C.; Melchioretto, P.; Gribaldo, L. Regulation of metallothioneins and ZnT-1 transporter expression in human hepatoma cells HepG2 exposed to zinc and cadmium. Toxicol. In Vitro 2010, 24, 370–374. [Google Scholar] [CrossRef]
- Urani, C.; Melchioretto, P.; Bruschi, M.; Fabbri, M.; Sacco, M.G.; Gribaldo, L. Impact of Cadmium on Intracellular Zinc Levels in HepG2 Cells: Quantitative Evaluations and Molecular Effects, Impact of Cadmium on Intracellular Zinc Levels in HepG2 Cells: Quantitative Evaluations and Molecular Effects. BioMed Res. Int. BioMed Res. Int. 2015, 2015, e949514. [Google Scholar]
- Pedersen, M.Ø.; Larsen, A.; Stoltenberg, M.; Penkowa, M. The role of metallothionein in oncogenesis and cancer prognosis. Prog. Histochem. Cytochem. 2009, 44, 29–64. [Google Scholar] [CrossRef] [PubMed]
- Hart, B.A.; Potts, R.J.; Watkin, R.D. Cadmium adaptation in the lung—A double-edged sword? Toxicology 2001, 160, 65–70. [Google Scholar] [CrossRef]
- Eneman, J.D.; Potts, R.J.; Osier, M.; Shukla, G.S.; Lee, C.H.; Chiu, J.-F.; Hart, B.A. Suppressed oxidant-induced apoptosis in cadmium adapted alveolar epithelial cells and its potential involvement in cadmium carcinogenesis. Toxicology 2000, 147, 215–228. [Google Scholar] [CrossRef]
- Potts, R.J.; Bespalov, I.A.; Wallace, S.S.; Melamede, R.J.; Hart, B.A. Inhibition of oxidative DNA repair in cadmium-adapted alveolar epithelial cells and the potential involvement of metallothionein. Toxicology 2001, 161, 25–38. [Google Scholar] [CrossRef]
- Satoh, M.; Kloth, D.M.; Kadhim, S.A.; Chin, J.L.; Naganuma, A.; Imura, N.; Cherian, M.G. Modulation of Both Cisplatin Nephrotoxicity and Drug Resistance in Murine Bladder Tumor by Controlling Metallothionein Synthesis. Cancer Res. 1993, 53, 1829–1832. [Google Scholar]
- Wülfing, C.; van Ahlen, H.; Eltze, E.; Piechota, H.; Hertle, L.; Schmid, K.-W. Metallothionein in bladder cancer: Correlation of overexpression with poor outcome after chemotherapy. World J. Urol. 2007, 25, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, Y.; Smith, C.; Weisz, D.; van Huizen, I.; Xuan, J.; Moussa, M.; Stitt, L.; Hideki, S.; Cherian, M.G.; Izawa, J.I. Metallothionein expression as prognostic factor for transitional cell carcinoma of bladder. Urology 2006, 67, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Southgate, J.; Masters, J.R.W.; Trejdosiewicz, L.K. Culture of Human Urothelium. In Culture of Epithelial Cells; John Wiley & Sons, Inc.: New York, NY, USA, 2002; pp. 381–399. ISBN 978-0-471-22120-3. [Google Scholar]
- Rubenwolf, P.; Southgate, J. Permeability of differentiated human urothelium in vitro. Methods Mol. Biol. 2011, 763, 207–222. [Google Scholar]
- Fishwick, C.; Higgins, J.; Percival-Alwyn, L.; Hustler, A.; Pearson, J.; Bastkowski, S.; Moxon, S.; Swarbreck, D.; Greenman, C.D.; Southgate, J. Heterarchy of transcription factors driving basal and luminal cell phenotypes in human urothelium. Cell Death Differ. 2017, 24, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Research 2015, 4, 1521. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, H.; Bray, N.L.; Puente, S.; Melsted, P.; Pachter, L. Differential analysis of RNA-seq incorporating quantification uncertainty. Nat. Methods 2017, 14, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Lobban, E.D.; Smith, B.A.; Hall, G.D.; Harnden, P.; Roberts, P.; Selby, P.J.; Trejdosiewicz, L.K.; Southgate, J. Uroplakin gene expression by normal and neoplastic human urothelium. Am. J. Pathol. 1998, 153, 1957–1967. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward or Reverse | Sequence (5′–3′) | Product Size (bp) |
|---|---|---|---|
| GAPDH | Forward | CAAGGTCATCCATGACAACTTTG | 90 |
| GAPDH | Reverse | GGGCCATCCACAGTCTTCTG | 90 |
| HMOX1 | Forward | CCAGCAACAAAGTGCAAGATTC | 102 |
| HMOX1 | Reverse | GTGTAAGGACCCATCGGAGAAG | 102 |
| MT-1A | Forward | CTCGAAATGGACCCCAACT | 219 |
| MT-1A | Reverse | ATATCTTCGAGCAGGGCTGTC | 219 |
| MT-1B | Forward | GGAACTCCAGGCTTGTCTTGG | 77 |
| MT-1B | Reverse | TTGCAGGAGGTACATTTG | 77 |
| MT-1E | Forward | TGCGCCGGCTCCTGCAAGTC | 118 |
| MT-1E | Reverse | ATGCCCCTTTGCAGACGCAGC | 118 |
| MT-1F | Forward | CCTGCACCTGCGCTGGTTCC | 110 |
| MT-1F | Reverse | ACAGCCCTGGGCACACTTGC | 110 |
| MT-1G | Forward | CTTCTCGCTTGGGAACTCTA | 309 |
| MT-1G | Reverse | AGGGGTCAAGATTGTAGCAAA | 309 |
| MT-1H | Forward | CCTCTTCTCTTCTCGCTTGG | 317 |
| MT-1H | Reverse | GCAAATGAGTCGGAGTTGTAG | 317 |
| MT-1M | Forward | CTAGCAGTCGCTCCATTTATCG | 180 |
| MT-1M | Reverse | CAGCTGCAGTTCTCCAACGT | 180 |
| MT-1X | Forward | GGACCCAACTGCTCCTGCTC | 151 |
| MT-1X | Reverse | TTTGCAGATGCAGCCCTGGGC | 151 |
| MT-2A | Forward | CCGACTCTAGCCGCCTCTT | 259 |
| MT-2A | Reverse | GTGGAAGTCGCGTTCTTTACA | 259 |
| MT-3 | Forward | AGTGCGAGGGATGCAAATG | 98 |
| MT-3 | Reverse | GCCTTTGCACACACAGTCCTT | 98 |
| SLC30A1 | Forward | GCATCAGTTTATGAGGCTGGTCCT | 352 |
| SLC30A1 | Reverse | CAGGCTGAATGGTAGTAGCGTGAA | 352 |
| Antigen | Clone | Host | Supplier | Dilution | Molecular Weight (kDa) |
|---|---|---|---|---|---|
| Beta-actin | A5441 | Mouse | Sigma Aldrich | 1:10 000 (WB) | 42 |
| MT-1A | B01P | Mouse | Abnova | 1:750 (WB) | 6 |
| MT-1M | 17281-AP | Rabbit | ProteinTech | 1 µg/mL (WB) | 6 |
| Antigen | Conjugate | Host | Supplier | Application |
|---|---|---|---|---|
| Anti-mouse IgG | Alexa 680 | Goat | Life Technologies | WB |
| Anti-rabbit IgG | Alexa 800 | Goat | Life Technologies | WB |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McNeill, R.V.; Mason, A.S.; Hodson, M.E.; Catto, J.W.F.; Southgate, J. Specificity of the Metallothionein-1 Response by Cadmium-Exposed Normal Human Urothelial Cells. Int. J. Mol. Sci. 2019, 20, 1344. https://doi.org/10.3390/ijms20061344
McNeill RV, Mason AS, Hodson ME, Catto JWF, Southgate J. Specificity of the Metallothionein-1 Response by Cadmium-Exposed Normal Human Urothelial Cells. International Journal of Molecular Sciences. 2019; 20(6):1344. https://doi.org/10.3390/ijms20061344
Chicago/Turabian StyleMcNeill, Rhiannon V., Andrew S. Mason, Mark E. Hodson, James W.F. Catto, and Jennifer Southgate. 2019. "Specificity of the Metallothionein-1 Response by Cadmium-Exposed Normal Human Urothelial Cells" International Journal of Molecular Sciences 20, no. 6: 1344. https://doi.org/10.3390/ijms20061344
APA StyleMcNeill, R. V., Mason, A. S., Hodson, M. E., Catto, J. W. F., & Southgate, J. (2019). Specificity of the Metallothionein-1 Response by Cadmium-Exposed Normal Human Urothelial Cells. International Journal of Molecular Sciences, 20(6), 1344. https://doi.org/10.3390/ijms20061344