Low Expression of IL-10 in Circulating Bregs and Inverted IL-10/TNF-α Ratio in Tears of Patients with Perennial Allergic Conjunctivitis: A Preliminary Study


 ,
,
Abstract
1. Introduction
2. Results
2.1. Frequency of CD19+IL-10+ B Cell Subsets in Peripheral Blood
2.2. Diminished Expression of IL-10 in CD19+CD38+ B Cells in Patients with Perennial Allergic Conjunctivitis
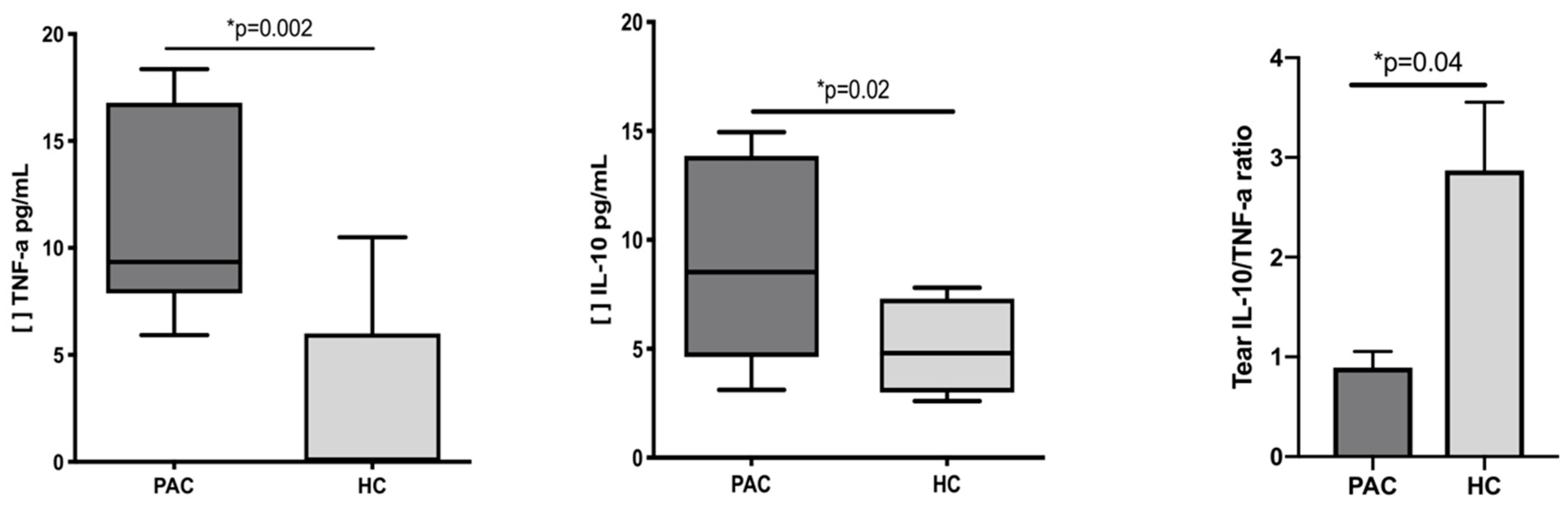
2.3. Increased Concentration of TNF-α and IL-10 in the Tears of Patients with Perennial Allergic Conjunctivitis
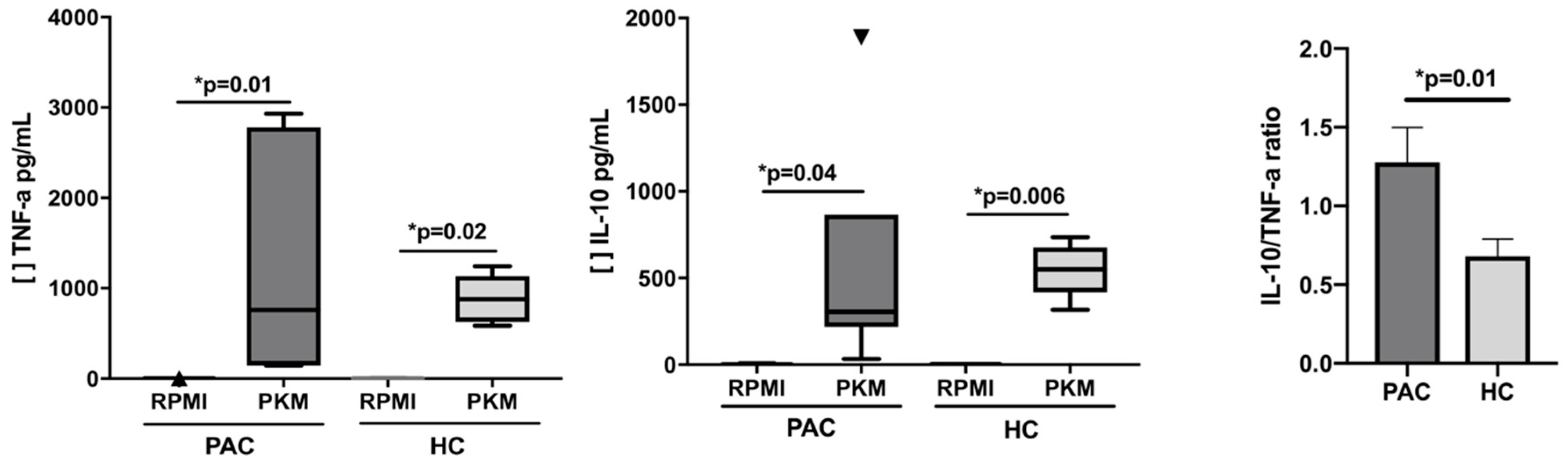
2.4. Functional Evaluation of B Cells after Mitogen Stimulation
3. Discussion
4. Materials and Methods
4.1. Patients and Health Controls
4.2. Monoclonal Antibodies and Reagents
4.3. Peripheral Blood Samples
4.4. Immunofluorescence Staining of Cell Surface Markers
4.5. Immunofluorescence Staining of Intracellular Markers
4.6. Flow Cytometric Analysis
4.7. Tear Samples
4.8. Serum Samples
4.9. Cell Cultures
4.10. Determination of Tear and Serum Cytokines
4.11. Determination of Total IgE (tIgE) and Specific IgE (sIgE)
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miraldi Utz, V.; Kaufman, A.R. Allergic eye disease. Pediatr. Clin. N. Am. 2014, 61, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Rosario, N.; Bielory, L. Epidemiology of allergic conjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Ezquerro, M.C.; Guerrero-Guerra, C.; Galán, C.; Serrano-Silva, N.; Guidos-Fogelbach, G.; Jiménez-Martínez, M.C.; Larenas-Linnemann, D.; López-Espinosa, E.D.; Ayala-Balboa, J. Pollen in the atmosphere of Mexico City and its impact on the health of the pediatric population. Atmos. Environ. 2018, 186, 198–208. [Google Scholar] [CrossRef]
- Passali, D.; Cingi, C.; Staffa, P.; Passali, F.; Muluk, N.B.; Bellussi, M.L. The International Study of the Allergic Rhinitis Survey: Outcomes from 4 geographical regions. Asia Pac. Allergy 2018, 8, e7. [Google Scholar] [CrossRef] [PubMed]
- Shaker, M.; Salcone, E. An update on ocular allergy. Curr. Opin. Allergy Clin. Immunol. 2016, 16, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Magaña, D.; Aguilar, G.; Linares, M.; Ayala-Balboa, J.; Santacruz, C.; Chávez, R.; Estrada-Parra, S.; Garfias, Y.; Lascurain, R.; Jiménez-Martínez, M.C. Intracellular IL-4, IL-5 and IFN-g as the main characteristic of CD4+CD30+ T cells after allergen-stimulation, in patients with vernal keratoconjunctivitis. Mol. Vis. 2015, 21, 443–450. [Google Scholar] [PubMed]
- Anderson, A.E.; Mackerness, K.J.; Aizen, M.; Carr, V.A.; Nguyen, D.; Du Pre, F.; Durham, S.R.; Robinson, D.S. Seasonal changes in suppressive capacity of CD4+ CD25+ T cells from patients with hayfever are allergen-specific and may result in part from expansion of effector T cells among the CD25+ population. Clin. Exp. Allergy 2009, 39, 1693–1699. [Google Scholar] [PubMed]
- Galicia-Carreón, J.; Santacruz, C.; Ayala-Balboa, J.; Robles-Contreras, A.; Perez-Tapia, S.M.; Garfias, Y.; Hong, E.; Jiménez-Martínez, M.C. An Imbalance Between Frequency of CD4+CD25+FOXP3+ Regulatory T Cells, and CCR4+ and CCR9+ Circulating Helper T cells is Associated with Active Perennial Allergic Conjunctivitis. Clin. Dev. Immunol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Palomares, O.; Martín-Fontecha, M.; Lauener, R.; Traidl-Hoffmann, C.; Cavkaytar, O.; Akdis, M.; Akdis, C.A. Regulatory T cells and immune regulation of allergic diseases: Roles of IL-10 and TGF-β. Genes Immunol. 2014, 15, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Poojary, K.V.; Kong, Y.M.; Farrar, M.A. Control of Th2-Mediated Inflammation by Regulatory T Cells. Am. J. Pathol. 2010, 177, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Ashour, H.M.; Niederkorn, J.Y. Expansion of B cells is necessary for the induction of T-cell tolerance elicited through the anterior chamber of the eye. Int. Arch Allergy Immunol. 2007, 144, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Pennati, A.; Ng, S.; Wu, Y.; Murphy, J.R.; Deng, J.; Rangaraju, S.; Asress, S.; Blanchfield, J.L.; Evavold, B.; Galipeau, J. Regulatory B Cells Induce Formation of IL-10-Expressing T Cells in Mice with Autoimmune Neuroinflammation. J. Neurosci. 2016, 36, 12598–12610. [Google Scholar] [CrossRef] [PubMed]
- Flores-Borja, F.; Bosma, A.; Ng, D.; Reddy, V.; Ehrenstein, M.R.; Isenberg, D.A.; Mauri, C. CD19+CD24hiCD38hi B cells maintain regulatory T cells while limiting TH1 and TH17 differentiation. Sci. Transl. Med. 2013, 5, 173ra23. [Google Scholar] [CrossRef] [PubMed]
- Fillatreau, S.; Sweenie, C.H.; McGeachy, M.J.; Gray, D.; Anderton, S.M. B cells regulate autoimmunity by provision of IL-10. Nat. Immunol. 2002, 3, 944. [Google Scholar] [CrossRef] [PubMed]
- Van de Veen, W.; Stanic, B.; Wirz, O.F.; Jansen, K.; Globinska, A.; Akdis, M. Role of regulatory B cells in immune tolerance to allergens and beyond. J. Allergy Clin. Immunol. 2016, 138, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Lundy, S.K.; Berlin, A.A.; Martens, T.F.; Lukacs, N.W. Deficiency of regulatory B cells increases allergic airway inflammation. Inflamm. Res. 2005, 54, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Bekeredjian-Ding, I.; Foermer, S.; Kirschning, C.J.; Parcina, M.; Heeg, K. Poke weed mitogen requires Toll-like receptor ligands for proliferative activity in human and murine B lymphocytes. PLoS ONE 2012, 7, e29806. [Google Scholar] [CrossRef] [PubMed]
- Bielory, L. Allergic and immunologic disorders of the eye. Part II: Ocular allergy. J. Allergy Clin. Immunol. 2000, 106, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Carreón, J.; Santacruz, C.; Hong, E.; Jiménez-Martínez, M.C. The ocular surface: From physiology to the ocular allergic diseases. Rev. Alerg. Mexico 2013, 60, 172–183. [Google Scholar]
- Miyazaki, D.; Kuo, C.H.; Tominaga, T.; Inoue, Y.; Ono, S.J. Regulatory function of CpG-activated B cells in late-phase experimental allergic conjunctivitis. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1626–1635. [Google Scholar] [CrossRef] [PubMed]
- Jaroszeski, M.J.; Radcliff, G. Fundamentals of flow cytometry. Mol. Biotechnol. 1999, 11, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Hulstaert, F.; Hannet, I.; Deneys, V.; Munhyeshuli, V.; Reichert, T.; De Bruyere, M.; Strauss, K. Age-related changes in human blood lymphocyte subpopulations. II. Varying kinetics of percentage and absolute count measurements. Clin. Immunol. Immunopathol. 1994, 70, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Morbach, H.; Eichhorn, E.M.; Liese, J.G.; Girschick, H.J. Reference values for B cell subpopulations from infancy to adulthood. Clin. Exp. Immunol. 2010, 162, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Valiathan, R.; Ashman, M.; Asthana, D. Effects of Ageing on the Immune System: Infants to Elderly. Scand. J. Immunol. 2016, 83, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Iwata, Y.; Matsushita, T.; Horikawa, M.; Dilillo, D.J.; Yanaba, K.; Venturi, G.M.; Szabolcs, P.M.; Bernstein, S.H.; Magro, C.M.; Williams, A.D.; Hall, R.P.; St Clair, E.W.; Tedder, T.F. Characterization of a rare IL-10-competent B-cell subset in humans that parallels mouse regulatory B10 cells. Blood 2011, 117, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Blair, P.A.; Noreña, L.Y.; Flores-Borja, F.; Rawlings, D.J.; Isenberg, D.A.; Ehrenstein, M.R.; Mauri, C. CD19(+)CD24(hi)CD38(hi) B cells exhibit regulatory capacity in healthy individuals but are functionally impaired in systemic Lupus Erythematosus patients. Immunity 2010, 32, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Pantoja, M.; López-Herrera, G.; Romero-Ramírez, H.; Santos-Argumedo, L.; Chávez-Rueda, A.K.; Hernández-Cueto, Á.; Flores-Muñoz, M.; Rodríguez-Alba, J.C. CD38 protein deficiency induces autoimmune characteristics and its activation enhances IL-10 production by regulatory B cells. Scand. J. Immunol. 2018, 87, e12664. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Qiu, J.; Yu, L.; Hu, X.; Zhao, P.; Jiang, Y. Increased numbers of CD5+CD19+CD1dhighIL-10+ Bregs, CD4+Foxp3+ Tregs, CD4+CXCR5+Foxp3+ follicular regulatory T (TFR) cells in CHB or CHC patients. J. Transl. Med. 2014, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.I.; Hong, S.H.; Cho, S.H.; Lee, W.J.; Lee, S.E. Induction of IL-10-producing CD1dhighCD5+ regulatory B cells following Babesia microti-infection. PLoS ONE 2012, 7, e46553. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Terao, T.; Osawa, T. Carbohydrate-binding specificity of pokeweed mitogens. Biochim. Biophys. Acta 1978, 538, 384–396. [Google Scholar] [CrossRef]
- Dewald, J.H.; Colomb, F.; Bobowski-Gerard, M.; Groux-Degroote, S.; Delannoy, P. Role of Cytokine-Induced Glycosylation Changes in Regulating Cell Interactions and Cell Signaling in Inflammatory Diseases and Cancer. Cells 2016, 29, 43. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Khosraviani, S.; Noel, S.; Mohan, D.; Donner, T.; Hamad, A.R. Interleukin-10 paradox: A potent immunoregulatory cytokine that has been difficult to harness for immunotherapy. Cytokine 2014, 74, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, D.F.; Zlotnik, A.; Vieira, P.; Mosmann, T.R.; Howard, M.; Moore, K.W.; O’Garra, A. IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J. Immunol. 1991, 146, 3444–3451. [Google Scholar] [PubMed]
- Thompson-Snipes, L.; Dhar, V.; Bond, M.W.; Mosmann, T.R.; Moore, K.W.; Rennick, D.M. Interleukin 10: A novel stimulatory factor for mast cells and their progenitors. J. Exp. Med. 1991, 173, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Elieh Ali Komi, D.; Rambasek, T.; Bielory, L. Clinical implications of mast cell involvement in allergic conjunctivitis. Allergy 2018, 73, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Curnow, S.J.; Zhan, H.; Calder, V.L. Multiple cytokines in human tear specimens in seasonal and chronic allergic eye disease and in conjunctival fibroblast cultures. Clin. Exp. Allergy 2006, 36, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Pelikan, Z. Cytological changes in tears during the secondary conjunctival response induced by nasal allergy. Br. J. Ophthalmol. 2012, 96, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Pelikan, Z. Cytokine profiles in tears accompanying the secondary conjunctival responses induced by nasal allergy. Curr. Eye Res. 2014, 39, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Skwor, T.A.; Atik, B.; Kandel, R.P.; Adhikari, H.K.; Sharma, B.; Dean, D. Role of secreted conjunctival mucosal cytokine and chemokine proteins in different stages of trachomatous disease. PLoS Negl. Trop. Dis. 2008, 2, e264. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Han, S.J.; Nam, S.M.; Yoon, S.C.; Ahn, J.M.; Kim, T.I.; Kim, E.K.; Seo, K.Y. Analysis of tear cytokines and clinical correlations in Sjögren syndrome dry eye patients and non-Sjögren syndrome dry eye patients. Am. J. Ophthalmol. 2013, 156, 247.e1–253.e1. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Sumi, T.; Fukuda, K.; Kumagai, N.; Nishida, T.; Yagita, H.; Ueno, H. Interleukin 10 and transforming growth factor beta contribute to the development of experimentally induced allergic conjunctivitis in mice during the effector phase. Br. J. Ophthalmol. 2006, 90, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wandu, W.S.; St Leger, A.; Kielczewski, J.; Wawrousek, E.F.; Chan, C.C.; Gery, I. Unlike Th1/Th17 cells, Th2/Th9 cells selectively migrate to the limbus/conjunctiva and initiate an eosinophilic infiltration process. Exp. Eye Res. 2018, 166, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Bemelmans, M.H.A.; van Tits, L.J.H.; Buurman, W.A. Tumor Necrosis Factor: Function, Release and Clearance. Crit. Rev. Immunol. 2017, 37, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Enríquez-de-Salamanca, A.; Calder, V.; Gao, J.; Galatowicz, G.; García-Vázquez, C.; Fernández, I.; Stern, M.E.; Diebold, Y.; Calonge, M. Cytokine responses by conjunctival epithelial cells: An in vitro model of ocular inflammation. Cytokine 2008, 44, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Motterle, L.; Bortolotti, M. Allergy and the eye. Clin. Exp. Immunol. 2008, 153 (Suppl. 1), 17–21. [Google Scholar] [CrossRef] [PubMed]
- Fukagawa, K.; Okada, N.; Fujishima, H.; Nakajima, T.; Takano, Y.; Tanaka, M.; Dogru, M.; Satake, Y.; Tsubota, K.; Saito, H. Corneal and conjunctival fibroblasts are major sources of eosinophil-recruiting chemokines. Allergol. Int. 2009, 58, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A. The central role of conjunctival mast cells in the pathogenesis of ocular allergy. Curr. Allergy Asthma Rep. 2002, 2, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Hamrah, P.; Shimazaki, J. Bilateral Alterations in Corneal Nerves, Dendritic Cells, and Tear Cytokine Levels in Ocular Surface Disease. Cornea 2016, 35 (Suppl. 1), S65–S70. [Google Scholar] [CrossRef] [PubMed]
- Van de Veen, W.; Stanic, B.; Yaman, G.; Wawrzyniak, M.; Söllner, S.; Akdis, D.G.; Rückert, B.; Akdis, C.A.; Akdis, M. IgG4 production is confined to human IL-10-producing regulatory B cells that suppress antigen-specific immune responses. J. Allergy Clin. Immunol. 2013, 131, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Van de Veen, W. The role of regulatory B cells in allergen immunotherapy. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Guo, H.; Liu, Z.; Peng, T.; Hu, X.; Han, M.; Yang, X.; Zhou, X.; Li, H. Analysis of Peripheral B Cell Subsets in Patients With Allergic Rhinitis. Allergy Asthma Immunol. Res. 2018, 10, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, J.T.; Angell, E. Paradoxical effects of IL-10 in endotoxin-induced uveitis. J. Immunol. 1995, 15, 4090–4094. [Google Scholar]
- Shen, J.; Taylor, N.; Duncan, L.; Kovesdi, I.; Bruder, J.T.; Forrester, J.V.; Dick, A.D. Ex vivo adenovirus mediated gene transfection of human conjunctival epithelium. Br. J. Ophthalmol. 2001, 85, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Stevenson, D.; Ritter, T.; Schechter, J.E.; Mircheff, A.K.; Kaslow, H.R.; Trousdale, M.D. Expression of IL-10 and TNF-inhibitor genes in lacrimal gland epithelial cells suppresses their ability to activate lymphocytes. Cornea 2002, 21, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Santacruz, C.; Linares, M.; Garfias, Y.; Loustaunau, L.M.; Pavon, L.; Perez-Tapia, S.M.; Jimenez-Martinez, M.C. Expression of IL-8, IL-6 and IL-1β in tears as a main characteristic of the immune response in human microbial keratitis. Int. J. Mol. Sci. 2015, 16, 4850–4864. [Google Scholar] [CrossRef] [PubMed]



 and
and  Corresponded to the outliers values. [ ] Concentration of cytokine.
and Corresponded to the outliers values. [ ] Concentration of cytokine.
Corresponded to the outliers values. [ ] Concentration of cytokine.
and Corresponded to the outliers values. [ ] Concentration of cytokine.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Characteristics | PAC | HC | p Value |
|---|---|---|---|
| (n = 16) MD (IQR) | (n = 8) MD (IQR) | ||
| Age | 12.5 (11–14) | 12 (11–15.5) | NS |
| Male | 13 (11–14) | 11 (11–13.3) | NS |
| Female | 11 (10.5–13.5) | 14.5 (8.5–16.8) | NS |
| TBUT (sg) | 4.5 (3–6) | 6 (3.5–8) | NS |
| Schirmer Test (mm) | 16.5 (9–27) | 20 (14.8–29.8) | NS |
| SPT (mm) to Der p | 6 (3–8) | 0 (0–0) | 0.0006 |
| IgA | 174 (140–211) | 171 (86–213) | NS |
| IgG | 1230 (1169–1377) | 1310 (649–1933) | NS |
| IgM | 116 (85–146) | 127 (103–516) | NS |
| tIgE | 404 (164–585) | 58.3 (29.3–182.5) | 0.002 |
| sIgE | 29.9 (11.4–62.9) | 0 (0–0) * | <0.0001 |
| B Cell Subsets | PAC | HC | p Value |
|---|---|---|---|
| (n = 16) | (n = 8) | ||
| MD (IQR) | MD (IQR) | ||
| CD19+ | 18.8 (16.9–21.9) | 17.7 (15.7–20.1) | 0.5177 |
| CD19+CD38+ | 84.4 (77.9–88.6) | 86 (71.9–91.8) | 0.9649 |
| CD19+CD38Lo | 61.5 (36.8–64.1) | 55.2 (40.6–60.5) | 0.2597 |
| CD19+CD38Med | 23.5 (14.2–31.3) | 17.5 (7.2–36.7) | 0.6650 |
| CD19+CD38Hi | 5 (3.0–11.9) | 5.25 (1.8–14.9) | 0.2834 |
| CD19+IL-10+ | 3.8 (2.7–4.6) | 3.3 (1.9–3.5) | 0.2198 |
| CD19+CD38Lo IL-10+ | 2.5 (1.6–4.2) | 2.3 (1.0–7.0) | 0.500 |
| CD19+CD38Med IL-10+ | 5 (2.6–6.7) | 4.8 (3.7–5.4) | 0.449 |
| CD19+CD38Hi IL-10+ | 4.7 (3.2–7.3) | 10.4 (3.9–15.3) | 0.419 |
| Tear Cytokines | PAC (n = 16) | HC (n = 8) | p Value |
|---|---|---|---|
| MD (IQR) | MD (IQR) | ||
| pg/mL | pg/mL | ||
| IL-1β | 32.46 (19.85–47.63) | 33.8 (18.02–38.79) | 0.4435 |
| IL-6 | 25.46 (14.61–40.41) | 19.95 (17.36–25.28) | 0.3333 |
| IL-8 | 167 (127–222.7) | 176 (158–245) | 0.3031 |
| IL-10 | 8.5 (4.6–13.9) | 4.8 (3.0–7.3) | 0.02 |
| IL-12p70 | 27.17 (7.9–39.53) | 18.6 (7.9–22.6) | 0.1750 |
| TNF-α | 9.34 (7.88–16.79) | 0 (0–6.0) * | 0.002 |
| SN Cytokines | PAC (n = 16) | HC (n = 8) | ||||
|---|---|---|---|---|---|---|
| MD (IQR) | MD (IQR) | |||||
| pg/mL | pg/mL | |||||
| RPMI | PKM | p | RPMI | PKM | p | |
| IL-1β | 6.6 (5.2–7.6) | 454.7 (108.8–1342) | 0.03 | 6.3 (5.9–6.9) | 830 (380–1161) | 0.01 |
| IL-6 | 3.8 (3.1–4.6) | 3078 (2137–8638) | 0.03 | 4.8 (4–9.8) | 5084 (2508–6197) | 0.002 |
| IL-8 | 919 (318–1043) | 20,237 (10,558–21,513) | 0.0003 | 1223 (501.5–1654) | 22,075 (17,873–24,910) | 0.004 |
| IL-10 | 0 (0–0.3) * | 304.8 (218–866) | 0.006 | 3.3 (0–4.6) * | 550.2 (418–675) | 0.04 |
| IL-12p70 | 4.6 (2.2–4.9) | 21 (4.5–50) | 0.02 | 3.5 (2.2–4.5) | 8.9 (4.8–47.3) | 0.03 |
| TNF-α | 0 (0–1.9) * | 877 (627–1131) | 0.01 | 0 (0–1.2) * | 757.6 (144.9–2932) | 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar, A.; Casanova-Méndez, I.; Pacheco-Quito, M.; Velázquez-Soto, H.; Ayala-Balboa, J.; Graue-Hernández, E.O.; Serafín-López, J.; Jiménez-Martínez, M.C. Low Expression of IL-10 in Circulating Bregs and Inverted IL-10/TNF-α Ratio in Tears of Patients with Perennial Allergic Conjunctivitis: A Preliminary Study. Int. J. Mol. Sci. 2019, 20, 1035. https://doi.org/10.3390/ijms20051035
Salazar A, Casanova-Méndez I, Pacheco-Quito M, Velázquez-Soto H, Ayala-Balboa J, Graue-Hernández EO, Serafín-López J, Jiménez-Martínez MC. Low Expression of IL-10 in Circulating Bregs and Inverted IL-10/TNF-α Ratio in Tears of Patients with Perennial Allergic Conjunctivitis: A Preliminary Study. International Journal of Molecular Sciences. 2019; 20(5):1035. https://doi.org/10.3390/ijms20051035
Chicago/Turabian StyleSalazar, Alberto, Israel Casanova-Méndez, Michele Pacheco-Quito, Henry Velázquez-Soto, Julio Ayala-Balboa, Enrique O. Graue-Hernández, Jeanet Serafín-López, and María C. Jiménez-Martínez. 2019. "Low Expression of IL-10 in Circulating Bregs and Inverted IL-10/TNF-α Ratio in Tears of Patients with Perennial Allergic Conjunctivitis: A Preliminary Study" International Journal of Molecular Sciences 20, no. 5: 1035. https://doi.org/10.3390/ijms20051035
APA StyleSalazar, A., Casanova-Méndez, I., Pacheco-Quito, M., Velázquez-Soto, H., Ayala-Balboa, J., Graue-Hernández, E. O., Serafín-López, J., & Jiménez-Martínez, M. C. (2019). Low Expression of IL-10 in Circulating Bregs and Inverted IL-10/TNF-α Ratio in Tears of Patients with Perennial Allergic Conjunctivitis: A Preliminary Study. International Journal of Molecular Sciences, 20(5), 1035. https://doi.org/10.3390/ijms20051035