Advances in Antigenic Peptide-Based Vaccine and Neutralizing Antibodies against Viruses Causing Hand, Foot, and Mouth Disease


Abstract
1. Introduction
2. Etiological Agents of HFMD
3. Neutralizing Epitope Mapping
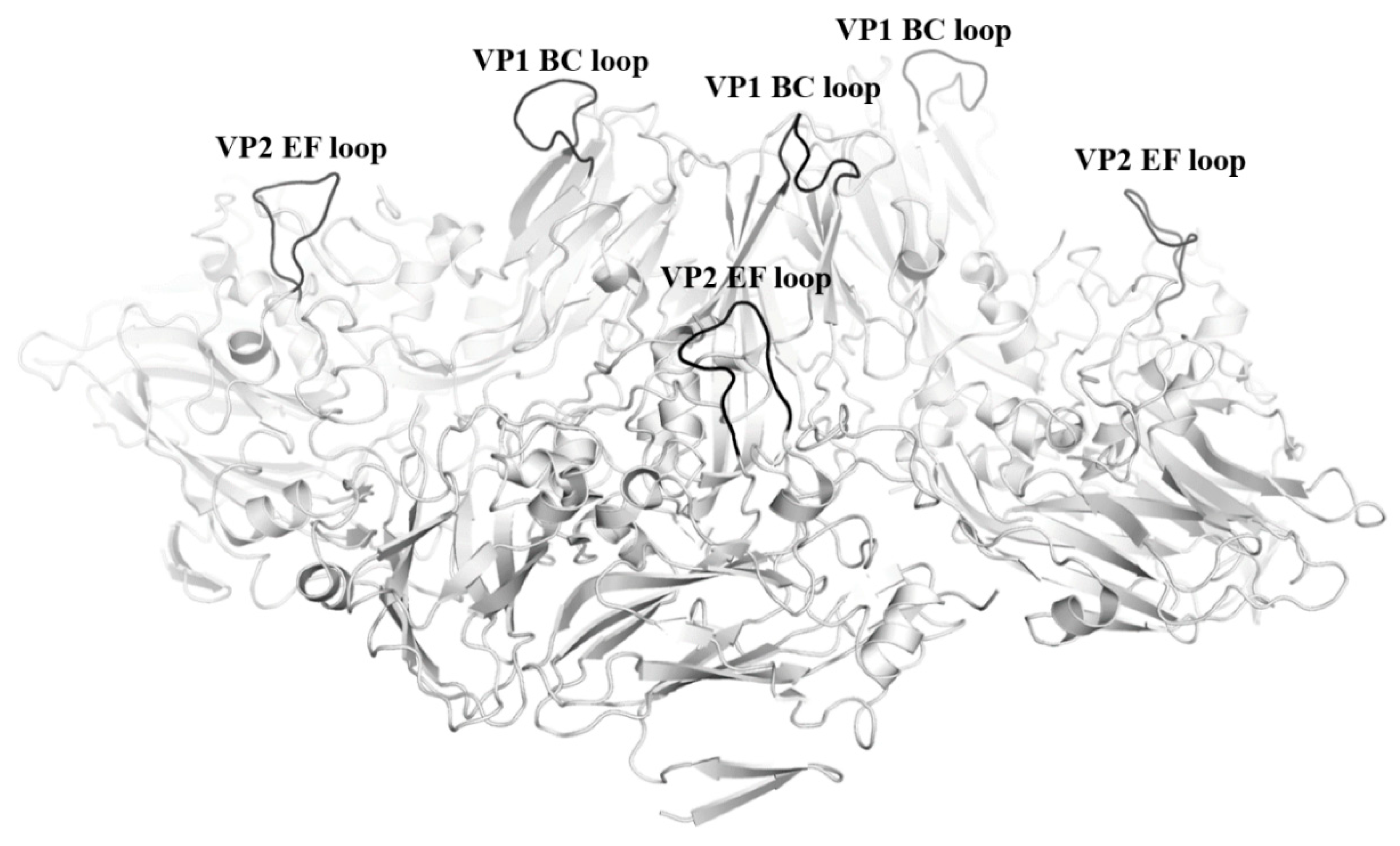
3.1. EV-A71 Neutralizing Epitope Mapping
3.2. CV-A16 Epitope Mapping
3.3. CV-A6 and CV-A10 Epitope Mapping
4. Antigenic Peptide-Based Vaccine
4.1. Peptide-Based Vaccines Against EV-A71
4.2. EV-A71/CV-A16 Peptide-Based Bivalent Vaccines
4.3. Peptide Insertion Sites in EV-A71 VLP
4.4. Tetravalent Peptide-Based HFMD Vaccine
5. Neutralizing Antibodies Against HFMD Etiological Agents
5.1. EV-A71 Neutralizing Antibodies
5.2. CV-A16, CV-A6 and CV-A10 Neutralizing Antibodies
5.3. Neutralizing Antibodies as Potential Prophylactic or Therapeutic Agents against Enteroviruses Causing HFMD
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADE | Antibody dependent enhancement |
| BEI | Binary ethyleneimine |
| CDR | Complementary Determining Region |
| CNS | Central Nervous System |
| CV-A16 | Coxsackievirus 16 |
| EMEA | European Medicine Agency |
| EV-A71 | Enterovirus A 71 |
| Fab | Fragment antigen-binding |
| FcR | Fc Receptor |
| FDA | Food and Drug Administration |
| HFMD | Hand, Foot and Mouth Disease |
| mTLNE | Multiple tandem linear neutralizing epitope |
| RD | Rhabdomyosarcoma |
| r.m.s.d | Root Mean Square Deviation |
| RNA | Ribonucleic Acid |
| Trx | Thioredoxin |
| UTR | Untranslated Region |
| VCV | Varicella-zoster virus |
| VLP | Virus-Like-Particle |
References
- Solomon, T.; Lewthwaite, P.; Perera, D.; Cardosa, M.J.; McMinn, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect. Dis. 2010, 10, 778–790. [Google Scholar] [CrossRef]
- Mao, Q.; Wang, Y.; Yao, X.; Bian, L.; Wu, X.; Xu, M.; Liang, Z. Coxsackievirus A16: Epidemiology, diagnosis, and vaccine. Hum. Vaccin. Immunother. 2014, 10, 360–367. [Google Scholar] [CrossRef]
- Chen, M.; He, S.; Yan, Q.; Xu, X.; Wu, W.; Ge, S.; Zhang, S.; Chen, M.; Xia, N. Severe hand, foot and mouth disease associated with coxsackievirus A10 infections in Xiamen, China in 2015. J. Clin. Virol. 2017, 93, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Anh, N.T.; Nhu, L.N.T.; Van, H.M.T.; Hong, N.T.T.; Thanh, T.T.; Hang, V.T.T.; Ny, N.T.H.; Nguyet, L.A.; Phuong, T.T.L.; Nhan, L.N.T.; et al. Emerging coxsackievirus A6 causing hand, foot and mouth disease, Vietnam. Emerg. Infect. Dis. 2018, 24, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Yee, P.T.I.; Poh, C.L. T cell immunity to enterovirus 71 infection in humans and implications for vaccine development. Int. J. Med. Sci. 2018, 15, 1143–1152. [Google Scholar] [CrossRef]
- Chen, L.; Yang, H.; Wang, C.; Yao, X.J.; Zhang, H.L.; Zhang, R.L.; He, Y.Q. Genomic characteristics of coxsackievirus A8 strains associated with hand, foot, and mouth disease and herpangina. Arch. Virol. 2016, 161, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wu, S.; Xiong, Y.; Li, T.; Wen, Z.; Yan, M.; Qin, K.; Liu, Y.; Wu, J. Co-circulation and genomic recombination of coxsackievirus A16 and enterovirus 71 during a large outbreak of hand, foot, and mouth disease in Central China. PLoS ONE 2014, 9, e96051. [Google Scholar] [CrossRef]
- Blomqvist, S.; Klemola, P.; Kaijalainen, S.; Paananen, A.; Simonen, M.L.; Vuorinen, T.; Roivainen, M. Co-circulation of coxsackieviruses A6 and A10 in hand, foot and mouth disease outbreak in Finland. J. Clin. Virol. 2010, 48, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Wang, Y.; Bian, L.; Xu, M.; Liang, Z. EV-A71 vaccine licensure: A first step for multivalent enterovirus vaccine to control HFMD and other severe diseases. Emerg. Microbes Infect. 2016, 5, e75. [Google Scholar] [CrossRef]
- Zhu, F.; Xu, W.; Xia, J.; Liang, Z.; Liu, Y.; Zhang, X.; Tan, X.; Wang, L.; Mao, Q.; Wu, J.; et al. Efficacy, Safety, and Immunogenicity of an Enterovirus 71 Vaccine in China. N. Engl. J. Med. 2014, 370, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.C.; Meng, F.Y.; Li, J.X.; Li, X.L.; Mao, Q.Y.; Tao, H.; Zhang, Y.T.; Yao, X.; Chu, K.; Chen, Q.H.; et al. Efficacy, safety, and immunology of an inactivated alum-adjuvant enterovirus 71 vaccine in children in China: A multicentre, randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2013, 381, 2024–2032. [Google Scholar] [CrossRef]
- Liu, L.; Mo, Z.; Liang, Z.; Zhang, Y.; Li, R.; Ong, K.C.; Wong, K.T.; Yang, E.; Che, Y.; Wang, J.; et al. Immunity and clinical efficacy of an inactivated enterovirus 71 vaccine in healthy Chinese children: A report of further observations. BMC Med. 2015, 13, 226. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.A.; Pallansch, M.A. Complete nucleotide sequence of enterovirus 71 is distinct from poliovirus. Virus Res. 1995, 39, 195–205. [Google Scholar] [CrossRef]
- Plevka, P.; Perera, R.; Cardosa, J.; Kuhn, R.J.; Rossmann, M.G. Crystal structure of human enterovirus 71. Science 2012, 336, 1274. [Google Scholar] [CrossRef] [PubMed]
- Shingler, K.L.; Yoder, J.L.; Carnegie, M.S.; Ashley, R.E.; Makhov, A.M.; Conway, J.F.; Hafenstein, S. The enterovirus 71 A-particle forms a gateway to allow genome release: A cryoEM study of picornavirus uncoating. PLoS Pathog. 2013, 9, e1003240. [Google Scholar] [CrossRef]
- Belnap, D.M.; Filman, D.J.; Trus, B.L.; Cheng, N.; Booy, F.P.; Conway, J.F.; Curry, S.; Hiremath, C.N.; Tsang, S.K.; Steven, A.C.; et al. Molecular tectonic model of virus structural transitions: The putative cell entry states of poliovirus. J. Virol. 2000, 74, 1342–1354. [Google Scholar] [CrossRef]
- Rossmann, M.G.; Arnold, E.; Erickson, J.W.; Frankenberger, E.A.; Griffith, J.P.; Hecht, H.J.; Johnson, J.E.; Kamer, G.; Luo, M.; Mosser, A.G.; et al. Structure of a human common cold virus and functional relationship to other picornaviruses. Nature 1985, 317, 145–153. [Google Scholar] [CrossRef]
- Tuthill, T.J.; Bubeck, D.; Rowlands, D.J.; Hogle, J.M. Characterization of early steps in the poliovirus infection process: Receptor-decorated liposomes induce conversion of the virus to membrane-anchored entry-intermediate particles. J. Virol. 2006, 80, 172–180. [Google Scholar] [CrossRef]
- Wang, X.; Peng, W.; Ren, J.; Hu, Z.; Xu, J.; Lou, Z.; Li, X.; Yin, W.; Shen, X.; Porta, C.; et al. A sensor-adaptor mechanism for enterovirus uncoating from structures of EV71. Nat. Struct. Mol. Biol. 2012, 19, 424–429. [Google Scholar] [CrossRef]
- Zhu, L.; Xu, K.; Wang, N.; Cao, L.; Wu, J.; Gao, Q.; Fry, E.E.; Stuart, D.I.; Rao, Z.; Wang, J.; et al. Neutralization mechanisms of two highly potent antibodies against human enterovirus 71. mBio 2018, 9, e01013–e01018. [Google Scholar] [CrossRef]
- Wu, C.N.; Lin, Y.C.; Fann, C.; Liao, N.S.; Shih, S.R.; Ho, M.S. Protection against lethal enterovirus 71 infection in newborn mice by passive immunization with subunit VP1 vaccines and inactivated virus. Vaccine 2001, 20, 895–904. [Google Scholar] [CrossRef]
- Foo, D.G.; Alonso, S.; Phoon, M.C.; Ramachandran, N.P.; Chow, V.T.; Poh, C.L. Identification of neutralizing linear epitopes from the VP1 capsid protein of Enterovirus 71 using synthetic peptides. Virus Res. 2007, 125, 61–68. [Google Scholar] [CrossRef]
- Liu, C.C.; Chou, A.H.; Lien, S.P.; Lin, H.Y.; Liu, S.J.; Chang, J.Y.; Guo, M.S.; Chow, Y.H.; Yang, W.S.; Chang, K.H.; et al. Identification and characterization of a cross-neutralization epitope of Enterovirus 71. Vaccine 2011, 29, 4362–4372. [Google Scholar] [CrossRef]
- Xu, L.; He, D.; Yang, L.; Li, Z.; Ye, X.; Yu, H.; Zhao, H.; Li, S.; Yuan, L.; Qian, H.; et al. A broadly cross-protective vaccine presenting the neighboring epitopes within the VP1 GH loop and VP2 EF loop of enterovirus 71. Sci. Rep. 2015, 5, 12973. [Google Scholar] [CrossRef]
- Aw-Yong, K.L.; Sam, I.C.; Koh, M.T.; Chan, Y.F. Immunodominant IgM and IgG epitopes recognized by antibodies induced in enterovirus A71-associated Hand, Foot and Mouth Disease patients. PLoS ONE 2016, 11, e0165659. [Google Scholar] [CrossRef]
- Gao, F.; Wang, Y.P.; Mao, Q.Y.; Yao, X.; Liu, S.; Li, F.X.; Zhu, F.C.; Yang, J.Y.; Liang, Z.L.; Lu, F.M.; et al. Enterovirus 71 viral capsid protein linear epitopes: Identification and characterization. Virol. J. 2012, 9, 26. [Google Scholar] [CrossRef]
- Kiener, T.K.; Jia, Q.; Lim, X.F.; He, F.; Meng, T.; Chow, V.T.; Kwang, J. Characterization and specificity of the linear epitope of the enterovirus 71 VP2 protein. Virol. J. 2012, 9, 55. [Google Scholar] [CrossRef]
- Xu, L.; He, D.; Li, Z.; Zheng, J.; Yang, L.; Yu, M.; Yu, H.; Chen, Y.; Que, Y.; Shih, J.W.; et al. Protection against lethal enterovirus 71 challenge in mice by a recombinant vaccine candidate containing a broadly cross-neutralizing epitope within the VP2 EF loop. Theranostics 2014, 4, 498–513. [Google Scholar] [CrossRef]
- Kiener, T.K.; Jia, Q.; Meng, T.; Chow, V.T.; Kwang, J. A novel universal neutralizing monoclonal antibody against enterovirus 71 that targets the highly conserved “knob” region of VP3 protein. PLoS Negl. Trop. Dis. 2014, 8, e2895. [Google Scholar] [CrossRef]
- Jia, Q.; Ng, Q.; Chin, W.; Meng, T.; Chow, V.T.K.; Wang, C.I.; Kwang, J.; He, F. Effective in vivo therapeutic IgG antibody against VP3 of enterovirus 71 with receptor-competing activity. Sci. Rep. 2017, 7, 46402. [Google Scholar] [CrossRef]
- Zhao, M.; Bai, Y.; Liu, W.; Xiao, X.; Huang, Y.; Cen, S.; Chan, P.K.; Sun, X.; Sheng, W.; Zeng, Y. Immunization of N terminus of enterovirus 71 VP4 elicits cross-protective antibody responses. BMC Microbiol. 2013, 13, 287. [Google Scholar] [CrossRef] [PubMed]
- Chong, P.; Guo, M.S.; Lin, F.H.; Hsiao, K.N.; Weng, S.Y.; Chou, A.H.; Wang, J.R.; Hsieh, S.Y.; Su, I.J.; Liu, C.C. Immunological and biochemical characterization of coxsackie virus A16 viral particles. PLoS ONE 2012, 7, e49973. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yan, K.; Feng, Y.; Huang, X.; Ku, Z.; Cai, Y.; Liu, F.; Shi, J.; Huang, Z. A virus-like particle vaccine for coxsackievirus A16 potently elicits neutralizing antibodies that protect mice against lethal challenge. Vaccine 2012, 30, 6642–6648. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Huang, X.; Liu, Q.; Huang, Z. Identification of conserved neutralizing linear epitopes within the VP1 protein of coxsackievirus A16. Vaccine 2013, 31, 2130–2136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, C.; Xiao, X.; Pang, L.; Shen, S.; Zhang, J.; Cen, S.; Yang, B.B.; Huang, Y.; Sheng, W.; et al. Phage Display-Derived Cross-Reactive Neutralizing Antibody against Enterovirus 71 and Coxsackievirus A16. Jpn. J. Infect. Dis. 2016, 69, 66–74. [Google Scholar] [CrossRef]
- Zhu, R.; Xu, L.; Zheng, Q.; Cui, Y.; Li, S.; He, M.; Yin, Z.; Liu, D.; Li, S.; Li, Z.; et al. Discovery and structural characterization of a therapeutic antibody against coxsackievirus A10. Sci. Adv. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Sun, Y.; Fan, J.; Zhu, B.; Cao, L.; Gao, Q.; Zhang, Y.; Liu, H.; Rao, Z.; Wang, X. Structures of coxsackievirus A10 unveil the molecular mechanisms of receptor binding and viral uncoating. Nat. Commun. 2018, 9, 4985. [Google Scholar] [CrossRef]
- Xu, L.; Zheng, Q.; Li, S.; He, M.; Wu, Y.; Li, Y.; Zhu, R.; Yu, H.; Hong, Q.; Jiang, J.; et al. Atomic structures of Coxsackievirus A6 and its complex with a neutralizing antibody. Nat. Commun. 2017, 8, 505. [Google Scholar] [CrossRef]
- Yang, L.; Mao, Q.; Li, S.; Gao, F.; Zhao, H.; Liu, Y.; Wan, J.; Ye, X.; Xia, N.; Cheng, T.; et al. A neonatal mouse model for the evaluation of antibodies and vaccines against coxsackievirus A6. Antiviral Res. 2016, 134, 50–57. [Google Scholar] [CrossRef]
- Li, W.; Joshi, M.D.; Singhania, S.; Ramsey, K.H.; Murthy, A.K. Peptide vaccine: Progress and challenges. Vaccines 2014, 2, 515–536. [Google Scholar] [CrossRef]
- Tirado, S.M.; Yoon, K.J. Antibody-dependent enhancement of virus infection and disease. Viral Immunol. 2003, 16, 69–86. [Google Scholar] [CrossRef]
- Han, J.F.; Cao, R.Y.; Deng, Y.Q.; Tian, X.; Jiang, T.; Qin, E.D.; Qin, C.F. Antibody dependent enhancement infection of enterovirus 71 in vitro and in vivo. Virol. J. 2011, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.M.; Chen, I.C.; Su, L.Y.; Huang, K.J.; Lei, H.Y.; Liu, C.C. Enterovirus 71 infection of monocytes with antibody-dependent enhancement. Clin. Vaccine Immunol. 2010, 17, 1517–1523. [Google Scholar] [CrossRef]
- Chen, I.C.; Wang, S.M.; Yu, C.K.; Liu, C.C. Subneutralizing antibodies to enterovirus 71 induce antibody-dependent enhancement of infection in newborn mice. Med. Microbiol. Immunol. 2013, 202, 259–265. [Google Scholar] [CrossRef]
- Cao, R.Y.; Dong, D.Y.; Liu, R.J.; Han, J.F.; Wang, G.C.; Zhao, H.; Li, X.F.; Deng, Y.Q.; Zhu, S.Y.; Wang, X.Y.; et al. Human IgG subclasses against enterovirus Type 71: Neutralization versus antibody dependent enhancement of infection. PLoS ONE 2013, 8, e64024. [Google Scholar] [CrossRef] [PubMed]
- Foo, D.G.; Alonso, S.; Chow, V.T.; Poh, C.L. Passive protection against lethal enterovirus 71 infection in newborn mice by neutralizing antibodies elicited by a synthetic peptide. Microbes Infect. 2007, 9, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Su, X.; Li, X.; Li, H.; Li, T.; Zhou, Z.; Zhong, T.; Zhou, R. Protection against enterovirus 71 with neutralizing epitope incorporation within adenovirus type 3 hexon. PLoS ONE 2012, 7, e41381. [Google Scholar] [CrossRef]
- Li, Y.X.; Zhao, H.; Cao, R.Y.; Deng, Y.Q.; Han, J.F.; Zhu, S.Y.; Ma, J.; Liu, L.; Qin, E.D.; Qin, C.F. Recombinant tandem multi-linear neutralizing epitopes of human enterovirus 71 elicited protective immunity in mice. Virol. J. 2014, 11, 79. [Google Scholar] [CrossRef]
- Oberste, M.S.; Penaranda, S.; Maher, K.; Pallansch, M.A. Complete genome sequences of all members of the species Human enterovirus A. J. Gen. Virol. 2004, 85, 1597–1607. [Google Scholar] [CrossRef]
- Sun, S.; Jiang, L.; Liang, Z.; Mao, Q.; Su, W.; Zhang, H.; Li, X.; Jin, J.; Xu, L.; Zhao, D.; et al. Evaluation of monovalent and bivalent vaccines against lethal Enterovirus 71 and Coxsackievirus A16 infection in newborn mice. Hum. Vaccin. Immunother. 2014, 10, 2885–2895. [Google Scholar] [CrossRef]
- Fan, C.; Ye, X.; Ku, Z.; Kong, L.; Liu, Q.; Xu, C.; Cong, Y.; Huang, Z. Beta-propiolactone inactivation of coxsackievirus A16 induces structural alteration and surface modification of viral capsids. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Zhao, H.; Li, H.Y.; Han, J.F.; Deng, Y.Q.; Zhu, S.Y.; Li, X.F.; Yang, H.Q.; Li, Y.X.; Zhang, Y.; Qin, E.D.; et al. Novel recombinant chimeric virus-like particle is immunogenic and protective against both enterovirus 71 and coxsackievirus A16 in mice. Sci. Rep. 2015, 5, 7878. [Google Scholar] [CrossRef]
- Ren, J.; Wang, X.; Zhu, L.; Hu, Z.; Gao, Q.; Yang, P.; Li, X.; Wang, J.; Shen, X.; Fry, E.E.; et al. Structures of coxsackievirus A16 capsids with native antigenicity: Implications for particle expansion, receptor binding, and immunogenicity. J. Virol. 2015, 89, 10500–10511. [Google Scholar] [CrossRef]
- Ren, J.; Wang, X.; Hu, Z.; Gao, Q.; Sun, Y.; Li, X.; Porta, C.; Walter, T.S.; Gilbert, R.J.; Zhao, Y.; et al. Picornavirus uncoating intermediate captured in atomic detail. Nat. Commun. 2013, 4, 1929. [Google Scholar] [CrossRef]
- Lyu, K.; Wang, G.C.; He, Y.L.; Han, J.F.; Ye, Q.; Qin, C.F.; Chen, R. Crystal structures of enterovirus 71 (EV71) recombinant virus particles provide insights into vaccine design. J. Biol. Chem. 2015, 290, 3198–3208. [Google Scholar] [CrossRef]
- Lyu, K.; He, Y.L.; Li, H.Y.; Chen, R. Crystal structures of yeast-produced enterovirus 71 and enterovirus 71/coxsackievirus A16 chimeric virus-like particles provide the structural basis for novel vaccine design against hand-foot-and-mouth disease. J. Virol. 2015, 89, 6196–6208. [Google Scholar] [CrossRef]
- Frietze, K.M.; Peabody, D.S.; Chackerian, B. Engineering virus-like particles as vaccine platforms. Curr. Opin. Virol. 2016, 18, 44–49. [Google Scholar] [CrossRef]
- Salmons, B.; Lim, P.Y.; Djurup, R.; Cardosa, J. Non-clinical safety assessment of repeated intramuscular administration of an EV-A71 VLP vaccine in rabbits. Vaccine 2018, 36, 6623–6630. [Google Scholar] [CrossRef]
- Arita, M.; Nagata, N.; Iwata, N.; Ami, Y.; Suzaki, Y.; Mizuta, K.; Iwasaki, T.; Sata, T.; Wakita, T.; Shimizu, H. An attenuated strain of enterovirus 71 belonging to genotype A showed a broad spectrum of antigenicity with attenuated neurovirulence in cynomolgus monkeys. J. Virol. 2007, 81, 9386–9395. [Google Scholar] [CrossRef]
- Wang, G.; Cao, R.Y.; Chen, R.; Mo, L.; Han, J.F.; Wang, X.; Xu, X.; Jiang, T.; Deng, Y.Q.; Lyu, K.; et al. Rational design of thermostable vaccines by engineered peptide-induced virus self-biomineralization under physiological conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 7619–7624. [Google Scholar] [CrossRef]
- Zhang, W.; Dai, W.; Zhang, C.; Zhou, Y.; Xiong, P.; Wang, S.; Ye, X.; Liu, Q.; Zhou, D.; Huang, Z. A virus-like particle-based tetravalent vaccine for hand, foot, and mouth disease elicits broad and balanced protective immunity. Emerg. Microbes Infect. 2018, 7, 94. [Google Scholar] [CrossRef]
- Blokhina, E.A.; Kuprianov, V.V.; Stepanova, L.A.; Tsybalova, L.M.; Kiselev, O.I.; Ravin, N.V.; Skryabin, K.G. A molecular assembly system for presentation of antigens on the surface of HBc virus-like particles. Virology 2013, 435, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Kratz, P.A.; Böttcher, B.; Nassal, M. Native display of complete foreign protein domains on the surface of hepatitis B virus capsids. Proc. Natl. Acad. Sci. USA 1999, 96, 1915–1920. [Google Scholar] [CrossRef]
- Skamel, C.; Ploss, M.; Bottcher, B.; Stehle, T.; Wallich, R.; Simon, M.M.; Nassal, M. Hepatitis B virus capsid-like particles can display the complete, dimeric outer surface protein C and stimulate production of protective antibody responses against Borrelia burgdorferi infection. J. Biol. Chem. 2006, 281, 17474–17481. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, R.; Xu, L.; Li, Y.; Li, S.; Yu, H.; Li, S.; Zhu, H.; Cheng, T.; Xia, N. A novel combined vaccine based on monochimeric VLP co-displaying multiple conserved epitopes against enterovirus 71 and varicella-zoster virus. Vaccine 2017, 35, 2728–2735. [Google Scholar] [CrossRef]
- Huang, K.Y.; Lin, J.J.; Chiu, C.H.; Yang, S.; Tsao, K.C.; Huang, Y.C.; Lin, T.Y. A Potent Virus-Specific Antibody-Secreting Cell Response to Acute Enterovirus 71 Infection in Children. J. Infect. Dis. 2015, 212, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Liu, C.C.; Lin, M.H.; Ho, H.M.; Yang, Y.T.; Chow, Y.H.; Chong, P.; Sia, C. Generation of murine monoclonal antibodies which cross-neutralize human enterovirus genogroup B isolates. J. Virol. Methods 2011, 173, 189–195. [Google Scholar] [CrossRef]
- Tan, S.H.; Ong, K.C.; Perera, D.; Wong, K.T. A monoclonal antibody to ameliorate central nervous system infection and improve survival in a murine model of human Enterovirus-A71 encephalomyelitis. Antiviral Res. 2016, 132, 196–203. [Google Scholar] [CrossRef]
- Chang, G.H.; Luo, Y.J.; Wu, X.Y.; Si, B.Y.; Lin, L.; Zhu, Q.Y. Monoclonal antibody induced with inactived EV71-Hn2 virus protects mice against lethal EV71-Hn2 virus infection. Virol. J. 2010, 7, 106. [Google Scholar] [CrossRef]
- Lim, X.F.; Jia, Q.; Khong, W.X.; Yan, B.; Premanand, B.; Alonso, S.; Chow, V.T.; Kwang, J. Characterization of an isotype-dependent monoclonal antibody against linear neutralizing epitope effective for prophylaxis of enterovirus 71 infection. PLoS ONE 2012, 7, e29751. [Google Scholar] [CrossRef]
- Deng, Y.Q.; Ma, J.; Xu, L.J.; Li, Y.X.; Zhao, H.; Han, J.F.; Tao, J.; Li, X.F.; Zhu, S.Y.; Qin, E.D.; et al. Generation and characterization of a protective mouse monoclonal antibody against human enterovirus 71. Appl. Microbiol. Biotechnol. 2015, 99, 7663–7671. [Google Scholar] [CrossRef]
- Lee, H.; Cifuente, J.O.; Ashley, R.E.; Conway, J.F.; Makhov, A.M.; Tano, Y.; Shimizu, H.; Nishimura, Y.; Hafenstein, S. A strain-specific epitope of enterovirus 71 identified by cryo-electron microscopy of the complex with fab from neutralizing antibody. J. Virol. 2013, 87, 11363–11370. [Google Scholar] [CrossRef]
- Lin, T.; Xianyu, L.; Lyu, S. Monoclonal neutralizing antibodies against EV71 screened from mice immunized with yeast-produced virus-like particles. Virol. Sin. 2015, 30, 208–213. [Google Scholar] [CrossRef]
- Li, X.; Mao, C.; Ma, S.; Wang, X.; Sun, Z.; Yi, Y.; Guo, M.; Shen, X.; Sun, L.; Bi, S. Generation of neutralizing monoclonal antibodies against Enterovirus 71 using synthetic peptides. Biochem. Biophys. Res. Commun. 2009, 390, 1126–1128. [Google Scholar] [CrossRef] [PubMed]
- Ng, Q.; He, F.; Kwang, J. Recent Progress towards Novel EV71 Anti-Therapeutics and Vaccines. Viruses 2015, 7, 6441–6457. [Google Scholar] [CrossRef]
- Almagro, J.; Fransson, J. Humanization of antibodies. Front. Biosci. 2008, 13, 1619–1633. [Google Scholar] [CrossRef]
- Wong, Y.H.; Kumar, A.; Liew, C.W.; Tharakaraman, K.; Srinivasaraghavan, K.; Sasisekharan, R.; Verma, C.; Lescar, J. Molecular basis for dengue virus broad cross-neutralization by humanized monoclonal antibody 513. Sci. Rep. 2018, 8, 8449. [Google Scholar] [CrossRef]
- Tan, Y.; Ng, Q.; Jia, Q.; Kwang, J.; He, F. A Novel Humanized Antibody Neutralizes H5N1 Influenza Virus via Two Different Mechanisms. J. Virol. 2015, 89, 3712–3722. [Google Scholar] [CrossRef]
- Laursen, N.S.; Friesen, R.H.E.; Zhu, X.; Jongeneelen, M.; Blokland, S.; Vermond, J.; van Eijgen, A.; Tang, C.; van Diepen, H.; Obmolova, G.; et al. Universal protection against influenza infection by a multidomain antibody to influenza hemagglutinin. Science 2018, 362, 598–602. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Virus | Region | Name and Position | Peptide Sequences | Epitopes | Ref. |
|---|---|---|---|---|---|
| EV-A71 | VP1 | PEP23 (aa 41–55) | TGEVPALQAAEIGAS | IgG | [25] |
| VP1–15 (aa 43–54) | KVPALQAAEIGA | IgG | [26] | ||
| VP1–14 (aa 40–51) | DTGEVPALQAAE | IgM | [26] | ||
| VP1 | PEP27 (aa 142–156) | PTGEVVPQLLQYMFV | IgM | [25] | |
| VP1 | SP55 (aa 163–177) | PESRESLAWQTATNP | IgG | [22] | |
| VP1 | SP70 (aa 208–222) | YPTFGEHKQEKDLEYC | IgM and IgG | [22] | |
| VP1 (aa 208–222) | YPTFGEHKQEKDLEYC | IgG | [24] | ||
| VP1–43 (aa 211–220) | FGEHKQEKDL | IgG | [23] | ||
| VP2 | VP2–28 (aa 136–150) | AGGTGTEDSHPPYKQ | IgG | [23] | |
| 7C7 (aa 142–146) | EDSHP | IgG | [28] | ||
| VP2 (aa 141–155) | TEDSHPPYKQTQPGA | IgG | [27] | ||
| VP4 | VP4N20 (aa 1–20) | GSQVSTQRSGSHENSNSATE | Neutralizing | [31] | |
| CV-A16 | VP1 | PEP32 (aa 94–108) | TMPTMGTQNTDGYAN | IgG | [34] |
| PEP37 (aa 109–123) | WDIDLMGYAQLRRKC | IgG | [34] | ||
| PEP55 (aa 163–177) | PTSRDSFAWQTATNP | IgG | [34] | ||
| PEP63 (aa 187–201) | PAQVSVPFMSPASAY | IgG | [34] | ||
| PEP71 (aa 211–225) | FGEHLQANDLDYGQC | IgG | [34] | ||
| PEP91 (aa 271–285) | YLFKTNPNYKGNDIK | IgG | [34] | ||
| VP3 | VP3–41 (aa 176–190) | HYRAHARAGYFDYYT | IgG | [32] | |
| VP4 | VP4N20 (aa 1–20) | GSQVSTQRSGSHENSNSASE | Neutralizing | [35] |
| Virus | Name | Epitope Type | Epitope/Region Recognised | Isotype | Ref. |
|---|---|---|---|---|---|
| EV-A71 | N1 | Linear | Recognized a section of the SP70 epitope. Neutralized 2 subgenotypes tested (B4 and B5). | IgG2a | [67] |
| N3 | Linear | Recognized a section of the SP70 epitope. Neutralized 2 subgenotypes tested (B4 and B5). | IgG2a | [67] | |
| N4 | Linear | Recognized a section of the SP70 epitope. Neutralized 2 subgenotypes tested (B4 and B5). | IgG2a | [67] | |
| N6 | Linear | Recognized a section of the SP70 epitope. Neutralized 2 subgenotypes tested (B4 and B5). | IgG2a | [67] | |
| BB1A5 | Linear | Recognized amino acids 136–155 in VP2 (AGGTGTEDSHPPYKQTQPGA). Neutralized B3, B4, C2 and C5 subgenotypes. | IgG2a | [28] | |
| 3D1 | Unknown | Recognized VP1. Neutralized all 8 subgenotypes tested (C1, C2, C3, C4, C5, B3, B4, and B5). | IgM | [68] | |
| 4E8 | Unknown | Recognized two peptides P25 (aa2 40–250) and P26 (aa 250–260) of VP1. Neutralized C4 subgenotype (only subgenotype tested). | IgG | [69] | |
| 10D3 | Conformational | Recognized the knob region in VP3. Neutralized all 11 subgenotypes of EV-A71. | IgM | [29] | |
| mAb51 | Linear | Recognized a portion of the SP70 epitope (KQEKD). Neutralized all 11 subgenotypes of EV-A71. | IgM | [70] | |
| 2G8 | Linear | Recognized the SP70 epitope. Neutralized C4 subgenotype. | IgM | [71] | |
| MA28-7 | Conformational | Recognized the region comprising Gly145, Glu98, Lys242 and Lys244 in VP1. Neutralized 6 genotypes tested (A, B1, B3, B4, and C2). | N/A | [72] | |
| D6 | Unknown | Neutralized subgenotype C4 (only subgenotype tested). | N/A | [53] | |
| A9 | Unknown | Neutralized subgenotype C4 (only subgenotype tested). | N/A | [53] | |
| 5H7 | Conformational | Recognized conformational epitope harbouring Ser74 in VP3. Neutralized all 11 subgenotypes tested. | IgG | [30] | |
| D4 | Unknown | Neutralized subgenotype C4 (only subgenotype tested). | IgG | [73] | |
| G12 | Unknown | Neutralized subgenotype C4 (only subgenotype tested). | IgG | [73] | |
| 22A12 | Linear | Neutralized subgenotype C4 (only subgenotype tested). | IgG | [74] | |
| VP4N20 mAb | Linear | Recognized the VP4N20 epitope in VP4. Neutralized EV-A71 tested (genogroup A and subgenotype C4) and CV-A16. | N/A | [35] | |
| CV-A16 | VP4N20 mAb | Linear | Recognized the VP4N20 epitope in VP4. Neutralized EV-A71 tested (genogroup A and subgenotype C4) and CV-A16. | N/A | [35] |
| C33 | Unknown | Recognized the SP70 epitope. Neutralized genotype B of CV-A16 (only one tested). | N/A | [53] | |
| CV-A6 | 1D5 | Conformational | Recognized a region in VP1 comprising BC, DE, EF, and HI loops. | IgG | [38,39] |
| CV-A10 | 2G8 | Conformational | Recognized the VP1 C-terminus, VP2 EF loop, and VP3 AB loop. | IgG | [36] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anasir, M.I.; Poh, C.L. Advances in Antigenic Peptide-Based Vaccine and Neutralizing Antibodies against Viruses Causing Hand, Foot, and Mouth Disease. Int. J. Mol. Sci. 2019, 20, 1256. https://doi.org/10.3390/ijms20061256
Anasir MI, Poh CL. Advances in Antigenic Peptide-Based Vaccine and Neutralizing Antibodies against Viruses Causing Hand, Foot, and Mouth Disease. International Journal of Molecular Sciences. 2019; 20(6):1256. https://doi.org/10.3390/ijms20061256
Chicago/Turabian StyleAnasir, Mohd Ishtiaq, and Chit Laa Poh. 2019. "Advances in Antigenic Peptide-Based Vaccine and Neutralizing Antibodies against Viruses Causing Hand, Foot, and Mouth Disease" International Journal of Molecular Sciences 20, no. 6: 1256. https://doi.org/10.3390/ijms20061256
APA StyleAnasir, M. I., & Poh, C. L. (2019). Advances in Antigenic Peptide-Based Vaccine and Neutralizing Antibodies against Viruses Causing Hand, Foot, and Mouth Disease. International Journal of Molecular Sciences, 20(6), 1256. https://doi.org/10.3390/ijms20061256