Unraveling the Molecular Mechanism of Selective Antimicrobial Activity of 2(5H)-Furanone Derivative against Staphylococcus aureus

 ,
, 
 ,
,  , and
, and
Abstract
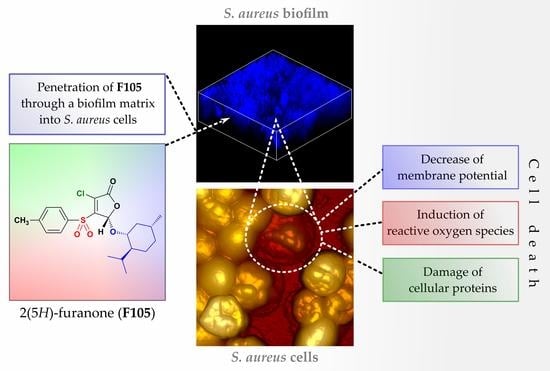
1. Introduction
2. Results
2.1. Antimicrobial Spectrum of F105
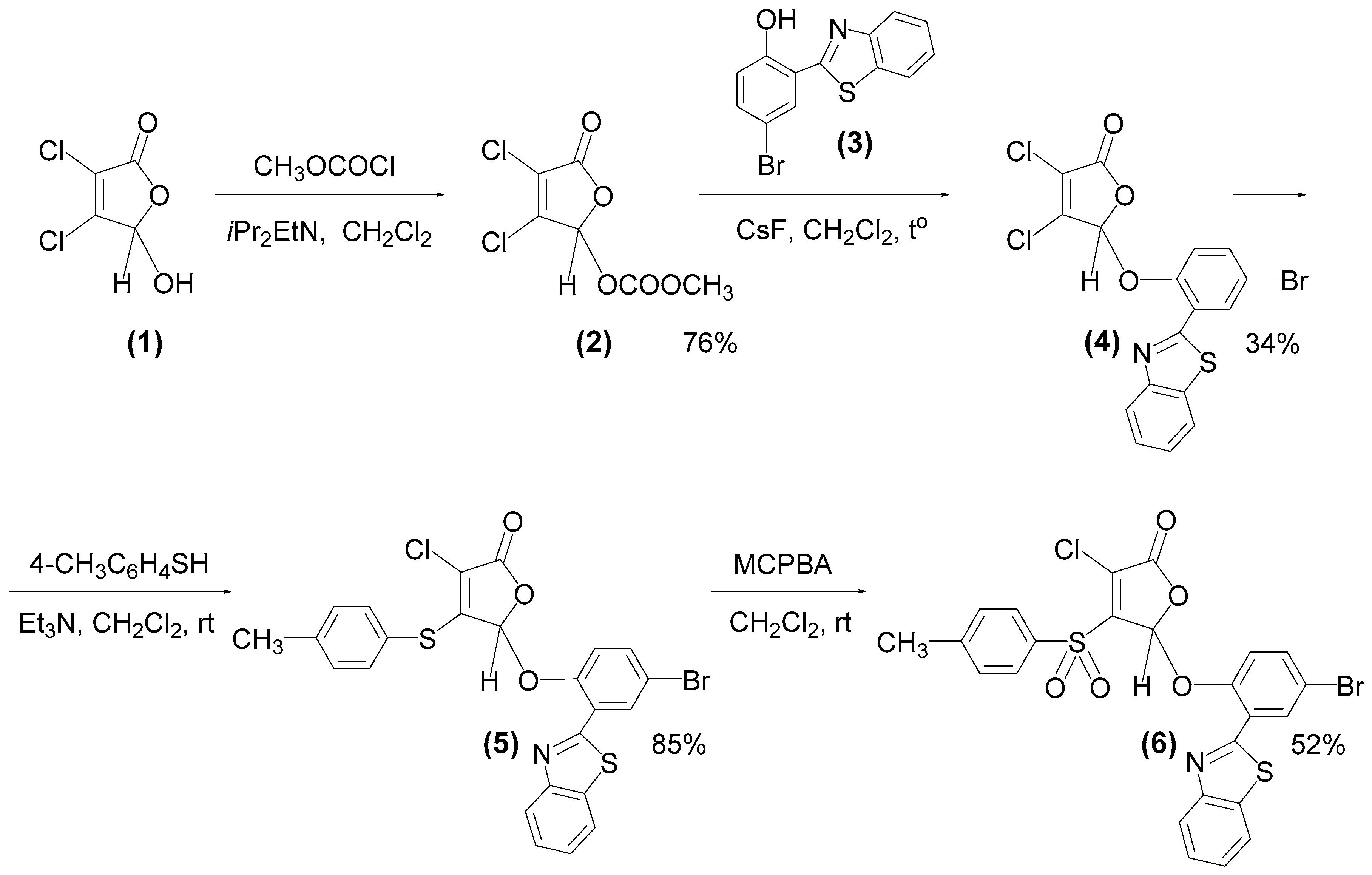
2.2. Synthesis of Fluorescent Compound 6 (F145)
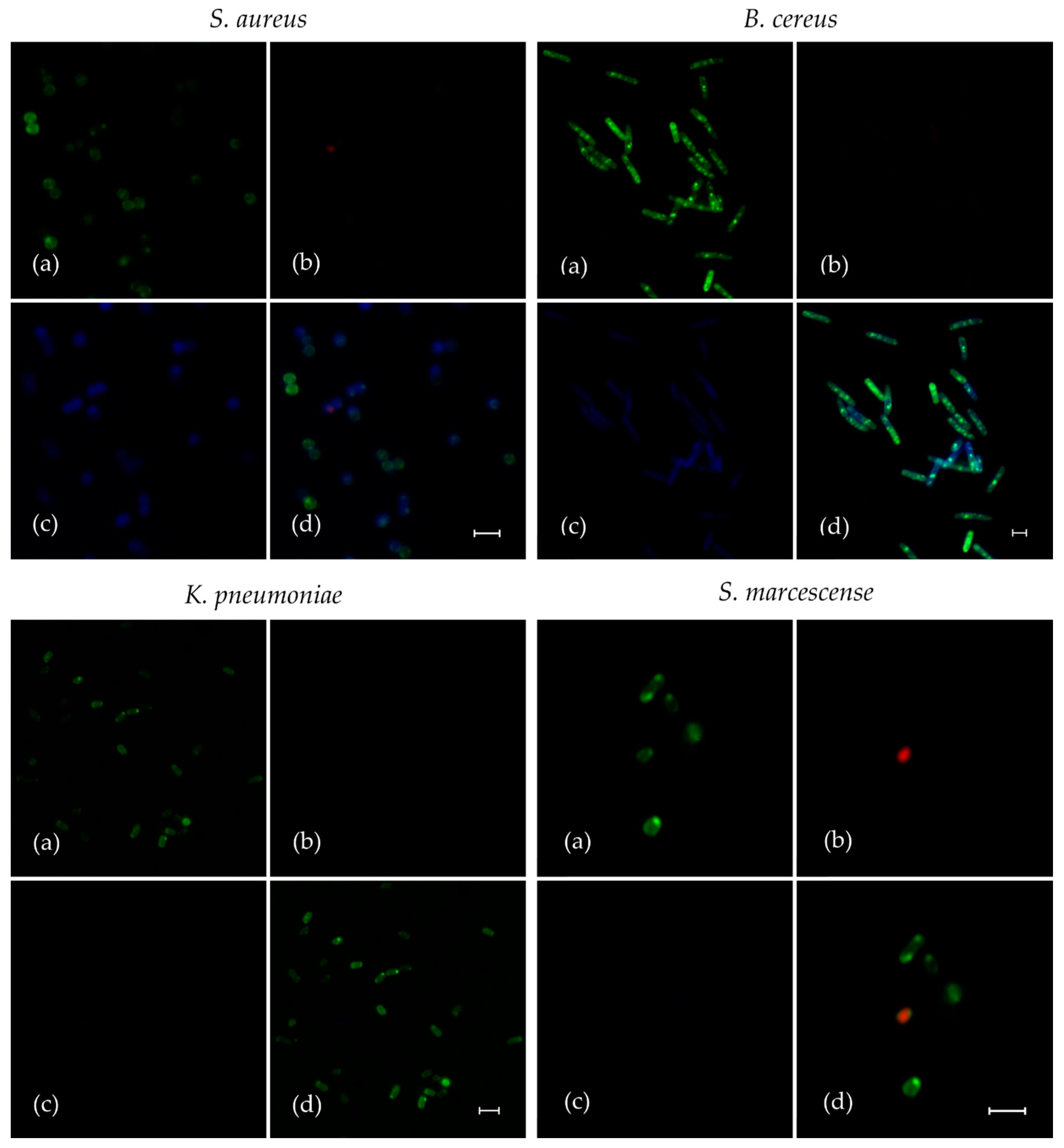
2.3. Penetration of F105 Analogue into Bacterial Cells
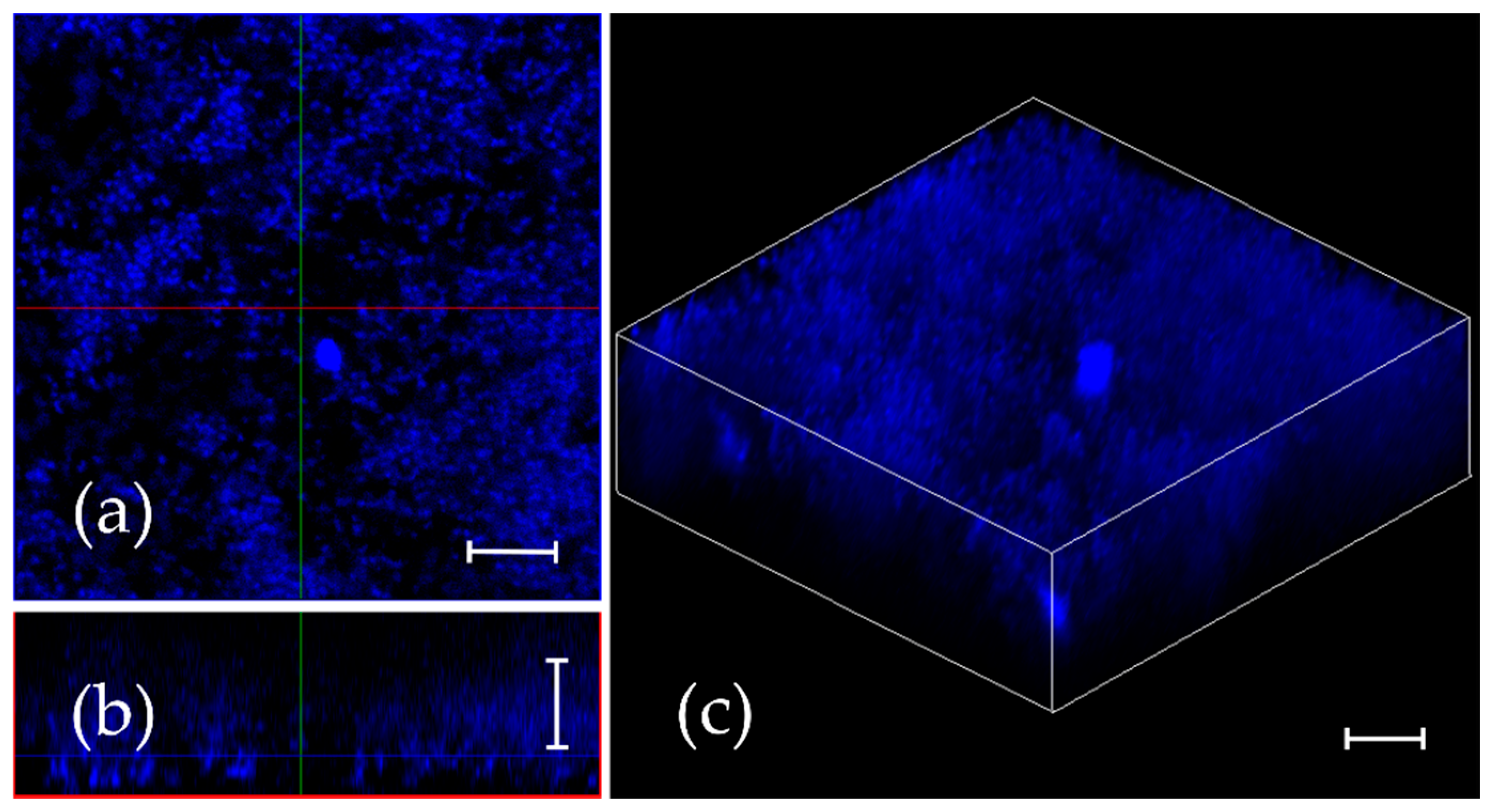
2.4. Penetration of F105 Analogue into S. aureus Biofilm
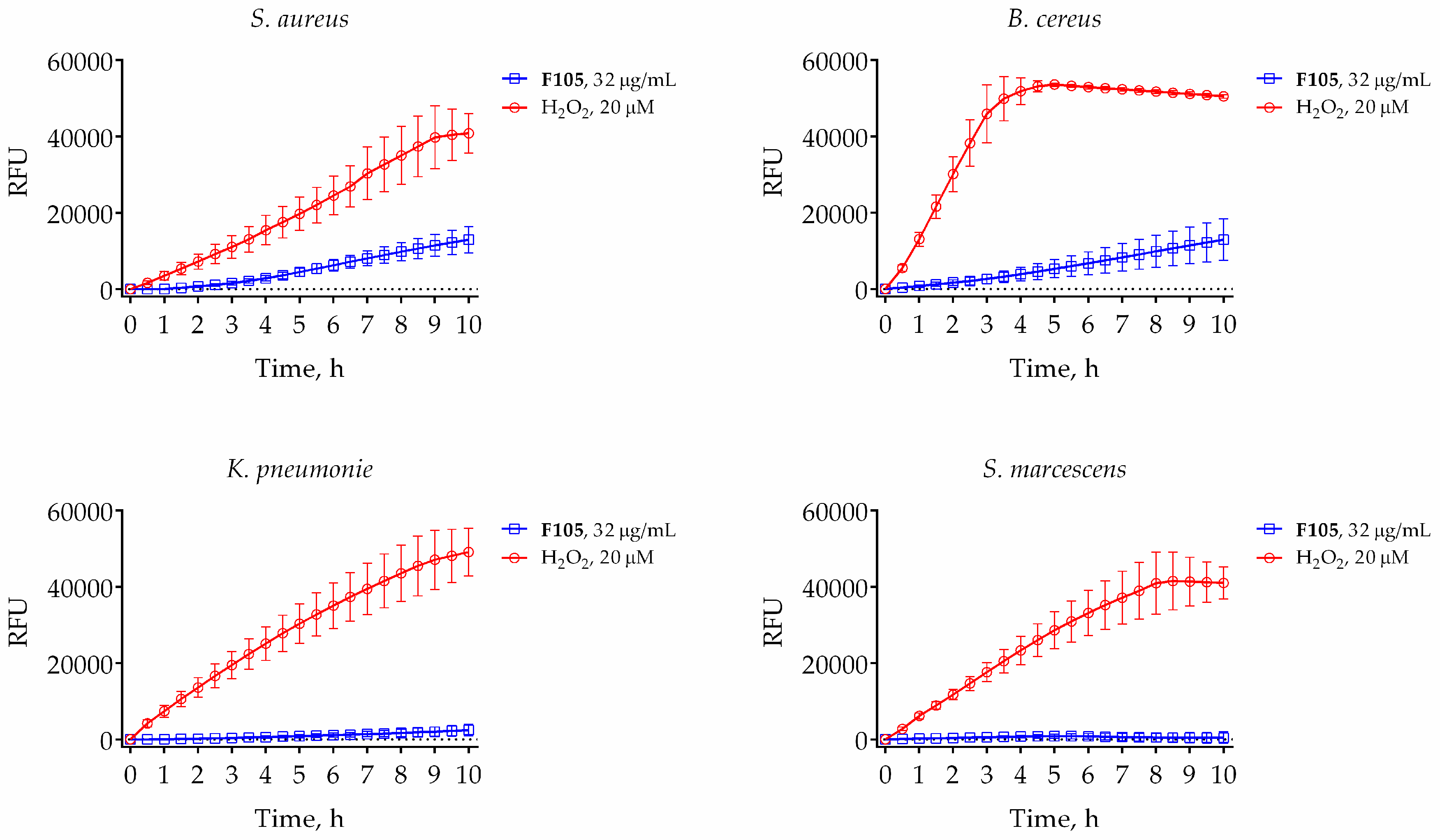
2.5. Reactive Oxygen Species (ROS) Induction
2.6. Effect of F105 on Membrane Potential
2.7. Effect of F105 on S. aureus Cell Integrity
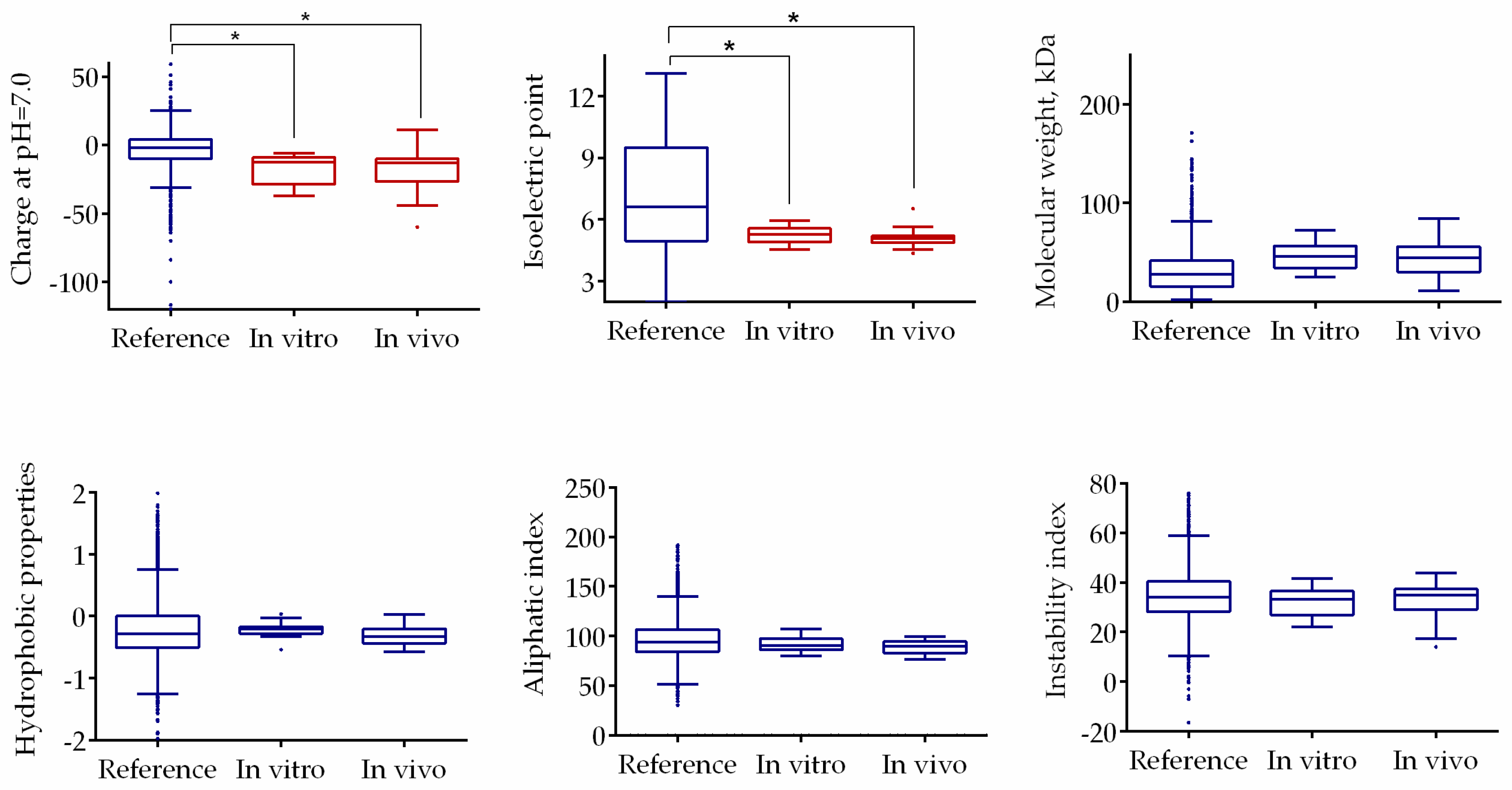
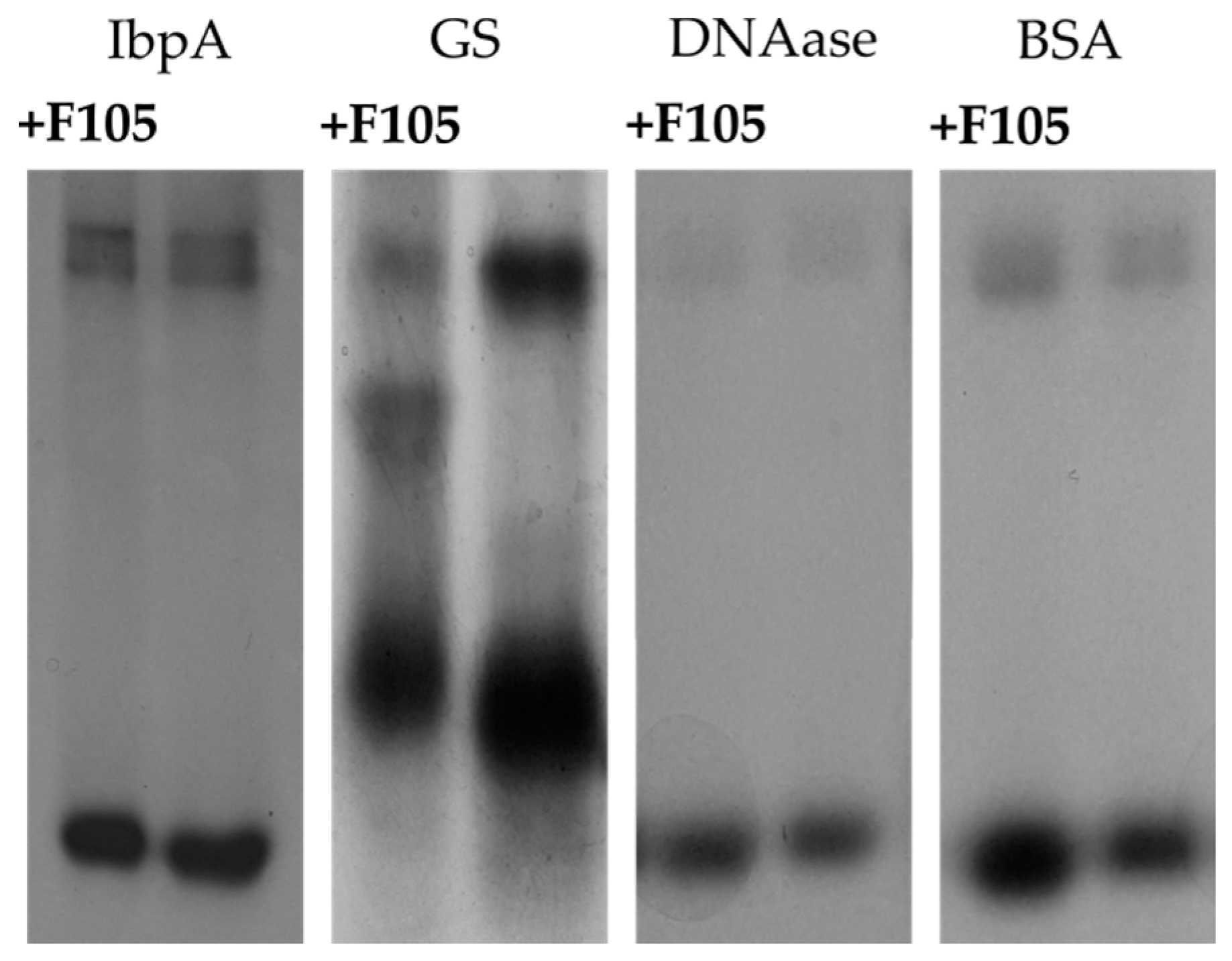
2.8. F105 Nonspecifically Interacts with a Number of Intracellular Proteins of S. aureus
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Determination of Minimal Inhibitory (MIC), Minimal Bactericidal Concentrations (MBC) and Resistance Development
4.3. Testing of the Resistance Development
4.4. Synthesis of 2(5H) Furanone Derivatives and General Chemical Experimental Procedures
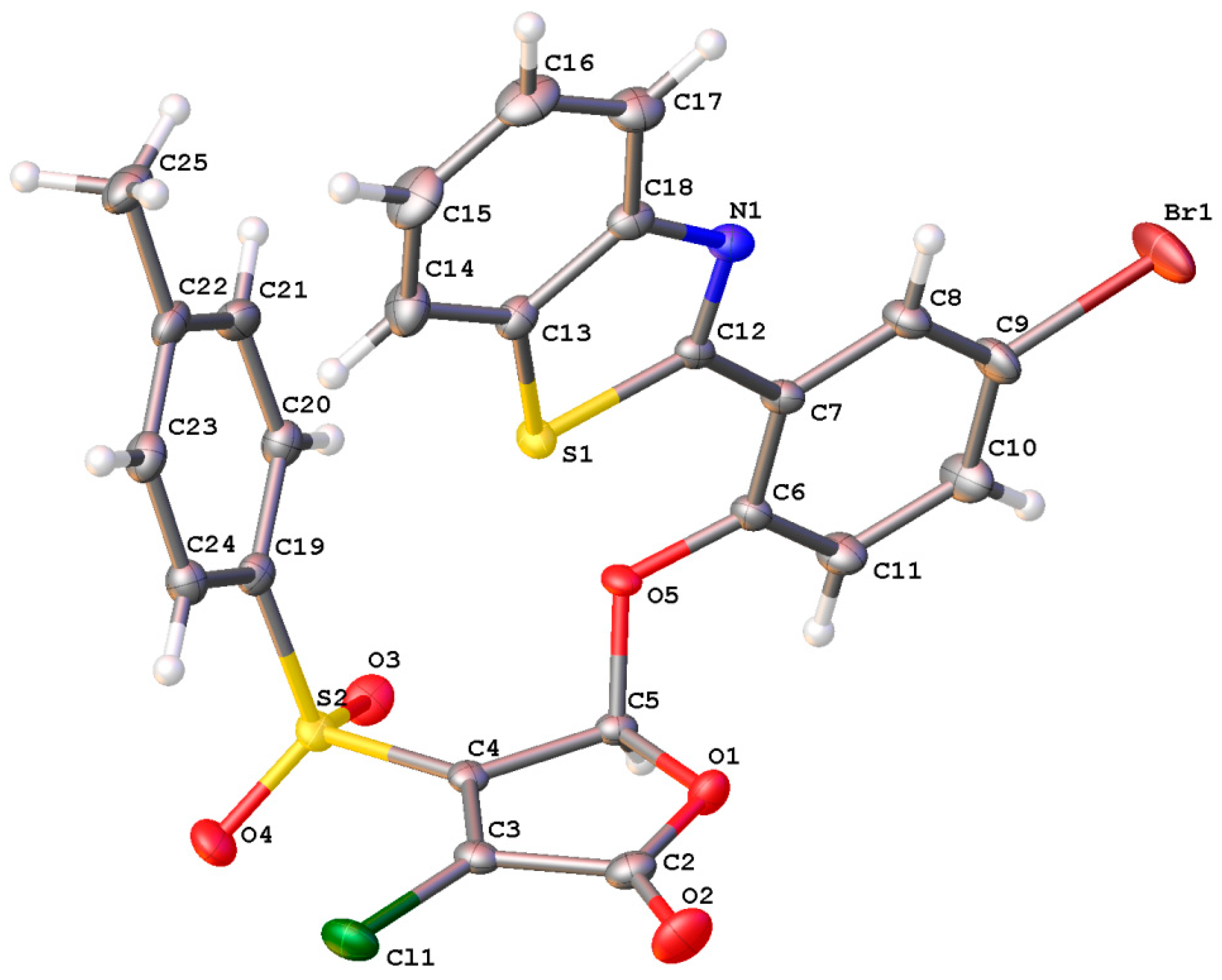
4.5. Single Crystal X-ray Analysis
4.6. Confocal Laser Scanning Microscopy
4.7. Atomic Force Microscopy
4.8. ROS Detection
4.9. Membrane Potential Evaluation
4.10. Proteomic Assays
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| F105 | 3-chloro-5(S)-[(1R,2S,5R)-2-isopropyl-5-methylcyclohexyloxy]-4-[4-methylphenylsulfonyl]-2(5H)-furanone |
| F145 | 5-[2-(Benzothiazol-2-yl)-4-bromophenoxy]-3-chloro-4-[(4-methylphenyl)sulfonyl]-2(5H)-furanone |
| BAC | Benzalkonium chloride |
| CFU | Colony forming units |
| RFU | Relative fluorescence unit |
| DCFDA | 2′,7′-dichlorofluorescin diacetate |
| DioC2(3) | 3,3′-diethyloxacarbocyanine iodide |
| DCFDA | 2′,7′-dichlorofluorescin diacetate |
| MBC | Minimal bactericidal concentration |
| CLSM | Confocal laser scanning microscopy |
| AFM | Atomic force microscopy |
| LC-MS | Liquid Chromatography Mass Spectrometry |
| MCPBA | m-Chloroperbenzoic acid |
| NMR | Nuclear magnetic resonance spectroscopy |
| HRMS | High-resolution mass spectrometry |
| TLC | Thin layer chromatography |
References
- Hodille, E.; Rose, W.; Diep, B.A.; Goutelle, S.; Lina, G.; Dumitrescu, O. The Role of Antibiotics in Modulating Virulence in Staphylococcus aureus. Clin. Microbiol. Rev. 2017, 30, 887–917. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, E.G.; Farulla, I.; Prignano, G.; Gallo, M.T.; Vespaziani, M.; Cavallo, I.; Sperduti, I.; Pontone, M.; Bordignon, V.; Cilli, L.; et al. Biofilm is a Major Virulence Determinant in Bacterial Colonization of Chronic Skin Ulcers Independently from the Multidrug Resistant Phenotype. Int. J. Mol. Sci. 2017, 18, 1077. [Google Scholar] [CrossRef] [PubMed]
- Estrela, A.B.; Abraham, W.-R. Combining Biofilm-Controlling Compounds and Antibiotics as a Promising New Way to Control Biofilm Infections. Pharmaceuticals 2010, 3, 1374–1393. [Google Scholar] [CrossRef]
- Chen, M.; Yu, Q.S.; Sun, H.M. Novel Strategies for the Prevention and Treatment of Biofilm Related Infections. Int. J. Mol. Sci. 2013, 14, 18488–18501. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Coenye, T. Quorum Sensing Inhibitors as Anti-Biofilm Agents. Curr. Pharm. Des. 2015, 21, 5–11. [Google Scholar] [CrossRef]
- Ren, D.C.; Sims, J.J.; Wood, T.K. Inhibition of biofilm formation and swarming of Escherichia coli by (5Z)-4-bromo-5(bromomethylene)-3-butyl-2(5H)-furanone. Environ. Microbiol. 2001, 3, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.C.; Bedzyk, L.A.; Setlow, P.; England, D.F.; Kjelleberg, S.; Thomas, S.M.; Ye, R.W.; Wood, T.K. Differential gene expression to investigate the effect of (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone on Bacillus subtilis. Appl. Environ. Microbiol. 2004, 70, 4941–4949. [Google Scholar] [CrossRef]
- Ledl, F.; Beck, J.; Sengl, M.; Osiander, H.; Estendorfer, S.; Severin, T.; Huber, B. Chemical pathways of the Maillard reaction. Prog. Clin. Biol. Res. 1989, 304, 23–42. [Google Scholar]
- Yujie, L.; Geng, X.; Huang, Y.C.; Li, Y.; Yang, K.Y.; Ye, L.H.; Chen, X.B.; Zhao, G.Q.; Yin, C. The Effect of Brominated Furanones on The Formation of Staphylococcus aureus Biofilm on PVC. Cell Biochem. Biophys. 2013, 67, 1501–1505. [Google Scholar] [CrossRef]
- Ngwane, A.H.; Panayides, J.L.; Chouteau, F.; Macingwana, L.; Viljoen, A.; Baker, B.; Madikane, E.; de Kock, C.; Wiesner, L.; Chibale, K.; et al. Design, synthesis, and In vitro antituberculosis activity of 2(5H)-Furanone derivatives. IUBMB Life 2016, 68, 612–620. [Google Scholar] [CrossRef]
- Gondela, E.; Walczak, K.Z. Synthesis and preliminary bioactivity assays of 3,4-dichloro-5-(omega-hydroxyalkylamino)-2(5H)-furanones. Eur. J. Med. Chem. 2010, 45, 3993–3997. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, M.; Riedel, K.; Rasmussen, T.B.; Heydorn, A.; Andersen, J.B.; Parsek, M.R.; Rice, S.A.; Eberl, L.; Molin, S.; Hoiby, N.; et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 2002, 148, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Lonn-Stensrud, J.; Landin, M.A.; Benneche, T.; Petersen, F.C.; Scheie, A.A. Furanones, potential agents for preventing Staphylococcus epidermidis biofilm infections? J. Antimicrob. Chemother. 2009, 63, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Trizna, E.Y.; Khakimullina, E.N.; Latypova, L.Z.; Kurbangalieva, A.R.; Sharafutdinov, I.S.; Evtyugin, V.G.; Babynin, E.V.; Bogachev, M.I.; Kayumov, A.R. Thio Derivatives of 2(5H)-Furanone As Inhibitors against Bacillus subtilis Biofilms. Acta Nat. 2015, 7, 102–107. [Google Scholar]
- Manefield, M.; de Nys, R.; Kumar, N.; Read, R.; Givskov, M.; Steinberg, P.; Kjelleberg, S.A. Evidence that halogenated furanones from Delisea pulchra inhibit acylated homoserine lactone (AHL)-mediated gene expression by displacing the AHL signal from its receptor protein. Microbiology 1999, 145, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Manefield, M.; Harris, L.; Rice, S.A.; De Nys, R.; Kjelleberg, S. Inhibition of luminescence and virulence in the black tiger prawn (Penaeus monodon) pathogen Vibrio harveyi by intercellular signal antagonists. Appl. Environ. Microbiol. 2000, 66, 2079–2084. [Google Scholar] [CrossRef] [PubMed]
- Manefield, M.; Rasmussen, T.B.; Henzter, M.; Andersen, J.B.; Steinberg, P.; Kjelleberg, S.; Givskov, M. Halogenated furanones inhibit quorum sensing through accelerated LuxR turnover. Microbiology 2002, 148, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Kayumov, A.R.; Khakimullina, E.N.; Sharafutdinov, I.S.; Trizna, E.Y.; Latypova, L.Z.; Hoang Thi, L.; Margulis, A.B.; Bogachev, M.I.; Kurbangalieva, A.R. Inhibition of biofilm formation in Bacillus subtilis by new halogenated furanones. J. Antibiot. 2015, 68, 297–301. [Google Scholar] [CrossRef]
- Zang, T.Z.; Lee, B.W.K.; Cannon, L.M.; Ritter, K.A.; Dai, S.J.; Ren, D.C.; Wood, T.K.; Zhou, Z.S. A naturally occurring brominated furanone covalently modifies and inactivates LuxS. Bioorg. Med. Chem. Lett. 2009, 19, 6200–6204. [Google Scholar] [CrossRef]
- Kuehl, R.; Al-Bataineh, S.; Gordon, O.; Luginbuehl, R.; Otto, M.; Textor, M.; Landmann, R. Furanone at Subinhibitory Concentrations Enhances Staphylococcal Biofilm Formation by luxS Repression. Antimicrob. Agents Chemother. 2009, 53, 4159–4166. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Trizna, E.Y.; Baidamshina, D.R.; Ryzhikova, M.N.; Sibgatullina, R.R.; Khabibrakhmanova, A.M.; Latypova, L.Z.; Kurbangalieva, A.R.; Rozhina, E.V.; Klinger-Strobel, M.; et al. Antimicrobial Effects of Sulfonyl Derivative of 2(5H)-Furanone against Planktonic and Biofilm Associated Methicillin-Resistant and -Susceptible Staphylococcus aureus. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Trizna, E.; Latypova, L.; Kurbangalieva, A.; Bogachev, M.; Kayumov, A. 2(5H)-Furanone Derivatives as Inhibitors of Staphylococcal Biofilms. Bionanoscience 2016. [Google Scholar] [CrossRef]
- Otsubo, T.; Minami, A.; Fujii, H.; Taguchi, R.; Takahashi, T.; Suzuki, T.; Teraoka, F.; Ikeda, K. 2-(Benzothiazol-2-yl)-phenyl-beta-D-galactopyranoside derivatives as fluorescent pigment dyeing substrates and their application for the assay of beta-D-galactosidase activities. Bioorg. Med. Chem. Lett. 2013, 23, 2245–2249. [Google Scholar] [CrossRef] [PubMed]
- Blazecka, P.G.; Belmont, D.; Curran, T.; Pflum, D.; Zhang, J. Further utilization of mucohalic acids: Palladium-free, regioselective etherification and amination of alpha,beta-dihalo gamma-methoxycarbonyloxy and gamma-acetoxy butenolides. Org. Lett. 2003, 5, 5015–5017. [Google Scholar] [CrossRef]
- Kurbangalieva, A.R.; Lodochnikova, O.A.; Devyatova, N.F.; Berdnikov, E.A.; Gnezdilov, O.I.; Litvinov, I.A.; Chmutova, G.A. Structural diversity of interaction products of mucochloric acid and its derivatives with 1,2-ethanedithiol. Tetrahedron 2010, 66, 9945–9953. [Google Scholar] [CrossRef]
- Kurbangalieva, A.R.; Devyatova, N.F.; Bogdanov, A.V.; Berdnikov, E.A.; Mannafov, T.G.; Krivolapov, D.B.; Litvinov, I.A.; Chmutova, G.A. Synthesis of novel arylthio derivatives of mucochloric acid. Phosphorus Sulfur Silicon Relat. Elem. 2007, 182, 607–630. [Google Scholar] [CrossRef]
- Latypova, L.Z.; Saigitbatalova, E.S.; Chulakova, D.R.; Lodochnikova, O.A.; Kurbangalieva, A.R.; Berdnikov, E.A.; Chmutova, G.A. Sulfides, sulfones, and sulfoxides of the furan-2(5H)-one series. synthesis and structure. Russ. J. Org. Chem. 2014, 50, 521–534. [Google Scholar] [CrossRef]
- Braoudaki, M.; Hilton, A.C. Mechanisms of resistance in Salmonella enterica adapted to erythromycin, benzalkonium chloride and triclosan. Int. J. Antimicrob. Agents 2005, 25, 31–37. [Google Scholar] [CrossRef]
- Kamatou, G.P.; Vermaak, I.; Viljoen, A.M.; Lawrence, B.M. Menthol: A simple monoterpene with remarkable biological properties. Phytochemistry 2013, 96, 15–25. [Google Scholar] [CrossRef]
- Narishetty, S.T.; Panchagnula, R. Effect of L-menthol and 1,8-cineole on phase behavior and molecular organization of SC lipids and skin permeation of zidovudine. J. Control. Release 2005, 102, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Shi, Q.S.; Huang, X.M.; Xie, X.B. The Three Bacterial Lines of Defense against Antimicrobial Agents. Int. J. Mol. Sci. 2015, 16, 21711–21733. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M.S.; Cooke, J.; Salib, R.J.; Holding, R.E.; Biggs, T.; Salamat, A.A.; Allan, R.N.; Newby, R.S.; Halstead, F.; Oppenheim, B.; et al. Reactive oxygen: A novel antimicrobial mechanism for targeting biofilm-associated infection. J. Glob. Antimicrob. Resist. 2017, 8, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Forrest, G.N.; Tamura, K. Rifampin Combination Therapy for Nonmycobacterial Infections. Clin. Microbiol. Rev. 2010, 23, 14–34. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.Q.; Wang, Z.Y.; Mei, W.J.; Fu, J.H.; Tan, Y.H.; Luo, S.H. Reaction of 5-alkoxy-3,4-dihalo-2(5H)-furanones with secondary amines: Expected versus unanticipated products and their preliminary bioactivity investigations. Monatshefte Chem. 2012, 143, 443–453. [Google Scholar] [CrossRef]
- Leclercq, R.; Canton, R.; Brown, D.F.; Giske, C.G.; Heisig, P.; MacGowan, A.P.; Mouton, J.W.; Nordmann, P.; Rodloff, A.C.; Rossolini, G.M.; et al. EUCAST expert rules in antimicrobial susceptibility testing. Clin. Microbiol. Infect. 2013, 19, 141–160. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Dieases (ESCMID). EUCAST Definitive Document E.Def 1.2, May 2000: Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect. 2000, 6, 503–508. [Google Scholar] [CrossRef]
- Locher, H.H.; Caspers, P.; Bruyere, T.; Schroeder, S.; Pfaff, P.; Knezevic, A.; Keck, W.; Ritz, D. Investigations of the Mode of Action and Resistance Development of Cadazolid, a New Antibiotic for Treatment of Clostridium difficile Infections. Antimicrob. Agents Chemother. 2014, 58, 901–908. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Pavlova, A.S.; Khabibrakhmanova, A.M.; Faizova, G.M.; Kurbangalieva, A.R.; Katsunori, T.; Trizna, E.Y.; Baidamshina, D.R.; Bogachev, M.I.; Kayumov, A.R. Targeting Bacillus cereus cells: Increasing efficiency of antimicrobials by the bornyl-possessing 2(5H)-furanone derivative. New Microbiol. 2019, 42, Epub ahead of print. [Google Scholar] [PubMed]
- Sheldrick, G.M. SHELXT—Integrated space-group and crystal-structure determination. Acta Crystallogr. Sect. A Found. Adv. 2015, 71, 3–8. [Google Scholar] [CrossRef]
- Sheldrick, G.M. Crystal structure refinement with SHELXL. Acta Crystallogr. Sect. C Struct. Chem. 2015, 71, 3–8. [Google Scholar] [CrossRef]
- Farrugia, L.J. WinGX and ORTEP for Windows: An update. J. Appl. Crystallogr. 2012, 45, 849–854. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during assembly of head of bacteriophage-T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Vishnyakov, I.E.; Levitskii, S.A.; Manuvera, V.A.; Lazarev, V.N.; Ayala, J.A.; Ivanov, V.A.; Snigirevskaya, E.S.; Komissarchik, Y.Y.; Borchsenius, S.N. The identification and characterization of IbpA, a novel alpha-crystallin-type heat shock protein from mycoplasma. Cell Stress Chaperones 2012, 17, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Hauf, K.; Kayumov, A.; Gloge, F.; Forchhammer, K. The Molecular Basis of TnrA Control by Glutamine Synthetase in Bacillus subtilis. J. Biol. Chem. 2016, 291, 3483–3495. [Google Scholar] [CrossRef] [PubMed]
- Vishnyakov, I.E.; Bogachev, M.I.; Salafutdinov, I.; Borchsenius, S.N.; Kayumov, A.R. The Temperature-Dependent Selectivity of Potential Interaction Partners for the Small Heat Shock Protein IbpA from Acholeplasma laidlawii. Bionanoscience 2016, 6, 437–442. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC, μg/mL | MBC, μg/mL | |
|---|---|---|
| S. aureus | 8 | 32 |
| S. epidermidis | 16 | 32 |
| B. cereus | 8 | 32 |
| B. subtilis | 16 | 32 |
| M. luteus | 8 | 32 |
| K. pneumoniae | >128 | >128 |
| S. marcescens | >128 | >128 |
| P. aeruginosa | >128 | >128 |
| E. coli | >128 | >128 |
| Control | F105, 32 μg/mL | BAC, 4 μg/mL |
|---|---|---|
| 7 × 109 | 2 × 105 | 5 × 105 |
| Protein | IbpA | GS | DNAase | BSA |
|---|---|---|---|---|
| Charge | −2.3 | −18.5 | −8.1 | −9.9 |
| pI | 5.7 | 5.1 | 5.3 | 6.2 |
| Relative mobility change index | 0.979 ± 0.004 | 0.971 ± 0.015 | 1.016 ± 0.005 | 1.017 ± 0.004 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharafutdinov, I.S.; Pavlova, A.S.; Akhatova, F.S.; Khabibrakhmanova, A.M.; Rozhina, E.V.; Romanova, Y.J.; Fakhrullin, R.; Lodochnikova, O.A.; Kurbangalieva, A.R.; Bogachev, M.I.; et al. Unraveling the Molecular Mechanism of Selective Antimicrobial Activity of 2(5H)-Furanone Derivative against Staphylococcus aureus. Int. J. Mol. Sci. 2019, 20, 694. https://doi.org/10.3390/ijms20030694
Sharafutdinov IS, Pavlova AS, Akhatova FS, Khabibrakhmanova AM, Rozhina EV, Romanova YJ, Fakhrullin R, Lodochnikova OA, Kurbangalieva AR, Bogachev MI, et al. Unraveling the Molecular Mechanism of Selective Antimicrobial Activity of 2(5H)-Furanone Derivative against Staphylococcus aureus. International Journal of Molecular Sciences. 2019; 20(3):694. https://doi.org/10.3390/ijms20030694
Chicago/Turabian StyleSharafutdinov, Irshad S., Anna S. Pavlova, Farida S. Akhatova, Alsu M. Khabibrakhmanova, Elvira V. Rozhina, Yulia J. Romanova, Rawil Fakhrullin, Olga A. Lodochnikova, Almira R. Kurbangalieva, Mikhail I. Bogachev, and et al. 2019. "Unraveling the Molecular Mechanism of Selective Antimicrobial Activity of 2(5H)-Furanone Derivative against Staphylococcus aureus" International Journal of Molecular Sciences 20, no. 3: 694. https://doi.org/10.3390/ijms20030694
APA StyleSharafutdinov, I. S., Pavlova, A. S., Akhatova, F. S., Khabibrakhmanova, A. M., Rozhina, E. V., Romanova, Y. J., Fakhrullin, R., Lodochnikova, O. A., Kurbangalieva, A. R., Bogachev, M. I., & Kayumov, A. R. (2019). Unraveling the Molecular Mechanism of Selective Antimicrobial Activity of 2(5H)-Furanone Derivative against Staphylococcus aureus. International Journal of Molecular Sciences, 20(3), 694. https://doi.org/10.3390/ijms20030694